Introduction
Acinetobacter baumannii (ABA) is an important
conditional pathogen in hospitals, and often causes infection in
critically ill patients (1,2). Carbapenems have always been considered
as an important class of antibiotics to treat various infections.
However, the proportion of carbapenem-resistant A. baumannii
(CRABA) has gradually increased in recent years (3). Data from the China Antimicrobial
Surveillance Network (2016) indicate that ABA ranked third among
all the isolates (4). From 2005 to
2016, the resistance rate of ABA to imipenem rose from 31.0-68.6%
and that to meropenem rose from 39.0-71.4% (5).
Integrons are closely associated with the multidrug
resistance of ABA (1). Integron
cassettes are mobile genetic materials in the bacterial genome that
carry components of site-specific recombination systems that can
integrate a number of drug resistance gene cassettes together to
form multiple drug resistance (6).
There are currently 10 types of integrons that have been discovered
and identified in humans, but only 5 integrons are associated with
drug resistance gene cassettes (7).
Class I integrons are the commonest type and their structures
include 5' and 3' conserved ends and an internal variable region
encoding antibiotic resistance genes (2). The gene sequence of class II integrons
is similar to that of class I integrons; the encoded integrin has
46% homology with class I. It is located on transposon Tn7 and its
derivatives and is a defective DNA integrase Intl gene (6), Class III integrons have been identified
on a resistant plasmid of Serratia marcescens that is
resistant to imipenem. Its integrase gene encodes 320 amino acids
and has 51% homology with integrase I (8). Class IV integrons are a special type of
integrons: They were discovered by Cambray et al (9) from Vibrio cholerae. These
integrons are 126 kb in length and contain ≥179 genes. The primary
function of these integrons is to encode adaptability genes. Class
V integrons are located on a complex transposon carried by
Vibrio salmonicidum (10).
At present, studies are mainly focusing on type I,
II and III integrons, as they serve a vital role in drug resistance
transmission. The product encoded by them can resist most clinical
antibiotics (10). Those antibiotics
can be divided into nine categories, of which class A are
carbocylase dilutes: Imipenem (IPM) and meropenem (MEM); class B
are aminoglycosides: Gentamicin (GM) and amikacin, (AN); class C
are quinolones: Ciprofloxacin (CIP), levofloxacin (LVX), and
ciprofloxacin (MXF); class D are cephalosporins: Cefazoxime (CZ),
ceftazidime (CAZ), cefotaxime (CTX) and cefepime (FEP); class E are
broad-spectrum penicillins: Ampicillin/sulbactam (SAM),
piperacillin/tazobactam (TZP), via ampicillin (AMC) and
piperacillin (PIP); class F are β-lactams: Aztreonam (ATM) and
amoxicillin/clavulanic acid (AM); class G are sulfonamides:
Compound neomin (SXT); class H are tetracyclinea: tetracycline
(TE); and class I are amides and alcohols: Chloramphenicol (C). The
resistance rate of integron-positive strains to β-lactams,
aminoglycosides and quinolone antibiotics is increased compared
with integron-negative strains (7).
In China, integrons with resistance gene cassettes are considered
as the main drug resistance factor, as the detection rate of
integrons is between 52.3-69.6% (11).
The present study collected CRABA and non-CRABA
clinical samples from different clinical departments in the Sixth
Affiliated Hospital of Guangzhou Medical University, investigated
the association between integron distribution and drug resistance
in ABA, and analyzed the homology among multilocus sequence types
(MLSTs) in ABA. ABA integrase genes, variable regions and 7
housekeeping genes from 96 strains were amplified and
sequenced.
Materials and methods
Strain conservation
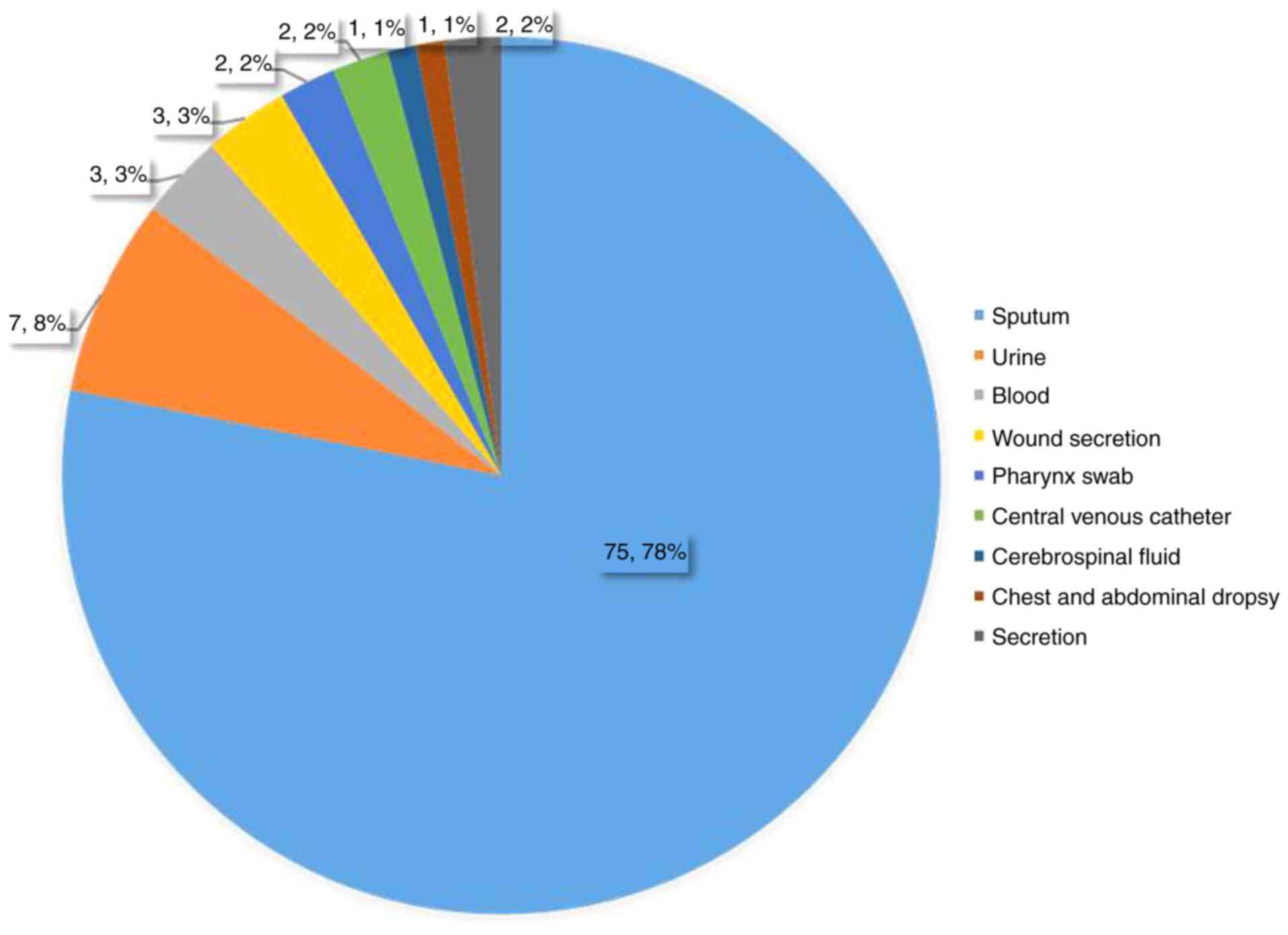
A total of 96 ABA strains were harvested and
separated from clinical samples between August 2014 and October
2018, and duplicates were eliminated. Samples were obtained from
the Intensive Care Unit (ICU), and departments of Neurology,
Infectious Diseases, Respiratory Medicine, Hematology,
neurosurgery, Urology, Neonatal and ENT. All the experiment are
performed under the permission of Medical Ethics committee of
Qingyuan People's Hospital. Quality control strains were
Escherichia coli ATCC25922, Staphylococcus aureus
ATCC25923 and Pseudomonas aeruginosa ATCC27853 (American
Type Culture Collection). Among the 96 ABA strains, 74 were CRABA
strains and 22 were non-CRABA strains (Fig. 1). Written informed consent was
obtained from all patients.
