Introduction
Diabetic cataract is a debilitating ophthalmic
disease and a major cause of visual impairment and blindness in
both developed and developing countries with increased incidence
and progression compared with other diabetes complications in
patients with diabetes mellitus (1-3).
In the UK, incidence rates of cataract were 20.4 (95% CI,
19.8-20.9) per 1,000 person-years (py) in patients with diabetes
and 10.8 (95% CI, 10.5-11.2) per 1,000 py in the general population
(1). Due to the increasing numbers
of cases of diabetes mellitus worldwide, the incidence of diabetic
cataract has steadily increased, especially in developing
countries, due to limited accesses to cataract surgery (3,4). A
recent epidemiological study reported that diabetic cataract was
also an early ocular complication in child and adolescent patients
with diabetes mellitus (5).
Although chataract surgery is widely applied for diabetic cataract
treatment, the pathogenic mechanisms underlying cataract
development in diabetes is still poorly understood, which makes it
challenging to prevent diabetic cataract at its initial stages
(4). In addition, a high
possibility of complications from cataract surgery has also been
observed in patients with diabetes mellitus, presenting a
challenging medical issue (4).
Thus, a complete understanding of the molecular pathogenic
mechanisms of diabetic cataract may facilitate the development of
novel modules for its prevention and treatment.
Previous studies have demonstrated that the
pathogenic development of diabetic cataract is associated with
abnormal metabolic regulation (4,6,7). For
instance, accumulation of sorbitol produced as a result of
enzymatic reduction of glucose by aldose reductase, has been
established as an initiating factor for diabetic cataract
pathogenesis (7,8). Specifically, the intracellular
deposition of the polyol sorbitol causes significant osmotic
changes in lens tissues, resulting in lens fiber degeneration and
liquefaction, eventually inducing sugar cataract formation, lens
swelling and opacities (4,9). In addition, the apoptosis of lens
epithelial cells (LECs) is also enhanced by the osmotic stress
induced by abnormal sorbitol accumulation in the lens, which is
also involved in diabetic cataract formation (10,11).
Normally, the mitochondrial oxygen consumption by human LECs
(HLECs) performs crucial roles in lens metabolism and maintenance
of a hypoxic environment around the lens, which is important for
lens transparency (12). Under
pathogenic conditions, high glucose effectively suppresses the
proliferation and induces apoptosis and oxidative stress of HLECs,
which is associated with lens opacities and cataract formation in
diabetes (12-14).
Therefore, the inhibition of LEC apoptosis and oxidative stress
under high glucose conditions is regarded as a potential strategy
for the prevention and treatment of diabetic cataract.
Autophagy refers to the cellular lysosomal
degradation processes responsible for the degradation and recycling
of abnormal proteins and damaged organelles, which is essential for
cell survival and development, and is associated with various human
diseases including diabetes mellitus (3,15). The
activation of the autophagy processes has been demonstrated to
potentially suppress cell apoptosis and oxidative stress under
specific conditions (16,17). For instance, the activation of the
autophagy process by eupatorine was demonstrated to suppress
arachidonic acid and iron-induced oxidative stress in human liver
cancer cells by alleviating mitochondrial dysfunction and
inhibiting reactive oxygen species (ROS) production (16). Notably, autophagy has also been
reported as a protective mechanism against oxidative stress in
pancreatic β cells and other insulin target tissues, including the
skeletal muscles, adipose tissue and liver (15). In another recent report, a novel
autophagy enhancer screened from a chemical library was effective
in suppressing the metabolic syndrome and diabetes mellitus in a
mouse model (18). Alterations of
the autophagy process in HLECs have also been implicated in
diabetic cataract progression (19); however, an effective regulator of
autophagy in HLECs associated with the pathogenesis of diabetic
cataract has not been established.
Resveratrol is a non-flavonoid polyphenol previously
isolated from the grape seed (Vitis vinifera L.), which
naturally occurrs as phytoalexin and is recommended for diabetic
patients as a dietary supplement to control blood sugar levels in
the body (20,21). It has been documented in the past 10
years that resveratrol is beneficial in the prevention and
treatment of various human disorders such as cancer,
cardiovascular, Alzheimer's, inflammatory bowel and fatty liver
diseases, as well as diabetes mellitus (20-22).
In addition, resveratrol can also regulate cellular oxidative
injury and autophagy processes under certain conditions; for
instance, resveratrol suppressed the oxidative injury in human
umbilical endothelial vein cells induced by oxidized low density
lipoprotein by activating the silencing information regulator 1
(SIRT1) and modulating the AMP-activated protein kinase α1
signaling pathway (23). In
addition, resveratrol has been recently demonstrated to induce
autophagy in non-small-cell lung cancer cells by activating the p38
mitogen-activated protein kinase (MAPK) and inhibiting the Akt/mTOR
signaling pathway (24). However,
the effects of resveratrol on the autophagy and oxidative damage in
HLECs associated with diabetic cataract pathogenesis are currently
unknown.
