1. Introduction
Virulence factors, the molecular weapons used by
pathogenic microorganisms to survive and proliferate within a host
have been a subject of intense study since the inception of germ
theory. While traditionally defined within the context of
infectious disease as components that modulate host-microbe
interactions to enhance host damage (1,2), the
classification of these factors is becoming increasingly complex. A
classic example is the AB toxin family, such as Diphtheria toxin,
which possesses dual functions: An enzymatic ‘A’ fragment that
drives pathogenicity and a receptor-binding ‘B’ fragment that
facilitates delivery (3,4).
However, an increasing body of evidence suggests
that similar microbial components produced by the commensal
microbiome can influence the development and progression of
non-infectious diseases, including cancer (5,6). The
distinction between a helpful symbiont and a harmful pathogen is
often fluid. In the context of cancer, a once-benign microbe may
acquire ‘virulent’ behavior during dysbiosis, contributing to
carcinogenesis (7). Rather than
acting solely as direct carcinogens termed ‘oncomicrobes’ [for
example, Helicobacter pylori (H. pylori) and human
papillomavirus (HPV)] numerous microorganisms function as
‘complicit microbes’. These facilitators [for example,
Fusobacterium nucleatum (F. nucleatum)] do not
necessarily initiate cancer but create a microenvironment conducive
to tumor development (8,9).
The present review offers a unique perspective by
moving beyond a catalogue of microbial species to focus on their
functional machinery. It was examined how the microbiome's
‘toolkit’ actively participates in the ‘Hallmarks of Cancer’
framework proposed by Hanahan and Weinberg (10-12).
This framework, which originally focused on host cell genetics, now
formally recognizes the microbiome as a key enabling characteristic
of the tumor microenvironment (TME) (10-12).
Whether classified as oncomicrobes or complicit microbes, these
organisms exploit an arsenal of virulence factors: Adhesins, toxins
and enzymes to rewire host signaling, degrade physical barriers,
and suppress immune responses.
The present review is organized by the functional
class of the virulence factor rather than the specific microbe. A
comprehensive summary of these factors categorized by their
functional class is provided in Table
I. Bacterial adhesins as the initiators of malignant
interaction were first explored, followed by secreted toxins as
molecular weapons. It was then detailed how degradative enzymes act
as an extracellular ‘demolition crew’ and how viral oncoproteins
hijack cellular machinery. Finally, structural components were
analyzed, including extracellular vesicles and metabolites, as
environmental modulators, before synthesizing these mechanisms into
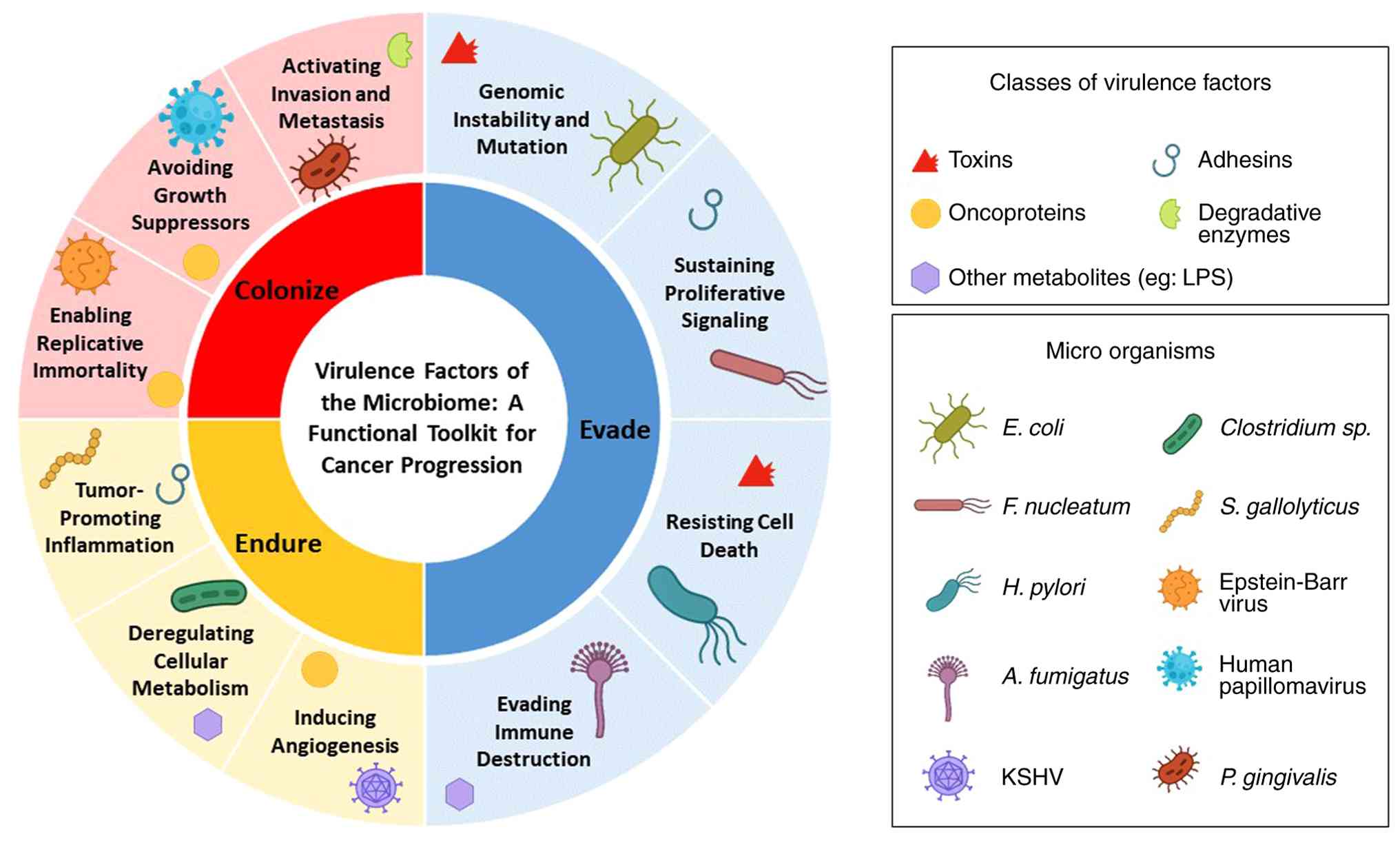
the ‘Evade-Endure-Colonize’ framework (illustrated in Fig. 1).