Instruments and reagents
Instruments included a Phoenix M50 automatic
bacterial detector (Becton, Dickinson and Company), a T100 Thermal
Cycler (Bio-Rad Laboratories, Inc.) and a Gel Dox XR+gel imaging
system (Bio-Rad Laboratories, Inc.). Reagents included Colombia
blood Agar plate (Guangzhou Dijing Microbiology Co., Ltd),
bacterial group DNA extraction kit, 2X Taq PCR MasterMix, Gel Red
nucleic acid dye and Marker I, II, III DNA Ladder (all from Tiangen
Biotech (Beijing) Co., Ltd.) and primers from Shanghai Shenggong
Bioengineering Technology Service Co., Ltd.
Total DNA extraction from
bacteria
The strains were stored in sterile milk at 4˚C and
resuscitated prior to use. Following separation and culturing on
blood-containing media, a sufficient number of bacterial colonies
was selected with aseptic cotton and inoculated in a 1.5-ml sterile
saline Eppendorf (EP) tube to prepare a bacterial suspension and
DNA was extracted in accordance with the protocol of the DNA
extraction kit, and transferred to aseptic 1.5 ml EP tube and
stored at -80˚C.
Primer design
Primers were designed as described previously
(12,13) and are listed in Tables I and II.
 | Table IThe primer sequence of three
integrase gene and variable area. |
Table I
The primer sequence of three
integrase gene and variable area.
| Target gene |
Oligonucleotide | Sequence
(5'→3') | Length, base
pairs |
|---|
| Int 1 | int 1-F |
GGTCAAGGATCTGGATTTCG | 493 |
| int 1-R |
ACATGCGTGTAAATCATCGTC |
| Int 2 | int 2-F |
CACGGATATGCGACAAAAAGGT | 789 |
| int 2-R |
GTAGCAAACGAGTGACGAAATG |
| Int 3 | int 3-F |
AGTGGGTGGCGAATGAGTG | 922 |
| int 3-R |
TGTTCTTGTATCGGCAGGTG |
| Int-variable
area | 5'-CS |
GGCATCCAAGCAGCAAG | variable |
| 3'-CS |
AAGCAGACTTGACCTGA |
| Table IIThe primer sequence of multilocus
sequence typing. |
Table II
The primer sequence of multilocus
sequence typing.
| Gene | Oligonucleotide and
sequence | Length/bp |
|---|
| gltA |
gltA(F):5'-AATTTACAGTGGCACATTAGGTCCC-3' | 722 |
|
gltA(R):5'-GCAGAGATACCAGCAGAGATACACG-3' |
| gyrB |
gyrB(F):5'-TGAAGGCGGCTTATCTGAGT-3' | 594 |
|
gyrB(R):5'-GCTGGGTCTTTTTCCTGACA-3' |
| gdhB |
gdhB(F)):5'-GCTACTTTTATGCAACAGAGCC-3' | 774 |
|
gdhB(R):5'-GTTGAGTTGGCGTATGTTGTGC-3' |
| recA |
recA(F):5'-CCTGAATCTTCYGGTAAAAC-3' | 425 |
|
recA(R):5'-GTTTCTGGGCTGCCAAACATTAC-3' |
| cpn60 |
cpn60(F):5'-GGTGCTCAACTTGTTCGTGA-3' | 640 |
|
cpn60(R):5'-CACCGAAACCAGGAGCTTTA-3' |
| Gpi |
gpi(F):5'-GAAATTTCCGGAGCTCACAA-3' | 456 |
|
gpi(R):5'-TCAGGAGCAATACCCCACTC-3' |
| rpoD |
rpoD(F):5'-ACCCGTGAAGGTGAAATCAG-3' | 672 |
|
rpoD(R):5'-TTCAGCTGGAGCTTTAGCAAT-3' |
PCR for integrase genes
The PCR mix for class I, II and III integrons
comprised: 12.5 µl 2X Taq PCR MasterMix, including Taq DNA
polymerase, dNTPs, reaction buffer and MgCl2, 0.5 µl
each of the forward and reverse primers, 0.5 µl DNA template and 25
µl ddH2O. The reaction conditions were: Initial
denaturation at 95˚C for 5 min, followed by 26 cycles of
denaturation at 94˚C for 30 sec, annealing at 62˚C for 30 sec and
extension at 72˚C for 1 min, and then a final extension at 72˚C for
5 min. The PCR products were visualized electrophoretically on a 2%
agarose gel for 45 min at 80 V, and visualized using a gel imaging
system (Bio-Rad Laboratories, Inc.).
PCR for the variable region
The PCR mix for variable regions of class I, II and
III integrons comprised: 12.5 µl 2X Taq PCR MasterMix, 0.5 µl each
of the forward and reverse primer, 1 µl DNA template and 25 µl
ddH2O. The reaction conditions were as follows: initial
denaturation at 94˚C for 5 min, followed by 35 cycles of
denaturation at 94˚C for 30 sec, annealing at 62˚C for 30 sec and
extension at 72˚C for 2 min, and a final extension at 72˚C for 7
min. The PCR products were visualized electrophoretically on a 1.2%
agarose gel for 35 min at 80 V and visualized using a gel imaging
system (Bio-Rad Laboratories, Inc.).
PCR for 7 housekeeping genes in
MLSTs
The PCR mix for 7 housekeeping genes was: 12.5 µl 2X
Taq PCR MasterMix, 0.5 µl each of the forward and reverse primer, 1
µl DNA template and 25 µl ddH2O. The reaction conditions
were: Initial denaturation at 94˚C for 5 min; 30 cycles of
denaturation at 94˚C for 1 min, annealing at 56˚C for 1 min and
extension at 72˚C for 2 min; final extension at 72˚C for 7 min. The
PCR products were visualized electrophoretically on a 1% agarose
gel (35 min, 65 V) and visualized using a gel imaging system
(Bio-Rad Laboratories, Inc.).
Sequence analysis of the variable
regions and 7 housekeeping genes
The PCR products (cut-off value >2) were sent to
Shanghai Bioengineering Company for sequencing, and reads were
corrected and spliced using Chromas software (version 2.6.6;
http://technelysium.com.au/wp/chromas/) and compared
and analyzed using BLAST (BLAST+ version 2.10.0; https://blast.ncbi.nlm.nih.gov/Blast.cgi) and reads
with the highest degree of coincidence were selected.
Genotyping analysis of ABA MLSTs
From the ABA MLST database (https://pubmlst.org/abaumannii/), the sequencing reads
were subjected to BLAST analysis and different allele values were
obtained. All strains were ranked, thus yielding an allelic
spectrum in the order of gltA, gyrB, gdhB,
recA, cpn60, gpi and rpoD.
