Introduction
Glioma is the most common primary brain tumor and
the prognosis of patients with advanced glioma is very poor
(1). Although a number of studies
have focused on exploring the pathogenesis of glioma (2–4), the
molecular mechanisms underlying its development and malignant
progression remain largely unknown. Therefore, it is necessary to
identify novel biomarkers in glioma, as well as reveal the
molecular mechanism that underlies glioma progression, which may be
beneficial for the early diagnosis and effective treatment of the
disease.
microRNAs (miRNAs or miRs), a class of endogenous
small RNAs, are primary regulators of gene expression, which
function by directly interacting with the 3′untranslational region
(3′UTR) of their target mRNAs, leading to translation, repression
or mRNA degradation (5–7). Through mediating the expression of
their target genes, miRNAs have been demonstrated to participate in
various cellular biological processes, including cell
proliferation, differentiation, apoptosis, survival, and invasion
(8–10). Furthermore, a large number of miRNAs
are dysregulated in certain types of human cancer, including glioma
and glioblastoma, and many have been reported to serve oncogenic or
tumor suppressive roles in the development and malignant
progression of glioma, including miR-365, miR-599 and miR-423
(11–13).
miR-200a is a member of the miR-200 family, that has
been demonstrated to serve a tumor suppressive or oncogenic role in
certain common types of human cancer, including gastric cancer,
esophageal cancer, endometrioid endometrial carcinoma,
hepatocellular carcinoma, breast cancer and cholangiocarcinoma
(14–19). For instance, miR-200a enhanced the
tumor necrosis factor-related apoptosis-inducing ligand-induced
apoptosis in gastric cancer cells (17). However, miR-200a promotes esophageal
cancer cell proliferation (18).
Additionally, certain target genes of miR-200a have been identified
in different types of human cancer, including A20 (17), collapsin response mediator protein 1
(18), zinc finger E-box binding
homeobox 2 (ZEB2) (15), phosphatase
and tens in homolog (14) and yes
associated protein 1 (YAP1) (16).
For instance, miR-200a suppresses the metastasis of a distinct
subpopulation of cells (obtained from flow cytometry) in
hepatocellular carcinoma by decreasing ZEB2 (15), but promotes anoikis resistance and
metastasis in human breast cancer by targeting YAP1 (16). Su et al (20) previously revealed that miR-200a
impairs glioma cell growth, survival and invasion by targeting
single-minded family basic helix-loop-helix transcription factor 2
(SIM2). Although a single miRNA generally has many target genes,
other targets of miR-200a may also serve primary roles in
glioma.
Forkhead box A1 (FOXA1), a member of the fork head
transcription factor family, has been demonstrated to participate
in the regulation of cell proliferation, differentiation and
motility (21,22). The aberrant expression of FOXA1 has
been observed in many types of human cancer and its oncogenic role
has also been elucidated (23,24). For
instance, the knockdown of FOXA1 inhibits the proliferation and
migration of pancreatic cancer cells (23). Furthermore, several miRs have been
revealed to serve a tumor suppressive role by targeting FOXA1
(25). For instance, miR-212
suppresses the proliferation and migration of osteosarcoma cells
via the inhibition of FOXA1 expression (25). However, to the best of our knowledge,
no previous studies have assessed whether FOXA1 is associated with
miR-200a-mediated glioma cells.
Therefore, the present study aimed to assess the
clinical significance of miR-200a expression in glioma and to
elucidate the regulatory mechanism of miR-200a underlying the
development and progression of glioma.
Materials and methods
Clinical tissue samples
The present study was approved by the Ethics
Committee of the Brain Hospital of Hunan Province (Changsha,
China). Glioma tissues (n=66) were collected from patients with
glioma (39 male, 27 female; age, 24–64 years; median age, 49.3
years) by surgical resection at the Brain Hospital of Hunan
Province between March 2013 and October 2016. Patients were
excluded from the current study if they received chemotherapy or
radiotherapy prior to surgical resection. A total of 8 patients
with glioma were diagnosed at World Health Organization stage II,
24 at stage III and 34 at stage IV. Furthermore, 15 normal brain
tissues were also collected from healthy patients (9 male, 6
female; age, 33–57 years; median age, 46.7 years) via surgical
resection at the same hospital between June 2013 and September
2016. Written informed consent was obtained from all patients. The
clinicopathologic characteristics of the patients are summarized in
Table I. Fresh tissues were stored
at −80°C prior to further use.
 | Table I.Association between miR-200a
expression and clinicopathological characteristics in patients with
glioma. |
Table I.