 | Table IKey microbial virulence factors and
their contributions to the hallmarks of cancer. |
Table I
Key microbial virulence factors and
their contributions to the hallmarks of cancer.
| Virulence factor
class | Specific virulence
factor | Microorganism | Host
target/mechanism of action | Consequence
(Hallmark of Cancer) | Associated
cancer(s) |
|---|
| Bacterial
adhesins | Fusobacterium
adhesin A | Fusobacterium
nucleatum | Binds E-cadherin,
leading to β-catenin release and nuclear translocation | Sustaining
proliferative signaling | Colorectal,
breast |
| | Fap2 | Fusobacterium
nucleatum | Binds TIGIT on
immune cells (immune evasion) and Gal-GalNAc on tumor cells
(metastasis) | Evading immune
destruction; activating invasion and metastasis | Colorectal,
breast |
| | Blood-group
antigen-binding adhesin/sialic acid-binding adhesin | Helicobacter
pylori | Bind Lewis b and
sialyl-Lewis x antigens for persistent colonization | Tumor-promoting
inflammation; enables toxin delivery | Gastric |
| Secreted
toxins | Colibactin | Escherichia
coli (pks+) | Alkylates host DNA,
causing double-strand breaks and a specific mutational
signature | Genomic instability
and mutation | Colorectal |
| | CagA | Helicobacter
pylori | Injected into
gastric cells; dysregulates signaling to disrupt cell polarity | Activating Invasion
and Metastasis (epithelial-mesenchymal tranistion) | Gastric |
| | Bacteroides
fragilis toxin | Bacteroides
fragilis (ETBF) | Metalloprotease
that cleaves E-cadherin, activating STAT3 signaling | Tumor-promoting
inflammation | Colorectal |
| Degradative
enzymes | Gingipains | Porphyromonas
gingivalis | Cysteine proteases
that degrade extracellular matrix components such as collagen and
fibronectin | Activating invasion
and metastasis | Oral,
pancreatic |
| | Collagenases | Clostridium,
Bacteroides | Degrade native
collagen fibers in the tumor stroma | Activating invasion
and metastasis | Pancreatic,
breast |
| Viral
oncoproteins | E6/E7 | Human
Papillomavirus | E6 degrades p53; E7
inactivates Rb | Resisting cell
death; avoiding growth suppressors | Cervical,
oropharyngeal |
| | Latent membrane
protein 1 | Epstein-Barr
virus | Constitutively
active CD40 mimic; activates NF-κB, JNK, and MAPK pathways | Enabling
replicative immortality | Lymphomas,
nasopharyngeal |
| Structural and
metabolic |
Lipopolysaccharide | Gram-negative
bacteria | Binds TLR4;
promotes inflammation and primes the lung pre-metastatic niche | Tumor-promoting
inflammation; activating invasion and metastasis | (Multiple
cancers) |
| | Fungal
β-glucans | Malassezia
species | Binds Dectin-1;
activates the complement cascade | Tumor-promoting
inflammation | Pancreatic |
| | Extracellular
Vesicles (microbial extracellular vesicles) | Various
bacteria | Long-distance
delivery of bioactive molecules; alters distant
microenvironments | Activating invasion
and metastasis | Lung |
| | Secondary bile
acids (for example, deoxycholic acid) | Clostridium
species | Induce reactive
oxygen species production, causing oxidative DNA damage | Genomic instability
and mutation | Colorectal |
2. Bacterial adhesins: Initiating the
malignant interaction
Bacterial adhesion represents the critical
initiating event in the interplay between the microbiome and
cancer. This molecular docking, mediated by surface proteins known
as adhesins, provides the anchor necessary for colonization and the
delivery of other virulence factors (13). Crucially, adhesion is not a passive
process; binding triggers signaling events that can directly
promote oncogenesis (14).
Fusobacterium adhesin A (FadA): A
direct activator of the Wnt/β-catenin pathway
The FadA protein, a signature virulence factor of
F. nucleatum, provides a direct link between a bacterial
protein and a core cancer pathway. FadA binds to E-cadherin on
colon epithelial cells (15),
triggering the internalization of the E-cadherin/β-catenin complex.
This releases β-catenin from the membrane, allowing it to
translocate to the nucleus and activate the Wnt signaling pathway.
This leads to the upregulation of oncogenes such as MYC and cyclin
D1 (CCND1), fueling the uncontrolled proliferation characteristic
of colorectal cancer (CRC) (16-19).
Fap2: A dual-function adhesin for
immune evasion and metastasis
F. nucleatum also employs Fap2, a protein
with dual pro-cancer functions. Firstly, Fap2 acts as an
immunomodulator by binding to the TIGIT receptor on natural killer
(NK) cells and cytotoxic T cells, delivering an inhibitory signal
that shields the tumor from immune destruction (20). Secondly, Fap2 functions as a
lectin, binding to Gal-GalNAc sugar moieties overexpressed on
cancer cells. This interaction allows F. nucleatum to
‘hitchhike’ on circulating tumor cells, facilitating their adhesion
to endothelial cells at distant sites and promoting metastasis
(21,22).
