Introduction
The most successful approach to treating a disease
has always been etiological therapy. In the case of bladder cancer,
however, this approach remains inapplicable, as the mechanism of
disease development has remained to be fully elucidated. Over 90%
of bladder tumours are transitional cell carcinomas - frequently
recurrent, but mostly non-invasive. However, they are a
heterogeneous group with at least two distinct subgroups with
different clinicopathological features-low-grade, non-infiltrating
cancers and high-grade, muscle-invasive cancers (1). In addition, based on their clinical
behavior, non-infiltrating tumours are subdivided into
non-progressive (70%) and progressive types (10-20%), whereas
high-grade, muscle-invasive cancers are subdivided into ones with a
relatively good prognosis and those with a poor prognosis with a
five-year survival rate of <50% (2). It is required to improve the current
knowledge on this clinical diversity and the specific molecular
mechanisms that underlie this variation in tumour behaviour.
Over 20% of non-invasive bladder tumours have been
indicated to have genomic imbalances, including losses in the short
(p) and long (q) chromosomal arm: 11p, 11q, 8p, 9, 17p, 3p and 12q
and gains in the 8q21, 13q21-q34, 1q31, 3q24-q26 and 1p22
chromosomes (3). Deletions in the
chromosomal regions 2q, 3p, 4p, 4q, 5q, 6q, 8p, 9p, 9q, 10q, 11p,
13q, 17p, 18q and Y and duplications/gains in 1q, 3q, 8q, 11q13,
17q and 20q and chromosome 7 are frequently detected (3-6).
In comparison, copy number variations are reported
in 30% of invasive bladder tumours, including elevated copy number
in the 1q23-24 chromosomal region, deletions in the 2q, 4q, 5p, 6q,
9p21.3, 8p23.1, 10p, 10q, 11p13 and 18q, as well as duplications in
the 6p22, 8p12, 8q22, 11q13, 19q13 and 20q regions (7). In 2003, Veltman et al (8) reported significant correlations
between copy number gain of CCNE1-containing regions and
gain of ERBB2, as well as a correlation between copy number
gain of CCND1 and deletion of TP53. So far, no
interdependence has been established between CNV findings and
tumour stage or grade.
Most of the studies related to quantitative genomic
changes were performed with low-resolution comparative genomic
hybridization (CGH). Studies on DNA imbalances in bladder cancer
performed using high-resolution microarray-based CGH (aCGH)
provided comprehensive information on quantitative genomic changes,
but due to the small sample size (usually between 7 and 40 bladder
cancer samples), they lacked statistical power (1,9-11).
The situation with expression microarray studies, which focused on
detecting changes in gene expression levels or mRNA levels, is
similar (12-15).
Despite the promising results from these studies, the current
understanding of the mechanisms involved in the progression of
bladder tumours remains insufficient. Advances in molecular methods
allowing for high-resolution locus-by-locus detection of CNVs may
possibly improve the understanding of the molecular pathology of
bladder cancer progression and facilitate the discovery of novel
drug targets and therapeutic approaches (16).
The aim of the present pilot study was to detect
CNVs related to tumour progression and metastasis of urothelial
carcinoma through whole-genome CNV scans of fresh frozen
samples.
Materials and methods
Ethics
This study was approved by the Ethical Committee of
the Medical University of Sofia (Sofia, Bulgaria; protocol No.
04/09/03/2018). Written informed consent and a questionnaire on
family history, as well as professional and environmental health
hazards, were obtained from all participants prior to tissue
collection.
Bladder cancer samples
A total of 30 bladder cancer samples from 6 females
and 24 males were collected for the present study. Samples were
collected for 12 months (January 2018 to January 2019) at the
Department of Urology, UMBALSM N.I. Pirogov and Department of
Urology, Tsaritsa Yoanna University Hospital. The clinical and
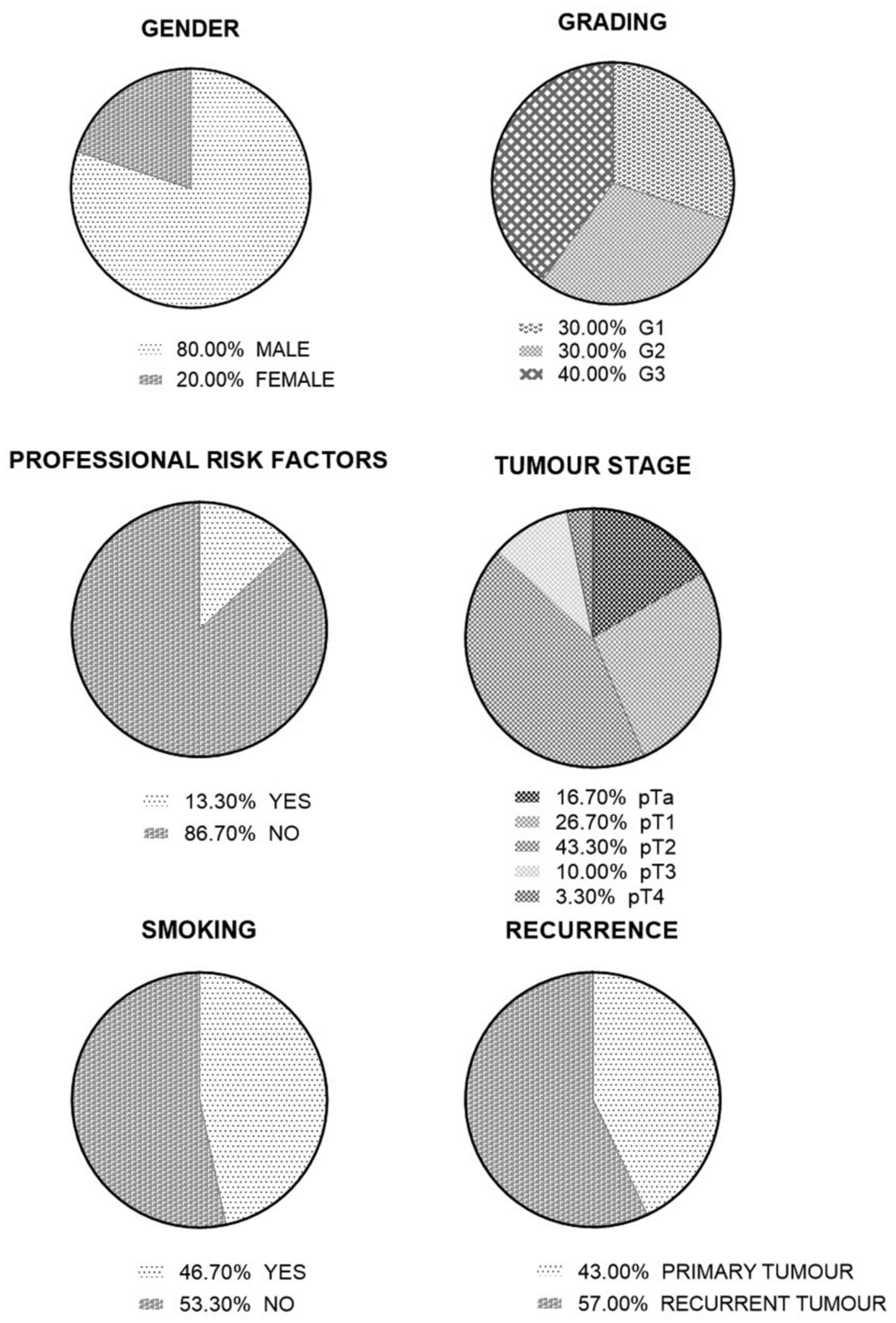
pathological characteristics of the studied cohort are described
according to the age of the patients, sex, smoking habits,
professional risk factors, tumor stage and grade (Table I).
 | Table ISummary of the clinical and
pathological characteristics of patients with bladder cancer used
in the present study. |
Table I
Summary of the clinical and
pathological characteristics of patients with bladder cancer used
in the present study.
| Parameter | Value |
|---|
| Mean age | 67.5±9.38
years |
| Smoking | 14 (46.7%) |
| Cigarette
consumption per day | 16.6±6.19 |
| Professional risk
factors | 4 (13.3%) |
| MMC therapy | 4 (13.3%) |
| BCG therapy | 1 (3.3%) |
| Grade | |
|
G1 | 9 (30%) |
|
G2 | 9 (30%) |
|
G3 | 12 (40%) |
| Recurrence | |
|
Recurrent | 17 (56.7%) |
|
Primary | 13 (43.3%) |
| Lymph Node
Metastasis | |
|
Present | 2 (6.7%) |
|
Absent | 28 (93.3%) |
aCGH
The samples were transported in sterile containers
at 4˚C to the genetic laboratory, where DNA was extracted using a
standard phenol-chloroform extraction protocol and stored at -20˚C.
Isolated DNA was quantitatively assessed spectrophotometrically
(NanoDrop® ND-2000c; Thermo Fisher Scientific, Inc.) and
qualitatively by horizontal low-voltage agarose gel electrophoresis
(Horizon 20-25; GibcoBRL; Thermo Fisher Scientific, Inc.).
Two independent platforms were used for the
detection of genomic imbalances: i) CytoChip ISCA 4x44K v1.0
(BlueGnome), scanned with an Agilent G2505 microarray scanner
(Agilent Technologies, Inc.) and analysed by BlueFuse Multi v3.1
(Illumina, Inc.) and ii) Infinium OncoArray-500K BeadChip
(Illumina, Inc.), scanned with iScan (Illumina, Inc.) and analysed
using KaryoStudio v.1.4 (Illumina, Inc.).