Whole-genome sequencing
Genomic DNA was extracted from the culture with a
QIAamp DNA kit according to the manufacturer's instructions
(Qiagen). Purified DNA was fragmented by sonication with a Covaris
S2 instrument (Covaris, Inc.). Indexed shotgun sequencing libraries
were prepared using the Kapa Library Preparation kit [Kapa
Biosystems (Pty) Ltd.], following the manufacturer's instructions.
All libraries were sequenced on HiSeq 2000 instruments (Illumina,
Inc.) using paired-end 101 bp reads with an index read of 9 bp.
SPAdes (version 3.13.1; with the following parameters, k
21,33,55,77,99 and 127) was used for assembly of reads (14,15). The
phylogenetic tree were constructed using core genome single
nucleotide polymorphism (CG-SNP) analysis with Parsnp (version 1.2;
with the following parameters, -x -c) (16), followed by displaying with iTOL
(http://itol.embl.de) (17). The scale represents relative genetic
distance.
Pulsed-field gel electrophoresis
(PFGE)
All the bacterial cells were suspended into a cell
suspension buffer (100 mM Tris-HCl, 100 mM EDTA, pH 8.0).
Proteinase K (20 µl) was added to 400 µl of the suspension along
with 400 µl 1% SeaKem Gold Agar. These were mixed quickly, and
approximately 300 µl was dispensed into prepared plug molds. Once
solidified, the plugs were placed into 1.5 ml cell lysis buffer (50
mM Tris-HCl, 50 mM EDTA, pH 8.0, 1% Sarcosyl) and 40 µl of
Proteinase K and incubated for 1.5 h at 54˚C in a shaking water
bath. The plugs were washed twice in ultrapure water for 15 min in
a 50˚C water bath followed by four washings in Tris-ETDA (TE)
buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0). The washed plugs were
stored in TE buffer at 4˚C. For PFGE, the plugs were cut into 3-mm
by 9-mm pieces and then digested in 173 µl of sterile water, 2 µl
of bovine serum albumin, 20 l of 10XReAct II buffer, and 5 µl of
XbaI (10 U/l) at 37˚C in a shaking water bath for 1.5 h. The plugs
were run in a 1% agarose gel using a CHEF III Pulsed-Field System
(Bio-Rad) in 0.5% Tris-borate-EDTA buffer (Sigma) at 10˚C. The
parameters were set with the initial switch time at 2.2 sec, the
final switch time at 64 sec, a voltage of 6 V/cm, and a duration of
21 h. The gel was stained with ethidium bromide and recorded on a
Gel Doc system (Bio-Rad Laboratories, Inc., Hercules, CA). The file
images were processed by BioNumerics software (Applied Maths BVBA,
Kortrijk, Belgium).
Statistical analysis
All statistical analysis was performed using the
SPSS software (Version 18.0; SPSS, Inc.). The antibiotic resistance
data between the integron-positive isolates and integron-negative
isolates was compared and the χ2 test was used to
calculate the P-value in terms of the resistant and susceptible
numbers. P<0.05 was considered to indicate a statistically
significant difference.
Results
Departmental distribution of ABA in
the Sixth Affiliated Hospital of Guangzhou Medical University,
Qingyuan People's Hospital
A total of 54 from 74 CRABA strains were obtained
from the ICUs; the remaining 9 strains were obtained from the
department of Neurology, 3 strains were obtained from the
department of Infectious Diseases and the remaining strains were
distributed among other departments. Of the 22 non-CRABA strains,
10 were obtained from the ICUs and the departments of Neurology and
Urology each contributed 3 strains, while the remaining strains
