Introduction
A complex signaling network composed of multiple
signaling molecules and pathways extensively and accurately
regulates the balance between cell proliferation, differentiation
and apoptosis (1), playing a
critical role in the normal development of cells and bions
(2). When this balance is broken,
abnormal cell apoptosis and growth, as well as pathological events
occur. Therefore, investigating the basic mechanisms responsible
for maintaining and destroying the balance between cell
proliferation, differentiation and apoptosis is significant for
understanding the processes of various physiological and
pathological events. Transforming growth factor-β (TGF-β) and its
mediated pathways are important parts of the signaling network that
regulates the physiological functions of bions and cells (3).
TGF-β belongs to a cytokine superfamily that
regulates cell proliferation, differentiation and apoptosis
(4). The activity center of TGF-β
is a 25-kDa dimer structure composed of two identical 12.5 kDa
peptide chains, linked by a disulfide bridge. At least five
subtypes of TGF-β, including TGF-β1, 2, 3, 4 and 5, have been
identified in mammals. The majority of cells express at least one
type of TGF-β (5). Therefore,
TGF-β is a multifunctional factor widely expressed in vivo.
One study demonstrated that TGF-β is widely involved in the
pathophysiological process and exerts significant effects on cell
proliferation and differentiation in vivo, production of
extracellular matrix, wound healing, embryonic development,
angiogenesis, fibrosis, apoptosis and the immune system (6). TGF-β1 has a major presence in the
human body and inhibition of the TGF-β1 signaling pathway may occur
as a result of mutation. Moreover, inactivation of TGF-β1 and its
receptor gene is one of the main causes of cancer development.
The TGF-β1 receptor is a transmembrane glycoprotein
present on the cell surface and is widely distributed on the
surface of normal tissues and tumor cells. TGF-β1 has five
receptors with unique structures (types I, II, III, IV and V),
among which types IV and V are identified only in pituitary cells
and bovine liver. The first three receptors are currently widely
studied. Receptor types I and II (TRβ I and TRβ II) are
transmembrane serine/threonine kinases that initiate intracellular
signaling via phosphorylation of Smads. Receptor types I and II are
composed of an extracellular ligand-binding domain and a domain
with serine/threonine kinase activity involved in intracellular
signal transduction. Receptor type III is not directly involved in
the process of signal transfer. TGF-β1 exerts an effect by binding
to TGF-β1 receptors. Studies have shown that the dimers formed by
receptor types I and II have a leading role in the TGF-β1 signaling
pathway (4). TGF-β1 is a
pleiotropic polypeptide that positively regulates biological
processes and negatively regulates cell proliferation,
differentiation and apoptosis. The biological effects of TGF-β1 are
dependent on cell type and physiological conditions. The absence of
TGF-β1 signaling in T cells causes spontaneous differentiation of T
cells and autoimmune disease (7),
suggesting that TGF-β1 signaling is required for maintaining the
inner balance of T cells. TGF-β1 strongly inhibits the
proliferation of epithelial cells. TGF-β1 has also been reported to
induce apoptosis in certain cells, including liver and liver tumor
cells (8). By contrast, TGF-β1 has
anti-apoptotic effects and is able to elevate the survival rate of
cells in certain cases. Research on the TGF-β family and their
mechanism of action has always gained significant interest due to
the importance, diversity and uncertainty of the role of
TGF-β1.
The mechanism of TGF-β1-mediated signal transduction
via interaction with receptors remains unclear. Smad-mediated
signaling pathways are considered the main signaling pathways by
which TGF-β1 produces its biological effects (9). Previously, a study identified that
extracellular signal-regulated kinases (ERKs), c-Jun N-terminal
kinases (JNKs) and p38 mitogen-activated protein kinases (MAPKs)
are involved in TGF-β-mediated signal transduction events (10) and have significant roles in a
variety of the biological effects mediated by TGF-β.
Apoptosis is a basic physiological mechanism for
maintaining homeostasis in an organism. The body eliminates
damaged, aged and mutant cells by apoptosis to maintain
physiological balance. An abnormal apoptosis level in tissues (too
high or too low) leads to the occurrence of various diseases. The
close correlation between apoptosis and disease has received focus
in the medical field. In addition, apoptosis and its mechanism have
been widely investigated, including the effect of TGF-β1 on the
mechanism of apoptosis. In T cells, TGF-β1 inhibits Fas
ligand-induced apoptosis (11).