A total of 20 bladder cancer samples were analysed
with the OncoArray-500K BeadChip, 12 cancer samples were analysed
with the CytoChip ISCA 4x44K v1.0 and two tumour samples were
analysed using both methods to confirm the robustness of the
results.
All procedures, including sample preparation, sample
processing, hybridization, scanning and data analysis, were
performed using the manufacturers' standard protocols.
Genotype/phenotype interrelation
The interconnection between the genomic alterations
and the clinical phenotype of tumours was assessed by a detailed
analysis of publicly available databases such as the Database of
Genomic Variants (DGV; http://projects.tcag.ca/variation/) and the Catalogue
of Somatic Mutations in Cancer (Cosmic; https://cancer.sanger.ac.uk/cosmic).
Results
Bladder cancer samples
The clinical and pathological characteristics of the
cohort are described in Table I and
Fig. 1. The individual data for
each case are presented in Table
SI. The mean age of the studied patients was 67.77±9.3 years. A
total of 46.7% of patients were smokers, with an average cigarette
consumption of 16.9±6.19 per day. Four patients (13.3%) had
professional risk factors related to the transport, oil and
chemical industries.
CNV analysis
A total of two of the studied samples exhibited copy
number changes in >70% of the genome. Therefore, they did not
meet the qualitative criteria of the platform used and were
excluded from any further analysis. Data from the remaining 28
samples were included in the next analytical steps. A total of 524
aberrations, including 175 losses and 349 gains, were detected
(Table II).
| Table IITotal number of detected CNVs (loss
and gain) distributed by tumor stage and grade. |
Table II
Total number of detected CNVs (loss
and gain) distributed by tumor stage and grade.
| Aberrations | Tumor samples
distributed by T and G | Detect CNVs
distributed by T and G | Aberrations per
tumor |
|---|
| pTa loss | 4 | 20 | 5 |
| pT1 loss | 7 | 40 | 5.7 |
| pT2 loss | 13 | 78 | 6 |
| pT3 loss | 3 | 30 | 10 |
| pT4 loss | 1 | 7 | 7 |
| pTa gain | 4 | 44 | 10.7 |
| pT1 gain | 7 | 24 | 3.3 |
| pT2 gain | 13 | 256 | 19.2 |
| pT3 gain | 3 | 19 | 5.7 |
| pT4 gain | 1 | 6 | 6 |
| G1 loss | 7 | 41 | 5.8 |
| G2 loss | 9 | 61 | 6.8 |
| G3 loss | 12 | 73 | 6.1 |
| G1 gain | 7 | 62 | 8.8 |
| G2 gain | 9 | 133 | 14.8 |
| G3 gain | 12 | 154 | 12.8 |
According to the tumour stage, the aberrations were
distributed as follows: Ta, n=64 (12.2%); T1, n=64 (12.2%); T2,
n=334 (63.7%); T3, n=49 (9.4); and T4, n=13 (2.5%). The mean number
of aberrations per tumour was as follows: Ta, 16 (range, 0-44); T1,
9 (range, 0-19); T2, 25.7 (range, 0-70); T3, 16.3 (range, 13-24);
and T4, 13 (only one sample). Additionally, the number of
aberrations were the highest for T2-stage tumours and the lowest
for T1-stage tumours (Tables II
and SII)
Detected aberrations were distributed according to
tumour grade as follows: 19.7% (n=103) in G1 tumours, 37% (n=194)
in G2 and 43.3% (n= 227) in G3 tumours. The total number of
aberrations in G3 tumours was more than twice as high as that in
tumours of the lowest grade G1, but the average number of
aberrations per tumour was similar in the different tumour classes,
with 14.7 aberrations in G1 (range, 1-45), 21.5 in G2 (range, 0-91)
and 18.9 in G3 (range, 0-37) (Tables
II and SII).
Only 10.7% (n=3) of the 28 cancer samples carried no
chromosomal aberrations. The remaining 89.3% (n=25) displayed
multiple chromosomal copy number changes. Of all imbalances, 396
(75.6%) were located in autosomal chromosomes. Autosomal
chromosomal regions were classified based on the number of detected
imbalances as follows: i) Group 1 (0-5 CNVs detected), representing
7 chromosomal regions that harboured 6.3% (n=25) of all genomic
imbalances, with 21q (n=1), 12p (n=2), 12q (n=4), 16p (n=4), 16q
(n=4), 11p (n=5) and 18p (n=5); ii) Group 2 (6-9 CNVs detected),
representing 14 chromosomal regions that harbour 27% (n=107) of all
genomic imbalances, with 2p (n=8), 3p (n=7), 3q (n=7), 7p (n=7), 8p
(n=8), 14q (n=9), 15q (n=6), 17p (n=7), 18q (n=8), 19p (n=6), 19q
(n=9), 20p (n=8), 20q (n=9) and 22q (n=8); iii) Group 3 (≥10 CNVs
detected), representing the remaining 18 autosomal chromosomal
regions harbouring 66.7% (n=264) of all genomic imbalances, with 1p
(n=12), 1q (n=16), 2q (n=15), 4p (n=11), 4q (n=16), 5p (n=10), 5q
(n=18), 6p (n=23), 6q (n=13), 7q (n=10), 8q (n=12), 9p (n=25), 9q
(n=23), 10p (n=13), 10q (n=10), 11q (n=13), 13q (n=10) and 17q
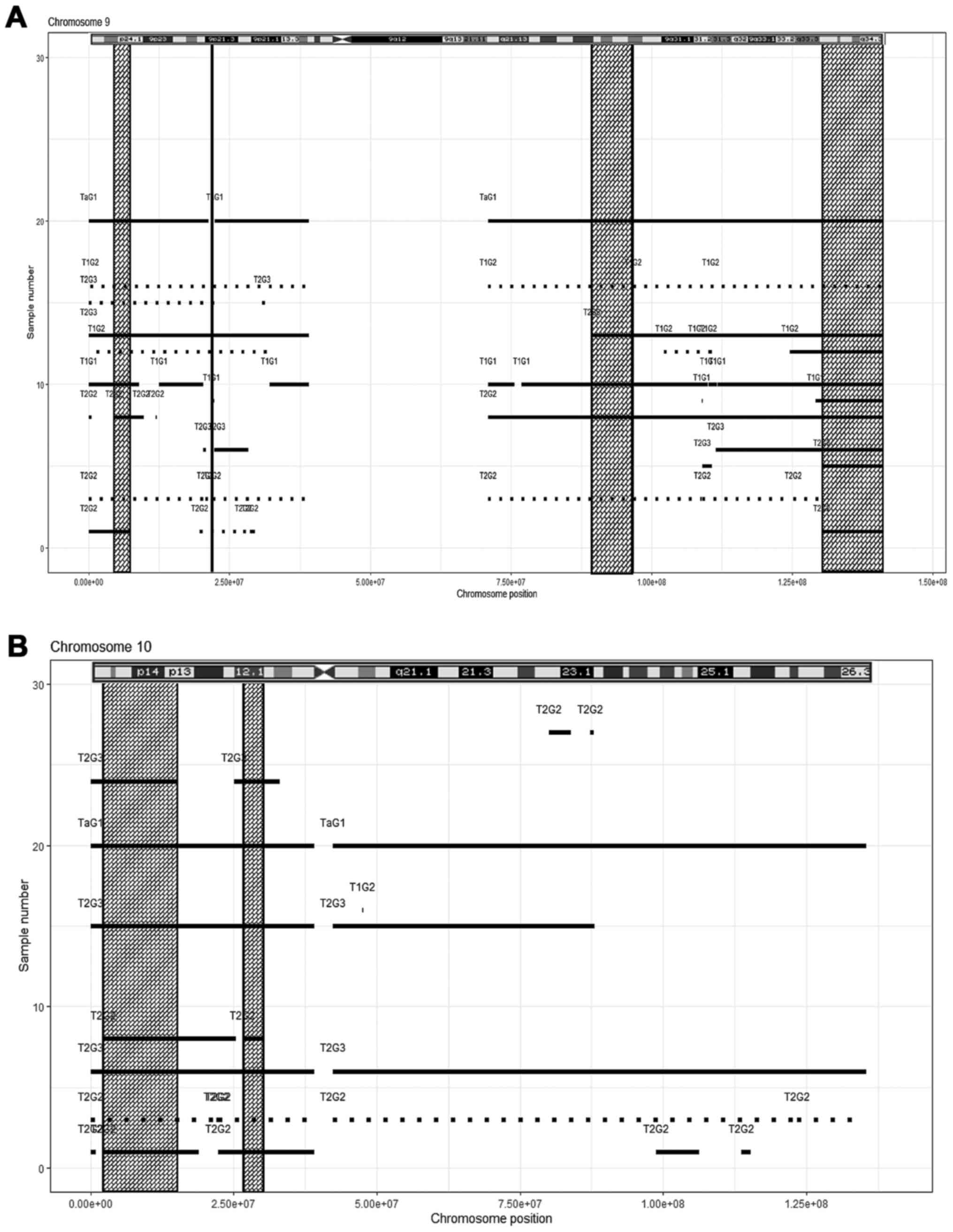
(n=14) (Table SII). Chromosome 9
was the most severely affected, displaying 25 CNVs in the short arm
and 23 in the long chromosomal arm, which accounted for 12.1% of
all autosomal aberrations.
The share of sex chromosomal imbalances stood at
24.4% (n=128). The highest frequency of CNVs per chromosome was
detected in the sex chromosomes, with 49 in the X (both losses and
gains) and 79 in the Y chromosome (only losses - partial or
complete) (Table SII).