were distributed among other departments. These observations
indicated that the ICUs have the highest number of contaminated
with ABA and are most prone to bacterial infections.
Antimicrobial resistance profile
Among the 96 ABA strains, the resistance rate of
strains harboring class I integrons towards tetracycline,
piperacillin, cefepime, ampicillin/sulbactam, meropenem,
gentamycin, levofloxacin and piperacillin/tazobactam was almost
100%, while that against ciprofloxacin, ceftazidime, cefotaxime and
imipenem was >90%. With the exception of
trimethoprim/sulfamethoxazole and amikacin, the resistance rate of
class I integron-positive strains to all antibiotics was increased
compared with of class I integron-negative strains. The results are
summarized in Table III.
| Table IIIComparison of drug resistance
phenotypes between 96 ABA profile I integron positive strains and
profile I integron negative strains. |
Table III
Comparison of drug resistance
phenotypes between 96 ABA profile I integron positive strains and
profile I integron negative strains.
| Antibacterials | D. R. R (%) | Profile I integron
positive strains (10 strains) | Profile I integron
negative strains (86 strains) | P-value |
|---|
| D. R. R (%) | I.R. (%) | S.R. (%) | D. R. R (%) | I.R. (%) | S.R. (%) |
|---|
| Ciprofloxacin | 85.4 | 90.0 | 10.0 | 0.0 | 84.9 | 0.0 | 15.1 | <0.05 |
| Tetracycline | 86.5 | 100.0 | 0.0 | 0.0 | 84.9 | 0.0 | 15.1 | <0.05 |
| Piperacillin | 89.6 | 100.0 | 0.0 | 0.0 | 88.4 | 0.0 | 11.6 | <0.05 |
| Ceftazidime | 85.4 | 90.0 | 0.0 | 10.0 | 84.9 | 1.2 | 14.0 | <0.05 |
| Cefotaxime | 83.3 | 90.0 | 0.0 | 10.0 | 82.6 | 8.1 | 9.3 | <0.05 |
| Cefepime | 87.5 | 100.0 | 0.0 | 0.0 | 86.1 | 1.2 | 12.8 | <0.05 |
|
Ampicillin/Sulbactam | 87.5 | 100.0 | 0.0 | 0.0 | 86.1 | 0.0 | 14.0 | <0.05 |
| Meropenem | 85.4 | 100.0 | 0.0 | 0.0 | 82.6 | 0.0 | 17.4 | <0.05 |
| Gentamycin | 86.5 | 100.0 | 0.0 | 0.0 | 84.9 | 0.0 | 15.1 | <0.05 |
| Imipenem | 83.3 | 90.0 | 0.0 | 10.0 | 82.6 | 0.0 | 17.4 | <0.05 |
| Levofloxacin | 86.5 | 100.0 | 0.0 | 0.0 | 84.9 | 0.0 | 15.1 | <0.05 |
| Bactrim | 71.9 | 30.0 | 0.0 | 70.0 | 76.7 | 0.0 | 23.3 | <0.05 |
| Colistin | 9.4 | 10.0 | 0.0 | 90.0 | 9.3 | 0.0 | 90.7 | <0.05 |
| Amikacin | 77.1 | 20.0 | 0.0 | 80.0 | 83.7 | 0.0 | 16.3 | <0.05 |
|
Piperacillin/tazobactam | 87.5 | 100.0 | 0.0 | 0.0 | 86.1 | 1.2 | 12.8 | <0.05 |
PFGE analysis and whole-genome
sequencing of 10 CRABA strains
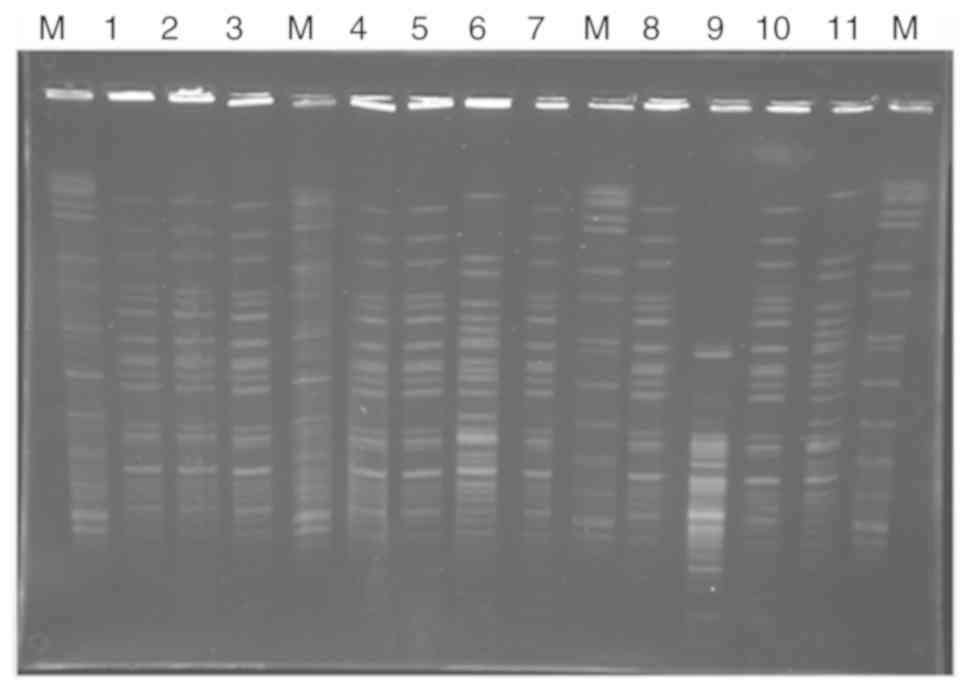
In this study, Ten Integron-positive strains could
be divided into four subtypes (A, B, C, and D) by PFGE (Fig. 2). Those strains harbored class I
integrons, of which 13.51% (10 of 74) were CRABA strains, and none
of them was non-CRABA strains. None of the 96 strains of ABA
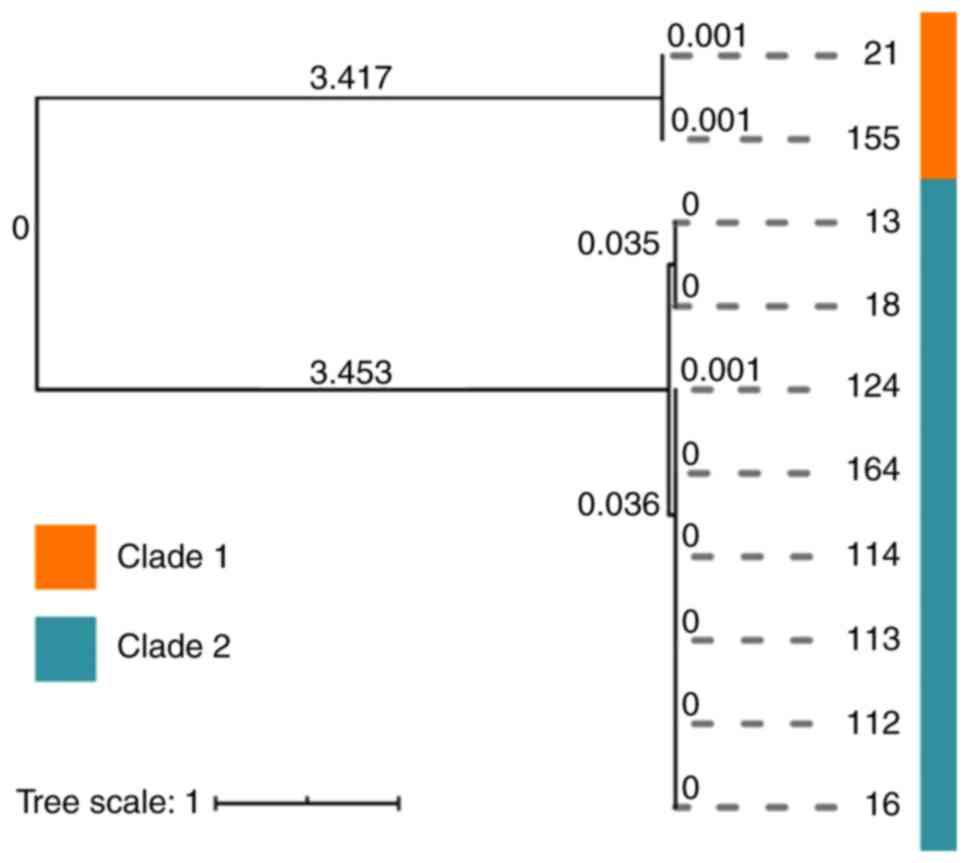
strains harbored class II and III integrons. A total of 10 A.
baumannii isolates were clustered into 2 main clades (Clade 1
and Clade 2), although the isolates had highly similar SNPs in each
clade, the genetic distance between the two clades was
comparatively high (Fig. 3).
Association between class I integrons
and antibacterial resistance
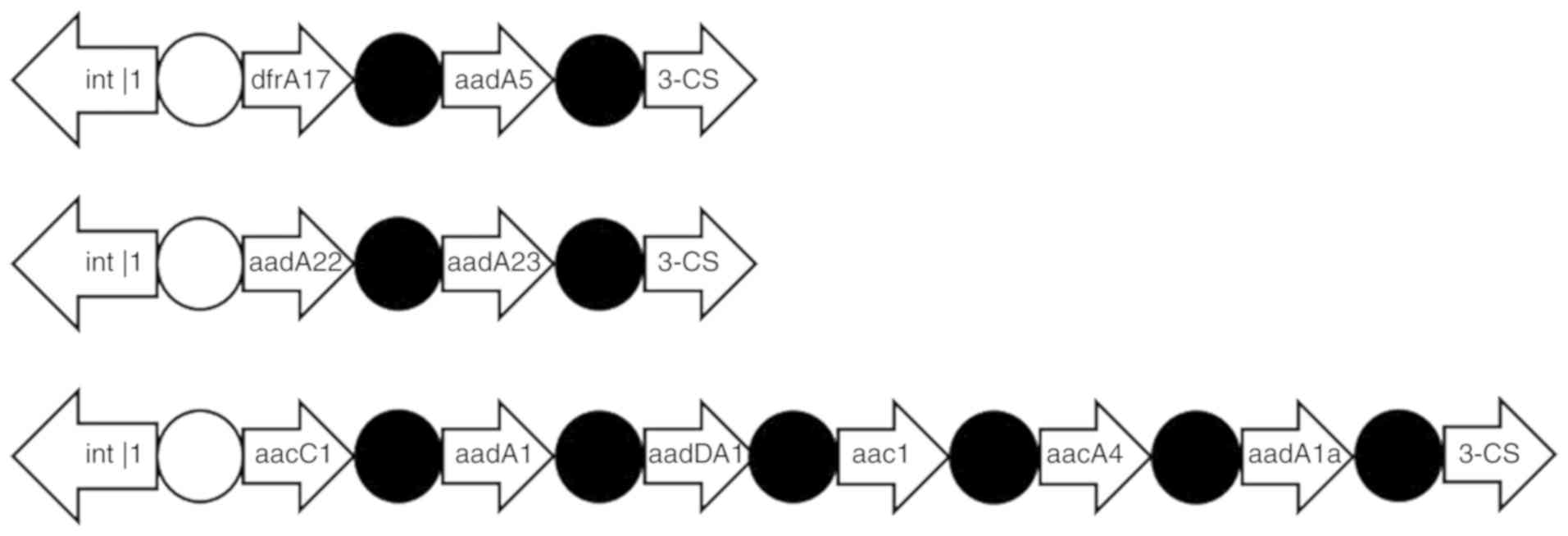
A total of 8 class I integron-positive strains and 1
class I integron-negative strain (n=9 strains) were sent for genome
sequencing and 10 drug resistance gene cassettes were detected
(Table IV). The integrated
substructure is presented in Fig. 4.