The present study aimed to verify the hypothesis
that resveratrol protects HLECs against high glucose-induced
oxidative injury by enhancing the cellular autophagy process.
Materials and methods
Cell culture
HLECs were purchased from Procell Life Science &
Technology Co., Ltd. (cat. no. CP-H127) and cultured in Dulbecco's
minimal essential medium (DMEM; cat. no. SH30022.01B; HyClone;
Cytiva) supplemented with 10% fetal bovine serum (FBS; cat. no.
10099-141; Gibco; Thermo Fisher Scientific, Inc.) and 5%
penicillin/streptomycin (Sigma-Aldrich; Merck KGaA) at 37˚C in a
humidified atmosphere with 5% CO2.
Cell treatment
To establish the cellular model of oxidative stress,
HLECs (1x105 cells/well) were plated into 6-well plates
and cultured at 37˚C for 8 h. The HLECs were then treated with 0,
5, 15, 20, 25 or 30 mM glucose (Sigma-Aldrich; Merck KGaA) for 24 h
under normal culture conditions. Then, the glucose-treated (25 mM)
HLECs were treated with 0, 1, 2 or 2.5 µM resveratrol (cat. no.
R5010; Sigma-Aldrich; Merck KGaA) for 24 h at 37˚C prior to
subsequent experiments.
For MAPK inhibition, glucose-treated (25 mM) and
resveratrol-treated (2 µM) HLECs were treated with 10 µM BIRB 796
(cat. no. S1574; Selleck Chemicals) for 24 h at 37˚C prior to
subsequent experiments.
Cell viability
The viability of cultured HLECs was detected using
the MTT Cell Proliferation and Cytotoxicity Assay kit (cat. no.
C0009; Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. Briefly, the cultured HLECs were
seeded into 96-well plates (2,000 cells/well) and cultured in DMEM
with 10% FBS containing the specified glucose concentrations for 24
h. Subsequently, cells in each well were incubated with 20 µl MTT
solution (cat. no. M-2128; Sigma-Aldrich; Merck KGaA) for 4 h under
normal culture conditions, followed by incubation with 100 µl DMSO
for 4 h under normal culture conditions until the formazan crystals
were completely dissolved. Finally, cell viability was determined
by measuring the absorbance at 570 nm using a plate reader. At
least three biological replicates were performed.
ROS content detection
The contents of ROS in the cultured HLECs were
measured by flow cytometry using the Cell-based ROS/Superoxide
Detection Assay kit (cat. no. ab139476; Abcam) according to the
manufacturer's instructions. Briefly, HLECs after specified
treatments were collected into 5 ml round-bottom polystyrene tubes,
washed with 1X Wash Buffer, resuspended in 500 µl ROS/Superoxide
Detection Solution and incubated for 30 min at 37˚C in the dark.
The ROS contents were determined using a flow cytometer (BD-C6,
CFLOW plus 1.0, Becton, Dickinson and Company) equipped with a blue
laser (488 nm). The ROS measurements were biologically repeated for
at least three times.
Western blot assay
For the detection of protein levels in cultured
HLECs, total proteins were extracted from cells after designated
treatments using a RIPA lysis buffer (cat. no. 89900; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
total protein contents were determined by a bicinchoninic acid
assay (cat. no. 23227; Thermo Fishier Scientific, Inc.).
Subsequently, 20 mg protein from each group was boiled at 100˚C for
5 min, separated by 10% SDS-PAGE and blotted onto PVDF membranes
(EMD Millipore). The membranes were blocked with a 5% fat-free milk
solution for 2 h at room temperature, incubated for 2 h at room
temperature with primary antibodies appropriately diluted in TBS-1%
Tween-20, and incubated with diluted secondary antibodies
(1:10,000) for 2 h at room temperature. The blots were visualized
by developing with the ECL Western Blotting Substrate (cat. no.
32106; Thermo Fishier Scientific, Inc.). GAPDH protein levels were
detected as the internal standard. The antibodies used were as
follows: Anti-LC3-I/II (cat. no. ABC929; Sigma-Aldrich; Merck
KGaA), anti-beclin-1 (cat. no. ab62557; Abcam), anti-P62 (cat. no.
ab56416; Abcam), anti-p38 MAPK (cat. no. 8690; Cell Signaling
Technology, Inc.), anti-phosphor-p38 MAPK (cat. no. 4511; Cell
Signaling Technology, Inc.), anti-GAPDH (ab9485; Abcam) and
HRP-conjugated goat anti-rabbit IgG (cat. no. ab205718; Abcam). At
least three biological replicates were performed for protein level
determination.
