To facilitate the understanding of the potential
roles of incretins, including GLP-1, GIP and GCG, in oncology, this
section provides a concise overview of the core physiology of
GLP-1, GIP and GCG. Although they share common signaling modules
such as cyclic adenosine monophosphate (cAMP), their receptor
distribution and principal metabolic functions underlie their
distinct influences on the TME (15,16).
Fig. 1 summarizes the fundamental
mechanisms of action of these three hormones.
Beyond the Gs pathway, GLP-1R engages multiple
signaling mechanisms. GLP-1R activates the PI3K/Foxo1 pathway to
promote β-cell survival, and can also recruit β-arrestins, which
regulate receptor trafficking and desensitization, and can also
modulate downstream extracellular signal-regulated kinases
(ERK1/2), thereby influencing β-cell function in a
context-dependent manner (28,29).
Additionally, GLP-1 attenuates palmitic acid-induced inflammation
by inactivating mTORC1 signaling, serving a pivotal role in
modulating insulin resistance, inflammation and autophagy (30). These diverse intracellular
signaling pathways are summarized in Fig. 1 (15,31).
In summary, GLP-1 signaling supports β-cell survival,
glucose-dependent insulin secretory responses and systemic
metabolic regulation. This suggests its dual role in oncology:
PI3K/Akt and ERK pathways activated by GLP-1RAs are classical
pro-survival pathways that may be utilized by tumor cells in some
contexts. GLP-1RAs exert strong systemic metabolic regulation and
immune regulation effects, which may indirectly inhibit tumor
growth. The ultimate effect of GLP-1R activation in a given cancer
will most likely depend on whether tumor cells express functional
GLP-1R and how GLP-1R downstream signaling is integrated with
oncogenic signals.
GIP is secreted by K-cells in the proximal small
intestine, and shares considerable functional overlap with
GLP-1(32). In pancreatic β-cells,
upon binding to its receptor [glucose-dependent insulinotropic
polypeptide receptor (GIPR)], GIP similarly elevates intracellular
cAMP levels, activating both PKA and Epac (15,33),
thereby enhancing glucose-stimulated insulin secretion (34). In adipose tissue, the lipogenic
effects of GIP appear to be context-dependent and influenced by
insulin availability and metabolic status (35). GIP promotes β-cell proliferation
and survival through the PI3K/Akt and MAPK/ERK pathways (36,37),
while inhibiting apoptosis via regulation of Bcl-2 and Bax
expression (38,39). These diverse intracellular pathways
are also illustrated in Fig. 1. A
key distinction between GIP and GLP-1 lies in the role of GIP in
adipose tissue, where it promotes fatty acid storage (40) and modulates lipolysis in an
insulin- and context-dependent manner (41). These metabolic effects may alter
nutrient availability and thereby influence tumor growth under
obese conditions in a context-dependent manner (42).
GCG is secreted by pancreatic α cells, and its
secretion is stimulated by hypoglycemic conditions or by elevated
levels of certain amine hormones such as epinephrine (43). As the primary counter-regulatory
hormone to insulin, it collaborates with insulin to maintain
glucose homeostasis (44). Upon
binding to the GCGR on hepatocytes, GCG couples with Gs and Gq
proteins (45). Activation of Gs
stimulates the cAMP/PKA signaling pathway, which drives hepatic
gluconeogenesis (46-48)
and glycogenolysis (49). The Gq
pathway activates phospholipase C (50), generating inositol trisphosphate
and diacylglycerol, thereby mobilizing intracellular calcium ions
(51). This signaling upregulates
key gluconeogenic enzymes, including phosphoenolpyruvate
carboxykinase and glucose-6-phosphatase (52,53).
Additionally, GCG stimulates lipolysis and ketogenesis (54), suppresses fatty acid synthesis
while enhancing fatty acid oxidation (55), improves mitochondrial function
(56) and reduces oxidative stress
(57). GCG also modulates hepatic
lipid metabolism through CREBH- and Insig-2-related transcriptional
programs and other acetylation-dependent regulatory mechanisms
(58). The intricate signaling
network of GCG in hepatocytes is detailed in Fig. 1 (59,60).
GCG is an energy mobilization hormone. GCG-driven
hepatic glucose export and lipolysis may provide abundant fuel for
tumor cells that are dependent on glucose and lipids. However, the
oncological significance of GCG signaling remains insufficiently
defined, and the currently available evidence does not support a
uniform conclusion that GCGR activation exerts a direct antitumor
effect (61). In this context, the
antitumor activity reported for incretin-based polyagonists should
not be interpreted as evidence for GCGR agonism alone, but rather
as an indication that coordinated multi-receptor targeting may
induce favorable systemic metabolic remodeling (62,63).
The apparent physiological opposition between GLP-1 and GCG does
not preclude pharmacological synergy, since GLP-1/GCG dual agonists
are engineered to achieve balanced receptor activation rather than
simple co-administration of two counter-regulatory hormones
(61,64). Such agents may retain the anorectic
and insulin-sensitizing effects of GLP-1 while utilizing the
effects of GCG on energy expenditure and lipid metabolism without
causing excessive hyperglycemia (65). In this context, their potential
antitumor relevance is biologically plausible, as they may improve
metabolic dysfunction, reduce obesity-related chronic inflammation,
and influence nutrient availability and metabolic reprogramming in
the TME (66-68).
Indirect support for this hypothesis comes from preclinical studies
of related incretin-based polyagonists. Tirzepatide, a dual
GIPR/GLP-1R agonist, has shown antitumor effects in models of EC
and colorectal cancer (CRC) (69),
whereas retatrutide, a triple GIPR/GLP-1R/GCGR agonist, has shown
antitumor activity in pancreatic and lung cancer models (70). Nevertheless, the currently
available evidence remains indirect and is insufficient to directly
demonstrate either a synergistic antitumor effect of GLP-1/GCG
co-agonism or an antitumor effect mediated through GCGR activation
alone. At present, the proposed antitumor relevance of GLP-1/GCG
co-agonism, and, more specifically, any antitumor effect
attributable to GCGR activation alone, should be regarded as a
mechanistic hypothesis requiring direct validation in
tumor-specific models.
GLP-1, GIP and GCG are key peptide hormones that
regulate metabolic homeostasis; however, they have distinct
origins, receptors and frequently opposing metabolic functions
(71). The clinical translation of
their receptor agonists has revealed divergent oncological
properties (72). GLP-1RAs are
well-established for managing T2D and obesity (61,73,74),
and exhibit emerging antitumor potential in related cancers, such
as colorectal and endometrial cancers (75,76).
The role of GIPR modulation in oncology remains poorly defined,
while investigations into GCGR agonism to increase whole-body
energy expenditure in obesity and metabolic disease are still in
progress, although its potential hyperglycemic effects necessitate
caution (61,66). Table
SI systematically compares the secretion, signaling
transduction and metabolic actions of GLP-1, GIP and GCG, and
summarizes their impacts on the TME offering a foundational
theoretical basis for their integrated assessment in cancer biology
(15,16,45,61,62,66).
This comparative analysis emphasizes that shared signaling pathways
can generate antagonistic physiological outcomes, a concept
critical for elucidating their microenvironment-dependent functions
in cancer (31,77).
Among the three hormones GLP-1, GIP and GCG, GLP-1
has been the most extensively studied in the field of malignant
tumors (74,78,79).
Thus, the antitumor potential of GLP-1RAs represents the primary
focus of the present review.
Studies have demonstrated that GLP-1RAs confer
therapeutic benefits in various T2D- or obesity-related conditions,
including polycystic ovary syndrome (80), metabolic syndrome (81), non-alcoholic fatty liver disease
(82) and obesity- or
diabetes-associated malignancies, such as gallbladder, ovarian and
esophageal cancers (73,79). However, their effects are not
uniformly beneficial. Under certain circumstances, these hormones
may exhibit pro-tumorigenic properties. The cAMP signaling pathway
serves as a central pathway shared by these three hormones, capable
of transmitting either pro- or anti-proliferative signals depending
on the cellular context (78,79).
For instance, GLP-1 has been shown to promote the progression of
medullary thyroid carcinoma (MTC) in rodent models (83). In some breast cancer models,
liraglutide enhances tumor cell proliferation via the NADPH oxidase
4 (NOX4)/reactive oxygen species (ROS)/VEGF signaling pathway
(84). Similarly, in EC,
upregulation of GLP-1R facilitates cell proliferation and migration
while conferring resistance to ferroptosis, thereby driving tumor
progression (85). These divergent
findings highlight that the net effect on tumorigenesis is
modulated by the interplay among tumor cell-intrinsic factors, host
systemic metabolism and pharmacological characteristics. Thus,
elucidating the contextual determinants underlying these dual
effects is essential for the safe and effective application of
these agents in oncology.
The TME is a complex ecosystem composed of tumor
cells, immune cells, fibroblasts and the extracellular matrix
(ECM). The TME serves as a dynamic interface, where interactions
between malignant and non-malignant components promote tumor
growth, invasion and metastasis (86,87).
The TME is characterized by nutrient deprivation, hypoxia and an
acidic milieu, driven by aberrant tumor vasculature and the Warburg
effect (a preference for aerobic glycolysis even in the presence of
oxygen), which collectively facilitate immune evasion by tumor
cells (88,89). The release of acidic metabolites
such as lactate, generated by tumor cells via glycolysis,
exacerbates local acidosis within the TME (90). This acidic environment not only
suppresses immune cell function but also activates pro-angiogenic
factors (91,92). Immune cells serve a pivotal role in
the TME. Cytotoxic CD8+ T cells serve as the primary
effectors of antitumor immunity (93,94);
however, their function is often suppressed by regulatory T cells
(Tregs), myeloid-derived suppressor cells (95) and tumor-associated macrophages
(TAMs) (96). Metabolic
competition for nutrients such as glucose and amino acids between
tumor cells and immune cells further impairs effective immune
responses (97). Given the potent
immunomodulatory and metabolic effects of GLP-1, GIP and GCG,
therapies targeting these pathways may influence the TME by
modulating nutrient competition and immune-cell function; however,
current mechanistic evidence is strongest for GLP-1-related agents
(98) Based on this premise, the
present review summarizes the important roles of GLP-1-related
agents within the tumor immune microenvironment, for which the
current evidence base is the most extensive, as shown in Fig. 2 (98,99).
T cells are core components of adaptive immunity and
central to antitumor immunotherapy. Cytotoxic T cells are primarily
characterized by CD8 expression, helper T cells by CD4 expression,
and Tregs by co-expression of CD25 and CD4 expression (100). During tumor progression,
cancer-associated fibroblasts, M2-type macrophages and Tregs can
establish an immunosuppressive barrier that counteracts
CD8+ T cell-mediated antitumor immunity (101). Therefore, effective antitumor
immunotherapy requires not only sufficient effector T cell activity
but also the maintenance of T cell fitness and persistence. Current
evidence suggests that the effects of GLP-1-related signaling on
T-cell biology are context dependent (102,103).
In inflammatory settings, this immunoregulatory
effect appears to be tissue-protective. Under alloimmune
conditions, GLP-1R-expressing T cells are predominantly enriched
within the exhausted CD8+ T cell population, where
GLP-1R signaling functions as a negative costimulatory pathway that
restrains excessive T-cell activation and tissue-damaging
inflammation (104). In a murine
model of nephrotoxic serum nephritis, GLP-1R activation suppressed
the proliferation of pro-inflammatory T helper 1 and T helper 17
cells, while reducing IL-6 production, thereby exerting renal
protective effects (105).
Furthermore, in intestinal intraepithelial lymphocytes, GLP-1R
activation inhibits proximal T cell receptor signaling in a
PKA-dependent manner, suppressing T cell activation and attenuating
local IFN-γ-driven inflammation (106). Through direct suppression of T
helper cell-mediated hyperinflammation and PKA-dependent modulation
of local immune responses, GLP-1RAs help protect normal tissues
(107). These findings support a
role for GLP-1R signaling in limiting T-cell-mediated
hyperinflammation and preserving immune homeostasis in normal
tissues.
This anti-inflammatory effect should not be
interpreted as uniformly detrimental to antitumor immunity
(104). GLP-1RAs appear to
support antitumor immunity mainly through remodeling of the
suppressive TME rather than through direct restoration of exhausted
intratumoral CD8+ T-cell function (108). In a syngeneic 4T1 breast cancer
model, semaglutide increased intratumoral T-cell infiltration,
promoted dendritic cell maturation, reduced FoxP3+ Tregs
and enhanced CD8+ T-cell cytotoxicity despite showing no
direct tumoricidal activity in vitro (108). Similarly, exenatide enhanced the
efficacy of programmed cell death protein-1 (PD-1) blockade in LLC-
and MC38-based lung and colon cancer models (109), and liraglutide did so in LLC- and
Hepa1-6-bearing mice, mainly by attenuating neutrophil
extracellular trap (NET)- and ROS-driven immunosuppression and
thereby permitting stronger CD8+ T-cell effector
responses (110).
In obesity-related tumor settings, the immunological
effects of GLP-1R agonism appear to be more pronounced. In
diet-induced obese mice bearing subcutaneous KPN1.1 tumors,
liraglutide reduced tumor burden, whereas in an autochthonous
KRAS/p53-driven non-small cell lung cancer model, semaglutide
improved tumor control and survival. These effects were accompanied
by increased intratumoral CD4+ T-cell and natural killer
(NK) cell frequencies, a shift of CD8+ and conventional
CD4+ T cells toward a memory-like phenotype, reduced
regulatory T-cell abundance, and improved antigen-presenting
cell-associated immune features (111).
Collectively, these findings support a role for
GLP-1RAs in strengthening antitumor T-cell immunity mainly through
TME remodeling, with particularly evident effects under
obesity-associated metabolic conditions (111). However, current evidence remains
insufficient to conclude that GLP-1 directly restores exhausted
CD8+ T-cell function or directly regulates the
PD-1/programmed death-ligand 1 axis within tumors.