Blood-group antigen-binding adhesin
(BabA)/sialic acid-binding adhesin (SabA) adhesin duo
H. pylori employ a complementary pair of
adhesins. BabA binds to Lewis b (Leb) antigens on
healthy gastric cells, establishing the chronic infection required
for delivering the CagA oncoprotein (23,24).
As infection induces inflammation, the gastric environment changes,
and H. pylori switches to SabA. SabA binds to sialyl-Lewis x
(sLex) antigens, which are upregulated on inflamed
tissue, creating a feedback loop that perpetuates chronic
inflammation (25).
PilG and fimbrial adhesins
Streptococcus gallolyticus utilizes the PilG
adhesin to bind collagen types I and IV, which are exposed in the
disorganized TME but hidden in healthy tissue. This allows the
bacterium to preferentially colonize colorectal tumors (26,27).
Similarly, fimbrial adhesins such as FimA [Porphyromonas
gingivalis (P. gingivalis) and FimH [adherent-invasive
Escherichia coli (E. coli)] bind to host integrins
and CEACAM6, respectively. These interactions activate Toll-like
receptors (TLRs) or stabilize tumor cell adhesion, driving chronic
inflammation and invasion (28-30).
Collectively, these adhesins demonstrate how
microbes overcome the first hurdle of carcinogenesis: Physical
persistence. However, they do more than simply hold on. By
targeting molecules such as E-cadherin (FadA), TIGIT (Fap2) and
CEACAM6 (FimH), these adhesins directly engage the ‘Proliferative
Signaling’ and ‘Avoiding Immune Destruction’ Hallmarks. Within the
‘Evade-Endure-Colonize’ framework, adhesins serve as primary tools
for the ‘Colonize’ phase, allowing microbes to establish a foothold
in the tumor niche and physically bridge cancer cells to metastatic
sites.
3. Secreted toxins: Molecular weapons
targeting host pathways
Beyond adhesion, microbes deploy secreted toxins,
specialized weapons that manipulate host biology from a distance.
These can be broadly categorized as genotoxins (which damage DNA)
or modulating toxins (which hijack signaling) (31).
Genotoxins: Colibactin and Cyto-lethal
Distending Toxin (CDT)
Colibactin, produced by pks+ E. coli, is a
potent alkylating agent that creates DNA adducts, leading to
double-strand breaks. It leaves a specific ‘mutational signature’
in human CRC genomes, serving as a molecular fingerprint of
bacterial activity (32).
Similarly, the CDT, found in H. pylori (gastric cancer) and
E. coli (CRC), functions as a DNase. It translocates to the
nucleus and cleaves chromosomal DNA, triggering cell cycle arrest
and genomic instability (33,34).
H. pylori toxin arsenal (CagA, VacA
and Tipα)
H. pylori injects the CagA oncoprotein
directly into host cells, where it disrupts cell polarity and
promotes epithelial-mesenchymal transition (EMT) (35). Concurrently, the secreted VacA
toxin disrupts epithelial barrier integrity and suppresses local
T-cell function (36,37), while Tipα binds to STAT3, driving
inflammation and proliferation (38).
Modulating toxins: Bacteroides
fragilis toxin (BFT), CPAF and Cytotoxic Necrotizing Factor 1
(CNF1)
BFT is a metalloprotease that cleaves E-cadherin.
This disrupts the intestinal barrier and activates Wnt signaling,
while also recruiting T helper 17 (Th17) cells to establish a
pro-tumorigenic inflammatory environment (39,40).
In non-gastrointestinal cancers, Chlamydia trachomatis
secretes CPAF, a protease that degrades pro-apoptotic proteins and
cell cycle regulators, promoting survival in cervical cells
(41,42). Additionally, CNF1 from E.
coli activates Rho GTPases, driving cytoskeletal rearrangement
and motility (43,44).
While diverse in mechanism, these toxins converge
functionally to enable the ‘Genomic Instability’ and
‘Tumor-Promoting Inflammation’ Hallmarks. Genotoxins such as
Colibactin and CDT directly mutagenize the host genome, providing
the genetic variation required for tumor evolution (the Endure
phase). Meanwhile, modulating toxins such as CagA and BFT dismantle
cell-cell junctions and induce EMT. This plasticity is essential
for cancer cells to detach from the primary tumor, initiating the
Evade phase of metastasis.
4. Degradative enzymes: Extracellular
demolition crew
While toxins target intracellular pathways,
microbial degradative enzymes target the extracellular matrix
(ECM), the physical barrier to invasion (45).
Gingipains and hyaluronidases
Gingipains, cysteine proteases from P.
gingivalis, degrade collagen and fibronectin. In oral squamous
cell carcinoma, this activity breaks down the basement membrane,
paving the way for invasion (28,46).
Similarly, hyaluronidases secreted by Staphylococcus and
Clostridium species cleave hyaluronic acid. This ‘liquefies’
the ECM in diverse cancer settings - from skin and breast cancer to
urogenital tract malignancies, reducing physical resistance to
cancer cell migration and facilitating angiogenesis (47,48).
Collagenases: Breaching the
barrier
Collagenases from bacteria such as Clostridium
histolyticum degrade the dense collagen scaffold of the ECM. In
the TME, this activity assists cancer cells in breaching the tumor
capsule and entering the vasculature (49). Interestingly, this mechanism is
being explored therapeutically to ‘soften’ desmoplastic tumors
(such as pancreatic cancer) to improve drug delivery (50).