Immunofluorescence (IF) assay
The expression of LC3B protein in cultured HLECs was
analyzed by IF for the evaluation of autophagy following the
designated treatments. Briefly, cell slides were fixed with 4%
paraformaldehyde for 15 min at room temperature and washed three
times with PBS for 5 min, followed by blocking with 3% BSA (cat.
no. ST025; Beyotime Institute of Biotechnology) for 1 h at room
temperature, overnight incubation with primary antibodies against
LC3B (1:200; cat. no. 2775; Cell Signaling Technology, Inc.) at
4˚C, three washes with PBS for 5 min and incubation with
fluorophore-conjugated secondary antibodies (Alexa
Fluor® 488; 1:200; cat. no. ab150081; Abcam) for 2 h in
the dark. The slides were finally mounted with the
Prolong® Gold Antifade reagent (cat. no. 9071; Cell
Signaling Technology, Inc.) and observed under a Leica DMI6000B
fluorescence microscope (Leica Microsystems, Inc.). Three
biological replicates were performed for analysis of LC3B
expression by immunofluorescence.
Transmission electron microscopy
The formation of autophagosomes in HLECs was
observed by transmission electron microscopy (TEM) as previously
described (23). Briefly, HLECs
grown on slides were fixed for 1 h with 2.5% glutaraldehyde (cat.
no. G7776; Sigma-Aldrich; Merck KGaA) dissolved in 0.1 mol/l PBS at
room temperature, incubated with 1% osmium tetroxide (cat. no.
O5500; Sigma-Aldrich; Merck KGaA) for 30 min at room temperature
and observed using a TecnaiG2 Spirit Bio TWIN transmission electron
microscope (120 kV; FEI; Thermo Fisher Scientific, Inc.). The TEM
experiments were performed at least three times.
Statistical analysis
Data are presented as the mean ± SD of at least
three biological replicates. The differences among groups were
analyzed using the SPSS 20.0 software package (IBM Corp.) by
one-way ANOVA followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Resveratrol reverses the high
glucose-induced suppression of HLEC viability
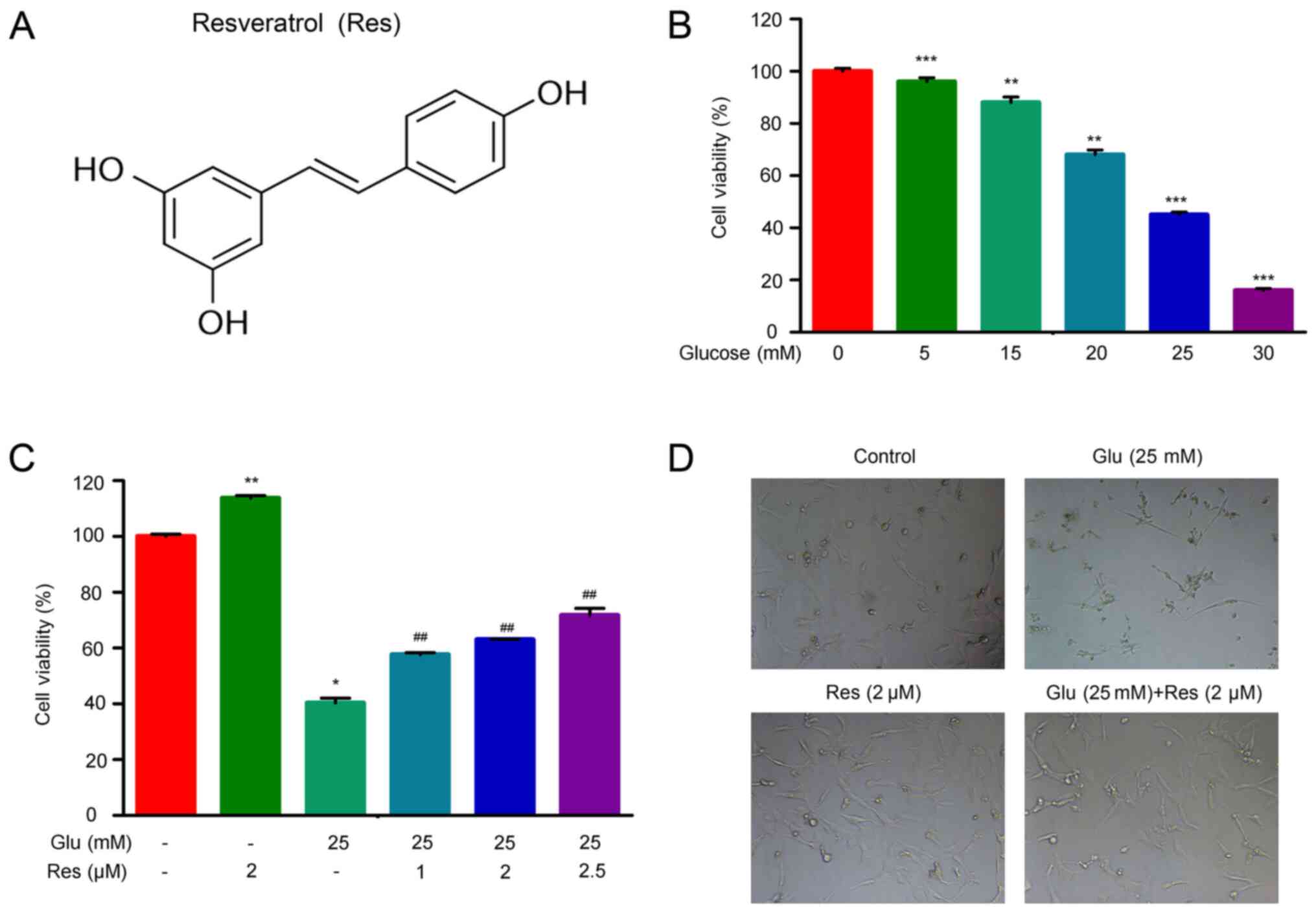
To determine the potential regulatory functions of