The role of neutrophils in cancer therapy remains
controversial. The N1 phenotype exhibits antitumor properties,
whereas the N2 phenotype promotes tumor progression (112). Studies have indicated that a key
mechanism of neutrophil-mediated tumor promotion involves the
formation of NETs, which can physically shield tumor cells from
cytotoxic lymphocytes and NK cells (113), while also inducing T-cell
exhaustion and functional impairment (114). In lung and liver cancer models,
the combination of liraglutide and exenatide with anti-PD-1 therapy
reduced ROS production, suppressed NET formation, and decreased the
levels of peripheral myeloperoxidase-DNA complexes and elastase.
These effects enhanced the cytotoxicity of CD8+ T cells
and their capacity to eliminate tumor cells, thereby augmenting the
antitumor efficacy of PD-1 inhibitors (109,110). Thus, in modulating neutrophil
activity, GLP-1RAs primarily exert indirect antitumor effects by
reducing NET formation. Furthermore, serum NET levels may serve as
a biomarker for predicting the efficacy of immunotherapy (115), offering valuable insights for
drug selection in patients with malignancies.
NK cells are innate cytotoxic lymphocytes that
surveil and eliminate tumor cells through the release of cytolytic
granules (116). NK cells also
coordinate with other immune cells via pro-inflammatory cytokines
and chemokines (117). Obesity
impairs NK cell function (118),
and GLP-1 analog therapy has been shown to restore NK cell effector
functions in individuals with obesity. After 6 months of treatment
with GLP-1 analogs, the levels of IFN-γ and granzyme B produced by
NK cells were markedly elevated, indicating partial restoration of
NK cell functionality (118).
Phenotypic differentiation of neutrophils is regulated by NK cells,
which can promote neutrophil polarization toward the N1 phenotype
through an IFN-γ-dependent mechanism (119). In NK cell-depleted mouse models,
neutrophils tend to adopt the N2 phenotype, characterized by
upregulated VEGF-A expression, thereby promoting tumor growth and
angiogenesis (119). Furthermore,
malignant tumors often exhibit hyperactivation of the IL-6/STAT3
signaling pathway, which dampens CD4+ T cell activity
and subsequently suppresses NK T cell function (120). In hepatocellular carcinoma (HCC)
models, liraglutide enhanced NK cell-mediated tumor lysis by
inhibiting the IL-6/STAT3 pathway (121). Thus, GLP-1 analogs not only
directly restore NK cell function but also augment antitumor
immunity through coordinated immune network interactions.
TAMs exhibit high plasticity and heterogeneous
responses to environmental stimuli (122), enabling their polarization into
either antitumor M1 or pro-tumor M2 phenotypes (123-125).
In established tumors, TAMs can further promote immunosuppression
and tumor progression by producing pro-angiogenic factors, such as
VEGF, and by releasing IL-10-rich signals that impair effective
cytotoxic T-cell responses (126). Multiple studies have demonstrated
that GLP-1RAs promote the polarization of uncommitted macrophages
toward an anti-inflammatory phenotype (127,128) and facilitate the transition from
M1 to M2 polarization (129,130). In patients with obesity and T2D,
GLP-1RA therapy reduces the levels of inflammatory cytokines
(TNF-α, IL-1β and IL-6) and the macrophage activation marker
soluble CD163, while increasing anti-inflammatory adiponectin
(131). Although M2 polarization
is generally associated with tissue repair, it exerts pro-tumor
effects in established cancers (132). GLP-1RA treatment enhances IL-10
secretion by M2 macrophages (133), an effect that exhibits strong
concentration dependence: Low IL-10 concentrations may facilitate
tumor immune evasion, whereas high concentrations promote
CD8+ T cell proliferation, activation and cytotoxicity
(134). The dosage and duration
of GLP-1RA treatment, as well as the inflammatory status of the
patient, collectively influence macrophage polarization (135). Thus, the dual effects of GLP-1RAs
in breast cancer and EC may be linked to individual variations in
macrophage polarization (136).
Further investigation is required to determine the net impact of
GLP-1RAs on TAMs and tumor outcomes. Identifying optimal dosing
regimens may help resolve the context-dependent duality of GLP-1RAs
in certain malignancies.
DCs serve a pivotal role in initiating and
regulating T cell-mediated antitumor immune responses by enhancing
the cytotoxicity of NK and T cells (137). Within the suppressive TME,
immature or dysfunctional DCs typically exhibit reduced major
histocompatibility complex (MHC) II and CD86 expression, which
limits antigen presentation and impairs effective T-cell priming
(138). Evidence indicates that
GLP-1RAs markedly augment DC-mediated adaptive immunity (108). In a 4T1 breast cancer mouse
model, semaglutide treatment markedly promoted the accumulation of
CD11c+ DCs in both the spleen and TME, while
upregulating the expression of surface MHC II molecules and the
co-stimulatory marker CD86, thereby facilitating DC maturation.
Mechanistically, semaglutide enhanced the mRNA expression levels of
antigen processing and presentation-related molecules (MHC I and
transporter associated with antigen processing) in DCs by
activating the PI3K/phosphorylated-Akt signaling pathway. These
findings demonstrated that GLP-1RAs strengthened the ability of DCs
to cross-present antigens and activate cytotoxic T lymphocytes,
underscoring their central role in antitumor immunity (108).
B cells are key components of humoral immunity,
contributing through antibody production and support of cellular
immune responses (139). B cells
are essential constituents of tertiary lymphoid structures, and
their primary functions include antigen presentation and antibody
secretion (140). Prognostically
relevant B-cell subsets include stress-responsive memory B cells
and tumor-associated atypical B cells (141). In gastric cancer, increased
infiltration of CD20+ B cells in gastric cancer tissues,
as detected by immunohistochemistry, is associated with improved
patient outcomes (142). However,
the role of B cells in GLP-1RA therapy remains unclear. Given that
their core function (antibody production) has limited direct links
to energy metabolism, to the best of our knowledge, no direct
studies have investigated interactions between B cells and
GLP-1RAs. Early genetic engineering research has demonstrated that
in transgenic mice constitutively expressing the exendin-4
precursor, lymphocyte infiltration occurred in multiple tissues.
Specifically, B220+ B cells accumulated in the pancreas
and liver, while CD4+ and CD8+ T cells were
increased in the liver and kidneys (143). These findings suggested that
sustained proexendin-4 expression may break immune tolerance and
simultaneously activate humoral and cellular immune responses.
Nevertheless, this model involved continuous, high-level transgene
expression across multiple tissues, which does not reflect the
pharmacokinetic profile of therapeutic GLP-1RAs. Furthermore, the
functional consequences of immune activation and underlying
mechanisms were not explored. However, the study implied that GLP-1
and its analogs may interact with various immune cells, warranting
further investigation.
In summary, GLP-1RAs modulate the tumor immune
microenvironment through multiple mechanisms. Documented antitumor
mechanisms include suppression of excessive inflammation and
potential functional restoration of CD8+ T cells,
reduction of NET formation in neutrophils to enhance
CD8+ T cell cytotoxicity, direct restoration of NK cell
function with indirect promotion of N1-polarized neutrophils, and
promotion of DC maturation and antigen presentation to reinforce
T-cell activation. Conversely, potential pro-tumor effects include
induction of anti-inflammatory macrophage polarization. The impact
on B cells remains undetermined.
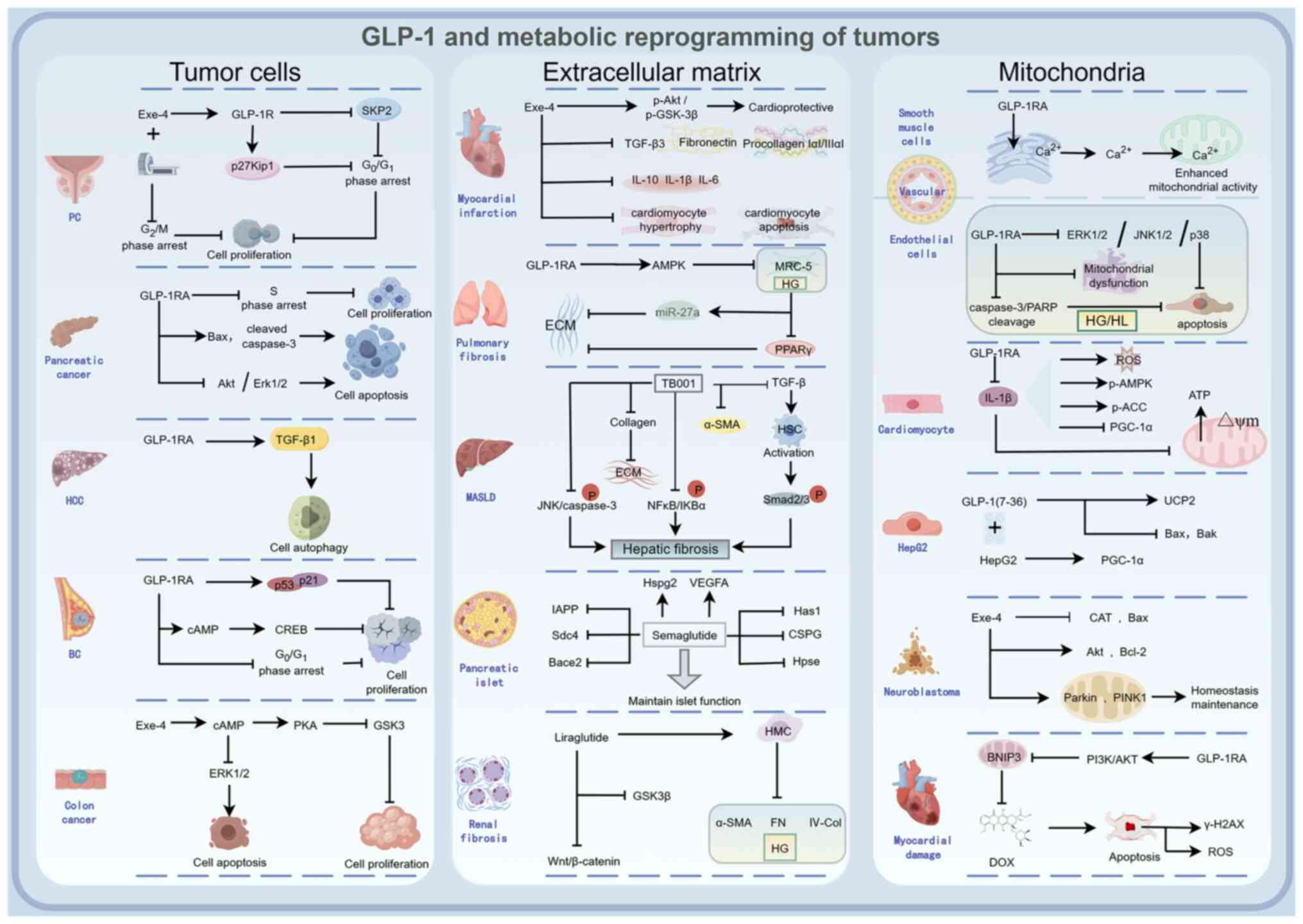
Metabolic reprogramming is a hallmark of cancer,
enabling rapid proliferation, survival and stress adaptation
(144). GLP-1RAs influence this
process through direct targeting of tumor cells, remodeling of the
ECM and modulation of mitochondrial function (Fig. 3).
GLP-1R expression levels exhibit an inverse
association with the Gleason score in prostate cancer (146). Exendin-4 reduces S-phase kinase
associated protein 2 expression and upregulates p27Kip1 in a
dose-dependent manner, inducing G0/G1 phase
arrest in ALVA-41 cells and thereby suppressing proliferation
(145). Combined treatment with
docetaxel and liraglutide decreases viability in the LNCaP prostate
cancer cell line, leading to cell cycle arrest and apoptosis, with
liraglutide enhancing chemotherapeutic efficacy and reducing drug
resistance (147). Furthermore,
exendin-4 potentiates the anti-proliferative effects of ionizing
radiation in vitro and in vivo, and enhances
radiation-induced G2/M phase arrest in a dose-dependent
manner, thereby improving radiosensitivity (148). Liraglutide reduces the
proliferation of HepG2 cells by inducing TGF-β1-mediated autophagy
and senescence without altering oxidative stress levels (149). GLP-1RAs and exendin-4 decrease
breast cancer cell viability and promote apoptosis via activation
of cAMP and CREB signaling (12).
Liraglutide inhibits the proliferation of MCF-7 human breast cancer
cells by ~48%, reduces colony formation, induces
G0/G1 phase arrest and ultimately suppresses
cancer cell proliferation (150).
In human pancreatic cancer cells, liraglutide inhibits
proliferation and promotes apoptosis accompanied by S-phase arrest
through activation of cAMP-mediated Akt and ERK1/2 signaling
pathways (151). Exendin-4
increases intracellular cAMP levels in colon cancer CT26 cells,
activates the PKA pathway, and inhibits the activity of signaling
kinases GSK3 and ERK1/2, consequently suppressing CT26 cell
proliferation (152).
Through the modulation of signaling pathways such as
the cAMP signaling pathway, GLP-1RAs induce cell cycle arrest and
apoptosis in various cancer cells and enhance sensitivity to
radiotherapy and chemotherapy. For example, exendin-4 has been
shown to potentiate the antiproliferative effects of ionizing
radiation and increase radiation-induced G2/M phase
arrest, whereas liraglutide enhances the effects of docetaxel in
prostate cancer cells. These findings suggest that GLP-1RAs may
have potential as adjuncts to radiotherapy or chemotherapy in
selected tumor models (148).