These enzymes function as the tumor's ‘demolition
crew’. By degrading the basement membrane and ECM, they directly
enable the ‘activating invasion and metastasis’ Hallmark. In the
metastatic cascade, these factors are critical for the transition
from the ‘Evade’ phase (local invasion) to the ‘Endure’ phase
(intravasation into blood vessels). Without this enzymatic
assistance, tumor cells would remain physically confined regardless
of their genetic mutations.
5. Viral oncoproteins: Master hijackers of
cellular machinery
Oncoviruses employ a strategy of genetic integration
and protein hijacking. Rather than damaging the cell from the
outside, viral oncoproteins seize control of core cellular
machinery (51,52).
HPV E6/E7 and HTLV-1 tax
The E6 and E7 proteins of high-risk HPV tear down
the p53 and Retinoblastoma (Rb) tumor suppressors, respectively.
This removes cell cycle checkpoints and prevents apoptosis, driving
the uncontrolled proliferation observed in cervical and
head-and-neck cancers (53,54).
The HTLV-1 Tax protein functions as a transcriptional activator,
driving the expression of IL-2 and its receptor to create a
malignant autocrine loop in T-cell leukemias (55).
EBV and hepatitis virus
oncoproteins
EBV's latent membrane protein 1 (LMP1) mimics a
constitutively active CD40 receptor, driving survival signaling via
NF-κB and MAPK pathways (56),
while EBV Nuclear Antigen 1 ensures viral persistence and immune
evasion (57). In liver cancer,
HBV X protein and HCV Core protein act as promiscuous regulators,
interacting with Wnt/β-catenin and generating reactive oxygen
species to promote both proliferation and genomic instability
(58-61).
Unlike bacteria that manipulate cells from the
exterior, oncoviruses bypass the ‘Evade’ phase and jump directly to
hijacking the cell's central command. By dismantling tumor
suppressors (p53 and Rb) and mimicking growth signals (vGPCR and
LMP1), these viral proteins directly enable the Hallmarks of
‘Enabling Replicative Immortality’ and ‘Sustaining Proliferative
Signaling’. This allows the infected cell to bypass natural
checkpoints, ensuring the survival and expansion required for the
‘Endure’ phase of malignancy.
6. Structural components and metabolites:
Environmental modulators
Beyond proteins, the microbiome influences cancer
through structural components and metabolic byproducts. These
factors modulate the ‘soil’ of the TME (62).
Structural components and
extracellular vesicles. Lipopolysaccharide (LPS) and
inflammation
LPS from Gram-negative bacteria activates TLR4,
driving NF-κB-mediated inflammation. Previous evidence highlights
the role of LPS in determining organotropism; circulating LPS can
‘prime’ the lungs for metastasis by upregulating inflammatory
adhesion molecules, creating a receptive ‘pre-metastatic niche’ for
breast cancer cells (63,64).
Fungal β-glucans: The microbiome is not
limited to bacteria; the fungal ‘mycobiome’ is also a key resident
of tumors. β-glucans, major structural components of the fungal
cell wall, are potent immunomodulators recognized by host receptors
such as Dectin-1(65). In
pancreatic cancer, fungi such as Malassezia migrate to the
pancreas, where their cell wall β-glucans activate the complement
cascade. This activation promotes inflammation and has been shown
to accelerate tumor progression (66).
Microbial extracellular vesicles (MEVs): An
increasing area of research focuses on MEVs - nanosized lipid
bilayers released by bacteria. These vesicles act as long-distance
delivery vehicles for virulence factors. In lung cancer, MEVs have
been shown to enter host cells and modulate signaling pathways that
suppress immune surveillance and promote EMT (67). Recent findings indicate that MEVs
can alter the lung microenvironment to favor tumor colonization,
representing a novel mechanism of host-microbe communication
(68).
Metabolites: Chemical language of
cancer
Secondary bile acids: Gut bacteria metabolize
primary bile acids into secondary forms such as deoxycholic acid.
In the liver, high levels of hydrophobic bile acids can induce DNA
damage and senescence in stellate cells, creating a
pro-inflammatory environment that facilitates hepatocellular
carcinoma and liver metastasis from CRC (69).
Short-chain fatty acids (SCFAs) and hydrogen
sulfide (H2S): While often protective, SCFAs
such as butyrate can be co-opted by cancer cells as an energy
source (Warburg effect) (70).
Similarly, H2S produced by F. nucleatum promotes
angiogenesis and fuels tumor cell mitochondrial metabolism
(71).
While proteins act as targeted weapons, these
structural and metabolic factors function as the ‘fertilizer’ for
the TME. By maintaining chronic inflammation (LPS and β-glucans)
and providing alternative fuel sources (SCFAs and H2S),
they enable the ‘Tumor-Promoting Inflammation’ and ‘Deregulating
Cellular Energetics’ hallmarks. Crucially, factors including
circulating LPS and MEVs act as long-range signals that prepare
distant organs for the ‘Colonize’ phase, establishing the
pre-metastatic niche before tumor cells arrive.
7. Future perspectives: Translating a
virulence-centric model into clinical impact
The shift to a virulence factor-centric framework
does more than reorganize the understanding of the microbiome's
role in cancer; it provides a direct, mechanistic roadmap for
intervention. This perspective aligns with the United Nations'
Sustainable Development Goal 3, specifically Target 3.4, which
calls for a one-third reduction in premature mortality from
non-communicable diseases, with cancer being a primary target
(72). By targeting the microbial
drivers of malignancy, the ‘toolkit’ described in the present
review, an entirely new front can be opened in this global
effort.