Among the autosomal chromosomal regions with a high
number of CNVs (≥10) (n=264), 79.9% of variations (n=211) were
chromosomal gains, while 20.1% (n=53) were losses. Five
copy-neutral aberrations (LOH variants) were detected (Table III). Furthermore, 19.7% (n=52) of
the copy number changes were <800 kB, frequently reported in DGV
as polymorphic findings, so they were classified as ‘benign’. Of
the remaining CNVs, 76.1% (n=201) were labelled as pathogenic and
4.2% (n=11) were classified as variants with uncertain
significance. According to the tumour stage, the aberrations were
distributed in the following way: Ta, n=25; T1, n=30; T2, n=191;
T3, n=1; and T4, n=6. According to the tumour grade, the majority
of aberrations were present in G2-grade tumours and were
distributed as follows: G1, n=40 (15.15%); G2, n=122 (46.21%); and
G3, n=102 (38.64%) (Table
SII).
| Table IIILOH regions detected in the bladder
cancer samples. |
Table III
LOH regions detected in the bladder
cancer samples.
| Stage | Grade | Chr. | Cytoband | Start | End | Length in bp |
|---|
| Т2 | G2 | 6 | p12.1-p11.1 | 55721511 | 58767335 | 3045824 |
| Т2 | G2 | 7 | q21.3 | 94372640 | 97676259 | 3303619 |
| Т2 | G2 | 7 | q22.1q22.2 | 99552168 | 104336782 | 4784614 |
| Т2 | G3 | 11 | q14.1q14.2 | 81417643 | 86071005 | 4653362 |
| Т2 | G3 | 17 | q25.1-q25.3 | 73593574 | 77382564 | 3788990 |
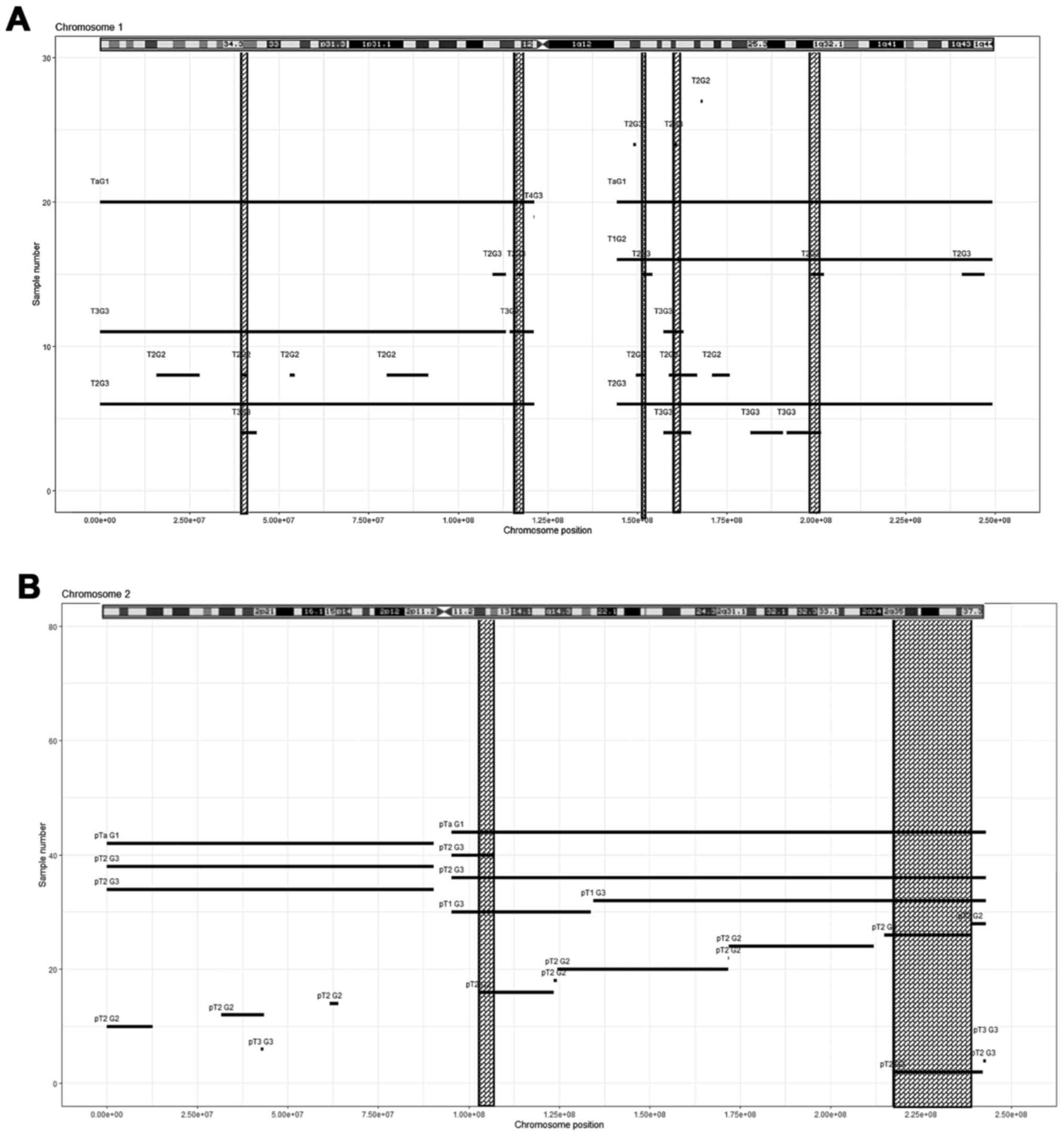
Among the pathogenic gains, 16.04 (n=45) had
overlapping regions in at least four different tumour samples,
mostly high-grade: Chromosome 1, five regions (Fig. 2A, Table
IV); chromosome 2, two regions (Fig. 2B, Table
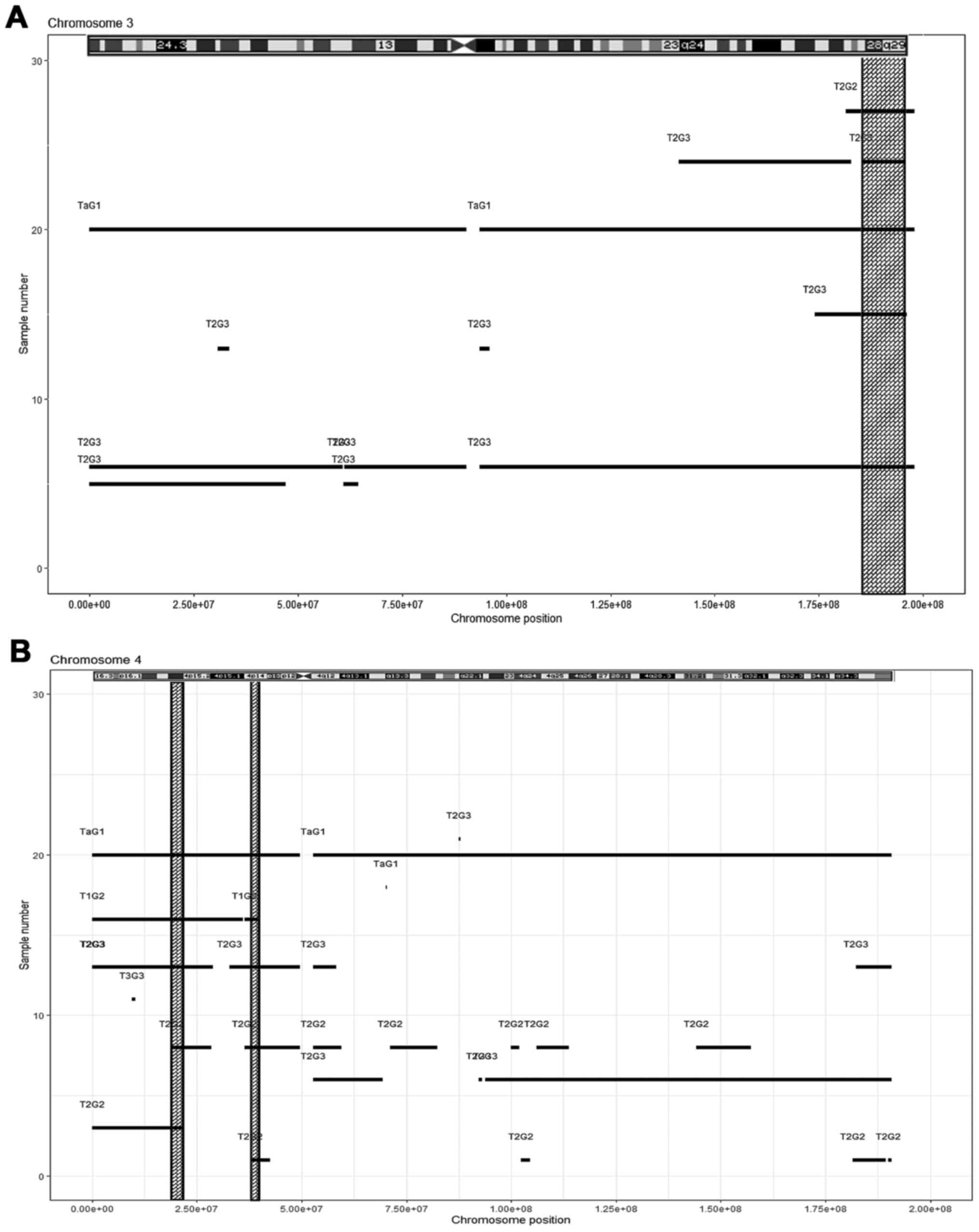
IV); chromosome 3, one region (Fig.
