Introduction
The benefit of bisphosphonates (BPs) is unequivocal
in the treatment of malignant bone neoplasias, such as multiple
myeloma, bone metastases (e.g., due to primary breast or prostate
cancer) or metabolic bone diseases, such as Paget’s disease and
severe osteoporosis.(1). Their
anti-resorptive mechanism results in a significant reduction of
skeletal-related events such as pain, hypercalcaemic episodes or
fractures that require radiation therapy or stabilizing operations,
thus improving the quality of life (2). BPs mainly inhibit bone resorption
through selective adsorption on mineral surfaces and subsequent
internalization via the bone-resorptive activity of osteoclasts
which interfere with various biochemical processes. The simpler,
non-nitrogen-containing BPs, such as clodronate, have the potential
to be metabolically incorporated into non-hydrolysable analogues of
adenosine triphosphate (ATP) that inhibit ATP-dependent
intracellular enzymes. By contrast, the more potent,
nitrogen-containing BPs, such as ibandronate, pamidronate and
zoledronate, inhibit farnesyl pyrophosphate synthase, a key enzyme
of the mevalonate pathway, thereby preventing the biosynthesis of
isoprenoid compounds that are essential for the post-translational
modification of small guanosine triphosphate (GTP)-binding proteins
(that are also GTPases), such as Rab, Rho and Rac (3–5). The
inhibition of protein prenylation and the disruption of the
function of these key regulatory proteins explain the loss of
osteoclast activity (6).
Besides the inhibition of osteoclast function, an
antiangiogenic effect of BPs has been throroughly investigated
(7–9). Sufficient tumor angiogenesis and
vessel formation is indispensable for tumor growth and progression
(10,11). In this context, the term
angiogenesis refers to sprouting from pre-existing vessels of
rather mature endothelial cells. Apart from this
bone-marrow-derived mechanism, circulating endothelial progenitor
cells (EPCs) bear the potential of the de novo creation of
primordial vessels, termed vasculogenesis (12–14).
Besides this biological function, EPCs demonstrated strong
paracrine effects in terms of the production of several cytokines
and growth factors, such as vascular endothelial growth factor
(VEGF), stromal cell-derived factor-1, insulin-like growth factor-1
and hepatocyte growth factor that increase angiogenesis and
neovascularization (15). EPCs have
been demonstrated to support short-, as well as long-term
neovascularization (14). The aim
of this study was to investigate the possible inhibitory effect of
different BPs on EPC-mediated neovascularization as a possible new
target for cancer therapy.
Materials and methods
EPC culture assay
Mononuclear cells (MNCs) were isolated by density
gradient centrifugation with Biocoll (Biochrom KG, Berlin, Germany)
from the peripheral blood of healthy human volunteers as previously
described (14). Immediately
following isolation, total MNCs (8×106 cells/ml medium)
were plated on 25-cm2 culture flasks coated with human
fibronectin (Sigma, Steinheim, Germany) and maintained in
endothelial basal medium (EBM) supplemented with EGM SingleQuots,
VEGF (100 ng/ml), and 20% FCS for 3 days. EPCs were obtained as a
byproduct from blood donors at our University hospital therefore no
approval from the ethics committee was required. This was an in
vitro study and only cell culture experiments were
conducted.
Migration assay
To examine the effect of bisphosphonates on EPC
migration, a 24-well Boyden chamber assay system (ThinCert™) was
used, according to the manufacturer’s instructions (Greiner
Bio-One, Frickenhausen, Germany). EPCs were incubated for 24 h with
varying concentrations of bisphosphonates. Cells were harvested,
washed twice in PBS and resuspended in EPC medium for adjustment to
a final concentration of 106 ml−1. EPCs were
stimulated to migrate from the upper to the lower chambers
subsequent to adding 10 ng/ml VEGF to the lower chambers. After 12
h, the cells were stained with fluorescent dye calcein-AM. The
culture medium was then removed from the inserts, while the inserts
were transferred onto a freshly prepared 24-well plate, containing
500 μl trypsin-EDTA/well. This plate was incubated for 10 min in a
cell culture incubator at 37°C with 5% CO2 with sporadic
agitation. The inserts were discarded and 200 μl of the
trypsin-EDTA solution, now containing the detached migratory cells,
was transferred to a flat-bottom, black 24-well plate. A
fluorescence plate reader was used for quantification.