A new frontier in diagnostics:
Functional risk profiling
As understanding deepens, the future of cancer
diagnostics lies in assessing the functional threat posed by the
microbiome rather than just its taxonomic composition. The presence
of the pks genomic island (encoding Colibactin) or specific alleles
of fadA or vacA could serve as powerful prognostic biomarkers
(73,74). Detecting the DNA of these virulence
factors in a tumor biopsy or ‘liquid biopsy’ could identify
patients at high risk for metastasis. For example, quantifying
F. nucleatum load via fadA detection in blood shows promise
for screening and predicting recurrence in CRC. Furthermore, the
composition of the gut microbiome is now a validated predictive
biomarker for patient response to immune checkpoint inhibitors
(75). The future will involve
developing precisely defined microbial signatures to predict
treatment responses and determine if microbiome modulation is
required before therapy begins.
Pioneering a new generation of
therapeutic strategies
Targeting the microbial drivers of cancer represents
a paradigm shift, offering therapies that complement and enhance
traditional oncology. These strategies can be grouped into three
main categories:
Disarming the pathogen (anti-virulence
therapy): This precise approach aims to neutralize specific
virulence factors without inducing broad-spectrum dysbiosis.
Strategies include small molecule inhibitors designed to block the
active sites of microbial enzymes, such as gingipains or bacterial
collagenases, to prevent tissue invasion (76). Additionally, monoclonal antibodies
could block critical adhesin-receptor interactions; for instance,
blocking the Fap2 adhesin to prevent it from binding TIGIT on NK
cells could restore antitumor immunity (77).
Precision microbiome editing: This strategy
aims to selectively remove harmful ‘oncomicrobes’ or introduce
beneficial ones. Bacteriophage therapy offers a highly specific
method to eliminate bacteria such as F. nucleatum while
leaving the beneficial microbiota unharmed (78). Furthermore, the development of
preventative vaccines against oncogenic agents such as H.
pylori or EBV remains a major goal for long-term cancer
prevention (79).
Reshaping the ecosystem: This strategy aims
to engineer the microbial community to support cancer therapy. This
includes next-generation probiotics engineered to produce
anti-inflammatory molecules or compete with pathogenic species
(80). It also involves metabolic
interventions, such as using prebiotics to favor butyrate-producing
bacteria or drugs that inhibit the conversion of primary to
secondary bile acids (81).
Charting the path forward: Key
research frontiers
To translate these concepts into clinical reality,
research must move from establishing correlations to proving
causation. While animal models provide compelling evidence, a major
hurdle remains in definitively proving that a specific microbial
virulence factor is a driver, not just a ‘passenger’, in human
cancer. This will necessitate sophisticated multi-omics analyses of
large, longitudinal patient cohorts. Furthermore, recreating the
complexity of the TME including hypoxia, immune cell infiltrate,
and polymicrobial communities, through advanced co-culture systems
and human tumor organoids will be essential for validating these
new therapeutic targets.
8. Conclusion
The evidence presented in the present review
reframes the role of the microbiome from a passive bystander to an
active and versatile participant in cancer progression. By
dissecting the microbial arsenal through the lens of its virulence
factors, a clear picture emerges: The microbiome provides a
functional ‘toolkit’ that cancer cells exploit to acquire and
enhance the Hallmarks of Cancer. This analysis reveals a crucial
pattern of functional convergence, where diverse microbes
repeatedly target core host pathways including NF-κB, Wnt/β-catenin
and p53 to facilitate the ‘Evade, Endure and Colonize’ phases of
metastasis.
In essence, the tumor is not a solitary entity but a
malignant ecosystem. Understanding its non-host members is a
critical frontier in oncology. The intricate relationship between
the microbiome and cancer is no longer a niche interest but a
central theme in modern cancer biology. By continuing to unravel
the functions of the microbial virulence toolkit, a powerful new
pillar to our strategies for preventing and treating metastatic
disease is ready to be introduced, directly contributing to the
global goals of reducing cancer mortality.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
KV conceptualized the study, performed the
literature search, and was a major contributor in writing the
manuscript. BCP supervised the work and critically revised the
manuscript for intellectual content. All authors read and approved
the final version of the manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript or to generate images, and subsequently,
the authors revised and edited the content produced by the
artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
References
|
1
|
Casadevall A and Pirofski LA: Virulence
factors and their mechanisms of action: The view from a
damage-response framework. J Water Health. 7 (Suppl 1):S2–S18.
2009.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Méthot PO and Alizon S: What is a
pathogen? Toward a process view of host-parasite interactions.
Virulence. 5:775–785. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cherubin P, Quiñones B and Teter K:
Cellular recovery from exposure to sub-optimal concentrations of AB
toxins that inhibit protein synthesis. Sci Rep.
8(2494)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Song J: Bacterial AB toxins and
host-microbe interactions. Adv Microb Physiol. 81:67–109.
2022.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ong HS and Yim HCH: Microbial factors in
inflammatory diseases and cancers. Adv Exp Med Biol. 1024:153–174.
2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Aggarwal N, Kitano S, Puah GRY, Kittelmann
S, Hwang IY and Chang MW: Microbiome and human health: Current
understanding, engineering, and enabling technologies. Chem Rev.
123:31–72. 2022.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dzutsev A, Badger JH, Perez-Chanona E, Roy
S, Salcedo R, Smith CK and Trinchieri G: Microbes and Cancer. Annu
Rev Immunol. 35:199–228. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
El Tekle G, Andreeva N and Garrett WS: The
role of the microbiome in the etiopathogenesis of colon cancer.
Annu Rev Physiol. 86:453–478. 2024.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sepich-Poore GD, Zitvogel L, Straussman R,
Hasty J, Wargo JA and Knight R: The microbiome and human cancer.
Science. 371(eabc4552)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hanahan D: Hallmarks of cancer: New
dimensions. Cancer Discov. 12:31–46. 2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Di Martino P: Bacterial adherence: Much
more than a bond. AIMS Microbiol. 4:563–566. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Pizarro-Cerdá J and Cossart P: Bacterial
adhesion and entry into host cells. Cell. 124:715–727.