GLP-1RAs have exhibited protective effects across
multiple organ fibrosis and injury models. In the cardiovascular
system, GLP-1RAs improve post-myocardial infarction remodeling by
enhancing Akt/GSK-3 phosphorylation and downregulating profibrotic
and inflammatory factors such as procollagen, TGF-β3 and
interleukins (156). In diabetic
kidney disease, liraglutide suppresses mesangial cell production of
fibronectin and type IV collagen via activation of the
Wnt/β-catenin pathway (157). In
diabetic lung injury, GLP-1RAs reduce ECM accumulation and
ameliorate pulmonary fibrosis via the AMP-activated protein kinase
(AMPK)/microRNA-27a/peroxisome proliferator-activated receptor γ
axis (158). Additionally, the
GLP-1R/GCGR dual agonist TB001 alleviates liver fibrosis by
inhibiting the TGF-β/Smad2/3 signaling pathway and hepatic stellate
cell activation, while modulating NF-κB and JNK pathways to
suppress inflammation and apoptosis (159). Semaglutide directly targets
pancreatic cancer-associated fibroblasts, inhibiting the expression
and activity of collagen prolyl-4-hydroxylase (P4HA1), reducing
collagen hydroxylation and ECM deposition. This softens the
physical barrier of the tumor stroma, promotes intratumoral
infiltration of CD3+ and CD8+ T cells,
remodels the immunosuppressive microenvironment, and suppresses
pancreatic cancer progression (159).
Through inhibition of key targets, such as TGF-β and
P4HA1, GLP-1RAs exert antifibrotic and immunomodulatory effects,
demonstrating protective potential in multiple organs, including
the heart, kidneys, liver and lungs (160). Furthermore, their ability to
remodel the pancreatic cancer ECM and enhance T-cell infiltration
offers novel strategies for cancer immunotherapy (161). Future research should focus on
combination therapies of GLP-1RAs with chemotherapy and immune
checkpoint inhibitors, and further explore their therapeutic
potential in other stroma-rich tumors.
Despite the Warburg effect, mitochondrial function
remains crucial for tumor cell survival, biosynthesis and redox
homeostasis (162). Mitochondrial
DNA mutations or dysfunction can drive tumorigenesis (163). The aberrant proliferation of
tumor cells relies on mitochondrial integrity and tricarboxylic
acid (TCA) cycle activity (164).
Furthermore, cancer cells exploit mutations in multiple TCA cycle
enzymes to facilitate immunosuppression and immune evasion
(165).
GLP-1RAs exert protective effects on mitochondrial
function across multiple experimental models. GLP-1RAs enhance
endoplasmic reticulum-mitochondrial coupling, promote calcium
transfer from the endoplasmic reticulum to mitochondria and
increase mitochondrial activity in vascular smooth muscle cells
(166). Under high-glucose and
high-lipid stress, GLP-1RAs reduce apoptosis and mitigate
mitochondrial dysfunction in human microvascular endothelial cells
by inhibiting ERK1/2, JNK and p38 pathways, while suppressing
caspase-3 and PARP cleavage (167). Liraglutide treatment ameliorates
IL-1β-induced ROS production in cardiomyocytes, protects against
IL-1β-induced loss of mitochondrial membrane potential and
decreased ATP generation, and counteracts IL-1β-induced alterations
in AMPK and acetyl-CoA carboxylase phosphorylation as well as the
reduction of peroxisome proliferator-activated receptor γ
coactivator-1α (PGC-1α), which is a key regulator whose
downregulation leads to cardiac mitochondrial dysfunction (168). In hepatocytes, GLP-1 upregulates
uncoupling protein 2 mRNA expression, downregulates Bax and Bak
mRNA levels, enhances PGC-1α expression, improves mitochondrial
antioxidant capacity, and inhibits apoptosis (169). GLP-1RAs increase mitochondrial
content in the soleus muscle of SDT fatty rats by upregulating
citrate synthase activity and cytochrome c oxidase subunit
5B protein expression (170).
Additionally, in SH-SY5Y neuroblastoma cells, exendin-4 treatment
upregulates the mitochondrial quality control regulators Parkin and
PTEN induced kinase 1, reduces Bax expression, enhances Akt and
Bcl-2 levels, and inhibits tumor cell apoptosis (83). Through the PI3K/AKT pathway,
GLP-1RAs downregulate mitochondrial BCL2 interacting protein 3
expression, reduce doxorubicin-induced ROS production and γ-H2AX
elevation, and ameliorate doxorubicin-induced cardiotoxicity
(171).
Evidence indicates that GLP-1RAs preserve
mitochondrial function by enhancing calcium uptake, improving
membrane potential, increasing ATP production, promoting
mitochondrial biogenesis, and reducing oxidative stress and
apoptosis (172,173). Therefore, combining conventional
antitumor therapies with GLP-1RAs may not only enhance treatment
efficacy and reduce drug resistance but also mitigate adverse
effects.
The therapeutic potential of incretin and
GCG-related drugs is under active investigation. In the field of
oncology, GLP-1RAs have predominantly demonstrated anticancer
effects, although a limited number of studies suggest potential
tumor-promoting properties (13,75).
However, current human evidence is derived mainly from metabolic
trials and pharmacoepidemiologic studies conducted at approved
therapeutic doses, rather than from oncology trials specifically
designed to induce tumor apoptosis (174-176).
In these clinical settings, 3.0 mg liraglutide once daily and 2.4
mg semaglutide once weekly are associated predominantly with
gastrointestinal adverse events, including nausea, diarrhea,
vomiting and constipation, which are generally mild-to-moderate and
transient (75,177-179).
Meta-analysis has not shown a significant increase in pancreatitis
or pancreatic cancer with GLP-1RAs, although gallbladder-related
events such as cholelithiasis warrant attention (180). Accordingly, the cancer-specific
evidence discussed in the present review should be interpreted by
distinguishing mechanistic findings from preclinical models and
safety data derived from clinically approved regimens. Relevant
evidence is summarized in Table
SII (preclinical studies) (13,69,151,152,181-190)
and Table SIII (clinical studies)
(73,175,179,180,182,191-199).
Where available, adverse effect information from cited human
studies is also summarized in Table
SIII and briefly noted in the corresponding cancer-specific
subsections.
The prescribing information for GLP-1RA lists MTC
as a contraindication, based on a preclinical rodent study
demonstrating that GLP-1R activation elevates cAMP levels in rodent
thyroid C-cells, stimulates calcitonin release, and promotes C-cell
hyperplasia and MTC development (200). However, in papillary thyroid
carcinoma, neither insulin nor GLP-1RAs markedly influence tumor
cell proliferation (201). A case
report from the LY2189265 diabetes trial described a woman with
previously unrecognized MTC who received dulaglutide (2.0 mg once
weekly) for 6 months without a significant change in serum
calcitonin levels (202).
Nevertheless, this single case is insufficient to establish the
safety of GLP-1RAs in human MTC. A real-world study suggested that
over a 5-year period, GLP-1RAs were associated with a reduced risk
of prostate, lung and colon cancer compared with metformin, but
might be linked to an elevated risk of thyroid cancer (203). By contrast, a large cohort study
reported that GLP-1RA use was not associated with a significant
increase in thyroid cancer risk compared with dipeptidyl
peptidase-4 inhibitors (191).
Additionally, randomized trial data and subsequent systematic
reviews have not shown a clear increase in thyroid cancer risk with
GLP-1RA treatment (193,204). From a safety perspective, the
available semaglutide data indicate that adverse events are
predominantly gastrointestinal, most commonly nausea, diarrhea,
vomiting and constipation, without a clear thyroid-specific
carcinogenic signal (193).
In conclusion, current evidence has not established
a causal relationship between GLP-1RAs and thyroid cancer. While
rodent data support continued caution in MTC, GLP-1RAs should not
be regarded as an absolute contraindication in other thyroid cancer
subtypes, provided that treatment is individualized and appropriate
monitoring is undertaken.
An early investigation indicated that GLP-1R
stimulation does not affect pancreatic cancer cell proliferation
(205). Liraglutide enhances
chemosensitivity in pancreatic cancer: By inhibiting the NF-κB
pathway, it improves the response of gemcitabine-resistanT cells to
chemotherapy, and this sensitization effect has been validated in
animal models (181).
A large historical cohort study of 543,595 patients
demonstrated that GLP-1RA treatment over 7 years did not increase
the incidence of pancreatic cancer, confirming its safety profile
in this malignancy (179).
Similarly, a large retrospective cohort study reported that
GLP-1RAs were associated with a decreased incidence of pancreatic
cancer in patients with type 2 diabetes mellitus (206). Clinically, current evidence does
not indicate an increased risk of pancreatitis or pancreatic cancer
with GLP-1RAs. However, gallbladder-related adverse events,
particularly cholelithiasis, should be considered when interpreting
their overall safety profile (180). In summary, current evidence
supports the beneficial role of GLP-1RAs in both the prevention and
treatment of pancreatic cancer.
A translational case-control study revealed that
proteasome subunit α type-2 (PSMA2) expression was upregulated in
12 human cancer types based on The Cancer Genome Atlas database. In
T2D-associated cervical cancer, GLP-1R expression was positively
associated with PSMA2 levels, but this association was absent in
non-T2D cervical cancer specimens (182). Subsequent preclinical experiment
in the same study demonstrated that under high-glucose conditions,
exendin-4 reduced phosphorylation of P65 and IκB, downregulated
PSMA2 expression, inhibited the NF-κB pathway, suppressed cell
proliferation, and reduced tumor volume in vivo. These
findings suggest that hyperglycemia may promote cervical cancer
progression via PSMA2 upregulation, while exendin-4 counteracts
this effect (182). In
conclusion, GLP-1RAs inhibit cervical cancer growth under
hyperglycemic conditions. Because the current evidence in cervical
cancer remains largely mechanistic and translational, to the best
of our knowledge, no cancer-specific clinical adverse effect data
are currently available for GLP-1RAs in this setting. Further
epidemiological and clinical studies are warranted to validate
these observations.
A preclinical study demonstrated that liraglutide
and exenatide suppress HCC cell proliferation and induce autophagy.
Compared with liraglutide, exenatide exhibits stronger inhibition
of mTOR in HepG2 cells (174). In
a mouse model of non-alcoholic fatty liver disease-associated HCC
(207), liraglutide prevented HCC
progression by modulating blood glucose levels and ameliorating
steatosis and inflammation in non-tumorous liver lesions.
Furthermore, liraglutide enhances NK cell-mediated cytotoxicity
through suppression of the IL-6/STAT3 signaling pathway in HCC
cells, thereby promoting immune surveillance (121). Multiple retrospective studies
have confirmed that GLP-1RAs are associated with reduced risks of
long-term adverse hepatic outcomes in patients with metabolic
dysfunction-associated steatotic liver disease or T2D and cirrhosis
(208), as well as lowered risks
of HCC development and hepatic decompensation in patients with T2D
(175,209). Notably, these studies mainly
reported hepatic outcomes and cancer incidence rather than
treatment-emergent adverse events, and detailed adverse effect data
were not consistently available. Overall, through direct
anti-proliferative effects, metabolic regulation and
immunomodulation (121,174,207,209), GLP-1RAs demonstrate promising
potential for clinical translation in HCC management.
Although initial hypotheses suggested that GLP-1RAs
might promote CRC via the Wnt/β-catenin pathway, this was not
substantiated. Nevertheless, the study by Sun et al
(208) advised caution in using
GLP-1RAs in high-risk populations. However, subsequent research has
demonstrated beneficial effects of GLP-1RAs in CRC. Liraglutide
inhibits CRC cell proliferation, migration and invasion by
regulating the PI3K/Akt/mTOR pathway (210), and attenuates high
glucose-induced proliferation by suppressing bone morphogenetic
protein 4 expression (211).
Cohort studies and genetic data analyses further support the
preventive and therapeutic potential of GLP-1RAs against CRC
(75,212). The available epidemiologic
studies in CRC mainly addressed cancer incidence or risk reduction,
whereas treatment-related adverse effects were not systematically
reported.
In contrast to the relatively consistent beneficial
profile of GLP-1RAs in CRC, the role of GCG appears to be more
heterogeneous. Yagi et al (84) demonstrated that GCG promoted the
proliferation of HT29, SW480, CT26 and CMT93 cells through
GCGR-dependent AMPK deactivation and MAPK activation, using 1.0 nM
GCG replenished every 24 h for up to 72 h. By contrast, Xu et
al (213), using CT26 and
SW480 models together with endothelial C166 cells and
patient-derived xenografts, reported that GCG administered at 10 nM
in vitro and 20 µg per mouse daily in vivo for ~14
days inhibited tumor angiogenesis and vascular mimicry and enhanced
chemotherapy efficacy, without measurable differences in blood
glucose levels among groups. Taken together, these discrepant
findings likely reflect differences in tumor model, target cell
compartment, glycemic context and exposure conditions, indicating
that the effects of GCG in CRC are context-dependent. These
differences suggest that GCG cannot be uniformly characterized as
either pro-tumorigenic or antitumorigenic in CRC; rather, its
biological effects may shift according to metabolic milieu and
cellular target, which has important implications for interpreting
preclinical data and for the cautious evaluation of GLP-1/GCG dual
agonists in CRC.
The role of GLP-1RAs in breast cancer therapy
remains controversial. Exendin-4 activates the cAMP/CREB signaling
pathway, thereby inhibiting cell proliferation and promoting
apoptosis. This supports the view that GLP-1 acts as a potent
inducer of cAMP and an inhibitor of breast cancer cell
proliferation (12). However,
these effects are not uniform across experimental settings, and
liraglutide has also been reported to promote triple-negative
breast cancer progression through the NOX4/ROS/VEGF axis when
administered at 100 nM in vitro and 400 µg/kg in vivo
(13). Taken together, these
findings suggest that the effects of GLP-1RAs in breast cancer are
context-dependent and may vary according to tumor subtype and
exposure conditions, warranting caution when extrapolating
high-dose preclinical observations to clinical safety. However, at
present, clinical adverse effect data specific to breast cancer
treatment settings remain limited, and the available human evidence
is derived mainly from cancer risk analyses rather than
oncology-focused intervention studies.