Association between miR-200a
expression and clinicopathological characteristics in patients with
glioma.
| Variables | Cases (n=66) | Low miR-200a
expression (n=36) | High miR-200a
expression (n=30) | P-value |
|---|
| Age (years) |
|
|
| 0.623 |
|
<55 | 29 | 17 | 12 |
|
|
≥55 | 37 | 19 | 18 |
|
| Sex |
|
|
| 0.618 |
|
Male | 39 | 20 | 19 |
|
|
Female | 27 | 16 | 11 |
|
| WHO stage |
|
|
| 0.003 |
|
II–III | 32 | 11 | 21 |
|
| IV | 34 | 25 | 9 |
|
Cell culture
Several common human glioma cell lines (U251-MG,
T98G, U-87MG Uppsala and U-373MG Uppsala) were purchased from the
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Normal human astrocyte (NHA) cells were
purchased from the American Type Culture Collection (Manassas, VA,
USA). These cells were cultured in Dulbecco's modified Eagle's
medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.)
at 37°C, 5% CO2 and 95% O2.
Cell transfection
Lipofectamine™ 2000 (Thermo Fisher
Scientific, Inc.) was used to perform cell transfection according
to the manufacturer's protocol. T98G and U251 cells were
transfected with 100 nM miR-200a mimic (cat. no. HmiR0002-MR04;
Guangzhou Fulengen Co., Ltd., Guangzhou, China), 100 nM miR-200a
inhibitor (cat. no. HmiR-AN0298-SN-10; Guangzhou Fulengen Co.,
Ltd.) and 100 nM miR negative control (miR-NC; cat. no.
CmiR0001-MR04; Guangzhou Fulengen Co., Ltd.). Cells were also
co-transfected with 100 nM miR-200a mimic and 100 nM blank pcDNA3.1
vector (cat. no. ZL00008; Yearthbio, Changsha, China) or with 100
nM miR-200a mimic and 100 nM FOXA1 plasmid (cat. no. ZL00371;
Yearthbio). Non-transfected cells were used as control. Further
experimentation was performed 48 h following transfection.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues or U251, T98G,
U-87MG Uppsala, U-373MG Uppsala and NHA cells using TRIzol reagent
(Thermo Fisher Scientific, Inc.) in accordance with the
manufacturer's protocol. Total RNA (1 µg) was then reverse
transcribed into cDNA using the PrimeScript Reverse Transcription
kit (Takara Biotechnology Co., Ltd., Dalian, China), according to
the manufacturer's protocol. qPCR reactions were performed using
the SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.) with a 7300
plus ABI system (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. For the assessment of mRNA expression,
GAPDH was used as internal reference. For the expression of
miR-200a, U6 was used as internal reference. The thermocycling
conditions were as follows: 95°C for 5 min, followed by 35 cycles
of 95°C for 15 sec and 60°C for 30 sec. Relative expression was
analyzed using the 2−ΔΔCq method (26). The primers for FOXA1 (cat. no.
HQP055418), GAPDH (cat. no. HQP006940), miR-200a (cat. no.
HmiRQP0298) and U6 (cat. no. HmiRQP9001) were purchased from
Guangzhou Fulengen Co., Ltd.
Dual luciferase reporter gene
assay
TargetScan 7.1 software (www.targetscan.org) was used to assess the putative
target genes of miR-200a. The mutant type (MT) or wild type (WT)
3′UTR of FOXA1 was amplified and cloned into the psiCHECK-2 vector
(Promega Corporation, Madison, WI, USA) to construct luciferase
reporter plasmids, which were obtained from Yearthbio. U251 and
T98G cells were co-transfected with the WT FOXA1 plasmid or the MT
FOXA1plasmid and miR-200a mimic or miR-NC, respectively, using
Lipofectamine™ 2000 (Thermo Fisher Scientific, Inc.).
Following transfection for 48 h, a dual-luciferase reporter assay
system (Promega Corporation) was used to examine the luciferase
activity, according to the manufacturer's protocol. Renilla
luciferase activity was normalized to Firefly luciferase
activity.
Western blot analysis
T98G and U251 Cells were lysed using radio
immunoprecipitation assay lysis buffer (Thermo Fisher Scientific,
Inc.). Protein concentration was determined using a BCA Protein
Assay kit (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Protein (50 µg per lane) was separated
using 12% SDS-PAGE and transferred onto polyvinylidene fluoride
membranes (Thermo Fisher Scientific, Inc.). Membranes were then
blocked with 5% evaporated milk at room temperature for 1 h and
subsequently incubated with rabbit anti-human FOXA1 (1:200; cat.
no. ab170933; Abcam, Cambridge, UK)or rabbit anti-human GAPDH
(1:200; cat. no. ab9485; Abcam) antibodies overnight at 4°C.
Membranes were further incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies
(1:5,000; cat. no. ab6721; Abcam) for 40 min at room temperature.