Domestic studies concerning the effect of TGF-β1 on the apoptosis
of epithelial cells of the small intestine are few. Thus, the
current study was designed to detect the expression of TGF-β1 in
the epithelial tissue of the rat small intestine in a
cold-restraint stress state, as well as the apoptosis of the
epithelial cells to clarify the effect of TGF-β1 on the apoptosis
of the rat small intestine epithelial cells and provide a
theoretical basis for an improved understanding of the role of
TGF-β1 in apoptosis.
Materials and methods
Animals
A total of 60 male Sprague-Dawley rats weighing
180–220 g were randomized into three groups: the normal control
(NC), cold-restraint stress (CS) and CS drug intervention (DCS)
groups. The rat model of cold-restraint stress was established as
follows: after fixation of limbs on a bracket, the rat was immersed
in cold water (constant temperature, 18°C), with sternum parallel
to the water surface. After 2 h of cold-restraint stress, the rat
was taken from the water and the restraint was removed. The rat was
then placed in a dry and warm cage until the normal state was
recovered. The CS and DCS groups were further randomized into five
subgroups according to the time points (2, 4, 8, 12 and 24 h) after
the establishment of a successful model. From 1 week before the
establishment of the CS model, the DCS group received a single dose
of 20% decorin (Shanghai Deli Biotechnology Co., Ltd., Shanghai,
China) at 0.5 ml/kg into the ranine vein at intervals of 12 h for 7
days, to specifically inhibit the activation of TGF-β1. The CS
models were then established for the CS and DCS groups. The
experiments were approved by Animal Ethics Committee of the First
Affiliated Hospital of Shihezi University (Shihezi, China).
Methods
The apoptotic genome in the ileal mucosa epithelial
cells was extracted using a rapid animal genomic DNA extraction kit
(Beijing extensive biological Tektronix Gene Technology Co., Ltd.,
Beijing, China) and then subjected to DNA agarose gel
electrophoresis. Apoptosis of the cells was detected using terminal
deoxynucleotidyl transferase deoxyuridine triphosphate (dUTP) nick
end labeling (TUNEL; kits were purchased from Roche Company, Basel,
Switzerland). The terminal segment of the ileal mucosal tissue was
embedded with paraffin and then sectioned. TGF-β1 was observed
using the immunohistochemistry streptavidin-peroxidase method (kits
were provided by Wuhan Boster Biological Engineering Co., Ltd.,
Wuhan, China).
Statistical analysis
SPSS 10.0 was used for statistical analysis. The
χ2 test was used for counting data. The t-test and
analysis of variance were used for measurement data. Spearman
correlation analysis was used for the correlation analysis.
P<0.05 and P<0.01 were considered to indicate a statistically
significant and highly statistically significant result,
respectively.
Results
Agarose gel electrophoresis of genomic
DNA
The bands of the DNA genome in the NC group were
detected near the wells. Characteristic ladder bands were detected
in the CS group using agarose gel electrophoresis at 8 and 12 h
after modeling. For the DCS group, the characteristic ladder bands
were not detected using agarose gel electrophoresis at each time
point.
Immunohistochemical staining results
The positive expression rates of TGF-β1 in the
epithelial tissue of the ileal mucous membrane at two of the time
points, 8 and 12 h, for the CS group, were significantly different
to that of the NC group (P<0.05; Table I).
 | Table IExpression of TGF-β1 in the epithelial
tissue of rat ileal mucous tissue in the CS group. |
Table I
Expression of TGF-β1 in the epithelial
tissue of rat ileal mucous tissue in the CS group.
| Group | Positive expression
rate (%) | χ2 | P-value |
|---|
| Control | 16.67 | | |
| 2 h | 22.72 | 0.024 | 0.578 |
| 4 h | 29.17 | 0.444 | 0.370 |
| 8 h | 59.09 | 7.053 | 0.009a |
| 12 h | 54.16 | 5.689 | 0.018a |
| 24 h | 27.27 | 0.262 | 0.434 |
No significant difference was observed in the
positive expression rate of TGF-β1 in the epithelial tissue of the
ileal mucous membrane at each time point in the DCS group compared
with that in the NC group (P>0.05; Table II).
| Table IIExpression of TGF-β1 in the epithelial
tissue of rat ileal mucous tissue in the DCS group. |
Table II
Expression of TGF-β1 in the epithelial
tissue of rat ileal mucous tissue in the DCS group.