Diabetes and obesity are established risk factors
for EC, with weight loss and glycemic control representing key
preventive strategies (85). Under
hyperglycemic conditions, exendin-4 partially restores cisplatin
sensitivity in Ishikawa and HEC1B EC cells, with treatment
performed at 10 nM for 48 h and accompanied by modulation of
apoptosis-related proteins (183). Semaglutide has been shown to
potentiate the antitumor effect of levonorgestrel in EC models,
including Ishikawa, Hec50 and KLE cells, as well as patient-derived
organoids from grade 1 endometrial carcinomas. Mechanistically,
this combination was associated with upregulation of nuclear and
membrane progesterone receptors and produced a more pronounced
reduction in viability than either agent alone; notably, the
organoid experiments were performed at 100 nM for 72 h, supporting
mechanistic interpretation of synergy but not direct extrapolation
to clinically established exposure margins (214). The GLP-1/GIP dual agonist
tirzepatide effectively reduced tumor growth in both obese and lean
mice, and improved obesity-associated serum adiponectin, leptin and
C-reactive protein levels in obese mice (69). Tirzepatide suppressed ErbB
signaling and glycolysis/gluconeogenesis in obese mice, while
enhancing O-linked glycosylation biosynthesis and phospholipase D
signaling in lean mice, collectively inhibiting EC progression
(69). However, not all
observations support a uniformly protective role in EC. In one
mechanistic study, GLP-1R expression was downregulated in EC
tissues and cells, and GLP1R activation was reported to inhibit EC
progression through activation of the cAMP/PKA pathway (184). Nevertheless, a later cohort study
in individuals with T2D suggested a higher risk of EC with
long-term use (>2 years) of GLP-1RAs (160). Thus, the role of GLP-1R signaling
in EC remains context-dependent and requires further
investigation.
A large cohort study of women with T2D revealed
that short-term use of GLP-1RAs and dipeptidyl peptidase-4 (DPP4)
inhibitors was not associated with reduced EC risk compared with
sulfonylureas. However, use for >2 years was associated with a
higher risk of EC for GLP-1RAs [hazard ratio (HR), 2.47; 95% CI,
1.37-4.43] and for DPP4 inhibitors (HR, 1.63; 95% CI 1.14-2.34)
(160). This observation reflects
a long-term cancer risk association rather than a
treatment-emergent adverse effect profile, and detailed adverse
event data were not reported in the cited study.
The mechanisms underlying these contradictory
effects remain unresolved. Studies on macrophages suggest that
GLP-1RAs promote M2 polarization, potentially favoring tumor
progression, which is a promising avenue for elucidating the dual
roles of GLP-1Ras (127-131,215).
Additionally, whether GIP receptor agonists can enhance the
anticancer efficacy of GLP-1RAs warrants investigation in cohort
and preclinical studies.
While responses to incretin-based therapies vary
across cancer types, analysis of existing data reveals several
shared mechanisms that may explain their particular potential in
obesity-associated cancers: i) Direct metabolic intervention: Tumor
cells, particularly those evolving in an obese microenvironment,
may exhibit heightened sensitivity to alterations in metabolic
conditions. By directly reducing circulating glucose and free fatty
acid levels, and potentially enhancing hepatic fatty acid oxidation
through GCGR agonism, these agents may deprive tumor cells of
essential nutrients, thereby interfering with metabolic
reprogramming processes (55,65,69,70).
ii) Reversal of hyperinsulinemia: This represents one of the
central mechanisms. Insulin and IGF-1 function as potent mitogenic
signals. GLP-1RAs may counteract growth-promoting milieu through
improved insulin sensitivity and reduced circulating insulin
levels, which is particularly relevant in obesity- and
diabetes-associated cancers, such as breast cancer, CRC and EC
(75,80,81,176). iii) Attenuation of chronic
inflammation: Chronic low-grade inflammation in obesity serves as a
catalyst for tumorigenesis. GLP-1RAs demonstrate anti-inflammatory
properties both systemically and within the TME (for example, by
modulating macrophages and T cells, and reducing NETs as described
in the present review), thereby suppressing the pro-tumor
inflammatory milieu (105,108-110,121).
iv) Modulation of adipokine secretion: Weight loss and adipose
tissue reduction induced by GLP-1RAs alter adipokine secretion
profiles, for example by shifting adiponectin- and leptin-related
signals, which can directly influence tumor cell proliferation and
invasion (69,131).
The present review systematically examines the
roles of incretin and GCG signaling in tumor biology and their
potential therapeutic value. By integrating fundamental research
with clinical evidence, the main conclusions are summarized
here.
Analysis of the basic biology of these three
hormones reveals their interconnected metabolic regulatory networks
mediated through shared signaling modules such as cAMP. Although
the fundamental mechanisms of GLP-1, GIP and GCG are outlined in
the present review, the subsequent discussion primarily focuses on
GLP-1RAs, since the direct oncologic evidence for GIP and GCG
remains comparatively limited.
Regarding the TME, current evidence indicates that
GLP-1RAs modulate antitumor immunity through multiple cellular
pathways. GLP-1RAs directly enhance NK cell cytotoxicity, promote
DC antigen presentation, and ameliorate the immunosuppressive
microenvironment by inhibiting NET formation and regulating
macrophage polarization, thereby offering novel perspectives for
combination immunotherapy. In addition, current evidence suggests
that GLP-1RAs support antitumor T-cell immunity mainly through
remodeling of the suppressive TME rather than through direct
restoration of exhausted intratumoral CD8+ T-cell
function (108-111).
GLP-1RAs also exert multidimensional and context-dependent effects
on tumor metabolism. In addition to restraining tumor cell
proliferation in selected preclinical models, they remodel the ECM
through anti-fibrotic actions, thereby facilitating immune cell
infiltration (156-159).
Simultaneously, their protective effects on
mitochondrial function may enhance sensitivity to conventional
therapy while potentially reducing treatment-related toxicity.
Current human safety data obtained under approved therapeutic
regimens are characterized predominantly by mild-to-moderate
gastrointestinal adverse events rather than generalized cytotoxic
toxicity, underscoring the need to distinguish preclinical
pro-apoptotic observations from clinically established safety
profiles.
Cancer-specific analyses confirm the
context-dependent nature of these effects. GLP-1RAs demonstrate
preventive and therapeutic potential in pancreatic cancer, hepatic
cancer and CRC, require caution in MTC, and exhibit paradoxical
effects in breast cancer and EC. The role of GCG further highlights
the influence of metabolic context on hormone function; however,
current evidence remains insufficient to support a direct antitumor
effect of GCGR activation alone.
In summary, targeting GLP-1R, GIPR and GCGR
signaling pathways provides novel directions for cancer therapy.
Future studies should prioritize investigating GIP and GCG in
oncology, while deepening the understanding of the dual effects of
GLP-1RAs, establishing effective biomarker systems and ultimately
enabling precise clinical applications.
Funding: No funding was received.
Not applicable.
MH drafted the manuscript. CJJ prepared the figures
and tables. CY discussed and critically revised the manuscript.
Data authentication is not applicable. All authors have read and
approved the final version of the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Shi R, Tang YQ and Miao H: Metabolism in
tumor microenvironment: Implications for cancer immunotherapy.
MedComm (2020). 1:47–68. 2020.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Pearson-Stuttard J, Papadimitriou N,
Markozannes G, Cividini S, Kakourou A, Gill D, Rizos EC, Monori G,
Ward HA, Kyrgiou M, et al: Type 2 diabetes and cancer: An umbrella
review of observational and mendelian randomization studies. Cancer
Epidemiol Biomarkers Prev. 30:1218–1228. 2021.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Mili N, Paschou SA, Goulis DG, Dimopoulos
MA, Lambrinoudaki I and Psaltopoulou T: Obesity, metabolic
syndrome, and cancer: Pathophysiological and therapeutic
associations. Endocrine. 74:478–497. 2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Andersen DK, Korc M, Petersen GM, Eibl G,
Li D, Rickels MR, Chari ST and Abbruzzese JL: Diabetes,
pancreatogenic diabetes, and pancreatic cancer. Diabetes.
66:1103–1110. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hu Y, Zeng N, Ge Y, Wang D, Qin X, Zhang
W, Jiang F and Liu Y: Identification of the shared gene signatures
and biological mechanism in type 2 diabetes and pancreatic cancer.
Front Endocrinol (Lausanne). 13(847760)2022.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Escala-Garcia M, Morra A, Canisius S,
Chang-Claude J, Kar S, Zheng W, Bojesen SE, Easton D, Pharoah PDP
and Schmidt MK: Breast cancer risk factors and their effects on
survival: A Mendelian randomisation study. BMC Med.
18(327)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Seino Y and Yabe D: Glucose-dependent
insulinotropic polypeptide and glucagon-like peptide-1: Incretin
actions beyond the pancreas. J Diabetes Investig. 4:108–130.
2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nauck MA, Quast DR, Wefers J and Meier JJ:
GLP-1 receptor agonists in the treatment of type 2
diabetes-state-of-the-art. Mol Metab. 46(101102)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhao S, Yan Z, Du Y, Li Z, Tang C, Jing L,
Sun L, Yang Q, Tang X, Yuan Y, et al: A GLP-1/glucagon
(GCG)/CCK2 receptors tri-agonist provides new therapy
for obesity and diabetes. Br J Pharmacol. 179:4360–4377.
2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bossart M, Wagner M, Elvert R, Evers A,
Hübschle T, Kloeckener T, Lorenz K, Moessinger C, Eriksson O,
Velikyan I, et al: Effects on weight loss and glycemic control with
SAR441255, a potent unimolecular peptide GLP-1/GIP/GCG receptor
triagonist. Cell Metab. 34:59–74.e10. 2022.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ligumsky H, Wolf I, Israeli S, Haimsohn M,
Ferber S, Karasik A, Kaufman B and Rubinek T: The peptide-hormone
glucagon-like peptide-1 activates cAMP and inhibits growth of
breast cancer cells. Breast Cancer Res Treat. 132:449–461.
2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liu ZZ, Duan XX, Yuan MC, Yu J, Hu X, Han
X, Lan L, Liu BW, Wang Y and Qin JF: Glucagon-like peptide-1
receptor activation by liraglutide promotes breast cancer through
NOX4/ROS/VEGF pathway. Life Sci. 294(120370)2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhang Y, Xu F, Liang H, Cai M, Wen X, Li X
and Weng J: Exenatide inhibits the growth of endometrial cancer
Ishikawa xenografts in nude mice. Oncol Rep. 35:1340–1348.
2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Mayendraraj A, Rosenkilde MM and Gasbjerg
LS: GLP-1 and GIP receptor signaling in beta cells-a review of
receptor interactions and co-stimulation. Peptides.
151(170749)2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Baggio LL and Drucker DJ: Biology of
incretins: GLP-1 and GIP. Gastroenterology. 132:2131–2157.
2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Gribble FM and Reimann F: Signalling in
the gut endocrine axis. Physiol Behav. 176:183–188. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Rorsman P and Ashcroft FM: Pancreatic
β-cell electrical activity and insulin secretion: Of mice and men.
Physiol Rev. 98:117–214. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Nauck MA, Kleine N, Orskov C, Holst JJ,
Willms B and Creutzfeldt W: Normalization of fasting hyperglycaemia
by exogenous glucagon-like peptide 1 (7-36 amide) in type 2
(non-insulin-dependent) diabetic patients. Diabetologia.
36:741–744. 1993.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Drucker DJ: Mechanisms of action and
therapeutic application of glucagon-like peptide-1. Cell Metab.
27:740–756. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Siraj MA, Mundil D, Beca S, Momen A,
Shikatani EA, Afroze T, Sun X, Liu Y, Ghaffari S, Lee W, et al:
Cardioprotective GLP-1 metabolite prevents ischemic cardiac injury
by inhibiting mitochondrial trifunctional protein-α. J Clin Invest.
130:1392–1404. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ussher JR and Drucker DJ: Glucagon-like
peptide 1 receptor agonists: Cardiovascular benefits and mechanisms
of action. Nat Rev Cardiol. 20:463–474. 2023.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Montrose-Rafizadeh C, Avdonin P, Garant
MJ, Rodgers BD, Kole S, Yang H, Levine MA, Schwindinger W and
Bernier M: Pancreatic glucagon-like peptide-1 receptor couples to
multiple G proteins and activates mitogen-activated protein kinase
pathways in Chinese hamster ovary cells. Endocrinology.
140:1132–1140. 1999.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Hilger D, Masureel M and Kobilka BK:
Structure and dynamics of GPCR signaling complexes. Nat Struct Mol
Biol. 25:4–12. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Holz GG: Epac: A new cAMP-binding protein
in support of glucagon-like peptide-1 receptor-mediated signal
transduction in the pancreatic beta-cell. Diabetes. 53:5–13.
2004.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Fletcher MM, Halls ML, Christopoulos A,
Sexton PM and Wootten D: The complexity of signalling mediated by
the glucagon-like peptide-1 receptor. Biochem Soc Trans.
44:582–588. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Merola A, Romagnolo A, Rosso M,
Lopez-Castellanos JR, Wissel BD, Larkin S, Bernardini A, Zibetti M,
Maule S, Lopiano L and Espay AJ: Orthostatic hypotension in
Parkinson's disease: Does it matter if asymptomatic? Parkinsonism
Relat Disord. 33:65–71. 2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu L, Rashid M and Wess J: Regulation of
GLP-1 and glucagon receptor function by β-arrestins in
metabolically important cell types. Biochemistry. 64:978–986.
2025.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zaïmia N, Obeid J, Varrault A, Sabatier J,
Broca C, Gilon P, Costes S, Bertrand G and Ravier MA: GLP-1 and GIP
receptors signal through distinct β-arrestin 2-dependent pathways
to regulate pancreatic β cell function. Cell Rep.
42(113326)2023.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Holmes B, Benavides-Serrato A, Freeman RS,
Landon KA, Bashir T, Nishimura RN and Gera J: mTORC2/AKT/HSF1/HuR
constitute a feed-forward loop regulating rictor expression and
tumor growth in glioblastoma. Oncogene. 37:732–743. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zheng Z, Zong Y, Ma Y, Tian Y, Pang Y,
Zhang C and Gao J: Glucagon-like peptide-1 receptor: Mechanisms and
advances in therapy. Signal Transduct Target Ther.