Colony-forming unit assay
To examine the effect of the colony-forming ability,
EPCs were cultured in MethoCult™ GF H84434 culture medium (STEMCELL
Technologies, Inc., Vancouver, BC, Canada) on 35-mm well culture
plates for 14 days, according to the manufacturer’s instructions.
Cultures were incubated at 37°C with 5% CO2, and scored
for colony formation after 14 days.
Statistical analysis
Continuous variables were presented as the mean ±
SEM. Comparisons between groups were analyzed by analysis of
variance (ANOVA, post hoc test, Tukey) for experiments with >2
subgroups, or the Student’s t-test (two-sided). The software SPSS
16.0 for Windows was used for calculations. P<0.05 was
considered to indicate statistically significant differences.
Results
Migration
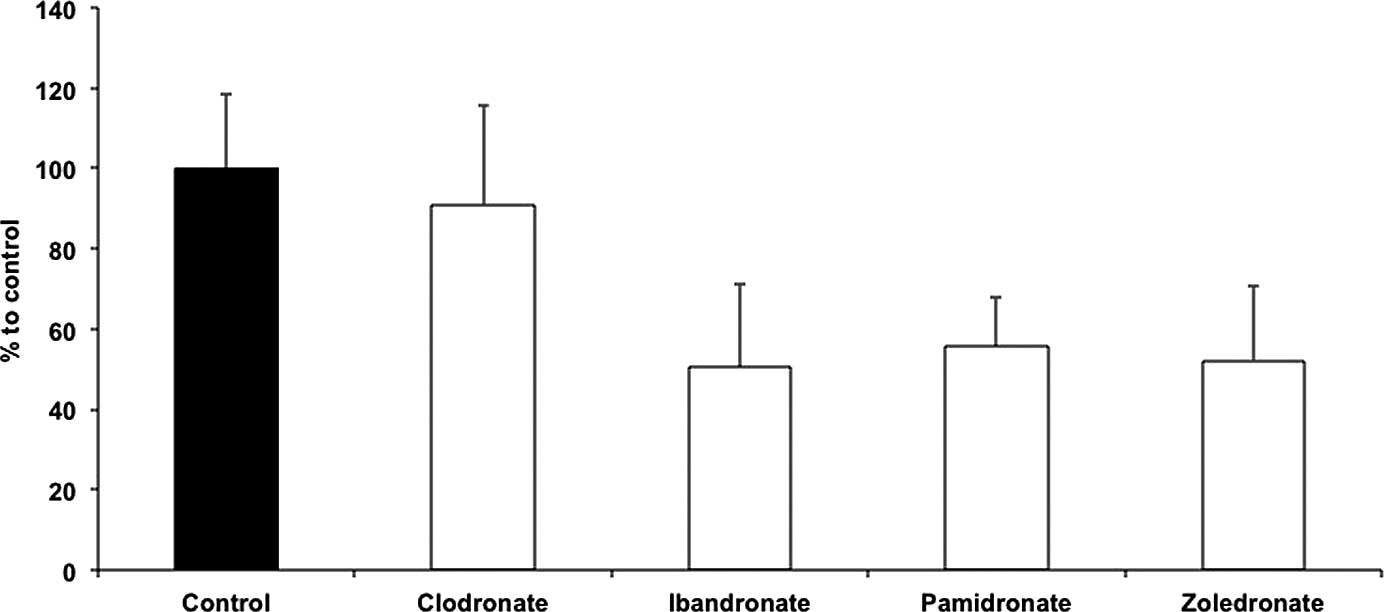
Results of the control group were set to 100%. EPC
migration was not reduced subsequent to clodronate treatment
(96.32±14.61%) compared to the control group. However, treatment
with ibandronate (45.5±6.53%), pamidronate (53.95±6.24%) and
zoledronate (51.29±5.07%) significantly affected the migration of
EPCs (P<0.01). No differences were observed in the migration
ability in the non-nitrogen BP clodronate group (Fig. 1).