| Group | Positive expression
rate (%) | χ2 | P-value |
|---|
| Control | 16.67 | | |
| 2 h | 25.00 | 0.108 | 0.511 |
| 4 h | 23.08 | 0.037 | 0.560 |
| 8 h | 32.14 | 0.840 | 0.275 |
| 12 h | 28.57 | 0.413 | 0.376 |
| 24 h | 22.72 | 0.024 | 0.578 |
Detection of apoptosis
A large number of apoptotic cells were observed
after 8 and 12 h in the CS group. The majority of the apoptotic
cells were located at the top of the epidermis of the mucous
membrane and a few apoptotic cells were located in the crypt. After
counting the number of apoptotic cells in the CN, CS and DCS groups
at each time point, the apoptotic index (AI) was calculated
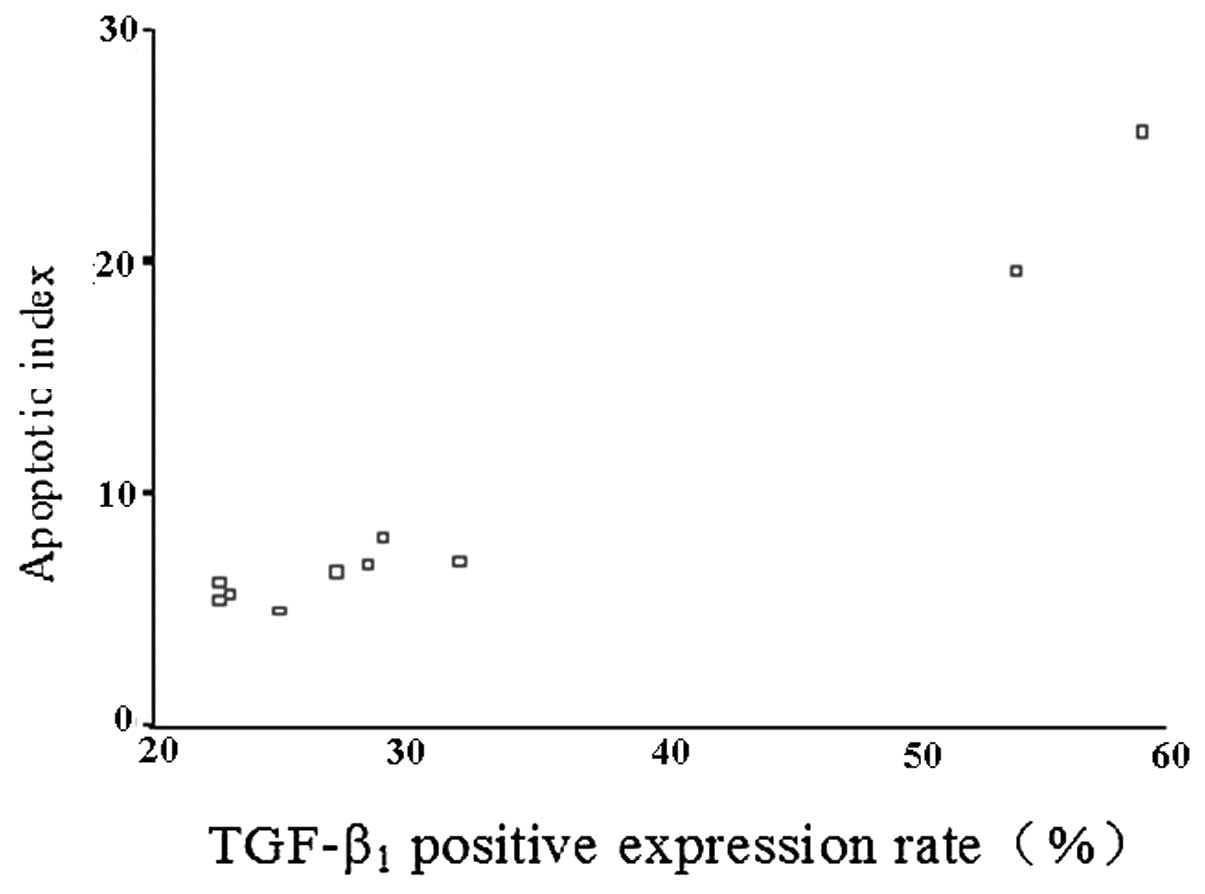
(Table III). Correlation analysis
of the positive expression rate of TGF-β1 and the AI of the
epithelial mucosa of the rat ileal mucous membrane was performed.