The actual drug resistance genes of 9 successfully sequenced
strains and the variable regions in the gene cassette are
summarized in Table IV. The drug
resistance rates of ABA class I integron-positive strains against
meropenem, imipenem, ceftazidime, cefotaxime, cefixime,
tetracycline, ciprofloxacin, cefazolin, piperacillin,
ampicillin/sulbactam, gentamicin, levofloxacin and
piperacillin/tazobactam were significantly increased compared with
the negative strains (P<0.05), indicating that the mechanism
underlying the acquisition of drug resistance in ABA strains was
closely associated with the presence of class I integrons. However,
the resistance rate of class I integron-negative ABA strains to a
number of antibiotics was also relatively high, indicating that
multidrug resistance among ABA strains is not only associated with
class I integrons but also with other drug resistance mechanisms,
which warrants further investigation.
| Table IVThe comparison between drug
resistance phenotype and variable region gene box. |
Table IV
The comparison between drug
resistance phenotype and variable region gene box.
| Sample number | Resistance
phenotype | The size of
variable Area (base pairs) | GM | AN | Bactrim | The combination of
variable region gene box |
|---|
| 8 | a, b, c, d, e, g,
h | 3000 | + | - | + | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
| 12 | a, b, c, d, e, g,
h | 3000 | + | - | + | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
| 24 | a, b, c, d, e,
h | 3000 | + | - | - | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
| 55 | a, b, c, d, e, g,
h | 3000 | + | + | + | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
| 60 | a, b, c, d, e, g,
h | 3000 | + | - | - | aacC1, aadA1,
aadDA1, aac1aacA4, aadA1a |
| 64 | a, b, c, d, e, g,
h | 3000 | + | + | - | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
| 67 | a, b, c, d, e, g,
h | 3000 | + | + | - | aadA22, aadA23 |
| 75 | a, b, c, d, e, g,
h | 3000 | + | - | - | dfrA17, aadA5 |
| 130 | a, b, c, d, e, g,
h | 3000 | + | - | - | aacC1, aadA1,
aadDA1, aac1, aacA4, aadA1a |
MLST data
A total of 21 sequence types (STs) were obtained
from 96 ABA strains, including 22 strains with ST208, 22 strains
with ST1145, 12 strains with ST1417 and 11 strains with ST195. The
MLSTs of CRABA and non-CRABA strains were compared, among which
ST208 and ST1145 were the most prominent STs of the CRABA strain
and ST1145 was the most prominent ST of the non-CRABA strain, while
others exhibited a scattered distribution (Table V). The genotypic distribution of each
branch is presented in Fig. 5.
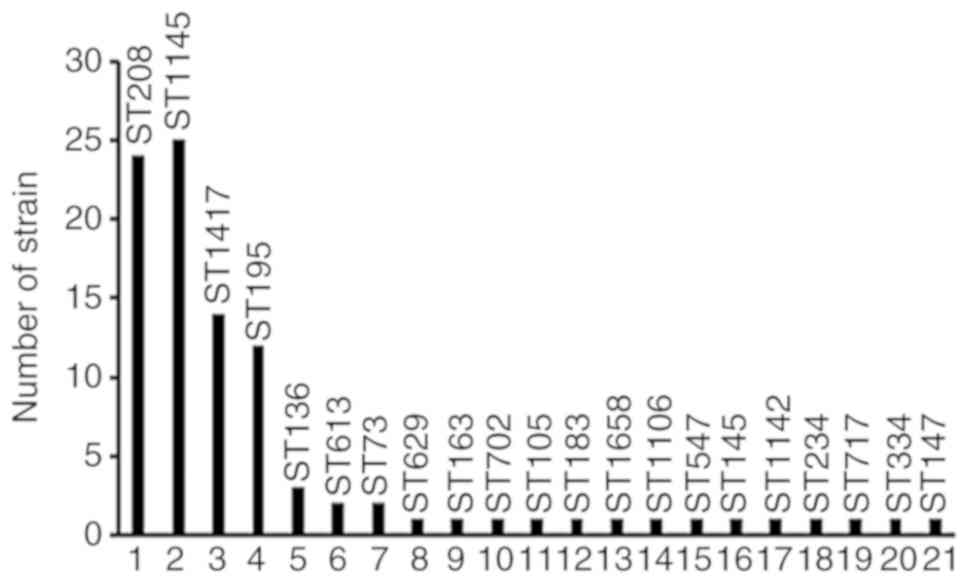 | Figure 5Distribution of various ST strains in
the Sixth Affiliated Hospital of Guangzhou Medical University. ST,
sequence type. 1. Intensive Care Unit, neurosurgery department,
Neurology department, and cardiology department; 2. Intensive Care
Unit, Tumor radiology department, Infectious Diseases department,
Neonatal pediatrics department, Hematology department, Neurological
surgery, and Respiratory Medicine department; 3. Intensive Care
Unit, Neurosurgery department, Burns and Plastic Surgery of
Department, Respiratory Medicine department, and Neurology
department; 4. Intensive Care Unit, Neurology department; 5.
Intensive Care Unit, Infectious Diseases department, and
Respiratory Medicine department; 6. Intensive Care Unit, Pediatrics
department; 7. Intensive Care Unit; 8. Intensive Care Unit; 9.
Neurology department; 10. Brain department; 11. Intensive Care
Unit; 12. Urological surgery; 13. Infectious diseases department;
14. Neonatal pediatrics department; 15. E.N.T department; 16.
Urological surgery department; 17. E.N.T department; 18. Intensive
Care Unit; 19. Intensive Care Unit; 20. Intensive Care Unit; 21.
Intensive Care Unit. |
| Table VNon-CRABA and CRABA strain multilocus
sequence typing results and distribution rate (%). |
Table V
Non-CRABA and CRABA strain multilocus
sequence typing results and distribution rate (%).