9(234)2024.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Gasbjerg LS, Helsted MM, Hartmann B,
Jensen MH, Gabe MBN, Sparre-Ulrich AH, Veedfald S, Stensen S, Lanng
AR, Bergmann NC, et al: Separate and combined glucometabolic
effects of endogenous glucose-dependent insulinotropic polypeptide
and glucagon-like peptide 1 in healthy individuals. Diabetes.
68:906–917. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Mcintosh CH, Widenmaier S and Kim SJ:
Glucose-dependent insulinotropic polypeptide signaling in
pancreatic β-cells and adipocytes. J Diabetes Investig. 3:96–106.
2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ding WG and Gromada J: Protein kinase
A-dependent stimulation of exocytosis in mouse pancreatic
beta-cells by glucose-dependent insulinotropic polypeptide.
Diabetes. 46:615–621. 1997.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kagdi S, Lyons SA and Beaudry JL: The
interplay of glucose-dependent insulinotropic polypeptide in
adipose tissue. J Endocrinol. 261(e230361)2024.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Ehses JA, Pelech SL, Pederson RA and
McIntosh CH: Glucose-dependent insulinotropic polypeptide activates
the Raf-Mek1/2-ERK1/2 module via a cyclic AMP/cAMP-dependent
protein kinase/Rap1-mediated pathway. J Biol Chem. 277:37088–37097.
2002.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Widenmaier SB, Ao Z, Kim SJ, Warnock G and
McIntosh CH: Suppression of p38 MAPK and JNK via Akt-mediated
inhibition of apoptosis signal-regulating kinase 1 constitutes a
core component of the beta-cell pro-survival effects of
glucose-dependent insulinotropic polypeptide. J Biol Chem.
284:30372–30382. 2009.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kim SJ, Winter K, Nian C, Tsuneoka M, Koda
Y and McIntosh CH: Glucose-dependent insulinotropic polypeptide
(GIP) stimulation of pancreatic beta-cell survival is dependent
upon phosphatidylinositol 3-kinase (PI3K)/protein kinase B (PKB)
signaling, inactivation of the forkhead transcription factor Foxo1,
and down-regulation of bax expression. J Biol Chem.
280:22297–22307. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Kim SJ, Nian C, Widenmaier S and McIntosh
CH: Glucose-dependent insulinotropic polypeptide-mediated
up-regulation of beta-cell antiapoptotic Bcl-2 gene expression is
coordinated by cyclic AMP (cAMP) response element binding protein
(CREB) and cAMP-responsive CREB coactivator 2. Mol Cell Biol.
28:1644–1656. 2008.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Asmar M, Asmar A, Simonsen L, Gasbjerg LS,
Sparre-Ulrich AH, Rosenkilde MM, Hartmann B, Dela F, Holst JJ and
Bülow J: The Gluco- and liporegulatory and vasodilatory effects of
glucose-dependent insulinotropic polypeptide (GIP) are abolished by
an antagonist of the human GIP receptor. Diabetes. 66:2363–2371.
2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Getty-Kaushik L, Song DH, Boylan MO,
Corkey BE and Wolfe MM: Glucose-dependent insulinotropic
polypeptide modulates adipocyte lipolysis and reesterification.
Obesity (Silver Spring). 14:1124–1131. 2006.PubMed/NCBI View Article : Google Scholar
|
|
42
|
No authors listed. Correction to:
‘Glucose-dependent insulinotropic polypeptide in incretin
physiology: Role in health and disease’. Endocr Rev.
46(954)2025.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Thorens B: Neuronal regulation of glucagon
secretion and gluconeogenesis. J Diabetes Investig. 13:599–607.
2022.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Lee YH, Wang MY, Yu XX and Unger RH:
Glucagon is the key factor in the development of diabetes.
Diabetologia. 59:1372–1375. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Capozzi ME, D'Alessio DA and Campbell JE:
. The past, present, and future physiology and pharmacology of
glucagon. Cell Metab. 34:1654–1674. 2022.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Li K, Qiu C, Sun P, Liu DC, Wu TJ, Wang K,
Zhou YC, Chang XA, Yin Y, Chen F, et al: Ets1-mediated acetylation
of FoxO1 Is critical for gluconeogenesis regulation during
feed-fast cycles. Cell Rep. 26:2998–3010.e5. 2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Dominy JE Jr, Lee Y, Jedrychowski MP, Chim
H, Jurczak MJ, Camporez JP, Ruan HB, Feldman J, Pierce K,
Mostoslavsky R, et al: The deacetylase Sirt6 activates the
acetyltransferase GCN5 and suppresses hepatic gluconeogenesis. Mol
Cell. 48:900–913. 2012.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Gertler AA and Cohen HY: SIRT6, a protein
with many faces. Biogerontology. 14:629–639. 2013.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Harney JA and Rodgers RL: Insulin-like
stimulation of cardiac fuel metabolism by physiological levels of
glucagon: Involvement of PI3K but not cAMP. Am J Physiol Endocrinol
Metab. 295:E155–E161. 2008.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Hansen LH, Gromada J, Bouchelouche P,
Whitmore T, Jelinek L, Kindsvogel W and Nishimura E:
Glucagon-mediated Ca2+ signaling in BHK cells expressing cloned
human glucagon receptors. Am J Physiol. 274:C1552–C1562.
1998.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Ozcan L, Wong CC, Li G, Xu T, Pajvani U,
Park SK, Wronska A, Chen BX, Marks AR, Fukamizu A, et al: Calcium
signaling through CaMKII regulates hepatic glucose production in
fasting and obesity. Cell Metab. 15:739–751. 2012.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Liu Y, Dentin R, Chen D, Hedrick S,
Ravnskjaer K, Schenk S, Milne J, Meyers DJ, Cole P, Yates J III, et
al: A fasting inducible switch modulates gluconeogenesis via
activator/coactivator exchange. Nature. 456:269–273.
2008.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Wondisford AR, Xiong L, Chang E, Meng S,
Meyers DJ, Li M, Cole PA and He L: Control of Foxo1 gene expression
by co-activator P300. J Biol Chem. 289:4326–4333. 2014.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Guzman CB, Zhang XM, Liu R, Regev A,
Shankar S, Garhyan P, Pillai SG, Kazda C, Chalasani N and Hardy TA:
Treatment with LY2409021, a glucagon receptor antagonist, increases
liver fat in patients with type 2 diabetes. Diabetes Obes Metab.
19:1521–1528. 2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Shi T, Fan GQ and Xiao SD: SIRT3 reduces
lipid accumulation via AMPK activation in human hepatic cells. J
Dig Dis. 11:55–62. 2010.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Perry RJ, Zhang D, Guerra MT, Brill AL,
Goedeke L, Nasiri AR, Rabin-Court A, Wang Y, Peng L, Dufour S, et
al: Glucagon stimulates gluconeogenesis by INSP3R1-mediated hepatic
lipolysis. Nature. 579:279–283. 2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Müller TD, Finan B, Clemmensen C, DiMarchi
RD and Tschöp MH: The new biology and pharmacology of glucagon.
Physiol Rev. 97:721–766. 2017.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kim H, Mendez R, Chen X, Fang D and Zhang
K: Lysine acetylation of CREBH regulates fasting-induced hepatic
lipid metabolism. Mol Cell Biol. 35:4121–4134. 2015.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Rodgers RL: Glucagon, cyclic AMP, and
hepatic glucose mobilization: A half-century of uncertainty.
Physiol Rep. 10(e15263)2022.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Wang H, Zhao M, Sud N, Christian P, Shen
J, Song Y, Pashaj A, Zhang K, Carr T and Su Q: Glucagon regulates
hepatic lipid metabolism via cAMP and Insig-2 signaling:
Implication for the pathogenesis of hypertriglyceridemia and
hepatic steatosis. Sci Rep. 6(32246)2016.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Winther JB and Holst JJ: Glucagon agonism
in the treatment of metabolic diseases including type 2 diabetes
mellitus and obesity. Diabetes Obes Metab. 26:3501–3512.
2024.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Wawrzak-Pienkowska K, Pienkowski T,
Golonko A, Krupa A, Warpechowski M, Plonski AF, Mojsak P, Daniluk
J, Dabrowski A and Kurek K: Incretin signaling at the crossroads of
metabolism, inflammation, and tumorigenesis: Implications for
obesity patients. Eur J Pharmacol. 1007(178290)2025.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Miousse IR: GLP-1 receptor agonists in the
context of cancer: The road ahead. Am J Physiol Cell Physiol.
328:C1822–C1828. 2025.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Spezani R and Mandarim-de-Lacerda CA: The
current significance and prospects for the use of dual receptor
agonism GLP-1/Glucagon. Life Sci. 288(120188)2022.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Zhou J, Cai X, Huang X, Dai Y, Sun L,
Zhang B, Yang B, Lin H, Huang W and Qian H: A novel glucagon-like
peptide-1/glucagon receptor dual agonist exhibits weight-lowering
and diabetes-protective effects. Eur J Med Chem. 138:1158–1169.
2017.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Del Prato S, Gallwitz B, Holst JJ and
Meier JJ: The incretin/glucagon system as a target for
pharmacotherapy of obesity. Obes Rev. 23(e13372)2022.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Parker VER, Robertson D, Erazo-Tapia E,
Havekes B, Phielix E, de Ligt M, Roumans KHM, Mevenkamp J, Sjoberg
F, Schrauwen-Hinderling VB, et al: Cotadutide promotes
glycogenolysis in people with overweight or obesity diagnosed with
type 2 diabetes. Nat Metab. 5:2086–2093. 2023.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Sanchez-Pino MD, Gilmore LA, Ochoa AC and
Brown JC: Obesity-associated myeloid immunosuppressive cells, key
players in cancer risk and response to immunotherapy. Obesity
(Silver Spring). 29:944–953. 2021.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Kong W, Deng B, Shen X, John C, Haag J,
Sinha N, Lee D, Sun W, Chen S, Zhang H, et al: Tirzepatide as an
innovative treatment strategy in a pre-clinical model of
obesity-driven endometrial cancer. Gynecol Oncol. 191:116–123.
2024.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Marathe SJ, Grey EW, Bohm MS, Joseph SC,
Ramesh AV, Cottam MA, Idrees K, Wellen KE, Hasty AH, Rathmell JC
and Makowski L: Incretin triple agonist retatrutide (LY3437943)
alleviates obesity-associated cancer progression. NPJ Metab Health
Dis. 3(10)2025.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Huang X, Liu J, Peng G, Lu M, Zhou Z,
Jiang N and Yan Z: Gut hormone multi-agonists for the treatment of
type 2 diabetes and obesity: Advances and challenges. J Endocrinol.
262(e230404)2024.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Valencia-Rincón E, Rai R, Chandra V and
Wellberg EA: GLP-1 receptor agonists and cancer: Current clinical
evidence and translational opportunities for preclinical research.
J Clin Invest. 135(e194743)2025.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Wang L, Xu R, Kaelber DC and Berger NA:
Glucagon-like peptide 1 receptor agonists and 13 obesity-associated
cancers in patients with type 2 diabetes. JAMA Netw Open.
7(e2421305)2024.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Ko A, Chang YC, Bahar F, Wang TH,
Xanthavanij N, Yu CC, Hsieh RJ, See XY, Lo SW, Song J, et al: Risk
for cancer with glucagon-like peptide-1 receptor agonists and dual
agonists: A systematic review and meta-analysis. Ann Intern Med.
179:216–229. 2026.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Wang L, Wang W, Kaelber DC, Xu R and
Berger NA: GLP-1 receptor agonists and colorectal cancer risk in
drug-naive patients with type 2 diabetes, with and without
overweight/obesity. JAMA Oncol. 10:256–258. 2024.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Silverii GA, Marinelli C, Bettarini C, Del
Vescovo GG, Monami M and Mannucci E: GLP-1 receptor agonists and
the risk for cancer: A meta-analysis of randomized controlled
trials. Diabetes Obes Metab. 27:4454–4468. 2025.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Yamanouchi D: The roles of incretin
hormones GIP and GLP-1 in metabolic and cardiovascular health: A
comprehensive review. Int J Mol Sci. 27(27)2025.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Ji L, He X, Min X, Yang H, Wu W, Xu H,
Chen J and Mei A: Glucagon-like peptide-1 receptor agonists in
neoplastic diseases. Front Endocrinol (Lausanne).
15(1465881)2024.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Lin A, Ding Y, Li Z, Jiang A, Liu Z, Wong
HZH, Cheng Q, Zhang J and Luo P: Glucagon-like peptide 1 receptor
agonists and cancer risk: advancing precision medicine through
mechanistic understanding and clinical evidence. Biomark Res.
13(50)2025.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Elkind-Hirsch KE, Chappell N, Shaler D,
Storment J and Bellanger D: Liraglutide 3 mg on weight, body
composition, and hormonal and metabolic parameters in women with
obesity and polycystic ovary syndrome: A randomized
placebo-controlled-phase 3 study. Fertil Steril. 118:371–381.
2022.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Sandsdal RM, Juhl CR, Jensen SBK, Lundgren
JR, Janus C, Blond MB, Rosenkilde M, Bogh AF, Gliemann L, Jensen
JB, et al: Combination of exercise and GLP-1 receptor agonist
treatment reduces severity of metabolic syndrome, abdominal
obesity, and inflammation: A randomized controlled trial.
Cardiovasc Diabetol. 22(41)2023.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Muzurović EM, Volčanšek Š, Tomšić KZ,
Janež A, Mikhailidis DP, Rizzo M and Mantzoros CS: Glucagon-like
peptide-1 receptor agonists and dual glucose-dependent
insulinotropic polypeptide/glucagon-like peptide-1 receptor
agonists in the treatment of obesity/metabolic syndrome,
prediabetes/diabetes and non-alcoholic fatty liver disease-current
evidence. J Cardiovasc Pharmacol Ther.