The results showed that the AI of the epithelial mucosa of rat
ileal mucous membrane in each time point in DCS group was lower
than the CS group, but still higher than the CN group. The positive
expression rate of TGF-β1 in the ileal mucous membrane of the rat
epithelial mucosa was positively correlated in the CS group and the
DCS group (r=0.980, P<0.05; Fig.
1).
| Table IIIApoptotic index (AI) of the epithelial
mucosa of the rat ileal mucous membrane at each time point. |
Table III
Apoptotic index (AI) of the epithelial
mucosa of the rat ileal mucous membrane at each time point.
| Group | 2 h | 4 h | 8 h | 12 h | 24 h |
|---|
| CN | 4.41±1.87 | 4.42±1.86 | 4.41±1.83 | 4.43±1.88 | 4.41±1.87 |
| CS | 5.47±1.36 | 8.14±3.32a | 25.69±8.09b | 19.65±6.61a | 6.65±2.95 |
| DCS | 4.92±2.36 | 5.68±2.59 | 7.13±2.70a | 6.91±2.13a | 6.25±2.71 |
Discussion
The cold stress reaction is a tension state of the
body from the normal state when stimulated by a cold environment.
The mechanism by which cold stimulation induces the
gastrointestinal stress response is complex and results in
gastrointestinal function and mucosal structural changes affecting
the nerves and body fluids, as well as other factors. Akimoto et
al(12) performed a study on
the correlation between gastrointestinal mucosal injury and
gastrointestinal microcirculation and identified that the
gastrointestinal stress response induced by cold stimulation is
essentially the process of ischemia-reperfusion of gastrointestinal
tissue. Another study demonstrated that the occurrence of apoptosis
in the mucosa of the small intestinal leads to the abnormal
expression of apoptosis-related genes and further increases the
incidence of apoptosis (13).
TGF-β1 promotes apoptosis. In liver cells, TGF-β1
induces apoptosis by activating p38 MAPK (14). TGF-β1 may promote the apoptosis of
the CD34 and CD34-DR cells of patients with chronic myeloid
leukemia; however, it does not increase the expression of the CD95
Fas receptor in leukemia cells. This condition suggests that TGF-β1
promotes the apoptosis of basal cells via a non-Fas-dependent
pathway (15). TGF-β1 promotes the
apoptosis of progenitor cells of normal and malignant B
lymphocytes; however, it does not change the expression of B cell
lymphoma 2 (Bcl-2) (16). The
mechanism by which TGF-β1 promotes the apoptosis of small
intestinal cells remains unclear.
In the current study, the immunohistochemical
detection of TGF-β1, as well as TUNEL and DNA agarose gel protein
electrophoresis for the detection of apoptosis, were performed on
rats at 2, 4, 8, 12 and 24 h after the establishment of the CS
model. The expression levels of TGF-β1 and apoptotic indices of the
ileal mucosa epithelial cells in the rats were dynamically
detected. The pathological changes of the mucosal epithelial tissue
of the rat terminal ileum were observed in the cold-restraint
stress state. After 8–12 h of cold-restraint stress, the villi of
the small intestine had a disordered arrangement; however, the
mucous membrane was complete and not necrotic. After 24 h under
cold-restraint stress, the villi were repaired and arranged
regularly and edema disappeared. The classic theory states that
apoptosis and necrosis are two mutually exclusive methods of cell
death. A previous study (17)
demonstrated that the discrimination between the two is not
absolute; they also often exhibit interrelated phenomena. The
results from the DNA agarose gel protein electrophoresis revealed
the characteristic ‘ladder’ bands of apoptosis after 8 and 12 h of
cold-restraint stress. At each time point, irregular film-like
bands that represent cell necrosis were not present. This result
suggests that the pathological changes of the epithelial cells of
the rat small intestine in the cold-restraint stress state were
mainly apoptosis, not necrosis. In this study, TUNEL technology was
used to detect the occurrence and distribution of epithelial cell
apoptosis. The results revealed that the incidence of apoptosis of
the small intestine epithelial cells in the cold-restraint stress
state at each time point was significantly higher than that of the
NC group. Apoptosis exists at each time point and peaks after 8-12
h of cold-restraint stress. In this study, the apoptotic cells were
mainly located at the top of the villi of the small intestine,
which corresponds with the results from a previous study (18). However, there is also a alternative
viewpoint that apoptosis of small intestine epithelial cells mainly
occurs in the crypt of the intestinal glands and only a small
amount of apoptotic cells are located on the surface of the
intestinal villi, indicating that the apoptosis of cells of the
crypt migrate upward (19).