| Number of
strains | Housekeeping
genes | ST types | Distribution rate %
(n/74) |
|---|
| gltA | gyrB | gdhB | recA | cpn60 | gpi | rpoD |
|---|
| CRABA strain | 22 | 1 | 3 | 3 | 2 | 2 | 97 | 3 | 208 | 29.73 |
| 22 | 1 | 3 | 3 | 102 | 2 | 97 | 3 | 1,145 | 29.73 |
| 12 | 1 | 3 | 3 | 102 | 2 | 96 | 3 | 1,417 | 16.22 |
| 11 | 1 | 3 | 3 | 2 | 2 | 96 | 3 | 195 | 14.86 |
| 3 | 1 | 3 | 3 | 2 | 2 | 16 | 3 | 136 | 4.05 |
| 1 | 15 | 48 | 58 | 42 | 36 | 54 | 41 | 613 | 1.35 |
| 1 | 1 | 81 | 3 | 102 | 2 | 16 | 3 | 1,658 | 1.35 |
| 1 | 1 | 3 | 3 | 102 | 2 | 169 | 3 | 1,142 | 1.35 |
| 1 | 61 | 12 | 2 | 28 | 1 | 158 | 5 | 717 | 1.35 |
| Non-CRABA
strain | 2 | 1 | 3 | 3 | 2 | 2 | 97 | 3 | 208 | 9.09 |
| 3 | 1 | 3 | 3 | 102 | 2 | 97 | 3 | 1,145 | 13.64 |
| 2 | 1 | 3 | 3 | 102 | 2 | 96 | 3 | 1,417 | 9.09 |
| 1 | 1 | 3 | 3 | 2 | 2 | 96 | 3 | 195 | 4.55 |
| 1 | 15 | 48 | 58 | 42 | 36 | 54 | 41 | 613 | 4.55 |
| 2 | 1 | 47 | 53 | 1 | 1 | 59 | 32 | 73 | 9.09 |
| 1 | 33 | 31 | 2 | 28 | 1 | 77 | 5 | 629 | 4.55 |
| 1 | 33 | 12 | 40 | 26 | 32 | 91 | 5 | 163 | 4.55 |
| 1 | 33 | 50 | 80 | 28 | 1 | 164 | 5 | 702 | 4.55 |
| 1 | 31 | 45 | 8 | 6 | 30 | 53 | 6 | 105 | 4.55 |
| 1 | 1 | 17 | 56 | 1 | 1 | 91 | 26 | 183 | 4.55 |
| 1 | 1 | 34 | 3 | 28 | 1 | 144 | 45 | 1,106 | 4.55 |
| 1 | 1 | 3 | 3 | 2 | 2 | 157 | 3 | 547 | 4.55 |
| 1 | 21 | 35 | 2 | 28 | 1 | 52 | 4 | 145 | 4.55 |
| 1 | 21 | 48 | 58 | 42 | 36 | 109 | 4 | 234 | 4.55 |
| 1 | 33 | 12 | 59 | 11 | 32 | 11 | 5 | 334 | 4.55 |
| 1 | 1 | 54 | 62 | 31 | 4 | 55 | 45 | 147 | 4.55 |
Discussion
Integrons contain specific recombinant sites, which
can propagate antibiotic resistance, enable transmission of
different drug resistance gene cassettes from bacterial drug
resistance genes among different strains and lead to
multidrug-resistance among bacteria (18). The most common integron type of ABA
are the class I integrons, which contains various drug resistance
genes and is closely associated with the acquisition of drug
resistance (19).
In the present study, class I, II and III integrons
and variable regions were sequenced in 96 ABA strains. Of these, 10
strains harbored class I integrons, with a positive rate of 10.42%,
and all of them were CRABA strains; none of them harbored class II
and III integrons. The results of the present study are in contrast
to those of a previous study, in which Goudarzi and Azimi
(20) reported that the prevalence
of class I and II integrons was 66.7 and 20%, respectively, and
that, for the first time, class III integrons were reported in 3
A. baumannii strains (2.9%). These results differ from the
present study, considering the difference of regional distribution
and the sample resources of A. baumannii strains. Although
the positive rate of class I integrons was relatively low, the
variable region was amplified and the drug resistance gene
cassettes were detected in 9 of 10 class I integron-positive
strains, all of which were CRABA strains. These results show that
the distribution of class I integrons in CRABA was markedly
different compared with the non-CRABA strains and that class I
integron-positive strains serve an important role in the
acquisition of drug resistance, thus providing essential and novel
clinical insights.
Drug sensitivity analysis indicated that the
resistance rate of ABA strains isolated from the Sixth Affiliated
Hospital of Guangzhou Medical University, Qingyuan People's
Hospital, was as high as 89% to broad-spectrum penicillin, >80%
to penicillin, carbapenem and cephalosporins of 3 or 4 generations
and >70% to aminoglycosides. These aforementioned drugs can be
used in accordance with the patient condition per clinical
guidelines. While strains resistant to >3 antimicrobial drugs
are considered multi-resistant, class I integron-positive strains
are more dangerous. The results from the present study show that
ABA, a multidrug-resistant bacterium, can be treated with few drugs
and that CRABA or non-CRABA strains can simultaneously cause
infections within hospitals over a long time period and spread
widely through different modes of infection, thereby resulting in
nosocomial infections and leading to the generation of
drug-resistant strains; this is of critical concern. In the present
study, CRABA carried the resistant cassettes that contained aacC1,
aadA1, aadDA1, aac1, aacA4, aadA1a, aadA22, aadA23, dfrA17 and
aadA5, and aacC1, aadA1, aadDA1, aac1, aacA4 and aadA1a were the
main strains in the Sixth Affiliated Hospital of Guangzhou Medical
University, Qingyuan People's Hospital. This is markedly different
from another study in China: Huang et al (21) reported that most of the cassettes
belong to a class I integron (136/144) encoding arr3, aacA4,
dfrA17, aadA5, aadB, cat, blaOXA10, aadA1, aadA2, dfrA and aacC1,
and AadA2-HP-dfrA were the prevalent strains in the Second
Affiliated Hospital of Chongqing Medical University, China.
Currently, numerous class I integron resistance gene cassettes are
available. In the present study, 10 types of drug resistance gene
cassettes were detected: aacC1, aac1, aadDA1, aadA1a, aacA4,
dfrA17, aadA5, aadA1, aadA22 and aadA23. Among these, aadA1, aadA2,
aadA5 and aadA22 encoded aminoglycoside nucleotransferase, which
imparted spectinycin and streptomycin resistance, aacA4 encoded 6
N-aminoglycan drugs, resulting in tobramycin and amikacin
resistance and dfrA12 and dfrA17 encoded sulfonamides, resulting in
sulfanilamide resistance. These results demonstrated that the drug
resistance rate of integron-positive strains was increased compared
with integron-negative strains. Furthermore, other bacterial drug
resistance mechanisms include alterations in bacterial
permeability, drug targets, and metabolic pathways, production of
enzymes that eliminate antibiotics, active excretion of the
antibiotics and inhibition of antibiotic entry into bacteria
cells.
Pulse-field gel electrophoresis is a standard method
for bacterial molecular typing and assessment of homology and is
often used for ABA genotyping; however, it is only suitable for
short-term epidemiological studies (<3 months), and the method
is difficult to standardize and digitize (22); by contrast, MLST has high resolution
and can be used to detect housekeeping gene sequences (23). Compared with other typing methods,
MLST yields more accurate and realistic results, is superior and
reproducible, and enables differential typing of different strains.
MLST has been widely used worldwide for molecular epidemiological
studies of bacteria, such as S. aureus (24), Vibrio parahaemolyticus
(25), E. coli (26) and others. Previous studies have
performed MLST to study the molecular epidemiological
characteristics of ABA and reported numerous well-known multidrug
resistance STs and clone complexes (cc) (27,28). The
PubMLST database (https://pubmlst.org/abaumannii/) shows that CC92 is
currently the most prominent clone complex and that the majority of
ABA multi-drug-resistant strains worldwide are CC92 (29,30).
ST92 is the ancestor of the CC92 clone complex, indicating that
other ST types have evolved from ST92(31). Strains with the CC92 clone complex
are difficult to eliminate with antibiotics, thereby leading to its
increased drug resistance (32,33);
however, this requires further verification. In China, a few
studies have reported this, although Han et al (34) demonstrated that in Xi'an, Shanxi
province, 4 ST types (ST195, ST218, ST368 and ST208) were
identified among 47 multidrug-resistant A. baumannii strains
and that ST368 and ST208 have the closest genetic relationship. Lin
et al (35) reported that in
Anhui Province the drug-resistance rates of 87 strains of A.
baumannii to imipenem and meropenem were 74.7 and 66.7%,
respectively; 87 strains were subdivided into 42 ST-types and 37
strains were demonstrated to be dominant types and sub-listed in
the ST2 category. The ST2 A. baumannii belongs to the clone
complex CC1.