resveratrol (Fig. 1A) on HLEC
viability associated with diabetic cataract formation, the present
study established the cellular model of HLEC oxidative stress by
treating cultured HLECs with 0, 5, 15, 20, 25 or 30 mM glucose
(Fig. 1B). Treatment with glucose
concentrations ≥15 mM resulted in a significant
concentration-dependent decrease of HLEC cell viability (Fig. 1B). Subsequently, HLECs treated with
25 mM glucose were treated with 0, 1, 2 or 2.5 µM resveratrol for
24 h. The results demonstrated that the decreases in HLEC viability
induced by high glucose were reversed by resveratrol treatment in a
concentration-dependent manner (Fig.
1C). Treatment with 2.5 µM resveratrol for 24 h increased the
25 mM glucose-treated HLEC viability by ~60% compared with that of
HLECs treated with glucose alone (Fig.
1C). Microscopy revealed that 25 mM glucose-treated HLECs lost
their normal cell morphological features (the normal crystal
epithelial cells are evenly distributed, with equal cell size and
full shape and the cells are oval or polygonal), which were
recovered by simultaneous treatment with 2 µM resveratrol (Fig. 1D). These results demonstrated that
resveratrol treatment effectively reversed the suppression of HLEC
viability induced by high glucose.
Resveratrol mitigates the high
glucose-induced oxidative damage in HLECs
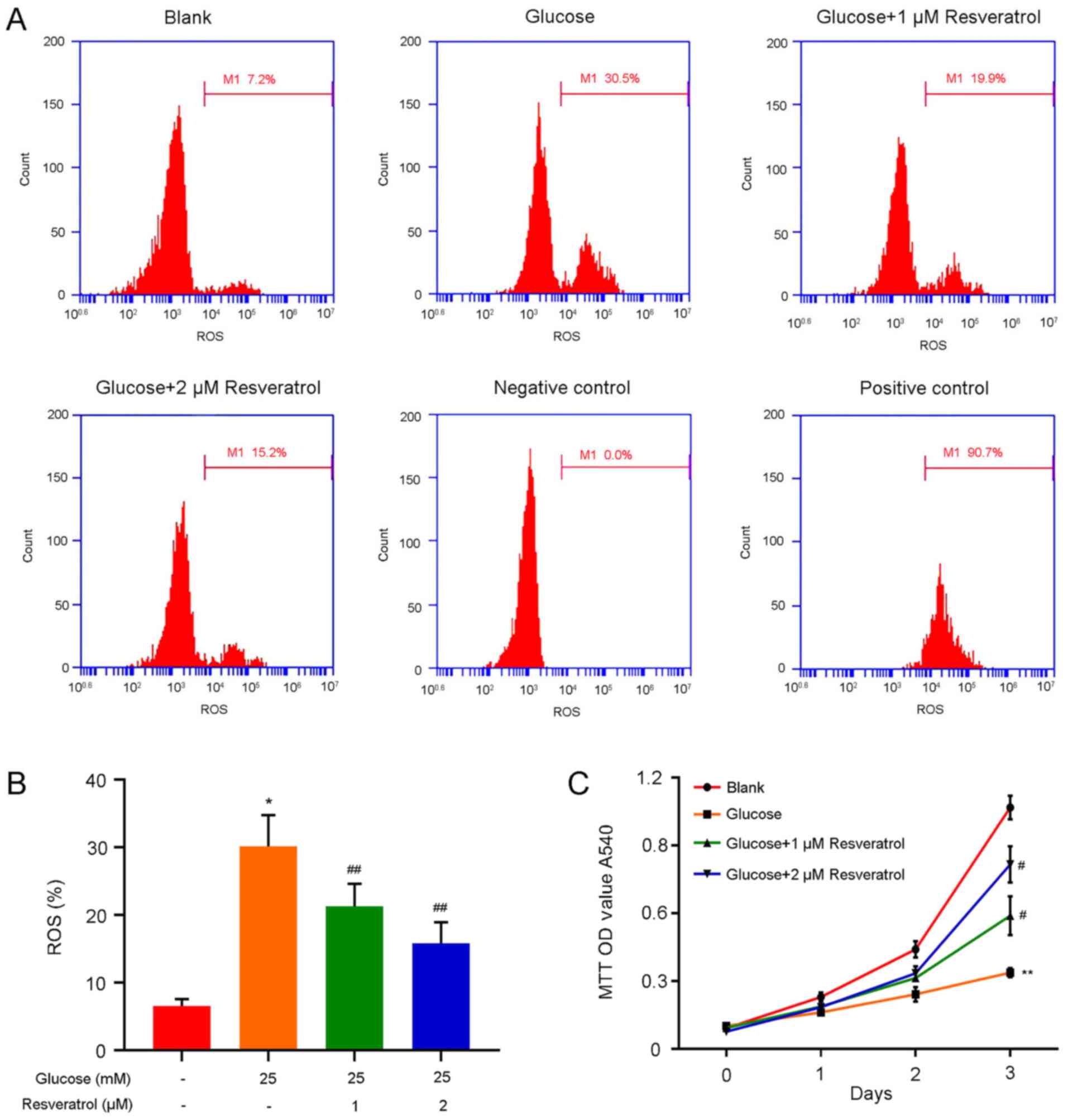
High glucose-induced oxidative injury in HLECs
mediates diabetic cataract pathogenesis (25). In the present study, the ROS
contents in HLECs under combined treatment with high glucose and
resveratrol were detected, and the results demonstrated that
treatment with 25 mM glucose induced a significant increase of ROS
contents in HLECs compared with those in the blank group (Fig. 2A). However, treatment with 1 or 2 µM
resveratrol reduced the ROS contents in HLECs under high glucose
treatment (Fig. 2A). In addition,
the proliferation of HLECs under above conditions was detected;
compared with that of untreated cells, the proliferation rate of
HLECs was repressed by treatment with 25 mM glucose, which was