In the current study, the expression of TGF-β1 was
detected by immunohistochemistry. We identified that TGF-β1 is
expressed in normal rat small intestine epithelial cells. After 2 h
of cold-restraint stress, the positive expression rate of TGF-β1
was slightly higher than that of the NC group, and tended to
increase with time. After 8 h, it reached a peak. In this study,
the experimental results revealed that the AI of the epithelial
cells of the rat small intestine is correlated with the positive
expression rate of TGF-β1. Continual slices of rat small intestine
epithelial tissue were observed. No significant differences were
identified in the positive expression rate of TGF-β1 at 2, 4 and 24
h after cold-restraint stress by immunohistochemical staining or in
the apoptosis rate compared with those of the NC group. Whereas,
after 8 and 12 h of cold-restraint stress, a significant difference
was observed compared with that of the NC group. Following
treatment with the proteoglycan, desmin, which specifically
inhibits the activation of TGF-β1, the rats were subjected to
cold-restraint stimulation. Following TUNEL and immunohistochemical
staining, the results revealed that the morphology of the rat small
intestine epithelium was not significantly different compared with
that of the CS group following the specific inhibition of TGF-β
activity. However, the positive expression rate of TGF-β1 and AI in
the epithelial cells was significantly decreased. The results of
the correlation analysis revealed that the apoptosis rate of the
rat small intestine epithelial cells is positively correlated with
the positive expression rate of TGF-β1. The results revealed that
the AI of the rat small intestine epithelial cells in the
cold-restraint stress state changes with the positive expression
rate of TGF-β1 in the epithelial tissue. TGF-β1 may induce the
apoptosis of rat small intestine epithelial cells. Our data also
revealed that the positive expression rate of TGF-β1 and the AI
were not significantly different after 2, 4 and 24 h of
cold-restraint stress from those of the control group. This result
may be due to the reduced expression of TGF-β1 in the cell stress
reaction at early and post-peak stages in the expression of
TGF-β1.
When TGF-β1 is secreted into the blood, the
epithelial cells of the small intestine cause apoptosis through the
paracrine mechanism, when the intracellular TGF-β1 levels increase.
The autocrine mechanism in cells is also able to induce apoptosis.
In this study, after injection with TGF-β1 inhibitor decorin, the
AI of the rat small intestine epithelial cells decreased along with
the positive expression level of TGF-β1. However, this was still
higher than that of the control rats in a normal state. This result
indicates that other mechanisms are able to induce and promote the
apoptosis of epithelial cells in the rat small intestine. However,
the specific mechanism of the TGF-β1-induced apoptosis of the rat
small intestine epithelial cells remains unclear. Studies have
shown that TGF-β1-induced apoptosis in the small intestine is
correlated with a high expression level of c-MYC and low expression
level of Bcl-2 (20). TGF-β1
causes G1 arrest through a variety of mechanisms (21), including preventing pRB
phosphorylation, inhibiting the protein expression of the G1 phase
of the cell cycle, inhibiting cyclin-dependent kinase (CDK)
activity and increasing CDK levels (22).
In summary, TGF-β1 inhibits cells from passing the
G1/S checkpoint through a variety of mechanisms, which arrests
cells in the G1 phase. However, the specific mechanism of the
TGF-β1-induced apoptosis of the epithelial cells of the rat small
intestine has not been investigated in this study due to limited
time and conditions, and requires further investigation.
The experimental results revealed that the
expression of TGF-β1 was increased in the rat small intestine
epithelial cells under cold-restraint stress and apoptosis
increased. TGF-β1 may be one of the cytokines that induces the
apoptosis of the epithelial cells of the mucous membrane in the
small intestine.