27(10742484221146371)2022.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Pandey S, Mangmool S, Madreiter-Sokolowski
CT, Wichaiyo S, Luangmonkong T and Parichatikanond W: Exendin-4
protects against high glucose-induced mitochondrial dysfunction and
oxidative stress in SH-SY5Y neuroblastoma cells through GLP-1
receptor/Epac/Akt signaling. Eur J Pharmacol.
954(175896)2023.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Yagi T, Kubota E, Koyama H, Tanaka T,
Kataoka H, Imaeda K and Joh T: Glucagon promotes colon cancer cell
growth via regulating AMPK and MAPK pathways. Oncotarget.
9:10650–10664. 2018.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Carugno J, Marbin SJ, LaganÀ AS, Vitale
SG, Alonso L, DI Spiezio Sardo A and Haimovich S: New development
on hysteroscopy for endometrial cancer diagnosis: State of the art.
Minerva Med. 112:12–19. 2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Kundu M, Butti R, Panda VK, Malhotra D,
Das S, Mitra T, Kapse P, Gosavi SW and Kundu GC: Modulation of the
tumor microenvironment and mechanism of immunotherapy-based drug
resistance in breast cancer. Mol Cancer. 23(92)2024.PubMed/NCBI View Article : Google Scholar
|
|
87
|
De Visser KE and Joyce JA: The evolving
tumor microenvironment: From cancer initiation to metastatic
outgrowth. Cancer Cell. 41:374–403. 2023.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Li Z, Pai R, Gupta S, Currenti J, Guo W,
Di Bartolomeo A, Feng H, Zhang Z, Li Z, Liu L, et al: Presence of
onco-fetal neighborhoods in hepatocellular carcinoma is associated
with relapse and response to immunotherapy. Nat Cancer. 5:167–186.
2024.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Ward PS and Thompson CB: Metabolic
reprogramming: A cancer hallmark even warburg did not anticipate.
Cancer Cell. 21:297–308. 2012.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Ghesquière B, Wong BW, Kuchnio A and
Carmeliet P: Metabolism of stromal and immune cells in health and
disease. Nature. 511:167–176. 2014.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Jing X, Yang F, Shao C, Wei K, Xie M, Shen
H and Shu Y: Role of hypoxia in cancer therapy by regulating the
tumor microenvironment. Mol Cancer. 18(157)2019.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Colegio OR, Chu NQ, Szabo AL, Chu T,
Rhebergen AM, Jairam V, Cyrus N, Brokowski CE, Eisenbarth SC,
Phillips GM, et al: Functional polarization of tumour-associated
macrophages by tumour-derived lactic acid. Nature. 513:559–563.
2014.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Gabrilovich DI, Ostrand-Rosenberg S and
Bronte V: Coordinated regulation of myeloid cells by tumours. Nat
Rev Immunol. 12:253–268. 2012.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Macintyre AN, Gerriets VA, Nichols AG,
Michalek RD, Rudolph MC, Deoliveira D, Anderson SM, Abel ED, Chen
BJ, Hale LP and Rathmell JC: The glucose transporter Glut1 is
selectively essential for CD4 T cell activation and effector
function. Cell Metab. 20:61–72. 2014.PubMed/NCBI View Article : Google Scholar
|
|
95
|
van Vlerken-Ysla L, Tyurina YY, Kagan VE
and Gabrilovich DI: Functional states of myeloid cells in cancer.
Cancer Cell. 41:490–504. 2023.PubMed/NCBI View Article : Google Scholar
|
|
96
|
You S, Li S, Zeng L, Song J, Li Z, Li W,
Ni H, Xiao X, Deng W, Li H, et al: Lymphatic-localized Treg-mregDC
crosstalk limits antigen trafficking and restrains anti-tumor
immunity. Cancer Cell. 42:1415–1433.e12. 2024.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Zheng Y, Yao Y, Ge T, Ge S, Jia R, Song X
and Zhuang A: Amino acid metabolism reprogramming: Shedding new
light on T cell anti-tumor immunity. J Exp Clin Cancer Res.
42(291)2023.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Lucente D, Bellino S and La Salvia A:
GLP-1 receptor agonists in solid tumour therapy: Exploring their
anticancer potential and underlying molecular pathways. Genes
(Basel). 16(1352)2025.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Kounatidis D, Vallianou NG, Karampela I,
Rebelos E, Kouveletsou M, Dalopoulos V, Koufopoulos P,
Diakoumopoulou E, Tentolouris N and Dalamaga M: Anti-diabetic
therapies and cancer: From bench to bedside. Biomolecules.
14(1479)2024.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Ostroumov D, Fekete-Drimusz N, Saborowski
M, Kühnel F and Woller N: CD4 and CD8 T lymphocyte interplay in
controlling tumor growth. Cell Mol Life Sci. 75:689–713.
2018.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Farhood B, Najafi M and Mortezaee K:
CD8+ cytotoxic T lymphocytes in cancer immunotherapy: A
review. J Cell Physiol. 234:8509–8521. 2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Wei J, Zheng W, Chapman NM, Geiger TL and
Chi H: T cell metabolism in homeostasis and cancer immunity. Curr
Opin Biotechnol. 68:240–250. 2021.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Deng S, Chen Z and Shi Y: Roles of
glucagon-like peptide 1 receptor agonists in immune cell biology
and autoimmune/autoinflammatory diseases. Cell Biosci.
15(137)2025.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Ben Nasr M, Usuelli V, Dellepiane S,
Seelam AJ, Fiorentino TV, D'Addio F, Fiorina E, Xu C, Xie Y,
Balasubramanian HB, et al: Glucagon-like peptide 1 receptor is a T
cell-negative costimulatory molecule. Cell Metab. 36:1302–1319.e12.
2024.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Moschovaki Filippidou F, Kirsch AH, Thelen
M, Kétszeri M, Artinger K, Aringer I, Schabhüttl C, Mooslechner AA,
Frauscher B, Pollheimer M, et al: Glucagon-like peptide-1 receptor
agonism improves nephrotoxic serum nephritis by inhibiting T-cell
proliferation. Am J Pathol. 190:400–411. 2020.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Wong CK, Yusta B, Koehler JA, Baggio LL,
McLean BA, Matthews D, Seeley RJ and Drucker DJ: Divergent roles
for the gut intraepithelial lymphocyte GLP-1R in control of
metabolism, microbiota, and T cell-induced inflammation. Cell
Metab. 34:1514–1531.e7. 2022.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Wong CK and Drucker DJ: Antiinflammatory
actions of glucagon-like peptide-1-based therapies beyond metabolic
benefits. J Clin Invest. 135(e194751)2025.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Stanisavljevic I, Pavlovic S, Simovic
Markovic B, Jurisevic M, Krajnovic T, Mijatovic S, Spasojevic M,
Mitrovic S, Corovic I and Jovanovic I: Semaglutide decelerates the
growth and progression of breast cancer by enhancing the acquired
antitumor immunity. Biomed Pharmacother. 181(117668)2024.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Chen D, Li Q, Liang H, Huang L, Zhou H,
Zheng X and Wang Z: Exenatide enhanced the antitumor efficacy on
PD-1 blockade by the attenuation of neutrophil extracellular traps.
Biochem Biophys Res Commun. 619:97–103. 2022.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Chen D, Liang H, Huang L, Zhou H and Wang
Z: Liraglutide enhances the effect of checkpoint blockade in lung
and liver cancers through the inhibition of neutrophil
extracellular traps. FEBS Open Bio. 14:1365–1377. 2024.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Pachimatla AG, Fitzgerald B, Ogidigo J,
Bhatia M, Smith RJ Jr, Ratnakaram K, Kalvapudi S, Vedire Y,
Washington D, Vethanayagam Rr R, et al: Glucagon-like peptide-1
receptor agonism improves lung cancer outcomes and tumor growth
control. JCI insight. 10(e195484)2025.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Fridlender ZG, Sun J, Kim S, Kapoor V,
Cheng G, Ling L, Worthen GS and Albelda SM: Polarization of
tumor-associated neutrophil phenotype by TGF-beta: ‘N1’ versus ‘N2’
TAN. Cancer Cell. 16:183–194. 2009.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Teijeira Á, Garasa S, Gato M, Alfaro C,
Migueliz I, Cirella A, de Andrea C, Ochoa MC, Otano I, Etxeberria
I, et al: CXCR1 and CXCR2 chemokine receptor agonists produced by
tumors induce neutrophil extracellular traps that interfere with
immune cytotoxicity. Immunity. 52:856–871.e8. 2020.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Kaltenmeier C, Yazdani HO, Morder K,
Geller DA, Simmons RL and Tohme S: Neutrophil extracellular traps
promote T cell exhaustion in the tumor microenvironment. Front
Immunol. 12(785222)2021.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Guo J, Shu T, Zhang H, Huang N, Ren J, Lin
L, Wu J, Wang Y, Huang Z, Bin J, et al: A combined model of serum
neutrophil extracellular traps, CD8+ T cells, and tumor
proportion score provides better prediction of PD-1 inhibitor
efficacy in patients with NSCLC. FEBS J. 291:3403–3416.
2024.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Coënon L, Geindreau M, Ghiringhelli F,
Villalba M and Bruchard M: Natural Killer cells at the frontline in
the fight against cancer. Cell Death Dis. 15(614)2024.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Chen S, Zhu H and Jounaidi Y:
Comprehensive snapshots of natural killer cells functions,
signaling, molecular mechanisms and clinical utilization. Signal
Transduct Target Ther. 9(302)2024.PubMed/NCBI View Article : Google Scholar
|
|
118
|
De Barra C, Khalil M, Mat A, O'Donnell C,
Shaamile F, Brennan K, O'Shea D and Hogan AE: Glucagon-like
peptide-1 therapy in people with obesity restores natural killer
cell metabolism and effector function. Obesity (Silver Spring).
31:1787–1797. 2023.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Ogura K, Sato-Matsushita M, Yamamoto S,
Hori T, Sasahara M, Iwakura Y, Saiki I, Tahara H and Hayakawa Y: NK
cells control tumor-promoting function of neutrophils in mice.
Cancer Immunol Res. 6:348–357. 2018.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Wang Y, Guo Z, Xu A, Fu Z, Hou Y, Tang K,
Li J, Zhao F and Chen L: Mechanistic insights into IL-6-mediated NK
cell dysfunction in NSCLC via the IRE1α-XBP1s-STAT3-UBE2S axis. NPJ
Precis Oncol. 9(361)2025.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Lu X, Xu C, Dong J, Zuo S, Zhang H, Jiang
C, Wu J and Wei J: Liraglutide activates nature killer
cell-mediated antitumor responses by inhibiting IL-6/STAT3
signaling in hepatocellular carcinoma. Transl Oncol.
14(100872)2021.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Li S, Yu J, Huber A, Kryczek I, Wang Z,
Jiang L, Li X, Du W, Li G, Wei S, et al: Metabolism drives
macrophage heterogeneity in the tumor microenvironment. Cell Rep.
39(110609)2022.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Laoui D, Van Overmeire E, Di Conza G,
Aldeni C, Keirsse J, Morias Y, Movahedi K, Houbracken I, Schouppe
E, Elkrim Y, et al: Tumor hypoxia does not drive differentiation of
tumor-associated macrophages but rather fine-tunes the M2-like
macrophage population. Cancer Res. 74:24–30. 2014.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Li M, Yang Y, Xiong L, Jiang P, Wang J and
Li C: Metabolism, metabolites, and macrophages in cancer. J Hematol
Oncol. 16(80)2023.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Cassetta L and Pollard JW: Targeting
macrophages: Therapeutic approaches in cancer. Nat Rev Drug Discov.
17:887–904. 2018.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Basak U, Sarkar T, Mukherjee S,
Chakraborty S, Dutta A, Dutta S, Nayak D, Kaushik S, Das T and Sa
G: Tumor-associated macrophages: An effective player of the tumor
microenvironment. Front Immunol. 14(1295257)2023.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Meurot C, Martin C, Sudre L, Breton J,
Bougault C, Rattenbach R, Bismuth K, Jacques C and Berenbaum F:
Liraglutide, a glucagon-like peptide 1 receptor agonist, exerts
analgesic, anti-inflammatory and anti-degradative actions in
osteoarthritis. Sci Rep. 12(1567)2022.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Vinué Á, Navarro J, Herrero-Cervera A,
García-Cubas M, Andrés-Blasco I, Martínez-Hervás S, Real JT, Ascaso
JF and González-Navarro H: The GLP-1 analogue lixisenatide
decreases atherosclerosis in insulin-resistant mice by modulating
macrophage phenotype. Diabetologia. 60:1801–1812. 2017.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Shiraishi D, Fujiwara Y, Komohara Y,
Mizuta H and Takeya M: Glucagon-like peptide-1 (GLP-1) induces M2
polarization of human macrophages via STAT3 activation. Biochem
Biophys Res Commun. 425:304–308. 2012.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Yang L, Chen L, Li D, Xu H and Chen J, Min
X, He M, Wu T, Zhong J, Yang H and Chen J: Effect of GLP-1/GLP-1R
on the polarization of macrophages in the occurrence and
development of atherosclerosis. Mediators Inflamm.
2021(5568159)2021.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Hogan AE, Gaoatswe G, Lynch L, Corrigan
MA, Woods C, O'Connell J and O'Shea D: Glucagon-like peptide 1
analogue therapy directly modulates innate immune-mediated
inflammation in individuals with type 2 diabetes mellitus.
Diabetologia. 57:781–784. 2014.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Wang S, Wang J, Chen Z, Luo J, Guo W, Sun
L and Lin L: Targeting M2-like tumor-associated macrophages is a
potential therapeutic approach to overcome antitumor drug
resistance. NPJ Precis Oncol. 8(31)2024.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Bruen R, Curley S, Kajani S, Crean D,
O'Reilly ME, Lucitt MB, Godson CG, McGillicuddy FC and Belton O:
Liraglutide dictates macrophage phenotype in apolipoprotein E null
mice during early atherosclerosis. Cardiovasc Diabetol.