recovered by treatment with 1 or 2 µM resveratrol (Fig. 2B). The alterations of the ROS
contents and proliferation rates of HLECs suggested that
resveratrol may act as an inhibitor of high glucose-induced
oxidative damage in HLECs.
Resveratrol promotes autophagy in
HLECs under high glucose
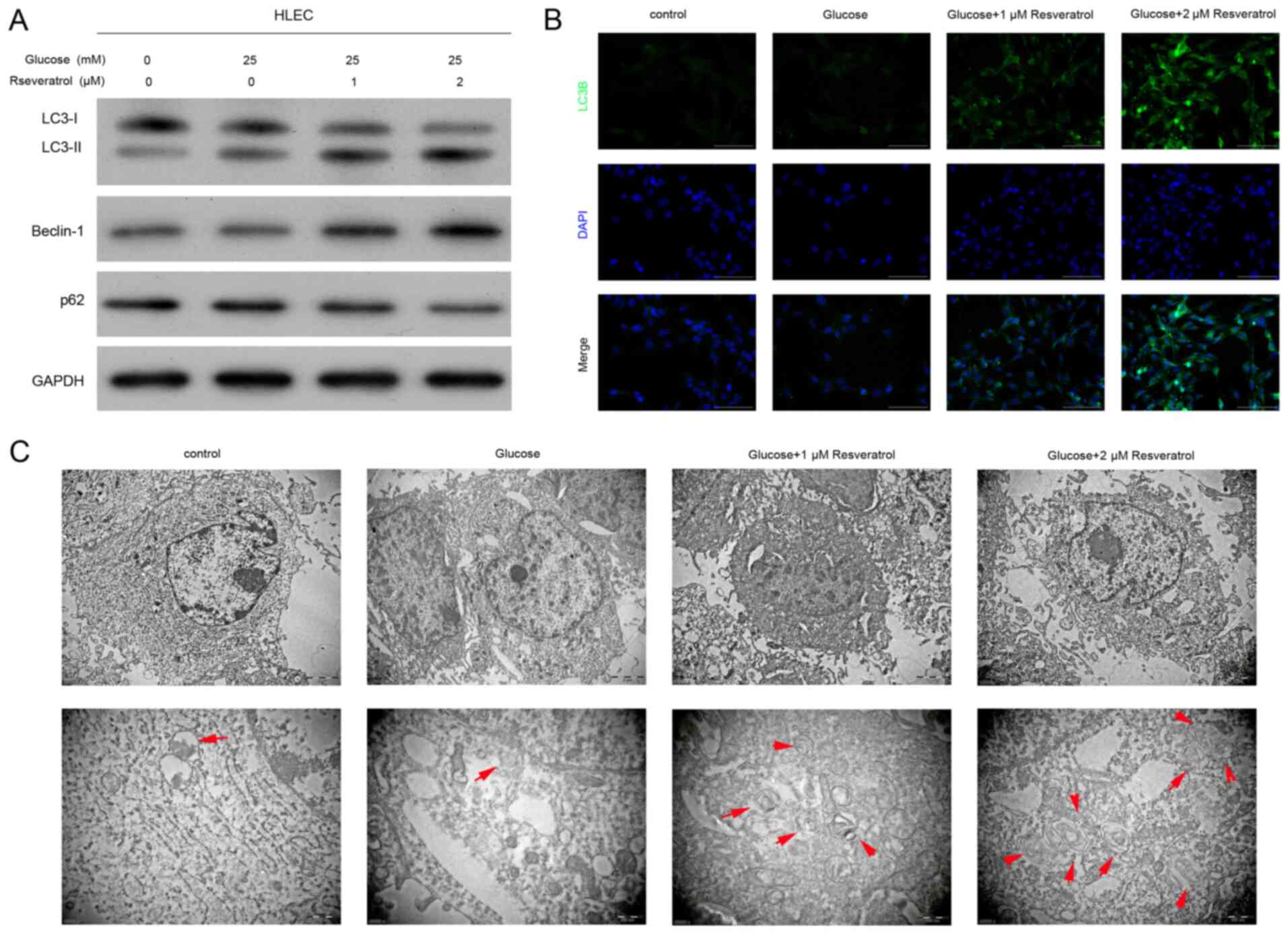
To explore the cellular processes underlying the
suppression of high glucose-induced HLEC oxidative damage by
resveratrol, the autophagy marker expression in HLECs treated with
high glucose and resveratrol was subsequently determined. The
protein expression of four autophagy markers was first detected by
western blotting, which demonstrated that LC3I and P62 protein
levels in cultured HLECs under high glucose treatment were reduced
by 1 or 2 µM resveratrol in a concentration-dependent manner
(Figs. 3A and S1). By contrast, the protein expression
levels of LC3II and beclin-1 in HLECs under high glucose conditions
were elevated by 1 or 2 µM resveratrol treatment (Fig. 3A). The IF assay demonstrated that
the levels of LC3B protein in HLECs under high glucose were
increased by treatment with 1 or 2 µM resveratrol compared with
those in untreated cells or cells treated with high glucose alone
(Fig. 3B). In addition, the
formation of autophagosomes in HLECs were analyzed by TEM. The
results demonstrated that a limited number of autophagosomes were
formed in untreated HLECs or those treated with high glucose alone;
however, a large number of autophagosomes was observed in HLEC
cells treated with high glucose and 1 or 2 µM resveratrol (Fig. 3C). These results suggested that
resveratrol treatment effectively activated autophagy in HLEC cells
under high glucose treatment.
Resveratrol activates the p38 MAPK
pathway to induce autophagy in HLECs under high glucose
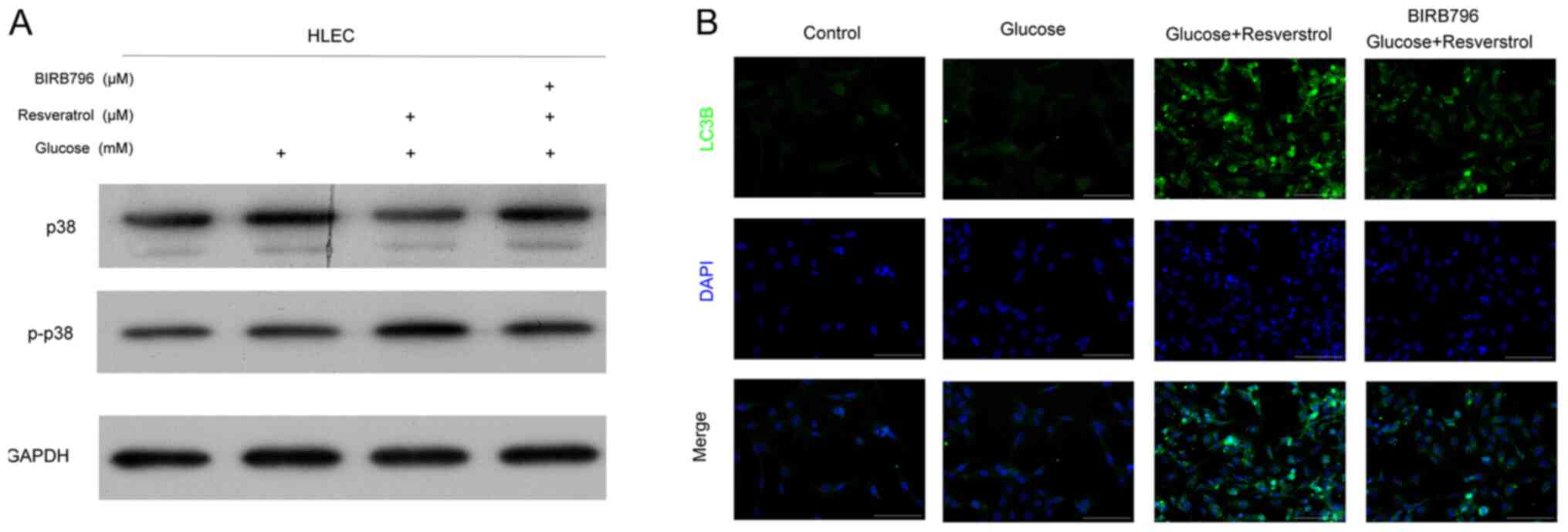
The p38 MAPK signaling pathway is involved in
autophagy development (26). The
results of the present study demonstrated that the total p38
protein levels in HLECs treated with high glucose and 2 µM