References
|
1.
|
Rahimi N, Tremblay E, McAdam L, Roberts A
and Elliott B: Autocrine secretion of TGF-beta 1 and TGF-beta 2 by
pre-adipocytes and adipocytes: a potent negative regulator of
adipocyte differentiation and proliferation of mammary carcinoma
cells. In Vitro Cell Dev Biol Anim. 34:412–420. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Liao JH, Chen JS, Chai MG, Zhao S and Song
JG: The involvement of p38 MAPK in transforming growth factor
beta1-induced apoptosis in murine hepatocytes. Cell Res. 11:89–94.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Hahm KB, Im YH, Lee C, et al: Loss of
TGF-beta signaling contributes to autoimmune pancreatitis. J Clin
Invest. 105:1057–1065. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Gorelik L and Flavell RA: Abrogation of
TGF beta signaling in T cells leads to spontaneous T cell
differentiation and auto-immune disease. Immunity. 121:171–181.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Yue J and Mulder KM: Transforming growth
factor-beta signal transduction in epithelial cells. Pharmacol
Ther. 91:1–34. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Dkhissi F, Rayna S, Jullien P and Lawrence
DA: Growth stimulation of murine fibroblasts by TGF-beta1 depends
on the expression of a functional p53 protein. Oncogene.
18:703–711. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Petruschke T, Rohrig K and Hauner H:
Transforming growth factor beta (TGF-beta) inhibits the
differentiation of human adipocyte precursor 532–536 cells in
primary culture. Int J Obes Relat Metab Disord. 18:23–35.
1994.PubMed/NCBI
|
|
8.
|
Rahimi N, Tremblay E, McAdam L, Roberts A
and Elliott B: Autocrine secretion of TGF-beta 1 and TGF-beta 2 by
pre-adipocytes and adipocytes: a potent negative regulator of
adipocyte differentiation and proliferation of mammary carcinoma
cells. In Vitro Cell Dev Biol Anim. 34:412–420. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Derynck R, Akhurst RJ and Balmain A:
TGF-beta signaling in tumor suppression and cancer progression. Nat
Genet. 29:117–129. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Ge B, Gram H, Di Padova F, et al:
MAPKK-independent activation of p38 alpha mediated by
TAB1-dependent autophosphorylation of p38alpha. Science.
295:1291–1294. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Hannun YA and Luberto C: Ceramide in the
eukaryotic stress response. Trends Cell Biol. 10:73–80. 2000.
View Article : Google Scholar
|
|
12.
|
Akimoto M, Hashimoto H, Shigemoto M and
Yokoyama I: Relationship between recurrence of gastric ulcer and
the microcirculation. J Cardiovasc Pharmacol. 31(Suppl 1):
S507–S508. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Hartsough MT and Mulder KM: Transforming
growth factor beta activation of p44mapk in proliferating cultures
of epithelial cells. Cell Res. 270:7117–7124. 1995.PubMed/NCBI
|
|
14.
|
Yamaguchi K, Shirakabe K, Shibuya H, et
al: Identification of a member of the MAPKKK family as a potential
mediator of TGF-beta signal transduction. Science. 270:2008–2011.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Shibuya H, Yamaguchi K, Shirakabe K, et
al: TAB1: an activator of the TAK 1 MAPKKK in TGF-beta signal
transduction. Science. 272:1179–1182. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Gram H, Di Padova F, Huang B, et al:
MAPKK-independent activation of p38 alpha mediated by
TAB1-dependent autophosphorylation of p38 alpha. Science.
295:1291–1294. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Larisch S, Yi Y, Lotan R, et al: A novel
mitochondrial septin-like protein, ARTS, mediates apoptosis
dependent on its P-loop motif. Nat Cell Biol. 2:915–921. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Okazaki T, Kondo T, Kitano T and Tashima
M: Diversity and complexity of ceramide signaling in apoptosis.
Cell Signal. 10:685–692. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Liu G, Kleine L and Hebert RL: Advances in
the signal trans duction of ceramide and related sphingolipids.
Crit Rev Clin Lab Sci. 36:511–573. 1999. View Article : Google Scholar
|
|
20.
|
Arsura M, FitzGerald MJ, Fausto N and
Sonenshein GE: Nuclear factor-kappaB/Rel blocks transforming growth
factor beta 1-induced of murine hepatocyte cell lines. Cell Growth
Differ. 8:1049–1059. 1997.PubMed/NCBI
|
|
21.
|
Li Z and Li J: Local expressions of
TGF-beta1, TGF-beta1RI, CTGF and Smad-7 in Helicobacter
pylori-associated gastritis. Scand J Gastroenterol.
41:1007–1012. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Azuma H, Ehata S, Miyazaki H, et al:
Effect of Smad7 expression on metastasis of mouse mammary carcinoma
JygMC(A) cells. J Natl Cancer Inst. 97:1734–1746. 2005. View Article : Google Scholar : PubMed/NCBI
|