16(143)2017.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Salkeni MA and Naing A: Interleukin-10 in
cancer immunotherapy: From bench to bedside. Trends Cancer.
9:716–725. 2023.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Bułdak Ł, Bołdys A, Skudrzyk E, Machnik G
and Okopień B: Liraglutide therapy in obese patients alters
macrophage phenotype and decreases their tumor necrosis factor
alpha release and oxidative stress markers-a pilot study.
Metabolites. 14(554)2024.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Rannikko JH and Hollmén M: Clinical
landscape of macrophage-reprogramming cancer immunotherapies. Br J
Cancer. 131:627–640. 2024.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Heras-Murillo I, Adán-Barrientos I, Galán
M, Wculek SK and Sancho D: Dendritic cells as orchestrators of
anticancer immunity and immunotherapy. Nat Rev Clin Oncol.
21:257–277. 2024.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Xiao Z, Wang R, Wang X, Yang H, Dong J, He
X, Yang Y, Guo J, Cui J and Zhou Z: Impaired function of dendritic
cells within the tumor microenvironment. Front Immunol.
14(1213629)2023.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Liu Z, Dai H, Cui X, Liu Y and Dong Z:
Importance of B cells (review). Int J Mol Med. 57(2)2026.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Fridman WH, Meylan M, Petitprez F, Sun CM,
Italiano A and Sautès-Fridman C: B cells and tertiary lymphoid
structures as determinants of tumour immune contexture and clinical
outcome. Nat Rev Clin Oncol. 19:441–457. 2022.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Yang Y, Chen X, Pan J, Ning H, Zhang Y, Bo
Y, Ren X, Li J, Qin S, Wang D, et al: Pan-cancer single-cell
dissection reveals phenotypically distinct B cell subtypes. Cell.
187:4790–4811.e22. 2024.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Sakimura C, Tanaka H, Okuno T, Hiramatsu
S, Muguruma K, Hirakawa K, Wanibuchi H and Ohira M: B cells in
tertiary lymphoid structures are associated with favorable
prognosis in gastric cancer. J Surg Res. 215:74–82. 2017.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Baggio LL, Holland D, Wither J and Drucker
DJ: Lymphocytic infiltration and immune activation in
metallothionein promoter-exendin-4 (MT-Exendin) transgenic mice.
Diabetes. 55:1562–1570. 2006.PubMed/NCBI View Article : Google Scholar
|
|
144
|
Jin HR, Wang J, Wang ZJ, Xi MJ, Xia BH,
Deng K and Yang JL: Lipid metabolic reprogramming in tumor
microenvironment: From mechanisms to therapeutics. J Hematol Oncol.
16(103)2023.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Moiz A, Filion KB, Tsoukas MA, Yu OHY,
Peters TM and Eisenberg MJ: The expanding role of GLP-1 receptor
agonists: A narrative review of current evidence and future
directions. EClinicalMedicine. 86(103363)2025.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Shigeoka T, Nomiyama T, Kawanami T,
Hamaguchi Y, Horikawa T, Tanaka T, Irie S, Motonaga R, Hamanoue N,
Tanabe M, et al: Activation of overexpressed glucagon-like
peptide-1 receptor attenuates prostate cancer growth by inhibiting
cell cycle progression. J Diabetes Investig. 11:1137–1149.
2020.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Eftekhari S, Montazeri H and Tarighi P:
Synergistic anti-tumor effects of Liraglutide, a glucagon-like
peptide-1 receptor agonist, along with Docetaxel on LNCaP prostate
cancer cell line. Eur J Pharmacol. 878(173102)2020.PubMed/NCBI View Article : Google Scholar
|
|
148
|
He W and Li J: Exendin-4 enhances
radiation response of prostate cancer. Prostate. 78:1125–1133.
2018.PubMed/NCBI View Article : Google Scholar
|
|
149
|
Krause GC, Lima KG, Dias HB, da Silva EFG,
Haute GV, Basso BS, Gassen RB, Marczak ES, Nunes RSB and de
Oliveira JR: Liraglutide, a glucagon-like peptide-1 analog, induce
autophagy and senescence in HepG2 cells. Eur J Pharmacol.
809:32–41. 2017.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Alanteet AA, Attia HA, Shaheen S, Alfayez
M and Alshanawani B: Anti-proliferative activity of glucagon-like
peptide-1 receptor agonist on obesity-associated breast cancer: The
impact on modulating adipokines' expression in adipocytes and
cancer cells. Dose Response. 19(1559325821995651)2021.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Zhao H, Wei R, Wang L, Tian Q, Tao M, Ke
J, Liu Y, Hou W, Zhang L, Yang J and Hong T: Activation of
glucagon-like peptide-1 receptor inhibits growth and promotes
apoptosis of human pancreatic cancer cells in a cAMP-dependent
manner. Am J Physiol Endocrinol Metab. 306:E1431–E1441.
2014.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Koehler JA, Kain T and Drucker DJ:
Glucagon-like peptide-1 receptor activation inhibits growth and
augments apoptosis in murine CT26 colon cancer cells.
Endocrinology. 152:3362–3372. 2011.PubMed/NCBI View Article : Google Scholar
|
|
153
|
Yuan Z, Li Y, Zhang S, Wang X, Dou H, Yu
X, Zhang Z, Yang S and Xiao M: Extracellular matrix remodeling in
tumor progression and immune escape: From mechanisms to treatments.
Mol Cancer. 22(48)2023.PubMed/NCBI View Article : Google Scholar
|
|
154
|
Saatci O, Kaymak A, Raza U, Ersan PG,
Akbulut O, Banister CE, Sikirzhytski V, Tokat UM, Aykut G, Ansari
SA, et al: Targeting lysyl oxidase (LOX) overcomes chemotherapy
resistance in triple negative breast cancer. Nat Commun.
11(2416)2020.PubMed/NCBI View Article : Google Scholar
|
|
155
|
Scavo MP, Lisco G, Depalo N, Rizzi F,
Volpe S, Arrè V, Carrieri L, Notarnicola M, De Nunzio V, Curri ML,
et al: Semaglutide modulates extracellular matrix production of
LX-2 cells via exosomes and improves metabolic
dysfunction-associated steatotic liver disease (MASLD). Int J Mol
Sci. 25(1493)2024.PubMed/NCBI View Article : Google Scholar
|
|
156
|
Robinson E, Cassidy RS, Tate M, Zhao Y,
Lockhart S, Calderwood D, Church R, McGahon MK, Brazil DP,
McDermott BJ, et al: Exendin-4 protects against post-myocardial
infarction remodelling via specific actions on inflammation and the
extracellular matrix. Basic Res Cardiol. 110(20)2015.PubMed/NCBI View Article : Google Scholar
|
|
157
|
Huang L, Lin T, Shi M, Chen X and Wu P:
Liraglutide suppresses production of extracellular matrix proteins
and ameliorates renal injury of diabetic nephropathy by enhancing
Wnt/β-catenin signaling. Am J Physiol Renal Physiol. 319:F458–F468.
2020.PubMed/NCBI View Article : Google Scholar
|
|
158
|
Liu J, Li X, Lu S, Zheng X, Zhang X and
Zhao W: Glucagon-like peptide-1 (GLP-1) improved diabetic lung
fibrosis via AMPK and microRNA-27a (miR-27a). Ann Transl Med.
9(492)2021.PubMed/NCBI View Article : Google Scholar
|
|
159
|
Song N, Xu H, Liu J, Zhao Q, Chen H, Yan
Z, Yang R, Luo Z, Liu Q, Ouyang J, et al: Design of a highly potent
GLP-1R and GCGR dual-agonist for recovering hepatic fibrosis. Acta
Pharm Sin B. 12:2443–2461. 2022.PubMed/NCBI View Article : Google Scholar
|
|
160
|
Yang F, Luo X, Li J, Lei Y, Zeng F, Huang
X, Lan Y and Liu R: Application of glucagon-like peptide-1 receptor
antagonists in fibrotic diseases. Biomed Pharmacother.
152(113236)2022.PubMed/NCBI View Article : Google Scholar
|
|
161
|
Cardoso LEM, Marinho TS, Martins FF,
Aguila MB and Mandarim-de-Lacerda CA: Treatment with semaglutide, a
GLP-1 receptor agonist, improves extracellular matrix remodeling in
the pancreatic islet of diet-induced obese mice. Life Sci.
319(121502)2023.PubMed/NCBI View Article : Google Scholar
|
|
162
|
Du H, Xu T, Yu S, Wu S and Zhang J:
Mitochondrial metabolism and cancer therapeutic innovation. Signal
Transduct Target Ther. 10(245)2025.PubMed/NCBI View Article : Google Scholar
|
|
163
|
Patel PS, Castelow C, Patel DS,
Bhattacharya SK, Kuscu C, Kuscu C, Makowski L, Eason JD and Bajwa
A: Mitochondrial role in oncogenesis and potential chemotherapeutic
strategy of mitochondrial infusion in breast cancer. Int J Mol Sci.
23(12993)2022.PubMed/NCBI View Article : Google Scholar
|
|
164
|
Zhang Y, Yan H, Wei Y and Wei X: Decoding
mitochondria's role in immunity and cancer therapy. Biochim Biophys
Acta Rev Cancer. 1879(189107)2024.PubMed/NCBI View Article : Google Scholar
|
|
165
|
Kiweler N, Delbrouck C, Pozdeev VI, Neises
L, Soriano-Baguet L, Eiden K, Xian F, Benzarti M, Haase L, Koncina
E, et al: Mitochondria preserve an autarkic one-carbon cycle to
confer growth-independent cancer cell migration and metastasis. Nat
Commun. 13(2699)2022.PubMed/NCBI View Article : Google Scholar
|
|
166
|
Morales PE, Torres G, Sotomayor-Flores C,
Peña-Oyarzún D, Rivera-Mejías P, Paredes F and Chiong M: GLP-1
promotes mitochondrial metabolism in vascular smooth muscle cells
by enhancing endoplasmic reticulum-mitochondria coupling. Biochem
Biophys Res Commun. 446:410–416. 2014.PubMed/NCBI View Article : Google Scholar
|
|
167
|
Liao P, Yang D, Liu D and Zheng Y: GLP-1
and ghrelin attenuate high glucose/high lipid-induced apoptosis and
senescence of human microvascular endothelial cells. Cell Physiol
Biochem. 44:1842–1855. 2017.PubMed/NCBI View Article : Google Scholar
|
|
168
|
Zhang L, Tian J, Diao S, Zhang G, Xiao M
and Chang D: GLP-1 receptor agonist liraglutide protects
cardiomyocytes from IL-1β-induced metabolic disturbance and
mitochondrial dysfunction. Chem Biol Interact.
332(109252)2020.PubMed/NCBI View Article : Google Scholar
|
|
169
|
Wang C, Li Q, Wang W, Guo L, Guo C, Sun Y
and Zhang J: GLP-1 contributes to increases in PGC-1α expression by
downregulating miR-23a to reduce apoptosis. Biochem Biophys Res
Commun. 466:33–99. 2015.PubMed/NCBI View Article : Google Scholar
|
|
170
|
Yamada S, Ogura Y, Inoue K, Tanabe J,
Sugaya T, Ohata K, Nagai Y, Natsuki Y, Hoshino S, Watanabe S, et
al: Effect of GLP-1 receptor agonist, liraglutide, on muscle in
spontaneously diabetic torii fatty rats. Mol Cell Endocrinol.
539(111472)2022.PubMed/NCBI View Article : Google Scholar
|
|
171
|
Li X, Luo W, Tang Y, Wu J, Zhang J, Chen
S, Zhou L, Tao Y, Tang Y, Wang F, et al: Semaglutide attenuates
doxorubicin-induced cardiotoxicity by ameliorating BNIP3-mediated
mitochondrial dysfunction. Redox Biol. 72(103129)2024.PubMed/NCBI View Article : Google Scholar
|
|
172
|
Old VJ, Davies MJ, Papamargaritis D,
Choudhary P and Watson EL: The effects of glucagon-like peptide-1
receptor agonists on mitochondrial function within skeletal muscle:
A systematic review. J Cachexia Sarcopenia Muscle.
16(e13677)2025.PubMed/NCBI View Article : Google Scholar
|
|
173
|
Luna-Marco C, de Marañon AM, Hermo-Argibay
A, Rodriguez-Hernandez Y, Hermenejildo J, Fernandez-Reyes M,
Apostolova N, Vila J, Sola E, Morillas C, et al: Effects of GLP-1
receptor agonists on mitochondrial function, inflammatory markers
and leukocyte-endothelium interactions in type 2 diabetes. Redox
Biol. 66(102849)2023.PubMed/NCBI View Article : Google Scholar
|
|
174
|
Krause GC, Lima KG, Levorse V, Haute GV,
Gassen RB, Garcia MC, Pedrazza L, Donadio MVF, Luft C and de
Oliveira JR: Exenatide induces autophagy and prevents the cell
regrowth in HepG2 cells. EXCLI J. 18:540–548. 2019.PubMed/NCBI View Article : Google Scholar
|
|
175
|
Wang L, Berger NA, Kaelber DC and Xu R:
Association of GLP-1 receptor agonists and hepatocellular carcinoma
incidence and hepatic decompensation in patients with type 2
diabetes. Gastroenterology. 167:689–703. 2024.PubMed/NCBI View Article : Google Scholar
|
|
176
|
Rothman SM, Yin H, Yu OHY, Pollak M and
Azoulay L: Incretin-based drugs and the incidence of endometrial
cancer among people with type 2 diabetes: Active comparator
new-user design. Drug Saf. 48:1023–1033. 2025.PubMed/NCBI View Article : Google Scholar
|
|
177
|
Kolotkin RL, Fujioka K, Wolden ML, Brett
JH and Bjorner JB: Improvements in health-related quality of life
with liraglutide 3.0 mg compared with placebo in weight management.