resveratrol were decreased compared with those in untreated cells
or cells treated with glucose alone (Fig. 4A). By contrast, the phosphorylated
p38 protein levels HLEC cells under high glucose treatment were
elevated by treatment with 2 µM resveratrol for 24 h (Fig. 4A). These alterations of p38
expression and phosphorylation induced by resveratrol were reversed
by the co-treatment with 10 µM MAPK inhibitor BIRB796 (Fig. 4A). Notably, IF results also
confirmed that the MAPK inhibitor BIRB796 suppressed the expression
of the autophagy marker LC3B in HLECs under high glucose and
resveratrol treatment (Fig. 4B).
These results demonstrated the role of resveratrol in promoting the
autophagy process in HLECs under high glucose conditions, which was
mediated by the activation of the p38 MAPK signaling pathway.
Discussion
Due to the crucial role of high glucose-induced
oxidative damage in the pathogenesis of diabetic cataract,
effective reagents capable of inhibiting HLEC oxidative damage have
been considered to be potential candidates for diabetic cataract
prevention and treatment (12-14).
Resveratrol, which is derived from grape seeds, has been
established as a potent antioxidant in various contexts, such as
inflammation and diabetes mellitus (20-22);
however, its effects on diabetic cataract remain unclear. In the
present study, it was hypothesized that resveratrol suppressed
diabetic cataract pathogenesis by regulating HLEC autophagy, which
was tested using high glucose-treated HLECs as an in vitro
model. The results of the present study demonstrated that
resveratrol effectively reversed the high glucose-induced
suppression of HLEC viability. In addition, the
high-glucose-induced oxidative stress and proliferation inhibition
in HLECs were significantly repressed by resveratrol. Using
multiple experimental assays, the present study demonstrated that
resveratrol treatment promoted autophagy in HLECs under high
glucose conditions, which was mediated by the resveratrol-induced
activation of the p38 MAPK signaling pathway. The molecular