2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Fardini Y, Wang X, Témoin S, Nithianantham
S, Lee D, Shoham M and Han YW: Fusobacterium nucleatum adhesin FadA
binds vascular-endothelial cadherin and alters endothelial
integrity. Mol Microbiol. 82:1468–1480. 2011.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Rubinstein MR, Wang X, Liu W, Hao Y, Cai G
and Han YW: Fusobacterium nucleatum promotes colorectal
carcinogenesis by modulating E-cadherin/β-catenin signaling via its
FadA adhesin. Cell Host Microbe. 14:195–206. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Guo P, Tian Z, Kong X, Yang L, Shan X,
Dong B, Ding X, Jing X, Jiang C, Jiang N and Yu Y: FadA promotes
DNA damage and progression of Fusobacterium nucleatum-induced
colorectal cancer through up-regulation of chk2. J Exp Clin Cancer
Res CR. 39(202)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Dadgar-Zankbar L, Elahi Z, Shariati A,
Khaledi A, Razavi S and Khoshbayan A: Exploring the role of
Fusobacterium nucleatum in colorectal cancer: Implications for
tumor proliferation and chemoresistance. Cell Commun Signal.
22(547)2024.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Rezasoltani S, Shams E, Piroozkhah M, Aidi
Y, Azizmohammad Looha M, Bagheri P, Behzadi Andouhjerdi R, Sadeghi
A, Rejali L and Nazemalhosseini-Mojarad E: FadA antigen of
Fusobacterium nucleatum: implications for ceRNA network in
colorectal cancer and adenomatous polyps progression. Discov Oncol.
16(58)2025.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Gur C, Ibrahim Y, Isaacson B, Yamin R,
Abed J, Gamliel M, Enk J, Bar-On Y, Stanietsky-Kaynan N,
Coppenhagen-Glazer S, et al: Binding of the Fap2 protein of
fusobacterium nucleatum to human inhibitory receptor TIGIT protects
tumors from immune cell attack. Immunity. 42:344–355.
2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Abed J, Emgård JE, Zamir G, Faroja M,
Almogy G, Grenov A, Sol A, Naor R, Pikarsky E, Atlan KA, et al:
Fap2 mediates fusobacterium nucleatum colorectal adenocarcinoma
enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host
Microbe. 20:215–225. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Parhi L, Alon-Maimon T, Sol A, Nejman D,
Shhadeh A, Fainsod-Levi T, Yajuk O, Isaacson B, Abed J, Maalouf N,
et al: Breast cancer colonization by Fusobacterium nucleatum
accelerates tumor growth and metastatic progression. Nat Commun.
11(3259)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Rad R, Gerhard M, Lang R, Schöniger M,
Rösch T, Schepp W, Becker I, Wagner H and Prinz C: The Helicobacter
pylori blood group antigen-binding adhesin facilitates bacterial
colonization and augments a nonspecific immune response. J Immunol.
168:3033–3041. 2002.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ohno T, Vallström A, Rugge M, Ota H,
Graham DY, Arnqvist A and Yamaoka Y: Effects of blood group
antigen-binding adhesin expression during helicobacter pylori
infection of mongolian gerbils. J Infect Dis. 203:726–735.
2011.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ong LL and Lin CH: Adhesion, infection,
and therapeutic treatment of Helicobacter pylori: A review on
current aspects and future promise. Discov Appl Sci.
6(323)2024.
|
|
26
|
Pasquereau-Kotula E, Martins M, Aymeric L
and Dramsi S: Significance of Streptococcus gallolyticus subsp.
gallolyticus association with colorectal cancer. Front Microbiol.
9(614)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Abdulamir AS, Hafidh RR and Abu Bakar F:
The association of Streptococcus bovis/gallolyticus with colorectal
tumors: The nature and the underlying mechanisms of its etiological
role. J Exp Clin Cancer Res. 30(11)2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chopra A, Bhat SG and Sivaraman K:
Porphyromonas gingivalis adopts intricate and unique molecular
mechanisms to survive and persist within the host: A critical
update. J Oral Microbiol. 12(1801090)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sheikh A and Fleckenstein JM: Interactions
of pathogenic Escherichia coli with CEACAMs. Front Immunol.
14(1120331)2023.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Nguyen D, Smolchek RA, Uruena JM, Sawyer
WG and Jobin C: 70274 TL1 team approach to investigating the
adhesin gene fimH in adherent invasive E. coli induced inflammation
and colorectal cancer development. J Clin Transl Sci. 5 (Suppl
1):S107–S108. 2021.
|
|
31
|
Ivleva EA and Grivennikov SI:
Microbiota-driven mechanisms at different stages of cancer
development. Neoplasia. 32(100829)2022.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Sadeghi M, Mestivier D and Sobhani I:
Contribution of pks+ Escherichia coli (E. coli) to Colon
Carcinogenesis. Microorganisms. 12(1111)2024.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Guerra L, Cortes-Bratti X, Guidi R and
Frisan T: The biology of the cytolethal distending toxins. Toxins
(Basel). 3:172–190. 2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lai YR, Chang YF, Ma J, Chiu CH, Kuo ML
and Lai CH: From DNA damage to cancer progression: potential
effects of cytolethal distending toxin. Front Immunol.
12(760451)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ansari S and Yamaoka Y: Helicobacter
pylori virulence factor cytotoxin-associated gene A (CagA)-mediated
gastric pathogenicity. Int J Mol Sci. 21(7430)2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Boncristiano M, Paccani SR, Barone S,
Ulivieri C, Patrussi L, Ilver D, Amedei A, D'Elios MM, Telford JL
and Baldari CT: The helicobacter pylori vacuolating toxin inhibits
T cell activation by two independent mechanisms. J Exp Med.