Concomitantly, multidrug-resistant ABA shares gene
homology in various regions in Europe via amplified fragment length
polymorphism analysis. Furthermore, European clones I, II and III
of multi-drug-resistant ABA are prevalent, among which European
clone II is the most prevalent (36). Higgins et al (37) genotyped CRABA strains from 32
countries via repetitive fragment PCR analysis and reported 8
worldwide clones, the European clone II being the most widely
distributed. Although the CC92 clone complex is widely prevalent
worldwide, different epidemic characteristics persist in different
regions. Zhong et al (32)
reported that ST92 and ST75 are the primary prevalent clones, while
another study reported that ST75 is probably the primary epidemic
type in Eastern China (38). He
et al (39) reported a novel
STn-1 (polymorphism 1-B1-3-2-2- 16-3, B1 being a novel allele
located at the gyrB locus) in Chengdu, in addition to ST92.
He et al (40) hypothesized
that ST138 and STn-1 may be the most widespread in Western China;
however, some other studies had already reported the primary
epidemic types in southern China. For instance, Fu et al
reported that CC92 was widespread in a number of cities in China
from 2004 to 2005, including ST75, ST90 and ST92(41).
In the present study, the changes in gyrB and
gpi were the most prominent, concurrent with previous
studies (42,43) and gyrB and gpi easily
undergo mutation and recombination during evolution. ST208 was
widely spread in the Sixth Affiliated Hospital of Guangzhou Medical
University, Qingyuan People's Hospital, accounting for 24 strains
(25%) of the CC92 clone, concurrent with previous nationwide and
international reports (31). The
proportions of ST1145 type and ST1417 were 26.04 and 14.58%,
respectively, potentially belonging to the new genotype and
warranting further investigation. Irrespective of the genotype, a
majority of the ABA strains were identified in the ICUs and
bacterial infections were most common in the ICUs, thus there is
the potential to lead to an outbreak, which warrants immediate
attention.
In conclusion, multidrug resistance in ABA strains
are closely associated with the presence of class I integrons. The
results of the present study highlighted the differences in STs
between CRABA and non-CRABA strains; CRABA strains containing
primarily ST208 and ST1145, and non-CRABA strains ST1145. The
results of the present study indicated that ABA in the Sixth
Affiliated Hospital of Guangzhou Medical University, Qingyuan
People's Hospital, is capable of horizontal transmission.
Furthermore, MLST can be used for clinical ABA genotyping. Owing to
multidrug resistance in ABA, further studies are required to
investigate the mechanism underlying bacterial drug resistance,
strictly monitor susceptible factors in hospitals, control
bacterial drug resistance, strengthen disinfection and isolation
methods in hospitals, improve safety awareness among the medical
staff and effectively control hospital infections.
Acknowledgements
We sincerely thank Shenzhen Center for Disease
Control and Prevention for PFGE analysis.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LX, SD and WW carried out the molecular genetic
studies, participated in the sequence alignment and drafted the
manuscript. YT and LC carried out the species identification. YL
and GZ participated in the susceptibility tests. JL and WT
participated in the PCR. BF conceived the study and participated in
its design and coordination, and helped to draft the manuscript.
All authors read and approved the final manuscript.
Funding
This study was supported by grants from the Natural
Science Foundation of China (grant no. 31770183) and the Medical
Science Technology Research Foundation of Guangdong (grant no.
A2015226), Guangdong Provincial Bureau of Traditional Chinese
Medicine (grant no. 20201407) and Qingyuan People's Hospital
Medical Scientific Research Fund Project (grant no. 20190209).
Ethics approval and consent to
participate
This study was approved by the Medical Ethics
committee of Qingyuan People's Hospital. Written informed consent
was obtained from each participant's legal guardian.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Da Silva GJ and Domingues S: Insights on
the horizontal gene transfer of carbapenemase determinants in the
opportunistic pathogen Acinetobacter baumannii.
Microorganisms. 4(E29)2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Antunes LC, Visca P and Towner KJ:
Acinetobacter baumannii: Evolution of a global pathogen.
Pathog Dis. 71:292–301. 2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lee CR, Lee JH, Park M, Park KS, Bae IK,
Kim YB, Cha CJ, Jeong BC and Lee SH: Biology of Acinetobacter
baumannii: Pathogenesis, antibiotic resistance mechanisms, and
prospective treatment options. Front Cell Infect Microbiol.
7(55)2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Fupin H, Guo Y, Zhu D, et al: CHINET
surveillance of bacterial resistance across China: Report of the
results in 2016. Chin J Infect Chemotherapy. 17:481–491. 2017.(In
Chinese).
|
|
5
|
Hu FP, Guo Y, Zhu DM, Wang F, Jiang XF, Xu
YC, Zhang XJ, Zhang CX, Ji P, Xie Y, et al: Resistance trends among
clinical isolates in China reported from CHINET surveillance of
bacterial resistance, 2005-2014. Clin Microbiol Infec. 22 (Suppl
1)(S9-S14)2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hall RM and Collis CM: Mobile gene
cassettes and integrons: Capture and spread of genes by
site-specific recombination. Mol Microbiol. 15:593–600.
1995.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Mazel D, Dychinco B, Webb VA and Davies J:
A distinctive class of integron in the Vibrio cholerae genome.
Science. 280:605–608. 1998.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Correia M, Boavida F, Grosso F, Salgado
MJ, Lito LM, Cristino JM, Mendo S and Duarte A: Molecular
characterization of a new class 3 integron in Klebsiella
pneumoniae. Antimicrob Agents Chemother. 47:2838–2843.
2003.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Cambray G, Guerout AM and Mazel D:
Integrons. Annu Rev Genet. 44:141–166. 2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Martinez-Freijo P, Fluit AC, Schmitz FJ,
Grek VS, Verhoef J and Jones ME: Class I integrons in Gram-negative
isolates from different European hospitals and association with
decreased susceptibility to multiple antibiotic compounds. J
Antimicrob Chemoth. 42:689–696. 1998.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen J, Li H, Yang J, Zhan R, Chen A and
Yan Y: Prevalence and characterization of integrons in multidrug
resistant Acinetobacter baumannii in eastern China: A
multiple-hospital study. Int J Environ Res Public Health.
12:10093–10105. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Machado E, Cantón R, Baquero F, Galán JC,
Rollán A, Peixe L and Coque TM: Integron content of
extended-spectrum-beta-lactamase-producing Escherichia coli strains
over 12 years in a single hospital in Madrid, Spain. Antimicrob
Agents Chemother. 49:1823–1829. 2005.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Urban C, Mariano N, Rahal JJ, Tay E, Ponio
C, Koprivnjak T and Weiss J: Polymyxin B-Resistant Acinetobacter
baumannii Clinical isolate susceptible to recombinant BPI and
cecropin P1. Antimicrob Agents Chemother. 45:994–995.
2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Bankevich A, Nurk S, Antipov D, Gurevich
AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S,
Prjibelski AD, et al: SPAdes: A new genome assembly algorithm and
its applications to single-cell sequencing. J Comput Biol.
19:455–477. 2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Nurk S, Bankevich A, Antipov D, Gurevich
AA, Korobeynikov A, Lapidus A, Prjibelski AD, Pyshkin A, Sirotkin
A, Sirotkin Y, et al: Assembling single-cell genomes and
mini-metagenomes from chimeric MDA products. J Comput Biol.