Colony-forming units
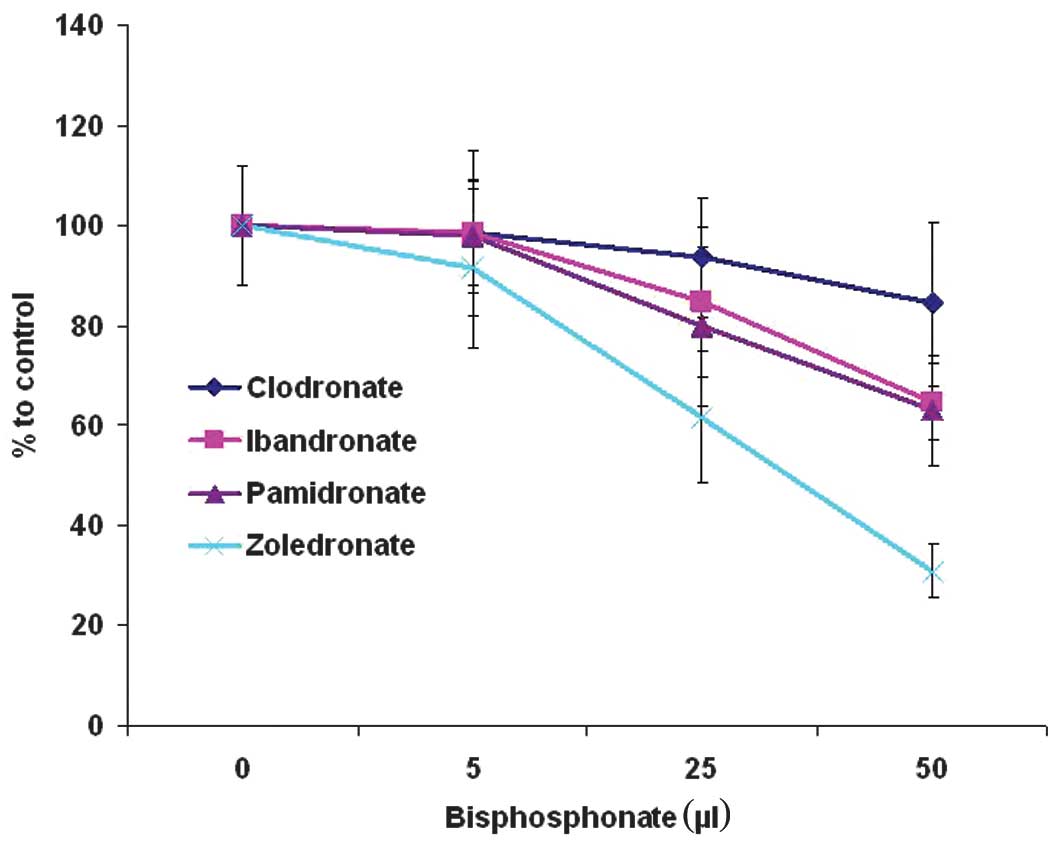
Fig. 2 shows the
colony-forming unit (CFU) ability during BP treatment. At a
concentration of 5 μmol BPs, no significant differences were
observed between the control group (57.59±0.55), clodronate
(55.22±0.44), ibandronate (55.11±3.97), pamidronate (54.88±3.27)
and zoledronate (51.22±1.92). Moreover, 25 μM clodronate
(53.22±4.2) did not demonstrate statistically significant
differences when compared to the control group. A tendency for
reduced CFU ability was observed at 25 μM ibandronate (48.22±5.71)
and pamidronate (45.44±4.77). Zoledronate (35.11±3.73) showed
statistically significant CFU results. Increased BP concentrations
of 50 μM showed notable inhibitory effects on CFU regarding
nitrogen-containing BPs ibandronate (38.66±1.52), pamidronate
(37.55±2.66) and zoledronate (18.55±2.11). No significant
difference was found between ibandronate and pamidronate. The most
marked negative effect on CFU was found for zoledronate, while the
lowest was for clodronate.
Discussion
BPs are common anti-bone-resorptive drugs with
clinical effectiveness against various types of cancer, when used
in combination with other conventional anticancer drugs (16–18).