mechanisms of the effects pf resveratrol in HLECs were finally
confirmed using the MAPK inhibitor BIRB796. Taken together, the
results of the present study characterized resveratrol as a new
drug candidate for diabetic cataract prevention and treatment,
which effectively suppressed high glucose-induced oxidative damage
by activating the p38 MAPK signaling to promote autophagy in
HLECs.
Natural chemical products have been established as a
valuable resource of novel therapeutic drugs for human disorders.
For instance, artemisinin, isolated from Artemisia annua L.,
has been explored as a life-saving anti-malaria drug (27) and has also been reported to possess
clinical values for other human pathogenic conditions such as
obesity and acute myeloid leukemia (28,29). A
number of such natural products have exhibited such multi-faceted
roles in human disease therapy, which has also been observed to
apply to resveratrol. Resveratrol was first identified as an
antioxidative reagent, which possessed additional biological
functions such as atherosclerosis prevention, hepatic
apolipoprotein and lipid synthesis modulation (28). In addition, resveratrol has been
reported to exert therapeutic effects on various human disorders
such as cancer, vascular disease, neurodegenerative and metabolic
diseases (30). The present study
demonstrated for the first time that resveratrol effectively
suppressed the inhibition of HLEC viability and oxidative stress
induced by high glucose treatment, indicating its application
potential as a new therapeutic drug for diabetic cataract
treatment. A previous study has reported that resveratrol represses
diabetes progression by modulating insulin action and pancreatic β
cell function (20). The results of
the present study further highlighted the value of resveratrol as
an inhibitor for diabetes mellitus and its complications.
The association of autophagy with human pathogenic
conditions including diabetes mellitus have been a research focus
during the past two decades, identifying autophagy as promising
target for drug development (31).