3A, Table IV); chromosome 4,
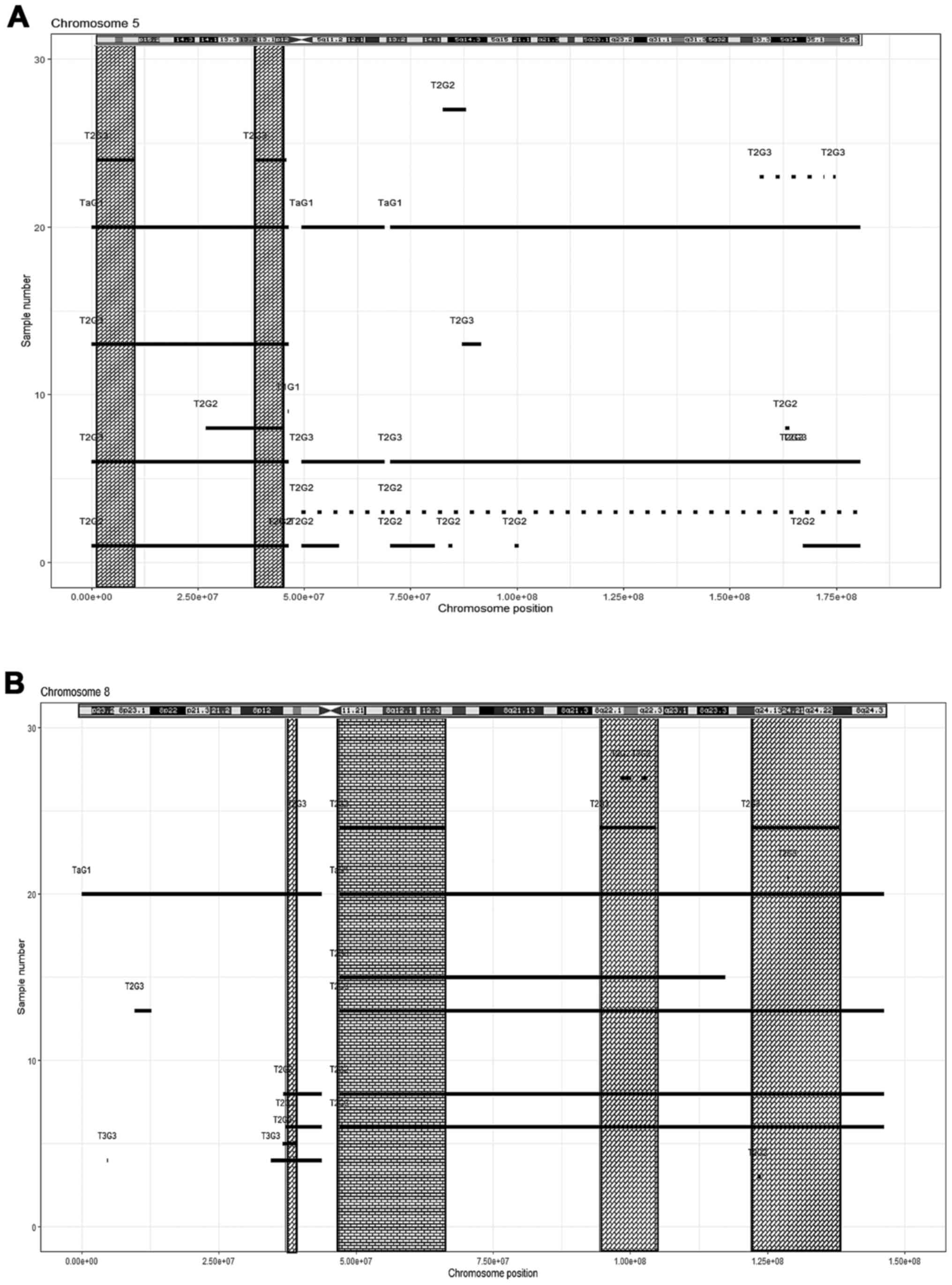
two regions (Fig. 3B, Table IV); chromosome 5, two regions
(Fig. 4A, Table IV); chromosome 8, four regions
(Fig. 4B, Table IV). In chromosome 9, four
overlapping chromosomal regions were detected, with both gains and
losses (Fig. 5A, Table IV). Furthermore, the following
overlapping regions were detected: Chromosome 10, two regions
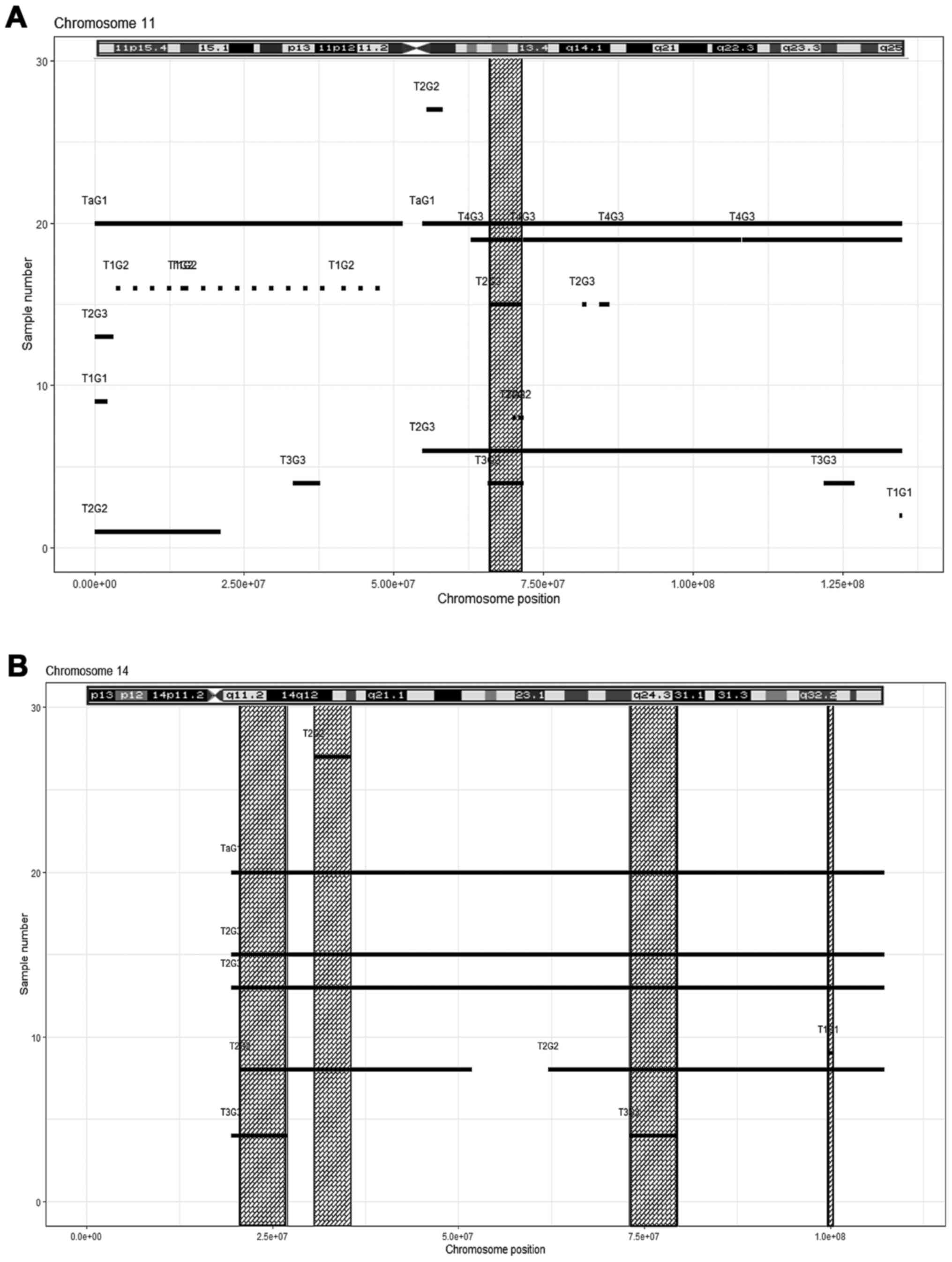
(Fig. 5B, Table IV); chromosome 11, one region
(Fig. 6A, Table IV); chromosome 14, four regions
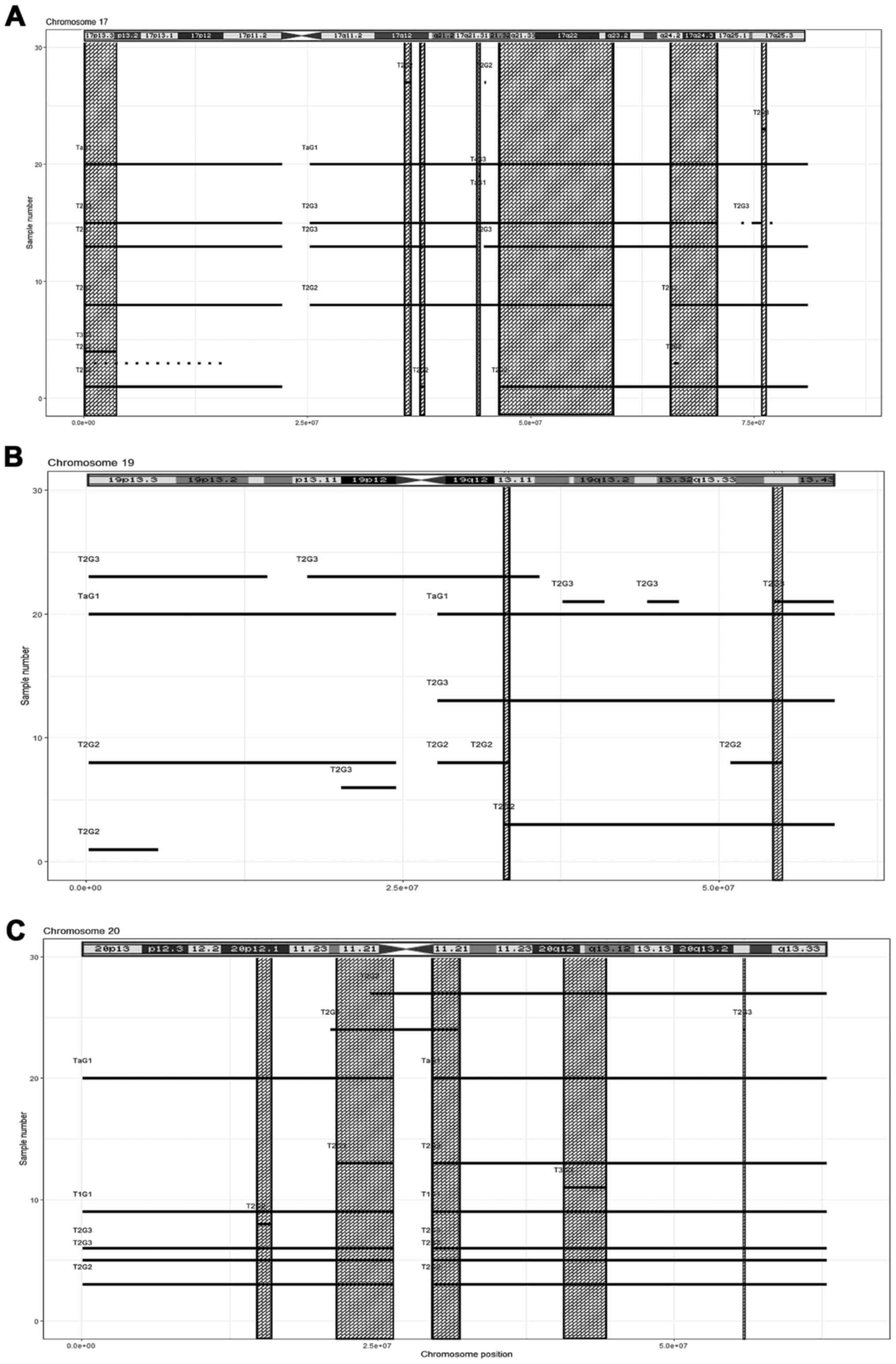
(Fig. 6B, Table IV); chromosome 17, seven regions
(Fig. 7A, Table IV); chromosome 19, two regions
(Fig. 7B, Table IV); and chromosome 20, five regions
(Fig. 7C, Table IV). The results obtained from the
follow-up analysis of genes from the pathogenic gain regions are
presented in Table V. This table
includes genes related to cancer treatment, resistance, initiation,
cell-cycle deregulation, tumor progression and metastases.