198:1887–1897. 2003.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Palframan SL, Kwok T and Gabriel K:
Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori
pathogenesis. Front Cell Infect Microbiol. 2(92)2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Suganuma M, Watanabe T, Sueoka E, Lim IK
and Fujiki H: Role of TNF-α-inducing protein secreted by
helicobacter pylori as a tumor promoter in gastric cancer and
emerging preventive strategies. Toxins (Basel).
13(181)2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Fang Y, Yan C, Zhao Q, Xu J, Liu Z, Gao J,
Zhu H, Dai Z, Wang D and Tang D: The roles of microbial products in
the development of colorectal cancer: A review. Bioengineered.
12:720–735. 2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wu S, Rhee KJ, Albesiano E, Rabizadeh S,
Wu X, Yen HR, Huso DL, Brancati FL, Wick E, McAllister F, et al: A
human colonic commensal promotes colon tumorigenesis via activation
of T helper type 17 T cell responses. Nat Med. 15:1016–1022.
2009.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Patton MJ, McCorrister S, Grant C,
Westmacott G, Fariss R, Hu P, Zhao K, Blake M, Whitmire B, Yang C,
et al: Chlamydial protease-like activity factor and type III
secreted effectors cooperate in inhibition of p65 nuclear
translocation. mBio. 7:e01427–16. 2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Waguia Kontchou C, Gentle IE, Weber A,
Schoeniger A, Edlich F and Häcker G: Chlamydia trachomatis inhibits
apoptosis in infected cells by targeting the pro-apoptotic proteins
Bax and Bak. Cell Death Differ. 29:2046–2059. 2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Guo Y, Wang J, Zhou K, Lv J, Wang L, Gao
S, Keller ET, Zhang ZS, Wang Q and Yao Z: Cytotoxic necrotizing
factor 1 promotes bladder cancer angiogenesis through activating
RhoC. FASEB J. 34:7927–7940. 2020.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Travaglione S, Fabbri A and Fiorentini C:
The Rho-activating CNF1 toxin from pathogenic E. coli: A risk
factor for human cancer development? Infect Agent Cancer.
3(4)2008.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Radisky ES: Extracellular proteolysis in
cancer: Proteases, substrates, and mechanisms in tumor progression
and metastasis. J Biol Chem. 300(107347)2024.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Gnanasekaran J, Binder Gallimidi A, Saba
E, Pandi K, Eli Berchoer L, Hermano E, Angabo S, Makkawi HA,
Khashan A, Daoud A, et al: Intracellular porphyromonas gingivalis
promotes the tumorigenic behavior of pancreatic carcinoma cells.
Cancers (Basel). 12(2331)2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Alfano M, Canducci F, Nebuloni M, Clementi
M, Montorsi F and Salonia A: The interplay of extracellular matrix
and microbiome in urothelial bladder cancer. Nat Rev Urol.
13:77–90. 2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Parnigoni A, Moretto P, Viola M, Karousou
E, Passi A and Vigetti D: Effects of hyaluronan on breast cancer
aggressiveness. Cancers (Basel). 15(3813)2023.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Ding X, Ting NLN, Wong CC, Huang P, Jiang
L, Liu C, Lin Y, Li S, Liu Y, Xie M, et al: Bacteroides fragilis
promotes chemoresistance in colorectal cancer, and its elimination
by phage VA7 restores chemosensitivity. Cell Host Microbe.
33:941–956.e10. 2025.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ebelt ND, Zamloot V, Zuniga E, Passi KB,
Sobocinski LJ, Young CA, Blazar BR and Manuel ER:
Collagenase-expressing salmonella targets major collagens in
pancreatic cancer leading to reductions in immunosuppressive
subsets and tumor growth. Cancers (Basel). 3(3565)2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Elkhalifa AME, Nabi SU, Shah OS, Bashir
SM, Muzaffer U, Ali SI, Wani IA, Alzerwi NAN, Elderdery AY, Alanazi
A, et al: Insight into oncogenic viral pathways as drivers of viral
cancers: Implication for effective therapy. Curr Oncol.
30:1924–1944. 2023.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Schiller JT and Lowy DR: An introduction
to virus infections and human cancer. Recent Results Cancer Res.
217:1–11. 2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Pešut E, Đukić A, Lulić L, Skelin J, Šimić
I, Milutin Gašperov N, Tomaić V, Sabol I and Grce M: Human
papillomaviruses-associated cancers: An update of current
knowledge. Viruses. 13(2234)2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Skelin J, Sabol I and Tomaić V: Do or Die:
HPV E5, E6 and E7 in cell death evasion. Pathogens.
11(1027)2022.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Azran I, Schavinsky-Khrapunsky Y and Aboud
M: Role of Tax protein in human T-cell leukemia virus type-I
leukemogenicity. Retrovirology. 1(20)2004.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Sides MD, Klingsberg RC, Shan B, Gordon
KA, Nguyen HT, Lin Z, Takahashi T, Flemington EK and Lasky JA: The
epstein-barr virus latent membrane protein 1 and transforming
growth factor-β1 synergistically induce epithelial-mesenchymal
transition in lung epithelial cells. Am J Respir Cell Mol Biol.