Apart from their direct effect on cancer cells, these drugs are
believed to affect angiogenesis (19). Tumors and ischemic tissue have the
potential to recruit EPCs from the bone marrow in order to increase
the number of sprouting vessels for blood supply by indirect
effects, such as paracrine secretion of growth factors including
VEGF or the direct differentiation of EPCs in mature endothelial
cells (20, 21). In patients with head and neck
cancer, for example, an elevated EPC concentration in peripheral
venous blood was demonstrated (22).
Therefore, the effect of different BPs on EPCs as an
option for cancer therapy was investigated. Consistent with
previous studies, results showed that the EPC concentration is
reduced by different BPs in a dose-dependent manner (8,9).
Particularly, pamidronate and zoledronate had a significant impact
on cell numbers compared to the control and clodronate groups.
Yamada et al(8) showed
increased EPC apoptosis rate subsequent to zoledronate treatment
in vitro. In this study, important biological functions,
such as migration and stem cell differentiation were demonstrated
to be impaired by BPs. Reduced migration capacity resulted in
impaired mobilization from bone marrow niche and homing of EPC
(23,24). EPC differentiation is a
pre-condition for clonal expansion and stem cell function. The
nitrogen-containing BP zoledronate was found to have a significant
negative effect on colony formation, while no effect was detected
by the non-nitrogen containing BP clodronate. Nitrogen-containing
and non-nitrogen-containing BPs have different ways of action
resulting in varying effects of BPs. Non-nitrogen-containing BPs,
such as clodronate, are built into non-hydrolysable ATP that
inhibit many different ATP-dependent intracellular enzymes
(25). Therefore, extremely high
concentrations are needed to completely inhibit special pathways.
By contrast, the nitrogen-containing BPs, such as pamidronate and
zoledronate, inhibit farnesyl pyrophosphate synthase, a key enzyme
of the mevalonate pathway (26).
This pathway is significant for the production of small G proteins,
such as Ras, Rac and Rho proteins, which are important for
intracellular structure and mechanisms, such as intracellular
transport (27,28), and may contribute to the greater
impact of nitrogen-containing BPs.
In conclusion, within the boundaries of in
vitro studies, these data support the theory of the
anti-angiogenetic component of nitrogen-containing BPs.
Investigations regarding the effect of BPs on the biology of EPC
are likely to provide insights into the role of BPs in inhibiting
tumor vasculogenesis, as well as a possible therapeutic option in
the future.
References
|
1
|
Neville-Webbe HL and Coleman RE:
Bisphosphonates and RANK ligand inhibitors for the treatment and
prevention of metastatic bone disease. Eur J Cancer. 46:1211–1222.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Coleman RE: Metastatic bone disease:
clinical features, pathophysiology and treatment strategies. Cancer
Treat Rev. 27:165–176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rogers MJ: New insights into the molecular
mechanisms of action of bisphosphonates. Curr Pharm Des.
9:2643–2658. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rogers MJ, Gordon S, Benford HL, et al:
Cellular and molecular mechanisms of action of bisphosphonates.
Cancer. 88:2961–2978. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dunford JE, Thompson K, Coxon FP, et al:
Structure-activity relationships for inhibition of farnesyl
diphosphate synthase in vitro and inhibition of bone resorption in
vivo by nitrogen-containing bisphosphonates. J Pharmacol Exp Ther.
296:235–242. 2001.
|
|
6
|
Fisher JE, Rogers MJ, Halasy JM, et al:
Alendronate mechanism of action: geranylgeraniol, an intermediate
in the mevalonate pathway, prevents inhibition of osteoclast
formation, bone resorption, and kinase activation in vitro. Proc
Natl Acad Sci USA. 96:133–138. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Santini D, Vincenzi B, Avvisati G, et al:
Pamidronate induces modifications of circulating angiogenetic
factors in cancer patients. Clin Cancer Res. 8:1080–1084.
2002.PubMed/NCBI
|
|
8
|
Yamada J, Tsuno NH, Kitayama J, et al:
Anti-angiogenic property of zoledronic acid by inhibition of
endothelial progenitor cell differentiation. J Surg Res.