Chemiluminescence was detected using the SuperSignal West Femto
Maximum Sensitivity substrate (Thermo Fisher Scientific, Inc.). The
quantity of protein was analyzed using Image J 1.45 software
(National Institutes of Health, Bethesda, MD, USA).
MTT cell proliferation assay
U251 and T98G cells (5×103 cells/well)
were plated into 96-well plates and cultured for different
durations (0, 24, 48 or 72 h) at 37°C in a humidified atmosphere
containing 5% CO2. A total of 10 µl MTT solution (5
mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added at
different time points (0, 24, 48 or 72 h). Cells were then
incubated at 37°C for 4 h. The supernatant was then removed and 100
µl dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added. The
absorbance value (optical density) was measured at 570 nm using a
microplate reader.
EdU incorporation cell viability
assay
U251 and T98G cells (5×103 cells/well)
were seeded into 96-well plates and incubated at 37°C for 24 h. A
total of 50 µM EdU (Guangzhou RiboBio Co., Ltd., Guangzhou, China)
was then added. Following this, cells were incubated at 37°C for 2
h. Cells were fixed with 4% paraformaldehyde for 30 min at room
temperature and then treated with 0.5% Triton-X-100 (Sigma-Aldrich;
Merck KGaA) for 30 min at room temperature. Subsequently, 100 µl
Apollo® Reaction Cocktail (Guangzhou RiboBio Co., Ltd.)
was added and cells were incubated at room temperature for 30 min.
Following staining with DAPI (5 µg/ml; Sigma-Aldrich; Merck KGaA)
at room temperature for 30 min, cells were visualized under a
fluorescent microscope (IX71; Olympus Corporation, Tokyo,
Japan).
Transwell invasion assay
Matrigel pre-coated Transwell chambers (BD
Biosciences, Franklin Lakes, NJ, USA) were utilized to examine cell
invasion. T98G and U251 cell suspensions (3×105
cells/ml) were prepared using DMEM. Subsequently, 300 µl cell
suspension was added into the upper chamber and 300 µl DMEM
supplemented with 10% FBS was added into the lower chamber. Cells
were incubated at 37°C for 24 h and the cells that did not invade
through the membrane were carefully removed. The transmigrated
cells were then fixed using 4% polyformaldehyde at room temperature
for 30 min, stained with crystal violet (Sigma-Aldrich; Merck KGaA)
at room temperature for 10 min and counted in five randomly
selected microscopic fields using a light microscope
(magnification, ×400). All experiments were performed in
triplicate.
Statistical analysis