| Table IVCommon CNV regions in bladder cancer
samples. |
Table IV
Common CNV regions in bladder cancer
samples.
| Chromosome | Aberration | Number of
samples | Cytobands | Start | Stop | Length (kb) | CNV change
(-/+) |
|---|
| 1 | Ab. 1 | 5 | p34.3p34.2 | 39548798 | 41088676 | 1539878 | + |
| 1 | Ab. 2 | 4 | p13.1p12 | 116361026 | 118042757 | 1681731 | + |
| 1 | Ab. 3 | 5 | q21.3 | 151271782 | 152259742 | 987960 | + |
| 1 | Ab. 4 | 6 | q23.2q23.3 | 160382554 | 162080328 | 1697775 | + |
| 1 | Ab. 5 | 5 | q31.3q32.1 | 198410156 | 201437832 | 3027676 | + |
| 2 | Ab. 6 | 5 | q11.2q12.2 | 102626416 | 107105269 | 4478853 | + |
| 2 | Ab. 7 | 5 | 35q37.3 | 217186741 | 238597790 | 24990400 | + |
| 3 | Ab. 8 | 5 | q27.2q29 | 185131783 | 195623848 | 10492065 | + |
| 4 | Ab. 9 | 5 | p15.31p15.2 | 19047617 | 21526226 | 2478609 | + |
| 4 | Ab. 10 | 5 | p14 | 36317970 | 39459154 | 3141184 | + |
| 5 | Ab. 11 | 5 | p15.33p15.2 | 1159888 | 10124258 | 8964370 | + |
| 5 | Ab. 12 | 6 | p13.1p12 | 38542259 | 44812566 | 6270307 | + |
| 8 | Ab. 13 | 5 | p11.23p11.22 | 36778072 | 39223462 | 2445390 | + |
| 8 | Ab. 14 | 6 | q11.1q13.1 | 46923445 | 66409897 | 19486452 | + |
| 8 | Ab. 15 | 6 | q22.1q22.3 | 94359808 | 104577357 | 10217549 | + |
| 8 | Ab. 16 | 5 | q24.12q24.23 | 121918835 | 138034801 | 16115966 | + |
| 9 | Ab. 7 | 4 | p24.2p24.1 | 4483189 | 7324382 | 2841193 | _ |
| 9 | Ab. 18 | 5 | p24.2p24.1 | 4483189 | 7324382 | 2841193 | + |
| 9 | Ab. 19 | 5 | p21.3 | 21746274 | 22004153 | 257879 | - |
| 9 | Ab. 20 | 6 | p21.3 | 21746274 | 22004153 | 257879 | + |
| 9 | Ab. 21 | 2 | q21.33q22.32 | 89345014 | 96677307 | 7332293 | - |
| 9 | Ab. 22 | 4 | q21.33q22.32 | 89345014 | 96677307 | 7332293 | + |
| 9 | Ab. 23 | 1 | q33.3q34.3 | 130265117 | 141098428 | 10833311 | - |
| 9 | Ab. 24 | 9 | q33.3q34.3 | 130265117 | 141098428 | 10833311 | + |
| 10 | Ab. 25 | 6 | p15.3p13 | 2291574 | 15075299 | 12783725 | + |
| 10 | Ab. 26 | 6 | p12.1p11.23 | 26530405 | 30212755 | 3682350 | + |
| 11 | Ab. 27 | 6 | q13.1q13.4 | 65782622 | 71287123 | 5504501 | + |
| 14 | Ab. 28 | 5 | q11.2q12 | 20652555 | 27002379 | 6349824 | + |
| 14 | Ab. 29 | 6 | q12q13.2 | 30527028 | 35537512 | 5010484 | + |
| 14 | Ab. 30 | 5 | q24.2q31.1 | 72962189 | 79510706 | 6548517 | + |
| 14 | Ab. 31 | 6 | q32.2 | 99762857 | 100398318 | 635461 | + |
| 17 | Ab. 32 | 6 | p13.3p13.2 | 13905 | 3676393 | 3662488 | + |
| 17 | Ab. 33 | 6 | q12 | 35958791 | 36593228 | 634437 | + |
| 17 | Ab. 34 | 6 | q12 | 37673211 | 38105334 | 432123 | |
| 17 | Ab. 35 | 6 | q21.31 | 44161441 | 44351452 | 190011 | + |
| 17 | Ab. 36 | 6 | q21.32q23.2 | 46545363 | 59277614 | 12732251 | + |
| 17 | Ab. 37 | 5 | q24.2q25.1 | 65584651 | 70979004 | 5394353 | + |
| 17 | Ab. 38 | 5 | q25.3 | 75816274 | 76303552 | 487278 | + |
| 19 | Ab.39 | 5 | q13.11 | 33013506 | 33370161 | 356655 | + |
| 19 | Ab. 40 | 5 | q13.42 | 54332068 | 54919859 | 587791 | + |
| 20 | Ab. 41 | 6 | p12.1 | 14715679 | 16003225 | 1287546 | + |
| 20 | Ab. 42 | 7 | p11.22p11.1 | 21559089 | 26285899 | 4726810 | + |
| 20 | Ab. 43 | 8 | q11.21 | 29530880 | 31803506 | 2272626 | + |
| 20 | Ab. 44 | 8 | q12q13.12 | 40678430 | 44278445 | 3600015 | + |
| 20 | Ab. 45 | 8 | q13.31 | 55804501 | 55957204 | 152703 | + |
| Table VGenes related to cancer invasion and
metastasis among the common gain pathogenic variant. |
Table V
Genes related to cancer invasion and
metastasis among the common gain pathogenic variant.