20:714–737. 2013.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Treangen TJ, Ondov BD, Koren S and
Phillippy AM: The Harvest suite for rapid core-genome alignment and
visualization of thousands of intraspecific microbial genomes.
Genome Biol. 15(524)2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Letunic I and Bork P: Interactive tree of
life (iTOL) v4: Recent updates and new developments. Nucleic Acids
Res. 47(W256-W259)2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Chen DQ, Jiang YT, Feng DH, Wen SX, Su DH
and Yang L: Integron mediated bacterial resistance and virulence on
clinical pathogens. Microb Pathog. 114:453–457. 2018. View Article : Google Scholar
|
|
19
|
Rowe-Magnus DA, Guerout AM and Mazel D:
Bacterial resistance evolution by recruitment of super-integron
gene cassettes. Mol Microbiol. 43:1657–1669. 2002.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Goudarzi M and Azimi H: Dissemination of
classes 1, 2, and 3 integrons in Acinetobacter baumannii
strains recovered from intensive care units using polymerase Chain
reaction-restriction fragment length polymorphism. Jundishapur J
Microbiol. 10(e13100)2017.PubMed/NCBI
|
|
21
|
Huang C, Long Q, Qian K, Fu T, Zhang Z,
Liao P and Xie J: Resistance and integron characterization of
Acinetobacter baumannii in a teaching hospital in Chongqing,
China. New Microbes New Infect. 8:103–108. 2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Tenover FC, Arbeit RD, Goering RV,
Mickelsen PA, Murray BE, Persing DH and Swaminathan B: Interpreting
chromosomal DNA restriction patterns produced by pulsed-field gel
electrophoresis: Criteria for bacterial strain typing. J Clin
Microbiol. 33:2233–2239. 1995.PubMed/NCBI
|
|
23
|
Maiden MC: Multilocus sequence typing of
bacteria. Annu Rev Microbiol. 60:561–588. 2006.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Miragaia M, Thomas JC, Couto I, Enright MC
and de Lencastre H: Inferring a population structure for
Staphylococcus epidermidis from multilocus sequence typing data. J
Bacteriol. 189:2540–2552. 2007.PubMed/NCBI View Article : Google Scholar
|
|
25
|
González-Escalona N, Martinez-Urtaza J,
Romero J, Espejo RT, Jaykus LA and DePaola A: Determination of
molecular phylogenetics of Vibrio parahaemolyticus strains by
multilocus sequence typing. J Bacteriol. 190:2831–2840.
2008.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Lacher DW, Steinsland H, Blank TE,
Donnenberg MS and Whittam TS: Molecular evolution of typical
enteropathogenic Escherichia coli: Clonal analysis by multilocus
sequence typing and virulence gene allelic profiling. J Bacteriol.
189:342–350. 2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Mugnier PD, Poirel L, Naas T and Nordmann
P: Worldwide dissemination of the blaOXA-23 carbapenemase gene of
Acinetobacter baumannii. Emerg Infect Dis. 16:35–40.
2010.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Diancourt L, Passet V, Nemec A, Dijkshoorn
L and Brisse S: The population structure of Acinetobacter
baumannii: Expanding multiresistant clones from an ancestral
susceptible genetic pool. PLoS One. 5(e10034)2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Spratt BG, Hanage WP and Feil EJ: The
relative contributions of recombination and point mutation to the
diversification of bacterial clones. Curr Opin Microbiol.
4:602–606. 2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Feil EJ, Holmes EC, Bessen DE, Chan MS,
Day NP, Enright MC, Goldstein R, Hood DW, Kalia A, Moore CE, et al:
Recombination within natural populations of pathogenic bacteria:
Short-term empirical estimates and long-term phylogenetic
consequences. Proc Natl Acad Sci USA. 98:182–187. 2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Karah N, Sundsfjord A, Towner K and
Samuelsen Ø: Insights into the global molecular epidemiology of
carbapenem non-susceptible clones of Acinetobacter
baumannii. Drug Resist Update. 15:237–247. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhong Q, Xu W, Wu Y and Xu H: Clonal
spread of carbapenem non-susceptible Acinetobacter baumannii
in an intensive care unit in a teaching hospital in China. Ann Lab
Med. 32:413–419. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Runnegar N, Sidjabat H, Goh HM, Nimmo GR,
Schembri MA and Paterson DL: Molecular epidemiology of
multidrug-resistant Acinetobacter baumannii in a single
institution over a 10-year period. J Clin Microbiol. 48:4051–4056.
2010.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Han L, Jie Q and Han SS: Clinical and
epidemiological characteristics of multidrug-resistant
Acinetobacter baumannii strains. J Xi'an Jiaotong
University. 38:872–877. 2017.PubMed/NCBI(In Chinese).
|
|
35
|
Lin W, Ying Y and Jiabin L: Multilocus
sequence typing of 87 strains of multidrug-resistant
Acinetobacter baumannii. Acta Universitatis Medicinalis
Anhui. 52:1480–1484. 2017.(In Chinese). PubMed/NCBI
|
|
36
|
van Dessel H, Dijkshoorn L, van der
Reijden T, Bakker N, Paauw A, van den Broek P, Verhoef J and Brisse
S: Identification of a new geographically widespread multiresistant
Acinetobacter baumannii clone from European hospitals. Res
Microbiol. 155:105–112. 2004.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Higgins PG, Dammhayn C, Hackel M and
Seifert H: Global spread of carbapenem-resistant Acinetobacter
baumannii. J Antimicrob Chemother. 65:233–238. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Liu S, Wang Y, Xu J, Li Y, Guo J, Ke Y,
Yuan X, Wang L, Du X, Wang Z, et al: Genome sequence of an
OXA23-producing, carbapenem-resistant Acinetobacter
baumannii strain of sequence type ST75. J Bacteriol.
194:6000–6001. 2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
He C, Xie Y, Fan H, Kang M, Tao C, Zhang
R, Hu Y, Chen Z and Wang L: Spread of imipenem-resistant
Acinetobacter baumannii of European clone II in Western
China. Int J Antimicrob Agents. 38:257–260. 2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
He C, Xie Y, Zhang L, Kang M, Tao C, Chen
Z, Lu X, Guo L, Xiao Y, Duo L and Fan H: Increasing imipenem
resistance and dissemination of the ISAba1-associated blaOXA-23
gene among Acinetobacter baumannii isolates in an intensive
care unit. J Med Microbiol. 60:337–341. 2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Fu Y, Zhou J, Zhou H, Yang Q, Wei Z, Yu Y
and Li L: Wide dissemination of OXA-23-producing
carbapenem-resistant Acinetobacter baumannii clonal complex
22 in multiple cities of China. J Antimicrob Chemother. 65:644–650.
2010.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Hamouda A, Evans BA, Towner KJ and Amyes
SG: Characterization of epidemiologically unrelated
Acinetobacter baumannii isolates from four continents by use
of multilocus sequence typing, pulsed-field gel electrophoresis,
and sequence-based typing of bla(OXA-51-like) genes. J Clin
Microbiol. 48:2476–2483. 2010.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Woo PC, Tsang AK, Wong AY, Chen H, Chu J,
Lau SK and Yuen KY: Analysis of multilocus sequence typing schemes
for 35 different bacteria revealed that gene loci of 10 bacteria
could be replaced to improve cost-effectiveness. Diagn Microbiol
Infect Dis. 70:316–323. 2011.PubMed/NCBI View Article : Google Scholar
|