Data are presented as the mean ± standard deviation.
GraphPad Prism version 5.01 software (GraphPad Software, Inc., La
Jolla, CA, USA) was used to perform statistical analysis.
Differences between two groups were analyzed using a Student's
t-test and differences among more than two groups were analyzed
using one-way analysis of variance followed by a Tukey's post-hoc
test. Glioma patients were divided into high and low miR-200a
expression groups, based on its mean expression value, and a
χ2 test was used to analyze the results presented in
Table I. A Pearson correlation test
was used to determine the relationship between miR-200a and
FOXA1expression. P<0.05 was considered to indicate a
statistically significant result.
Results
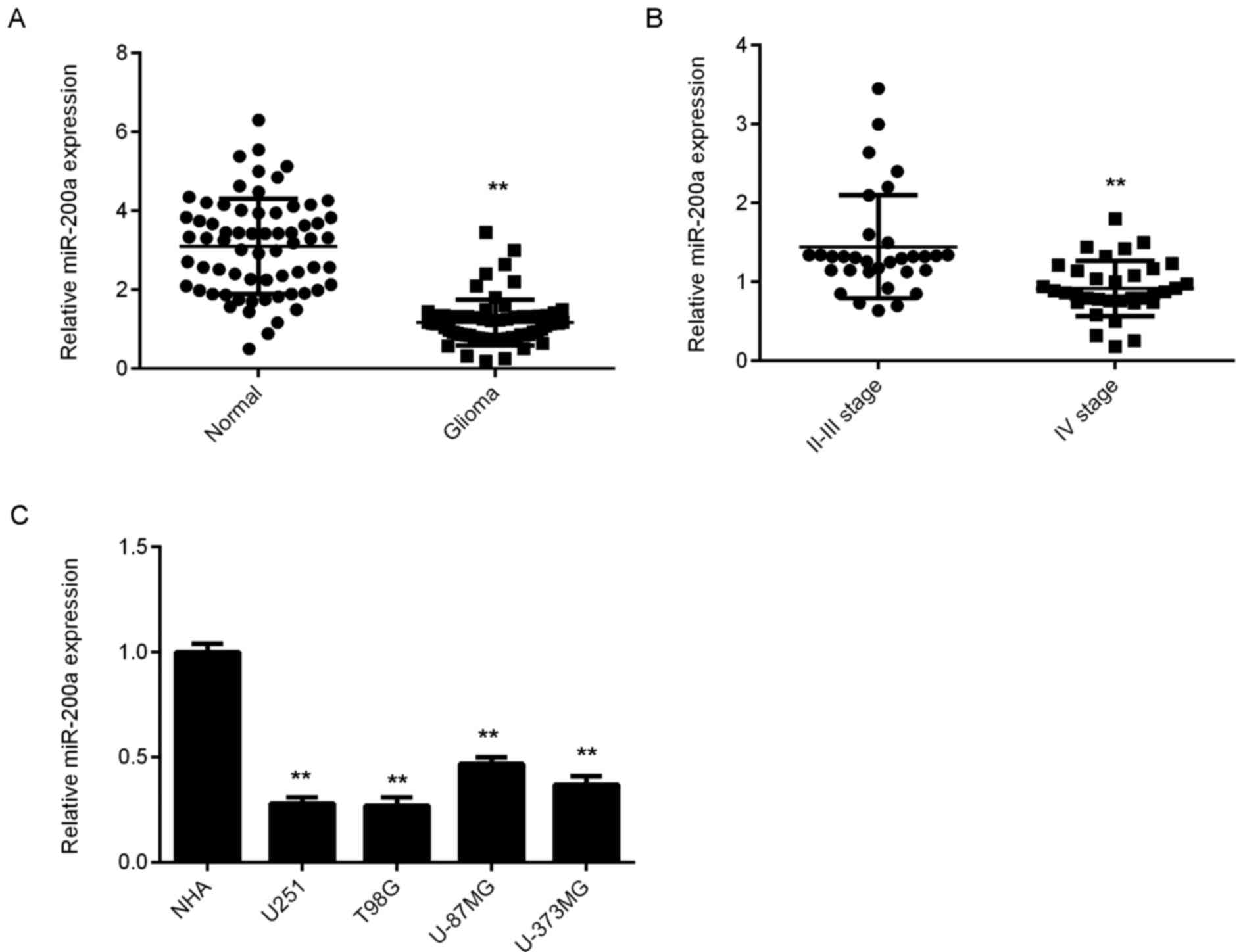
miR-200a is downregulated in
glioma
RT-qPCR was used to assess the expression of
miR-200a in glioma and normal brain tissues. As presented in
Fig. 1A, the levels of miR-200a were
significantly lower in glioma tissues compared with normal brain
tissues. Furthermore, grade IV glioma tissues demonstrated
significantly lower miR-200a levels when compared with grade II–III
glioma tissues (Fig. 1B).
Subsequently, glioma patients were divided into high and low
miR-200a expression groups, based on its mean expression value. As
presented in Table I, the low
expression of miR-200a was significantly associated with advanced
clinical stage in glioma. In addition, miR-200a levels were
significantly reduced in several common glioma cell lines compared
with those in NHA cells (Fig. 1C).
As T98G and U251 exhibited the lowest expression of miR-200a, they
were used for the following experiments.
miR-200a inhibits glioma cell
proliferation, survival, and invasion
T98G and U251 cells were transfected with miR-200a
mimics to upregulate the expression of miR200a. The results
indicated that miR-200a levels were significantly increased in the
miR-200a group compared with the miR-NC group. However,
transfection with miR-NC did not significantly affect the
expression of miR-200a (Fig. 2A).
The effects of miR-200a upregulation on glioma cells proliferation,
survival and invasion were then assessed. As presented in Fig. 2B-D, the overexpression of miR-200a
markedly reduced T98G and U251 cell proliferation (at 48 and 72 h),
viability and invasion. These findings suggest that miR-200a
demonstrated suppressive effects on the proliferation, survival and
invasion of glioma cells.