| Array | Gene name (gene
symbol) |
|---|
|
arr1p34.3p34.2(39548798-41088676)x3 | Microtubule actin
crosslinking factor 1 (MACF1) |
|
arr1p34.3p34.2(39548798-41088676)x3 | L-myc-1
proto-oncogene (MYCL) |
|
arr1p34.3p34.2(39548798-41088676)x3 | Basic
helix-loop-helix (bHLH) |
|
arr1p34.3p34.2(39548798-41088676)x3 | Hes-related family
bHLH transcription factor (HEYL) |
|
arr1p13.1p12(116361026-118042757)x3 | Prostaglandin F2
receptor inhibitor (PTGFRN) |
|
arr1q21.3(151271782-152259742)x3 | S100 calcium
binding protein A1 (S100A1) |
|
arr1q21.3(151271782-152259742)x3 | Hornerin
(HRNR) |
|
arr1q21.3(151271782-152259742)x3 | Regulatory factor
X5 (RFX5) |
|
arr1q21.3(151271782-152259742)x3 | Proteasome 20S
subunit beta 4p (PSMB4) |
|
arr1q23.2q23.3(160382554-162080328)x3 | Nectin cell
adhesion molecule 4 (NECTIN4) |
|
arr1q23.2q23.3(160382554-162080328)x3 |
Beta-1,4-galactosyltransferase 3
(B4GALT3) |
|
arr1q23.2q23.3(160382554-162080328)x3 | Olfactomedin like
2B (OLFML2B) |
|
arr1q31.3q32.1(198410156-201437832)x3 | Ladinin 1
(LAD1) |
|
arr1q31.3q32.1(198410156-201437832)x3 | Nuclear receptor
subfamily 5 group A member 2 (NR5A2) |
|
arr1q31.3q32.1(198410156-201437832)x3 | Kinesin family
member 14 (KIF14) |
|
arr2q11.2q12.2(102626416-107105269)x3 | POU class 3
homeobox 3 (POU3F3) |
|
arr2q11.2q12.2(102626416-107105269)x3 | Four and a half LIM
domains 2 (FHL2) |
|
arr2q11.2q12.2(102626416-107105269)x3 | NCK adaptor protein
2 (NCK2) |
|
arr2q35q37.3(217186741-238597790)x3 | Wnt family member 6
(WNT6) |
|
arr2q35q37.3(217186741-238597790)x3 | CCR4-NOT
transcription complex subunit 9 (CNOT9) |
|
arr2q35q37.3(217186741-238597790)x3 | Serine/threonine
kinase 16 (STK16) |
|
arr2q35q37.3(217186741-238597790)x3 | Actin related
protein 2/3 complex subunit 2 (ARPC2) |
|
arr2q35q37.3(217186741-238597790)x3 | Disrupted in renal
carcinoma 3 (DIRC3) |
|
arr2q35q37.3(217186741-238597790)x3 | EPH receptor A4
(EPHA4) |
|
arr2q35q37.3(217186741-238597790)x3 | Paired box 3
(PAX3) |
|
arr2q35q37.3(217186741-238597790)x3 | Serpin family E
member 2 (SERPINE2) |
|
arr2q35q37.3(217186741-238597790)x3 | ADP ribosylation
factor like GTPase 4C (ARL4C) |
|
arr3q27.2q29(185131783-195623848)x3 | Mitogen-activated
protein kinase kinase kinase 13 (MAP3K13) |
|
arr3q27.2q29(185131783-195623848)x3 | Insulin like growth
factor 2 mRNA binding protein 2 (IGF2BP2) |
|
arr3q27.2q29(185131783-195623848)x3 | Transformer 2 beta
homolog (TRA2B) |
|
arr3q27.2q29(185131783-195623848)x3 | Replication factor
C subunit 4 (RFC4) |
|
arr3q27.2q29(185131783-195623848)x3 | Ribosomal protein
L39 like (RPL39L) |
|
arr3q27.2q29(185131783-195623848)x3 | BCL6 transcription
repressor (BCL6) |
|
arr3q27.2q29(185131783-195623848)x3 | MicroRNA 944
(MIR944) |
|
arr3q27.2q29(185131783-195623848)x3 | MicroRNA 5692c-1
(MIR5692C1) |
|
arr3q27.2q29(185131783-195623848)x3 | Hes family bHLH
transcription factor 1 (HES1) |
|
arr3q27.2q29(185131783-195623848)x3 | Carboxypeptidase N
subunit 2 (CPN2) |
|
arr4p15.31p15.2(19047617-21526226)x3 | MicroRNA 218-1
(MIR218-1) |
|
arr4p14(36317970–39459154)x3 | Toll like receptor
6 (TLR6) |
|
arr5p15.33p15.2(1159888–10124258)x3 | CLPTM1 like
(CLPTM1L) |
|
arr5p15.33p15.2(1159888–10124258)x3 | Iroquois homeobox 2
(IRX2) |
|
arr5p13.1p12(38542259-44812566)x3 | Oncostatin M
receptor (OSMR) |
|
arr5p13.1p12(38542259-44812566)x3 | Poly(A) binding
protein interacting protein 1 (PAIP1) |
|
arr5p13.1p12(38542259-44812566)x3 | Chromosome 5 open
reading frame 34 (C5orf34) |
|
arr5p13.1p12(38542259-44812566)x3 | Nicotinamide
nucleotide transhydrogenase (NNT) |
|
arr8p11.23p11.22(36778072-39223462)x3 | Phospholipid
phosphatase 5 (PLPP5) |
|
arr8p11.23p11.22(36778072-39223462)x3 | BAG cochaperone 4
(BAG4) |
|
arr8q11.1q13.1(46923445-66409897)x3 | CCAAT enhancer
binding protein delta (CEBPD) |
|
arr8q11.1q13.1(46923445-66409897)x3 | Lysophospholipase 1
(LYPLA1) |
|
arr8q11.1q13.1(46923445-66409897)x3 | Long intergenic
non-protein coding RNA 1606 (LINC01606) |
|
arr8q11.1q13.1(46923445-66409897)x3 | MIR124-2 host gene
(MIR124-2HG) |
|
arr8q11.1q13.1(46923445-66409897)x3 | Syndecan binding
protein (SDCBP) |
|
arr8q22.1q22.3(94359808-104577357)x3 | MicroRNA 5680
(MIR5680) |
|
arr8q22.1q22.3(94359808-104577357)x3 | Collagen triple
helix repeat containing 1 (CTHRC1) |
|
arr8q24.12q24.23(121918835-138034801)x3 | Hyaluronan synthase
2 (HAS2) |
|
arr8q24.12q24.23(121918835-138034801)x3 | Annexin A13
(ANXA13) |
|
arr8q24.12q24.23(121918835-138034801)x3 | NADH:ubiquinone
oxidoreductase subunit B9 (NDUFB9) |
|
arr9q33.3q34.3(130265117-141098428)x3 | SH2 domain
containing 3C (SH2D3C) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Endoglin
(ENG) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Cyclin dependent
kinase 9 (CDK9) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Leucine rich repeat
containing 8 subunit A (LRRC8A) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Vav guanine
nucleotide exchange factor 2 (VAV2) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Protein phosphatase
1 regulatory subunit 26 (PPP1R26) |
|
arr9q33.3q34.3(130265117-141098428)x3 | EGF like domain
multiple 7 (EGFL7) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Exonuclease 3'-5'
domain containing 3 (EXD3) |
|
arr9q33.3q34.3(130265117-141098428)x3 | NOTCH regulated
ankyrin repeat protein (NRARP) |
|
arr9q33.3q34.3(130265117-141098428)x3 | RAB, member RAS
oncogene family like 6 (RABL6) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Prostaglandin D2
synthase (PTGDS) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Calcium
voltage-gated channel subunit alpha1 B (ACNA1B) |
|
arr9q33.3q34.3(130265117-141098428)x3 | DEAD-box helicase
31 (DDX31) |
|
arr9q33.3q34.3(130265117-141098428)x3 | Small nucleolar RNA
host gene 7 (SNHG7) |
|
arr10p15.3p13(2291574-15075299)x3 | Kruppel like factor
6 (KLF6) |
|
arr10p15.3p13(2291574-15075299)x3 | RNA binding motif
protein 17 (RBM17) |
|
arr10p15.3p13(2291574-15075299)x3 |
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase
3 (PFKFB3) |
|
arr10p15.3p13(2291574-15075299)x3 | Kin17 DNA and RNA
binding protein (KIN) |
|
arr10p12.1p11.23(26530405-30212755)x3 | Microtubule
associated serine/threonine kinase like (MASTL) |
|
arr11q13.1q13.4(65782622-71287123)x3 | Carnitine
palmitoyltransferase 1A (CPT1A) |
|
arr11q13.1q13.4(65782622-71287123)x3 | MAS related GPR
family member D (MRGPRD) |
|
arr11q13.1q13.4(65782622-71287123)x3 | Fibroblast growth
factor 3 (FGF3) |
|
arr11q13.1q13.4(65782622-71287123)x3 | Fibroblast growth
factor 4 (FGF4) |
|
arr14q11.2q12(20652555-27002379) | NDRG family member
2 (NDRG2) |
|
arr14q11.2q12(20652555-27002379) | Methyltransferase
like 3 (METTL3) |
|
arr14q11.2q12(20652555-27002379) | Chromodomain
helicase DNA binding protein 8 (CHD8) |
|
arr14q11.2q12(20652555-27002379) | Proteasome 20S
subunit beta 5 (PSMB5) |
|
arr14q11.2q12(20652555-27002379) | Protein arginine
methyltransferase 5 (PRMT5) |
|
arr14q11.2q12(20652555-27002379) | DDB1 and CUL4
associated factor 11 (DCAF11) |
|
arr14q12q13.2(30527028-35537512)x3 | Egl-9 family
hypoxia inducible factor 3 (EGLN3) |
|
arr14q12q13.2(30527028-35537512)x3 | Sorting nexin 6
(SNX6) |
|
arr14q24.2q31.1(72962189-79510706)x3 | PNMA family member
1 (PNMA1) |
|
arr14q24.2q31.1(72962189-79510706)x3 | Activator of HSP90
ATPase activity 1 (AHSA1) |
|
arr14q32.2(99762857-100398318)x3 | MicroRNA 5698
(MIR5698) |
| arr
17p13.3p13.2(13905-3676393)x3 | Reticulon 4
receptor like 1 (RTN4RL1) |
| arr
17p13.3p13.2(13905-3676393)x3 | Rouble C2 domain
beta (DOC2B) |
|
arr17q12(35958791-36593228)x3 | TBC1 domain family
member 3 (TBC1D3) |
|
arr17q12(37673211-38105334)x3 | Migration and
invasion enhancer 1 (MIEN1) |
|
arr17q12(37673211-38105334)x3 | Growth factor
receptor bound protein 7 (GRB7) |
|
arr17q21.32q23.2(46545363-59277614)x3 | Homeobox B7
(HOXB7) |
|
arr17q21.32q23.2(46545363-59277614)x3 | Speckle type
BTB/POZ protein (SPOP) |
|
arr17q21.32q23.2(46545363-59277614)x3 | Distal-less
homeobox 4 (DLX4) |
|
arr17q21.32q23.2(46545363-59277614)x3 | MicroRNA 454
(MIR454) |
|
arr17q21.32q23.2(46545363-59277614)x3 | ANKRD40 C-terminal
like (ANKRD40CL) |
|
arr17q21.32q23.2(46545363-59277614)x3 | Sperm associated
antigen 9 (SPAG9) |
|
arr17q21.32q23.2(46545363-59277614)x3 | A-kinase anchoring
protein 1 (AKAP1) |
|
arr17q21.32q23.2(46545363-59277614)x3 | Tripartite motif
containing 37 (TRIM37) |
|
arr17q24.2q25.1(65584651-70979004)x3 | Karyopherin subunit
alpha 2 (KPNA2) |
|
arr17q24.2q25.1(65584651-70979004)x3 | Mitogen-activated
protein kinase kinase 6 (MAP2K6) |
|