44:852–862. 2011.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Münz C: Epstein-barr virus nuclear antigen
1: From immunologically invisible to a promising T cell target. J
Exp Med. 199:1301–1304. 2004.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Yang SZ, Zhang LD, Zhang Y, Xiong Y, Zhang
YJ, Li HL, Li XW and Dong JH: HBx protein induces EMT through c-Src
activation in SMMC-7721 hepatoma cell line. Biochem Biophys Res
Commun. 382:555–560. 2009.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Wang F, Song H, Xu F, Xu J, Wang L, Yang
F, Zhu Y and Tan G: Role of hepatitis B virus non-structural
protein HBx on HBV replication, interferon signaling, and
hepatocarcinogenesis. Front Microbiol. 14(1322892)2023.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Li T, Li D, Cheng L, Wu H, Gao Z, Liu Z,
Jiang W, Gao YH, Tian F, Zhao L and Wang S: Epithelial-mesenchymal
transition induced by hepatitis C virus core protein in
cholangiocarcinoma. Ann Surg Oncol. 17:1937–1944. 2010.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Mani H, Yen JH, Hsu HJ, Chang CC and Liou
JW: Hepatitis C virus core protein: Not just a nucleocapsid
building block, but an immunity and inflammation modulator. Tzu Chi
Med J. 34:139–147. 2021.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Pérez Escriva P, Correia Tavares
Bernardino C and Letellier E: De-coding the complex role of
microbial metabolites in cancer. Cell Rep.
44(115358)2025.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Wu X, Qian S, Zhang J, Feng J, Luo K, Sun
L, Zhao L, Ran Y, Sun L, Wang J and Xu F: Lipopolysaccharide
promotes metastasis via acceleration of glycolysis by the nuclear
factor-κB/snail/hexokinase3 signaling axis in colorectal cancer.
Cancer Metab. 9(23)2021.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Li S, Xu X, Jiang M, Bi Y, Xu J and Han M:
Lipopolysaccharide induces inflammation and facilitates lung
metastasis in a breast cancer model via the prostaglandin E2-EP2
pathway. Mol Med Rep. 11:4454–4462. 2015.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Batbayar S, Lee DH and Kim HW:
Immunomodulation of fungal β-Glucan in host defense signaling by
dectin-1. Biomol Ther (Seoul). 20:433–445. 2012.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Wang H, Capula M, Krom BP, Yee D,
Giovannetti E and Deng D: Of fungi and men: Role of fungi in
pancreatic cancer carcinogenesis. Ann Transl Med.
8(1257)2020.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Ke J, Zhang CJ, Wang LZ, Xie FS, Wu HY, Li
T, Bian CW and Wu RL: Lipopolysaccharide promotes cancer cell
migration and invasion through METTL3/PI3K/AKT signaling in human
cholangiocarcinoma. Heliyon. 10(e29683)2024.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Jang JY, Seo JH, Choi JJ, Ryu HJ, Yun H,
Ha DM and Yang J: Insight into microbial extracellular vesicles as
key communication materials and their clinical implications for
lung cancer (Review). Int J Mol Med. 56:1–11. 2025.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Nguyen TT, Ung TT, Kim NH and Jung YD:
Role of bile acids in colon carcinogenesis. World J Clin Cases.
6:577–588. 2018.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Donohoe DR, Collins LB, Wali A, Bigler R,
Sun W and Bultman SJ: The Warburg effect dictates the mechanism of
butyrate-mediated histone acetylation and cell proliferation. Mol
Cell. 48:612–626. 2012.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Davies J, Mayer MJ, Juge N, Narbad A and
Sayavedra L: Bacteroides thetaiotaomicron enhances H2S production
in Bilophila wadsworthia. Gut Microbes. 16(2431644)2024.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Sustainable Development Goals (SDG 3). U N
West Eur. Goal 3: Ensure healthy lives and promote well-being for
all at all ages. https://www.un.org/sustainabledevelopment/health/.
|
|
73
|
El Khadir M, Alaoui Boukhris S, Benajah
DA, El Rhazi K, Ibrahimi SA, El M, Harmouch T, Nejjari C, Mahmoud
M, Benlemlih M and Bennani B: VacA and CagA status as biomarker of
two opposite end outcomes of helicobacter pylori infection (Gastric
Cancer and Duodenal Ulcer) in a moroccan population. PLoS One.
12(e0170616)2017.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Zuraik AA, Daboul Y, Awama MA, Yazigi H,
Kayasseh MA and Georges M: Rapid detection of FadA in Fusobacterium
nucleatum using the quantitative LAMP colorimetric phenol red
method in stool samples from colorectal cancer patients. Sci Rep.
14(13739)2024.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Yan J, Yang L, Ren Q, Zhu C, Du H, Wang Z,
Qi Y, Xian X and Chen D: Gut microbiota as a biomarker and
modulator of antitumor immunotherapy outcomes. Front Immunol.
15(1471273)2024.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Olsen I and Potempa J: Strategies for the
inhibition of gingipains for the potential treatment of
periodontitis and associated systemic diseases. J Oral Microbiol:
Aug 18, 2014 (Epub ahead of print). doi: 10.3402/jom.v6.24800.
|
|
77
|
Tsao LC, Force J and Hartman ZC:
Mechanisms of therapeutic antitumor monoclonal antibodies. Cancer
Res. 81:4641–4651. 2021.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Hibstu Z, Belew H, Akelew Y and Mengist
HM: Phage therapy: A different approach to fight bacterial
infections. Biologics. 16:173–186. 2022.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Kohli AS, Sanyal S, Kaushal RS and Dwivedi
M: An insight into immunological therapeutic approach against
cancer: Potential anticancer vaccines. Curr Genomics. 26:175–190.
2025.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Abouelela ME and Helmy YA: Next-generation
probiotics as novel therapeutics for improving human health:
Current trends and future perspectives. Microorganisms.
12(430)2024.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Anwer EKE, Ajagbe M, Sherif M, Musaibah
AS, Mahmoud S, ElBanbi A and Abdelnaser A: Gut microbiota secondary
metabolites: Key roles in GI tract cancers and infectious diseases.
Biomedicines. 13(100)2025.PubMed/NCBI View Article : Google Scholar
|