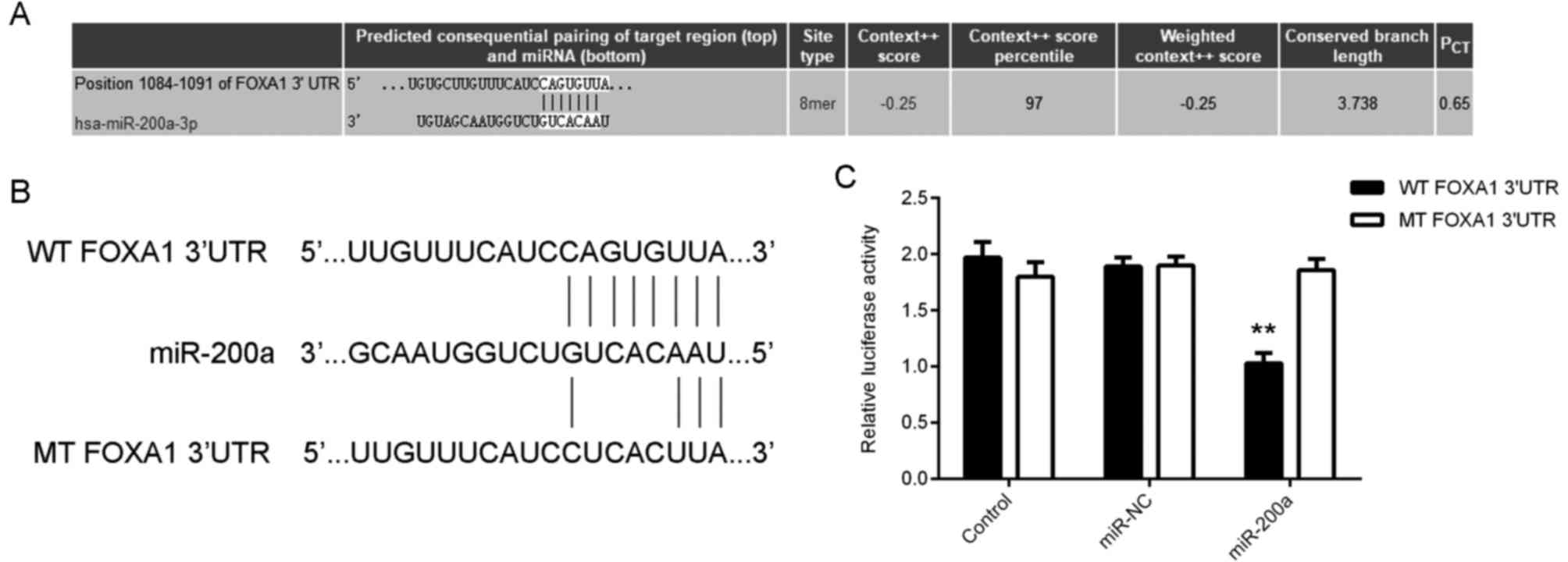
FOXA1 is a direct target gene of
miR-200a in glioma cells
The potential downstream target genes of miR-200a in
glioma were then assessed. TargetScan software data indicated that
the 3′UTR of FOXA1 mRNA contained the binding sequences of miR-200a
(Fig. 3A). To confirm this
prediction, the present study constructed the WT and MT FOXA1 3′UTR
luciferase reporter plasmids (Fig.
3B). Dual luciferase reporter gene assay data indicated that
the luciferase activity was significantly reduced in glioma cells
co-transfected with miR-200a mimics and WT reporter plasmids, when
compared with cells transfected with WT reporter plasmids. However,
this result was reversed following transfection with the MT
reporter plasmid (Fig. 3C and D).
These data indicate that FOXA1 is a target gene of miR-200a in
glioma cells.
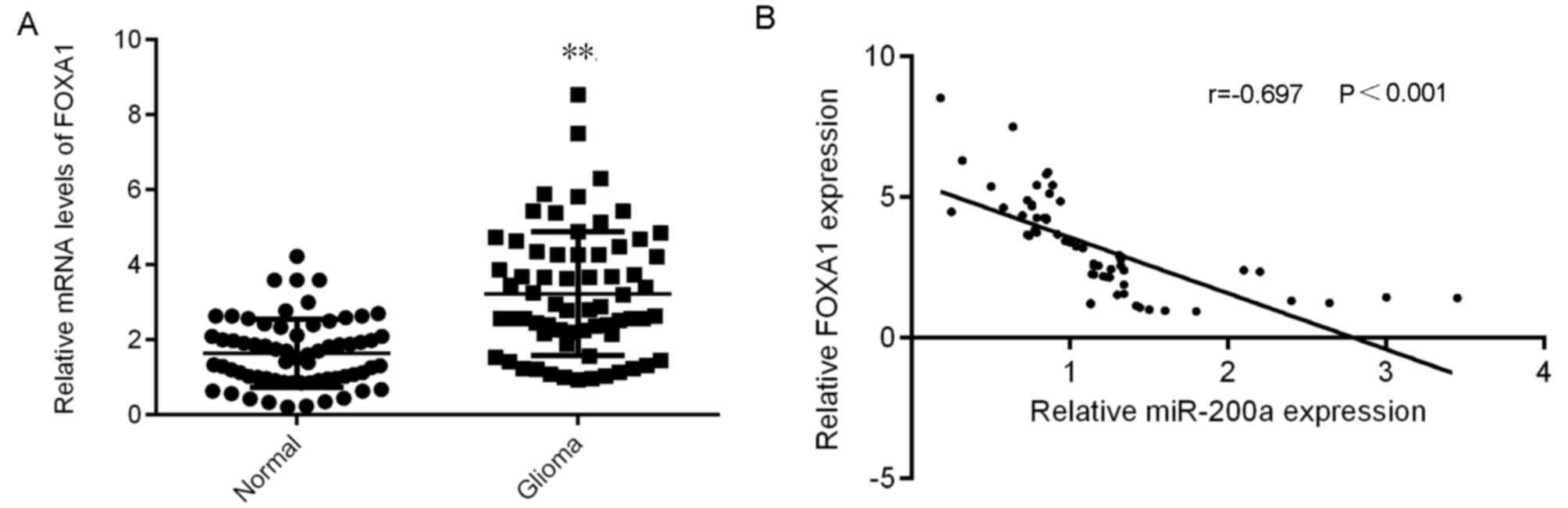
FOXA1 is upregulated in glioma and
inversely correlates to miR-200a expression
The expression of FOXA1 in glioma was further
assessed. The results demonstrated that FOXA1 was significantly
upregulated in glioma tissues compared with normal brain tissues
(Fig. 4A). A negative correlation
between FOXA1 and miR-200a expression in glioma tissues was also
identified (Fig. 4B). Therefore,
FOXA1was demonstrated to be upregulated in glioma, which may be due
to the downregulation of miR-200a.