arr17q24.2q25.1(65584651-70979004)x3 | AS1 SOX9 antisense
RNA 1 (SOX9) |
|
arr17q25.3(75816274-76303552)x3 | TNRC6C antisense
RNA 1 (TNRC6C-AS1) |
|
19q13.42(54332068-54919859)x3 | Leukocyte
immunoglobulin like receptor B2 (LILRB2) |
|
19q13.42(54332068-54919859)x3 | CCR4-NOT
transcription complex subunit 3 (CNOT3) |
|
arr20p12.1(14715679-16003225)x3 | Mono-ADP
ribosylhydrolase 2 (MACROD2) |
|
arr20p11.22p11.1(21559089-26285899)x3 | Ninein like
(NINL) |
|
arr20p11.22p11.1(21559089-26285899)x3 | GINS complex
subunit 1 (GINS1) |
|
arr20q11.21(29530880-31803506)x3 | PLAG1 like zinc
finger 2 (PLAGL2) |
|
arr20q12q13.12(40678430-44278445)x3 | Serine and arginine
rich splicing factor 6 (SRSF6) |
|
arr20q12q13.12(40678430-44278445)x3 | Semenogelin 1
(SEMG1) |
|
arr20q12q13.12(40678430-44278445)x3 | Translocase of
outer mitochondrial membrane 34 (TOMM34) |
|
arr20q13.31(55804501-55957204)x3 | Ribonucleic acid
export 1 (RAE1) |
Comparison between the two CNV
detection platforms
The two samples analysed with both platforms
demonstrated no difference in the detected chromosomal
abnormalities. Due to the higher resolution of the Infinium
OncoArray-500K BeadChip (Illumina, Inc.), the coordinates of the
detected aberrations were mapped comparatively more precisely. The
region arr16q23.1q24 (74356681-88675439)x1, designated as one
region by CytoChip ISCA 4x44K v1.0, was recognized as two regions,
namely arr16q23.1(74500123-75740477)x1 and arr16q23.1
(75766088-77151891)x0, by the Infinium OncoArray-500K BeadChip. The
latter platform detected an additional chromosomal region,
arr16p12.1p11 (24577766-35173765)x3, which was below the resolution
of the CytoChip ISCA 4x44K v1.0 arrays. Only the design of the
Infinium OncoArray-500K BeadChip allowed for the detection of LOH
regions.
LOH variants
Among the 19 tumour samples analysed with the
Infinium OncoArray-500K BeadChip, five LOH variants were detected
in chromosomes 6, 7, 11 and 17 Table
V. These variants contained 176 genes (Table SIII).
Discussion
In the present study, a CNV analysis of 30 bladder
cancer samples was performed. Despite the small number of patients
in the present cohort, the epidemiological data obtained
demonstrated the role of external hazardous factors in the
development of bladder cancer. More than 46% of patients
demonstrated unhealthy smoking habits and 13.3% had professional
risk factors. These findings are consistent with results of
previously conducted larger studies (17).
The high number of detected CNVs testifies the high
level of genomic instability observed in both high-grade tumours
(G3) and low-grade tumours (G1), and is in concordance with results
of previous studies (1). The
results obtained with regard to the role of Y-chromosome imbalances
in bladder cancer were consistent with the insight gained in a
study that demonstrated a strong tendency of Y-chromosome loss
(18), but were partially in
contrast to the studies of Conconi et al (1) from 2014 and Panani and Roussos
(19) from 2006, where Y-chromosome
amplification was detected. Due to the small size of the Y
chromosome, the phenomenon of age-related loss of the Y chromosome
and the development of bladder cancer in adulthood (20), it may be assumed that the loss of
the Y chromosome in bladder cancer is a non-specific
phenomenon.
A high number of genes related to cancer treatment,
resistance, initiation and cell-cycle deregulation were located in
the regions with pathogenic gains. Genes related to invasion and
metastasis, in line with the aims of the present study, were the
focus of the consequent data analysis. These genes are discussed
below.
In tree CNV regions in chromosome 9, both losses and
gains were detected: arr9p24.2p24.1 (4483189-7324382)x1,
arr9p24.2p24.1 (4483189-7324382)x3 arr9p21.3 (21746274-22004153)x1,
arr9p21.3 (21746274-22004153)x3, arr9q21.33q22.32
(89345014-96677307)x1 and arr9q21.33q22.32 (89345014-96677307)x3.
These could be unspecific events due to high level of genomic
instability in those regions. In the fourth region of chromosome 9,
arr9q33.3q34.3(130265117-141098428), nine gains and only one loss
were observed; thus, the further analysis so only focused on gain
variants. The genes that may be related to invasion and metastasis
in this region were SH2D3C, ENG, CDK9,
LRRC8A, VAV2, PPP1R26, EGFL7,
EXD3, NRARP, RABL6, PTGDS,
DDX31, SNHG7 and CACNA1B.
Among the regions with a gain in chromosome 17,
arr17q21.31(44161441-44351452)x3, a systemic gain in DGV was
commonly present and thus, it may be classified as a likely benign
variant. Among the other gained regions, the genes that may be
related to tumour progression and metastasis were RTN4RL1,
DOC2B, TBC1D3, MIEN1, GRB7,
HOXB7, SPOP, DLX4, MIR454,
ANKRD40CL, SPAG9, AKAP1, TRIM37,
KPNA2, MAP2K6, SOX9 and TNRC6C-AS1.
In the gained region of chromosome 19, only
LILRB2 and CNOT3 genes in
arr19q13.42(54332068-54919859)x3 were indicated to be potentially
related to cancer progression.
Certain genes detected in the regions with gains
discussed above have already been reported to be involved in
bladder cancer genesis and progression. These genes included the
following: PFDN2 located on 1q23.3, detected in the urinary
DNA with aCGH technique (21);
COL6A3 located in 2q37.3, which promotes
epithelial-mesenchymal transition in bladder cancer cells via the
TGF-β/Smad pathway (22);
DNER in 2q36.3, involved in proliferation, migration and
invasion by regulating the activation of the PI3K/AKT pathway
(23). OSMR in 5p13.1 was
indicated to be closely associated with cell growth and
differentiation, inflammation and enhancement of metastatic
capacity in urinary bladder cancer (24). MYC, located in 8q24.21, has
been reported in numerous studies to be involved in cell growth and
migration in bladder cancer (25).
Genes DDX31 and SNHG7, which are oncogenes that have
been reported to be dysregulated in various tumour types, have also
been previously indicated to be involved in bladder cancer
metastases (26,27). Both are located in the ‘gain’
chromosomal region in the terminal end of chromosome 9,
arr9q33.3q34.3(130265117-141098428). The SLC39A11 located in
17q24.3-q25.1 has been reported to be associated with survival of
patients with bladder cancer (28).
The enhanced methylation status of PRAC1 in 17q21.32 has been
linked to a high recurrence rate and progression in patients with
bladder cancer (29). Chen et
al (30) indicated that the
KAT7 in 17q21.33, promotes cell proliferation in bladder
cancer samples.
The genes detected in the LOH regions may be
investigated in further studies and functional analyses regarding
their potential tumour-suppressor role may be performed.
One limitation of the present study is the small
sample size. However, even in this small cohort, a large quantity
of aberrations were detected, which are difficult to classify
without any strict statistical, histological and bioinformatical
criteria, which unfortunately have not yet been developed.
Additionally, a relatively small sample cohort may lead to
statistical errors with overrepresentation of certain aberrations
and therefore, the prevalence rate of particular chromosomal
aberrations may not be representative of bladder cancer imbalances
in general. Another limitation of the study is that the analysis of
genomic imbalances was performed at the DNA level only, so the next
step should be gene expression analysis or functional investigation
of identified genes that have not already been reported elsewhere
to be involved in bladder cancer. Despite the limitations of the
present study, the results obtained are of significant scientific
value, as they provide certain mechanisms that may be responsible
for tumour invasion caused by genetic imbalances related to the
activation of genes with metastatic and proliferative potential. A
total of 42 recurrent CNVs, mostly in high-grade bladder tumours,
were detected in chromosomes 1-5, 8-11, 14, 17, 19 and 20.
Furthermore, in the present study, genes potentially related to the
metastatic potential of uroepithelial tumours were identified that
may be further studied as possible targets for precision therapy.
Finally, five LOH variants in high-grade bladder cancer tumours
were described.
In conclusion, the present study demonstrated that
applying genomic approaches to bladder cancer research is crucial
for furthering the current knowledge on the progression of the
disease and the inclusion of these technologies as part of routine
patient care, thus accelerating the implementation of a
personalized therapeutic approach.