Clin Obes. 6:233–242. 2016.PubMed/NCBI View Article : Google Scholar
|
|
178
|
Wharton S, Calanna S, Davies M, Dicker D,
Goldman B, Lingvay I, Mosenzon O, Rubino DM, Thomsen M, Wadden TA
and Pedersen SD: Gastrointestinal tolerability of once-weekly
semaglutide 2.4 mg in adults with overweight or obesity, and the
relationship between gastrointestinal adverse events and weight
loss. Diabetes Obes Metab. 24:94–105. 2022.PubMed/NCBI View Article : Google Scholar
|
|
179
|
Dankner R, Murad H, Agay N, Olmer L and
Freedman LS: Glucagon-like peptide-1 receptor agonists and
pancreatic cancer risk in patients with type 2 diabetes. JAMA Netw
Open. 7(e2350408)2024.PubMed/NCBI View Article : Google Scholar
|
|
180
|
Monami M, Nreu B, Scatena A, Cresci B,
Andreozzi F, Sesti G and Mannucci E: Safety issues with
glucagon-like peptide-1 receptor agonists (pancreatitis, pancreatic
cancer and cholelithiasis): Data from randomized controlled trials.
Diabetes Obes Metab. 19:1233–1241. 2017.PubMed/NCBI View Article : Google Scholar
|
|
181
|
Zhao HJ, Jiang X, Hu LJ, Yang L, Deng LD,
Wang YP and Ren ZP: Activation of GLP-1 receptor enhances the
chemosensitivity of pancreatic cancer cells. J Mol Endocrinol.
64:103–113. 2020.PubMed/NCBI View Article : Google Scholar
|
|
182
|
Mao D, Cao H, Shi M, Wang CC, Kwong J, Li
JJX, Hou Y, Ming X, Lee HM, Tian XY, et al: Increased co-expression
of PSMA2 and GLP-1 receptor in cervical cancer models in type 2
diabetes attenuated by exendin-4: A translational case-control
study. EBioMedicine. 65(103242)2021.PubMed/NCBI View Article : Google Scholar
|
|
183
|
Zhang Y, Cheng J, Li J, He J, Li X and Xu
F: The GLP-1R agonist exendin-4 attenuates hyperglycemia-induced
chemoresistance in human endometrial cancer cells through
ROS-mediated mitochondrial pathway. Front Oncol.
11(793530)2021.PubMed/NCBI View Article : Google Scholar
|
|
184
|
Li W, Gu Y, Liu S, Ruan F and Lv W: GLP1R
inhibits the progression of endometrial carcinoma through
activation of cAMP/PKA pathway. J Clin Lab Anal.
36(e24604)2022.PubMed/NCBI View Article : Google Scholar
|
|
185
|
Byrd RA, Sorden SD, Ryan T, Pienkowski T,
LaRock R, Quander R, Wijsman JA, Smith HW, Blackbourne JL, Rosol
TJ, et al: Chronic toxicity and carcinogenicity studies of the
long-acting GLP-1 receptor agonist dulaglutide in rodents.
Endocrinology. 156:2417–2428. 2015.PubMed/NCBI View Article : Google Scholar
|
|
186
|
Mao T, Zhang C, Yang S, Bi Y, Li M and Yu
J: Semaglutide alters gut microbiota and improves NAFLD in db/db
mice. Biochem Biophys Res Commun. 710(149882)2024.PubMed/NCBI View Article : Google Scholar
|
|
187
|
Chen BD, Zhao WC, Jia QA, Zhou WY, Bu Y,
Wang ZZ, Wang F, Wu WJ and Wang Q: Effect of the GLP-1 analog
exendin-4 and oxaliplatin on intrahepatic cholangiocarcinoma cell
line and mouse model. Int J Mol Sci. 14:24293–24304.
2013.PubMed/NCBI View Article : Google Scholar
|
|
188
|
Kissow H, Hartmann B, Holst JJ, Viby NE,
Hansen LS, Rosenkilde MM, Hare KJ and Poulsen SS: Glucagon-like
peptide-1 (GLP-1) receptor agonism or DPP-4 inhibition does not
accelerate neoplasia in carcinogen treated mice. Regul Pept.
179:91–100. 2012.PubMed/NCBI View Article : Google Scholar
|
|
189
|
Iwaya C, Nomiyama T, Komatsu S, Kawanami
T, Tsutsumi Y, Hamaguchi Y, Horikawa T, Yoshinaga Y, Yamashita S,
Tanaka T, et al: Exendin-4, a glucagonlike peptide-1 receptor
agonist, attenuates breast cancer growth by inhibiting NF-κB
activation. Endocrinology. 158:4218–4232. 2017.PubMed/NCBI View Article : Google Scholar
|
|
190
|
Ligumsky H, Amir S, Arbel Rubinstein T,
Guion K, Scherf T, Karasik A, Wolf I and Rubinek T: Glucagon-like
peptide-1 analogs activate AMP kinase leading to reversal of the
Warburg metabolic switch in breast cancer cells. Med Oncol.
41(138)2024.PubMed/NCBI View Article : Google Scholar
|
|
191
|
Dai H, Li Y, Lee YA, Lu Y, George TJ,
Donahoo WT, Lee KP, Nakshatri H, Allen J, Guo Y, et al: GLP-1
receptor agonists and cancer risk in adults with obesity. JAMA
Oncol. 11:1186–1193. 2025.PubMed/NCBI View Article : Google Scholar
|
|
192
|
Pasternak B, Wintzell V, Hviid A, Eliasson
B, Gudbjörnsdottir S, Jonasson C, Hveem K, Svanström H, Melbye M
and Ueda P: Glucagon-like peptide 1 receptor agonist use and risk
of thyroid cancer: Scandinavian cohort study. BMJ.
385(e078225)2024.PubMed/NCBI View Article : Google Scholar
|
|
193
|
Bezin J, Gouverneur A, Pénichon M, Mathieu
C, Garrel R, Hillaire-Buys D, Pariente A and Faillie JL: GLP-1
receptor agonists and the risk of thyroid cancer. Diabetes Care.
46:384–390. 2023.PubMed/NCBI View Article : Google Scholar
|
|
194
|
Feier CVI, Vonica RC, Faur AM, Streinu DR
and Muntean C: Assessment of thyroid carcinogenic risk and safety
profile of GLP1-RA semaglutide (ozempic) therapy for diabetes
mellitus and obesity: A systematic literature review. Int J Mol
Sci. 25(4346)2024.PubMed/NCBI View Article : Google Scholar
|
|
195
|
Newsome PN, Buchholtz K, Cusi K, Linder M,
Okanoue T, Ratziu V, Sanyal AJ, Sejling AS and Harrison SA:
NN9931-4296 Investigators. A placebo-controlled trial of
subcutaneous semaglutide in nonalcoholic steatohepatitis. N Engl J
Med. 384:1113–1124. 2021.PubMed/NCBI View Article : Google Scholar
|
|
196
|
Abrahami D, Yin H, Yu OHY, Pollak MN and
Azoulay L: Incretin-based drugs and the incidence of colorectal
cancer in patients with type 2 diabetes. Epidemiology. 29:246–253.
2018.PubMed/NCBI View Article : Google Scholar
|
|
197
|
Figlioli G, Piovani D, Peppas S, Pugliese
N, Hassan C, Repici A, Lleo A, Aghemo A and Bonovas S:
Glucagon-like peptide-1 receptor agonists and risk of
gastrointestinal cancers: A systematic review and meta-analysis of
randomized controlled trials. Pharmacol Res.
208(107401)2024.PubMed/NCBI View Article : Google Scholar
|
|
198
|
Piccoli GF, Mesquita LA, Stein C, Aziz M,
Zoldan M, Degobi NAH, Spiazzi BF, Lopes Junior GL, Colpani V and
Gerchman F: Do GLP-1 receptor agonists increase the risk of breast
cancer? A systematic review and meta-analysis. J Clin Endocrinol
Metab. 106:912–921. 2021.PubMed/NCBI View Article : Google Scholar
|
|
199
|
Sun Y, Liu Y, Dian Y, Zeng F, Deng G and
Lei S: Association of glucagon-like peptide-1 receptor agonists
with risk of cancers-evidence from a drug target Mendelian
randomization and clinical trials. Int J Surg. 110:4688–4694.
2024.PubMed/NCBI View Article : Google Scholar
|
|
200
|
Wang L, Wang Q, Li L, Kaelber DC and Xu R:
Glucagon-like peptide-1 receptor agonists and pancreatic cancer
risk: Target trial emulation using real-world data. J Natl Cancer
Inst. 117:476–485. 2025.PubMed/NCBI View Article : Google Scholar
|
|
201
|
Bjerre Knudsen L, Madsen LW, Andersen S,
Almholt K, de Boer AS, Drucker DJ, Gotfredsen C, Egerod FL,
Hegelund AC, Jacobsen H, et al: Glucagon-like peptide-1 receptor
agonists activate rodent thyroid C-cells causing calcitonin release
and C-cell proliferation. Endocrinology. 151:1473–1486.
2010.PubMed/NCBI View Article : Google Scholar
|
|
202
|
He L, Zhang S, Zhang X, Liu R, Guan H and
Zhang H: Effects of insulin analogs and glucagon-like peptide-1
receptor agonists on proliferation and cellular energy metabolism
in papillary thyroid cancer. Onco Targets Ther. 10:5621–5631.
2017.PubMed/NCBI View Article : Google Scholar
|
|
203
|
Sherman SI, Kloos RT, Tuttle RM,
Pontecorvi A, Völzke H, Harper K, Vance C, Alston JT, Usborne AL,
Sloop KW and Lakshmanan M: No calcitonin change in a person taking
dulaglutide diagnosed with pre-existing medullary thyroid cancer.
Diabet Med. 35:381–385. 2018.PubMed/NCBI View Article : Google Scholar
|
|
204
|
Wang J and Kim CH: Differential risk of
cancer associated with glucagon-like peptide-1 receptor agonists:
Analysis of real-world databases. Endocr Res. 47:18–25.
2022.PubMed/NCBI View Article : Google Scholar
|
|
205
|
Espinosa De Ycaza AE, Brito JP, McCoy RG,
Shao H and Singh Ospina N: Glucagon-like peptide-1 receptor
agonists and thyroid cancer: A narrative review. Thyroid.
34:403–418. 2024.PubMed/NCBI View Article : Google Scholar
|
|
206
|
Koehler JA and Drucker DJ: Activation of
glucagon-like peptide-1 receptor signaling does not modify the
growth or apoptosis of human pancreatic cancer cells. Diabetes.
55:1369–1379. 2006.PubMed/NCBI View Article : Google Scholar
|
|
207
|
Kojima M, Takahashi H, Kuwashiro T, Tanaka
K, Mori H, Ozaki I, Kitajima Y, Matsuda Y, Ashida K, Eguchi Y and
Anzai K: Glucagon-like peptide-1 receptor agonist prevented the
progression of hepatocellular carcinoma in a mouse model of
nonalcoholic steatohepatitis. Int J Mol Sci.
21(5722)2020.PubMed/NCBI View Article : Google Scholar
|
|
208
|
Sun Y, Fan L, Meng J, Zhang F, Zhang D and
Mei Q: Should GLP-1 receptor agonists be used with caution in high
risk population for colorectal cancer? Med Hypotheses. 82:255–256.
2014.PubMed/NCBI View Article : Google Scholar
|
|
209
|
Elsaid MI, Li N, Firkins SA, Rustgi VK,
Paskett ED, Acharya C, Reddy KR, Chiang CW and Mumtaz K: Impacts of
glucagon-like peptide-1 receptor agonists on the risk of adverse
liver outcomes in patients with metabolic dysfunction-associated
steatotic liver disease cirrhosis and type 2 diabetes. Aliment
Pharmacol Ther. 59:1096–1110. 2024.PubMed/NCBI View Article : Google Scholar
|
|
210
|
Tong G, Peng T, Chen Y, Sha L, Dai H,
Xiang Y, Zou Z, He H and Wang S: Effects of GLP-1 receptor agonists
on biological behavior of colorectal cancer cells by regulating
PI3K/AKT/mTOR signaling pathway. Front Pharmacol.
13(901559)2022.PubMed/NCBI View Article : Google Scholar
|
|
211
|
Ma B, Wang X, Ren H, Li Y, Zhang H, Yang M
and Li J: High glucose promotes the progression of colorectal
cancer by activating the BMP4 signaling and inhibited by
glucagon-like peptide-1 receptor agonist. BMC Cancer.
23(594)2023.PubMed/NCBI View Article : Google Scholar
|
|
212
|
Zhu C, Lai Y, Liu C, Teng L, Zhu Y, Lin X,
Fu X, Lai Q, Liu S, Zhou X and Fang Y: Comprehensively prognostic
and immunological analyses of GLP-1 signaling-related genes in
pan-cancer and validation in colorectal cancer. Front Pharmacol.
15(1387243)2024.PubMed/NCBI View Article : Google Scholar
|
|
213
|
Xu Y, Ni F, Sun D, Peng Y, Zhao Y, Wu X,
Li S, Qi X, He X, Li M, et al: Glucagon enhances chemotherapy
efficacy by inhibition of tumor vessels in colorectal cancer. Adv
Sci (Weinh). 11(e2307271)2024.PubMed/NCBI View Article : Google Scholar
|
|
214
|
Hagemann AR, Hagemann IS, Mutch DG, Devor
EJ, Malmrose PK, Zhang Y, Morrison AM, Thiel KW and Leslie KK:
Enhancing progestin therapy with a glucagon-like peptide 1 agonist
for the conservative management of endometrial cancer. Cancers
(Basel). 17(598)2025.PubMed/NCBI View Article : Google Scholar
|
|
215
|
Bułdak Ł, Machnik G, Bułdak RJ, Łabuzek K,
Bołdys A, Belowski D, Basiak M and Okopień B: Exenatide (a GLP-1
agonist) expresses anti-inflammatory properties in cultured human
monocytes/macrophages in a protein kinase A and B/Akt manner.
Pharmacol Rep. 68:329–337. 2016.PubMed/NCBI View Article : Google Scholar
|