miR-200a negatively regulates FOXA1
expression in glioma cells
The effect of miR-200a on the expression of FOXA1in
glioma cells was assessed. The results demonstrated that FOXA1 mRNA
and protein expression was significantly reduced following miR-200a
overexpression (Fig. 5A and B).
miRNAs not only affect mRNA expression, but also function to
inhibit protein translation (5–7). Due to
this double effect, transfection with miR-200a mimics demonstrated
a greater reduction in FOXA1 protein expression when compared with
mRNA expression. T98G and U251 cells were transfected with
amiR-200a inhibitor to knockdown its expression. RT-qPCR data
revealed that miR-200a expression was significantly reduced in the
miR-200a inhibitor group compared with the control group.
Furthermore, no marked difference in miR-200a expression between
the NC inhibitor and control group was identified (Fig. 5C). As presented in Fig. 5D and E, the expression of FOXA1 mRNA
and protein was significantly increased in the miR-200a inhibitor
group compared with the NC inhibitor group. These findings suggest
that the expression of FOXA1 was negatively regulated by miR-200a
in T98G and U251 cells.
FOXA1 reduces the inhibitory effects
of miR-200a on the malignant phenotypes of glioma cells
Rescue experiments were performed to assess whether
FOXA1 acted as a downstream effecter of miR-200a in glioma cells.
miR-200a-overexpressing T98G and U251 cells were transfected with
FOXA1 expression plasmid or blank vectors (acting as the negative
control group). Following transfection, FOXA1 protein levels were
markedly higher in the miR-200a+FOXA1 group compared with those in
the miR-200a+blank group. It was also demonstrated that the
proliferation (at 48 and 72 h), viability and invasion of glioma
cells were significantly increased in the miR-200a+FOXA1 group
compared with those in the miR-200a+blank group (Fig. 6B-D). These results indicated that
FOXA1 rescued the inhibitory effects of miR-200a by affecting the
proliferation, viability and invasion of glioma cells.
Discussion
The molecular mechanism of miR-200a underlying
glioma progression remains largely unknown. However, the present
study demonstrated that the level of miR-200a was significantly
reduced in glioma tissues and cell lines. Low miR-200a expression
was also negatively correlated with advanced stages of glioma.
Furthermore, miR-200a overexpression inhibited glioma cell
proliferation, survival and invasion. Results also identified that
FOXA1 was a target gene of miR-200a in glioma cells and the
increased expression of FOXA1 was negatively correlated with the
decreased expression of miR-200a in glioma tissues. FOXA1
expression was also negatively regulated by miR-200a in glioma
cells and the overexpression of FOXA1 rescued the inhibitory
effects of miR-200a on the malignant phenotypes of glioma
cells.
Previous studies have demonstrated that miR-200a
serves key roles in multiple types of human cancer; however,
studies that focus on miR-200a function in glioma are limited
(20,27,28).
Berthois et al (28) revealed
that miR-200a was downregulated in glioblastoma multiforme (grade
IV glioma) when compared with grade II–III glioma. These results
were consistent with the data of the present study. Furthermore,
Berthois et al (28)
demonstrated that miR-200a levels were significantly higher in
Timozolamine (TMZ)-responsive glial tumor cells compared to
TMZ-unresponsive glial tumor cells in primary culture and that the
overexpression of miR-200a in glioblastoma (GBM) cells promoted
TMZ-sensitivity. Therefore, miR-200a may serve as a promising
therapeutic target for GBM treatment. The reduced expression of
miR-200a may also be due to the increased expression of the DNA
repair enzyme O(6)-methylguanine methyltransferase (28) and long non-coding RNA activated by
TGF-β, which has been demonstrated to serve an oncogenic role in
glioma (27).