151:115–120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ziebart T, Pabst A, Klein MO, et al:
Bisphosphonates: restrictions for vasculogenesis and angiogenesis:
inhibition of cell function of endothelial progenitor cells and
mature endothelial cells in vitro. Clin Oral Investig. 15:105–111.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rak JW, St Croix BD and Kerbel RS:
Consequences of angiogenesis for tumor progression, metastasis and
cancer therapy. Anticancer Drugs. 6:3–18. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Asahara T, Masuda H, Takahashi T, et al:
Bone marrow origin of endothelial progenitor cells responsible for
postnatal vasculogenesis in physiological and pathological
neovascularization. Circ Res. 85:221–228. 1999. View Article : Google Scholar
|
|
13
|
Asahara T, Murohara T, Sullivan A, et al:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ziebart T, Yoon CH, Trepels T, et al:
Sustained persistence of transplanted proangiogenic cells
contributes to neovascularization and cardiac function after
ischemia. Circ Res. 103:1327–1334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Urbich C, Heeschen C, Aicher A, et al:
Cathepsin L is required for endothelial progenitor cell-induced
neovascularization. Nat Med. 11:206–213. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Efstathiou E, Bozas G, Kostakopoulos A, et
al: Combination of docetaxel, estramustine phosphate, and
zoledronic acid in androgen-independent metastatic prostate cancer:
efficacy, safety, and clinical benefit assessment. Urology.
65:126–130. 2005. View Article : Google Scholar
|
|
17
|
Bertelli G, Heouaine A, Arena G, et al:
Weekly docetaxel and zoledronic acid every 4 weeks in
hormone-refractory prostate cancer patients. Cancer Chemother
Pharmacol. 57:46–51. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vogt U, Bielawski KP, Bosse U and
Schlotter CM: Breast tumour growth inhibition in vitro
through the combination of
cyclophosphamide/metotrexate/5-fluorouracil,
epirubicin/cyclophosphamide, epirubicin/paclitaxel, and
epirubicin/docetaxel with the bisphosphonates ibandronate and
zoledronic acid. Oncol Rep. 12:1109–1114. 2004.
|
|
19
|
Wood J, Bonjean K, Ruetz S, et al: Novel
antiangiogenic effects of the bisphosphonate compound zoledronic
acid. J Pharmacol Exp Ther. 302:1055–1061. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Janic B and Arbab AS: The role and
therapeutic potential of endothelial progenitor cells in tumor
neovascularization. Sci World J. 10:1088–1099. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
De Falco E, Porcelli D, Torella AR, et al:
SDF-1 involvement in endothelial phenotype and ischemia-induced
recruitment of bone marrow progenitor cells. Blood. 104:3472–3482.
2004.PubMed/NCBI
|
|
22
|
Brunner M, Thurnher D, Heiduschka G, Grasl
M, Brostjan C and Erovic BM: Elevated levels of circulating
endothelial progenitor cells in head and neck cancer patients. J
Surg Oncol. 98:545–550. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Aicher A, Zeiher AM and Dimmeler S:
Mobilizing endothelial progenitor cells. Hypertension. 45:321–325.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hristov M, Erl W and Weber PC: Endothelial
progenitor cells: mobilization, differentiation, and homing.
Arterioscler Thromb Vasc Biol. 23:1185–1189. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Frith JC, Monkkonen J, Blackburn GM,
Russell RG and Rogers MJ: Clodronate and liposome-encapsulated
clodronate are metabolized to a toxic ATP analog, adenosine
5′-(beta, gamma-dichloromethylene) triphosphate, by mammalian cells
in vitro. J Bone Miner Res. 12:1358–1367. 1997.PubMed/NCBI
|
|
26
|
Nogawa M, Yuasa T, Kimura S, et al:
Zoledronic acid mediates Ras-independent growth inhibition of
prostate cancer cells. Oncol Res. 15:1–9. 2005.PubMed/NCBI
|
|
27
|
Bifulco M: Role of the isoprenoid pathway
in ras transforming activity, cytoskeleton organization, cell
proliferation and apoptosis. Life Sci. 77:1740–1749. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Crick DC, Andres DA and Waechter CJ: Novel
salvage pathway utilizing farnesol and geranylgeraniol for protein
isoprenylation. Biochem Biophys Res Commun. 237:483–487. 1997.
View Article : Google Scholar : PubMed/NCBI
|