Supplementary Material
Clinical and pathological
characteristics of patients with bladder cancer.
Number of detected CNVs distributed by
chromosomal arm (p and q), tumour stage and grade.
Genes located in the detected LOH
regions.
Acknowledgements
Bulgarian Ministry of Education and Science under
the National Program for Research ‘Young Scientists and
Postdocotral Students’.
Funding
Funding: This work was funded by the Bulgarian Ministry of
Education (grant no. DM13/4, 2017) and the Medical University of
Sofia (grant no. D-60/03.05.2018).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
OA was involved in the conception of the study,
sample processing and writing of the manuscript. BM, SR and CS were
involved in tumour sample collection. ZH and DN were involved in
DNA isolation. LB, RV and SH were involved in CNV detection. VS and
MG analysed the data and prepared the figures. DS and RS were
involved in statistical analyses. DT was involved in study design
and manuscript revision. OA, LB and RV confirm the authenticity of
all the raw data. All the authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethical Committee of
the Medical University of Sofia (Sofia, Bulgaria; protocol no.
04/09/03/2018). Written informed consent was obtained from the
participants prior to tissue collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Conconi D, Panzeri E, Redaelli S, Bovo G,
Viganò P, Strada G, Dalprà L and Bentivegna A: Chromosomal
imbalances in human bladder urothelial carcinoma: Similarities and
differences between biopsy samples and cancer stem-like cells. BMC
Cancer. 14(646)2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kawanishi H, Takahashi T, Ito M, Matsui Y,
Watanabe J, Ito N, Kamoto T, Kadowaki T, Tsujimoto G, Imoto I, et
al: Genetic analysis of multifocal superficial urothelial cancers
by array-based comparative genomic hybridisation. Br J Cancer.
97:260–266. 2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Kallioniemi A, Kallioniemi OP, Citro G,
Sauter G, DeVries S, Kerschmann R, Caroll P and Waldman F:
Identification of gains and losses of DNA sequences in primary
bladder cancer by comparative genomic hybridization. Genes
Chromosomes Cancer. 12:213–219. 1995.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sauter G: Rudolf Virchow Prize 1997.
Molecular cytogenetic analysis of superficial urothelial cancer of
the bladder. Verh Dtsch Ges Pathol. 81:18–27. 1997.PubMed/NCBI(In German).
|
|
5
|
Voorter C, Joos S, Bringuier PP, Vallinga
M, Poddighe P, Schalken J, du Manoir S, Ramaekers F, Lichter P and
Hopman A: Detection of chromosomal imbalances in transitional cell
carcinoma of the bladder by comparative genomic hybridization. Am J
Pathol. 146:1341–1354. 1995.PubMed/NCBI
|
|
6
|
Zaharieva BM, Simon R, Diener PA,
Ackermann D, Maurer R, Alund G, Knönagel H, Rist M, Wilber K,
Hering F, et al: High-throughput tissue microarray analysis of
11q13 gene amplification (CCND1, FGF3, FGF4, EMS1) in urinary
bladder cancer. J Pathol. 201:603–608. 2003.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Huang WC, Taylor S, Nguyen TB, Tomaszewski
JE, Libertino JA, Malkowicz SB and McGarvey TW: KIAA1096, a gene on
chromosome 1q, is amplified and overexpressed in bladder cancer.
DNA Cell Biol. 21:707–715. 2002.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Veltman JA, Fridlyand J, Pejavar S, Olshen
AB, Korkola JE, DeVries S, Carroll P, Kuo WL, Pinkel D, Albertson
D, et al: Array-based comparative genomic hybridization for
genome-wide screening of DNA copy number in bladder tumors. Cancer
Res. 63:2872–2880. 2003.PubMed/NCBI
|
|
9
|
Bellmunt J, Selvarajah S, Rodig S, Salido
M, de Muga S, Costa I, Bellosillo B, Werner L, Mullane S, Fay AP,
et al: Identification of ALK gene alterations in urothelial
carcinoma. PLoS One. 9(e103325)2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Scaravilli M, Asero P, Tammela TL,
Visakorpi T and Saramäki OR: Mapping of the chromosomal
amplification 1p21-22 in bladder cancer. BMC Res Notes.
7(547)2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Weilandt M, Koch A, Rieder H, Deenen R,
Schwender H, Niegisch G and Schulz WA: Target genes of recurrent
chromosomal amplification and deletion in urothelial carcinoma.
Cancer Genomics Proteomics. 11:141–153. 2014.PubMed/NCBI
|
|
12
|
Ewald JA, Downs TM, Cetnar JP and Ricke
WA: Expression microarray meta-analysis identifies genes associated
with Ras/MAPK and related pathways in progression of
muscle-invasive bladder transition cell carcinoma. PLoS One.
8(e55414)2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Hussain SA, Palmer DH, Syn WK, Sacco JJ,
Greensmith RM, Elmetwali T, Aachi V, Lloyd BH, Jithesh PV, Arrand
J, et al: Gene expression profiling in bladder cancer identifies
potential therapeutic targets. Int J Oncol. 50:1147–1159.
2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Mengual L, Burset M, Ars E, Lozano JJ,
Villavicencio H, Ribal MJ and Alcaraz A: DNA microarray expression
profiling of bladder cancer allows identification of noninvasive
diagnostic markers. J Urol. 182:741–748. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Sanchez-Carbayo M: Use of high-throughput
DNA microarrays to identify biomarkers for bladder cancer. Clin
Chem. 49:23–31. 2003.PubMed/NCBI View
Article : Google Scholar
|
|
16
|
de Ravel TJ, Devriendt K, Fryns JP and
Vermeesch JR: What's new in karyotyping? The move towards array
comparative genomic hybridisation (CGH). Eur J Pediatr.
166:637–643. 2007.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Letasiova S, Medve'ova A, Sovcikova A,
Dušinská M, Volkovová K, Mosoiu C and Bartonová A: Bladder cancer,
a review of the environmental risk factors. Environ Health. 11
Suppl 1(Suppl 1)(S11)2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Minner S, Kilgue A, Stahl P, Weikert S,
Rink M, Dahlem R, Fisch M, Höppner W, Wagner W, Bokemeyer C, et al:
Y chromosome loss is a frequent early event in urothelial bladder
cancer. Pathology. 42:356–359. 2010.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Panani AD and Roussos C: Sex chromosome
abnormalities in bladder cancer: Y polysomies are linked to
PT1-grade III transitional cell carcinoma. Anticancer Res.
26(1A):319–323. 2006.PubMed/NCBI
|
|
20
|
Forsberg LA: Loss of chromosome Y (LOY) in
blood cells is associated with increased risk for disease and
mortality in aging men. Hum Genet. 136:657–663. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Lopez V, Gonzalez-Peramato P, Suela J,
Serrano A, Algaba F, Cigudosa JC, Vidal A, Bellmunt J, Heredero O
and Sánchez-Carbayo M: Identification of prefoldin amplification
(1q23.3-q24.1) in bladder cancer using comparative genomic
hybridization (CGH) arrays of urinary DNA. J Transl Med.
11(182)2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huang Y, Li G, Wang K, Mu Z, Xie Q, Qu H,
Lv H and Hu B: Collagen Type VI Alpha 3 Chain promotes
epithelial-mesenchymal transition in bladder cancer cells via
transforming growth factor β (TGF-β)/Smad pathway. Med Sci Monit.
24:5346–5354. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Liang Y, Luo H, Zhang H, Dong Y and Bao Y:
Oncogene Delta/Notch-Like EGF-Related receptor promotes cell
proliferation, invasion, and migration in hepatocellular carcinoma
and predicts a poor prognosis. Cancer Biother Radiopharm.
33:380–386. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Deng S, He SY, Zhao P and Zhang P: The
role of oncostatin M receptor gene polymorphisms in bladder cancer.
World J Surg Oncol. 17(30)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Sun J, Zhang H, Tao D, Xie F, Liu F, Gu C,
Wang M, Wang L, Jiang G, Wang Z and Xiao X: CircCDYL inhibits the
expression of C-MYC to suppress cell growth and migration in
bladder cancer. Artif Cells Nanomed Biotechnol. 47:1349–1356.
2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen Y, Peng Y, Xu Z, Ge B, Xiang X, Zhang
T, Gao L, Shi H, Wang C and Huang J: Knockdown of lncRNA SNHG7
inhibited cell proliferation and migration in bladder cancer
through activating Wnt/β-catenin pathway. Pathol Res Pract.
215:302–307. 2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Daizumoto K, Yoshimaru T, Matsushita Y,
Fukawa T, Uehara H, Ono M, Komatsu M, Kanayama HO and Katagiri T: A
DDX31/Mutant-p53/EGFR Axis promotes multistep progression of
Muscle-Invasive bladder cancer. Cancer Res. 78:2233–2247.
2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wu L, Chaffee KG, Parker AS, Sicotte H and
Petersen GM: Zinc transporter genes and urological cancers:
Integrated analysis suggests a role for ZIP11 in bladder cancer.
Tumour Biol. 36:7431–7437. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Kim YW, Yoon HY, Seo SP, Lee SK, Kang HW,
Kim WT, Bang HJ, Ryu DH, Yun SJ, Lee SC, et al: Clinical
implications and prognostic values of prostate cancer
susceptibility candidate methylation in primary nonmuscle invasive
bladder cancer. Dis Markers. 2015(402963)2015.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen Z, Zhou L, Wang L, Kazobinka G, Zhang
X, Han X, Li B and Hou T: HBO1 promotes cell proliferation in
bladder cancer via activation of Wnt/β-catenin signaling. Mol
Carcinog. 57:12–21. 2018.PubMed/NCBI View Article : Google Scholar
|