Su et al (20)
demonstrated that SIM2 was a target gene of miR-200a in glioma
cells, as the inhibition of miR-200a resulted in the upregulation
of SIM2 in glioma cells and promoted glioma cell motility. It was
also revealed that miR-200a knockdown promoted glioma growth in a
mouse model (20). As one miRNA has
many target genes (5), other target
genes of miR-200 a may also exist and serve important roles in
glioma. The current study identified FOXA1 as a novel target gene
of miR-200a and revealed that the expression of FOXA1 was
negatively regulated by miR-200a in glioma cells. FOXA1 has
previously been reported to function as an oncogene in certain
types of common human cancer, including prostate (23), endometrial (21) and breast cancer (29). For instance, FOXA1 enhanced the
proliferation and migration of prostate cancer cells by modulating
ELL-associated factor 2 regulation of androgen receptor
transcriptional activity (23).
Furthermore, Wang et al (30)
previously revealed that the expression of FOXA1 in glioma tissues
was significantly higher than those in corresponding non-neoplastic
brain tissues, which was consistent with the results of the current
study. In addition, the present study demonstrated that the
expression of FOXA1 was upregulated in high-grade glioma tissue
compared with low-grade tissue and that the overall survival of
patients with glioma that exhibited high FOXA1 expression was
shorter than those with low FOXA1 (30). This indicates that the upregulation
of FOXA1 may serve a key role in glioma progression (30). The present study identified a
negative correlation between FOXA1 and miR-200a expression in
glioma tissues. These findings indicate that the reduced expression
of miR-200a may contribute to the increased expression of FOXA1 in
glioma tissues. Further investigation revealed that FOXA1
overexpression rescued the inhibitory effects of miR-200a on the
proliferation, viability and invasion of glioma cells. Thus, FOXA1
may act as a key downstream effecter of miR-200a in glioma
cells.
To the best of our knowledge, this is the first
study which demonstrates that miR-200a inhibits glioma cell
survival, proliferation and invasion partly through the inhibition
of FOXA1 expression. These results indicate that miR-200a may be a
potential therapeutic candidate for glioma.
Acknowledgements
Not applicable.
Funding
Not funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
KL and XC designed the present study and wrote the
manuscript. BL, YZ and BZ collected clinical tissues. XC, LY, PY,
WK, HH, and ET performed clinical and cell experimentation.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Brain Hospital of Hunan Province (Changsha,
China). Written informed consent was obtained from all
patients.
Patient consent for publication
Written informed consent was obtained for all
patients.
Conflict of interest
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song H, Zhang Y, Liu N, Wan C, Zhang D,
Zhao S, Kong Y and Yuan L: miR-92b regulates glioma cells
proliferation, migration, invasion, and apoptosis via PTEN/Akt
signaling pathway. J Physiol Biochem. 72:201–211. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gu J, Xu R, Li Y, Zhang J and Wang S:
MicroRNA-218 modulates activities of glioma cells by targeting
HMGB1. Am J Transl Res. 8:3780–3790. 2016.PubMed/NCBI
|
|
4
|
Marumoto T and Saya H: Molecular biology
of glioma. Adv Exp Med Biol. 746:2–11. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ambros V: microRNAs: Tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou Y, Yang C, Wang K, Liu X and Liu Q:
MicroRNA-33b inhibits the proliferation and migration of
osteosarcoma cells via targeting hypoxia-inducible factor-1α. Oncol
Res. 25:397–405. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang M, Zhai X, Ge T, Yang C and Lou G:
MiR-181a-5p promotes proliferation and invasion, and inhibits
apoptosis of cervical cancer cells via regulating inositol
polyphosphate-5-phosphatase A (INPP5A). Oncol Res. 26:703–712.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Li J, Xu C and Zhang X:
MicroRNA-139-5p inhibit cell proliferation and invasion by
targeting RHO-associated Coiled-coil containing protein kinase 2 in
ovarian cancer. Oncol Res. Jun 14–2017.(Epub ahead of print).
|
|
10
|
Wang C, Zhou B, Liu M, Liu Y and Gao R:
miR-126-5p restoration promotes cell apoptosis in cervical cancer
by targeting Bcl2l2. Oncol Res. 25:463–470. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu Y, Zhao H, Rao M and Xu S:
MicroRNA-365 inhibits proliferation, migration and invasion of
glioma by targeting PIK3R3. Oncol Rep. 37:2185–2192. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang T, Ma G, Zhang Y, Huo H and Zhao Y:
miR-599 inhibits proliferation and invasion of glioma by targeting
periostin. Biotechnol Lett. 39:1325–1333. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li S, Zeng A, Hu Q, Yan W, Liu Y and You
Y: miR-423-5p contributes to a malignant phenotype and temozolomide
chemoresistance in glioblastomas. Neuro Oncol. 19:55–65. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoneyama K, Ishibashi O, Kawase R, Kurose
K and Takeshita T: miR-200a, miR-200b and miR-429 are Onco-miRs
that target the PTEN gene in endometrioid endometrial carcinoma.
Anticancer Res. 35:1401–1410. 2015.PubMed/NCBI
|
|
15
|
Yang X, Wang J, Qu S, Zhang H, Ruan B, Gao
Y, Ma B, Wang X, Wu N, Li X, et al: MicroRNA-200a suppresses
metastatic potential of side population cells in human
hepatocellular carcinoma by decreasing ZEB2. Oncotarget.
6:7918–7829. 2015.PubMed/NCBI
|
|
16
|
Yu SJ, Hu JY, Kuang XY, Luo JM, Hou YF, Di
GH, Wu J, Shen ZZ, Song HY and Shao ZM: MicroRNA-200a promotes
anoikis resistance and metastasis by targeting YAP1 in human breast
cancer. Clin Cancer Res. 19:1389–1399. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo T, Zhang Y, Qu X, Che X, Li C, Fan Y,
Wan X, Ma R, Hou K, Zhou H, et al: miR-200a enhances TRAIL-induced
apoptosis in gastric cancer cells by targeting A20. Cell Biol Int.
42:506–514. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zang Y, Tai Y, Wan B and Jia X:
miR-200a-3p promotes the proliferation of human esophageal cancer
cells by post-transcriptionally regulating cytoplasmic collapsin
response mediator protein-1. Int J Mol Med. 38:1558–1564. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng F, Jiang J, Yu Y, Tian R, Guo X, Li
X, Shen M, Xu M, Zhu F, Shi C, et al: Direct targeting of
SUZ12/ROCK2 by miR-200b/c inhibits cholangiocarcinoma
tumourigenesis and metastasis. Br J Cancer. 109:3092–3104. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su Y, He Q, Deng L, Wang J, Liu Q, Wang D,
Huang Q and Li G: MiR-200a impairs glioma cell growth, migration,
and invasion by targeting SIM2-s. Neuroreport. 25:12–17.
2014.PubMed/NCBI
|
|
21
|
Qiu M, Bao W, Wang J, Yang T, He X, Liao Y
and Wan X: FOXA1 promotes tumor cell proliferation through AR
involving the Notch pathway in endometrial cancer. BMC Cancer.
14:782014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Robinson JL and Carroll JS: FoxA1 is a key
mediator of hormonal response in breast and prostate cancer. Front
Endocrinol (Lausanne). 3:682012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo W, Keener AL, Jing Y, Cai L, Ai J,
Zhang J, Fisher AL, Fu G and Wang Z: FOXA1 modulates EAF2
regulation of AR transcriptional activity, cell proliferation, and
migration in prostate cancer cells. Prostate. 75:976–987. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Horimoto Y, Arakawa A, Harada-Shoji N,
Sonoue H, Yoshida Y, Himuro T, Igari F, Tokuda E, Mamat O, Tanabe
M, et al: Low FOXA1 expression predicts good response to
neo-adjuvant chemotherapy resulting in good outcomes for luminal
HER2-negative breast cancer cases. Br J Cancer. 112:345–351. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu J, Chen B, Yue B and Yang J:
MicroRNA-212 suppresses the proliferation and migration of
osteosarcoma cells by targeting forkhead box protein A1. Exp Ther
Med. 12:4135–4141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma CC, Xiong Z, Zhu GN, Wang C, Zong G,
Wang HL, Bian EB and Zhao B: Long non-coding RNA ATB promotes
glioma malignancy by negatively regulating miR-200a. J Exp Clin
Cancer Res. 35:902016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Berthois Y, Delfino C, Metellus P, Fina F,
Nanni-Metellus I, Al Aswy H, Pirisi V, Ouafik L and Boudouresque F:
Differential expression of miR200a-3p and miR21 in grade II-III and
grade IV gliomas: Evidence that miR200a-3p is regulated by
O6-methylguanine methyltransferase and promotes
temozolomide responsiveness. Cancer Biol Ther. 15:938–950. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng L, Qian B, Tian D, Tang T, Wan S,
Wang L, Zhu L and Geng X: FOXA1 positively regulates gene
expression by changing gene methylation status in human breast
cancer MCF-7 cells. Int J Clin Exp Pathol. 8:96–106.
2015.PubMed/NCBI
|
|
30
|
Wang L, Qin H, Li L, Feng F, Ji P, Zhang
J, Li G, Zhao Z and Gao G: Forkhead-box A1 transcription factor is
a novel adverse prognosis marker in human glioma. J Clin Neurosci.
20:654–658. 2013. View Article : Google Scholar : PubMed/NCBI
|