Autophagy in pancreatic β cells is also associated with diabetes
mellitus development and progression (32). For instance, the knockout of
autophagy-related protein (ATG) 7 gene specifically in β cells
causes reduced β-cell mass and decreased pancreatic insulin
synthesis and secretion, impaired response to obesity-induced ER
stress, as well as hypoinsulinemia and hyperglycemia symptoms in a
mouse model (33). However, the
association between autophagy in LECs and diabetic cataract
formation remains unclear. In the present study, resveratrol
promoted autophagy in HLECs under high glucose conditions, which
mediated its anti-diabetic cataract functions. LC3II protein is
produced by the binding of soluble LC3I with lipid components
during autophagosome formation (34). Beclin-1 is a component of the class
III phosphoinositide 3-kinase complex and performs key roles in the
induction of the autophagy process as a cytoprotective mechanism
(35). In addition, p62 protein has
been characterized as a ubiquitin- and LC3-binding protein that
undergoes selective degradation in response to autophagy (36). The results of the present study
demonstrated that the protein levels of LC3II and beclin-1 were
increased during resveratrol-induced autophagy in HLECs compared
with normal glucose levels, accompanied by a significant decrease
in the levels of LC3I and p62. These results suggested an
autophagy-promoting role for resveratrol in HLECs during diabetic
cataract development, and thus revealed HLEC autophagy as a
promising target for new anti-diabetic cataract drug screening.
The signaling pathway responsible for modulating
autophagy under high glucose conditions in HLECs was also explored
in the present study. A previous study has demonstrated that
autophagy formation is regulated by the p38 MAPK signaling pathway,
which is activated by phosphorylation and binds with the
p38-interacting protein to modulate ATG9 protein activity during
autophagosome formation (37). In
the present study, the phosphorylation of p38 protein was promoted
by resveratrol in HLECs under high glucose conditions, which
indicated that resveratrol-induced HLEC autophagy was mediated by
the p38 MAPK and its associated signaling pathways. Notably, a
previous study has also demonstrated that a number of biological
roles of resveratrol were mediated by the activation of the
NAD+-dependent deacetylase SIRT1(38). In addition, a recent study has
reported that SIRT1 was associated with LEC proliferation and
apoptosis in a mouse model of diabetic cataract (39). The involvement of SIRT1 and other
signaling pathways in the HLEC oxidative damage-inhibiting roles of
resveratrol require further investigation.
In summary, the results of the present study
demonstrated that the plant-derived chemical resveratrol inhibited
high glucose-induced oxidative damage in HLECs, which was mediated
by the activation of autophagy through modulating the p38 MAPK
signaling pathway. These results suggested that resveratrol may be
a promising candidate anti-diabetic cataract drug and provided a
basis for its application in diabetic cataract prevention and
treatment.
Supplementary Material
Quantitative analysis of p62 and LC3I
of western blotting. **P<0.01 and
***P<0.001. LC3, microtubule-associated protein 1
light chain 3.
Acknowledgements
Not applicable.
Funding
This study was supported by The Inner Mongolia Autonomous Region
Natural Science Fund Project (grant no. 2018LH08064).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PC designed the study. PC, ZY and ZH performed the
experiments. ZY and ZH performed data analysis and interpretation.
PC wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Becker C, Schneider C, Aballéa S, Bailey
C, Bourne R, Jick S and Meier C: Cataract in patients with diabetes
mellitus-incidence rates in the UK and risk factors. Eye (Lond).
32:1028–1035. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wong IYH, Ni MY, Wong IOL, Fong N and
Leung GM: Saving sight in China and beyond: The lifeline express
model. BMJ Glob Health. 3(e000766)2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ferraiolo A, Bordone C, Ramone A, Gorlero
F, Gustavino C and Cordera R: Universal versus risk factor
screening for gestational diabetes mellitus. Clin Exp Obstet
Gynecol. 45:53–57. 2018.
|
|
4
|
Pollreisz A and Schmidt-Erfurth U:
Diabetic cataract- pathogenesis, epidemiology and treatment. J
Ophthalmol. 2010(608751)2010.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Šimunović M, Paradžik M, Škrabić R, Unić
I, Bućan K and Škrabić V: Cataract as early ocular complication in
children and adolescents with type 1 diabetes mellitus. Int J
Endocrinol. 2018(6763586)2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Obrosova IG, Chung SS and Kador PF:
Diabetic cataracts: Mechanisms and management. Diabetes Metab Res
Rev. 26:172–180. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Snow A, Shieh B, Chang KC, Pal A, Lenhart
P, Ammar D, Ruzycki P, Palla S, Reddy GB and Petrash JM: Aldose
reductase expression as a risk factor for cataract. Chem Biol
Interact. 234:247–253. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bhadada SV, Vyas VK and Goyal RK:
Protective effect of Tephrosia purpurea in diabetic cataract
through aldose reductase inhibitory activity. Biomed Pharmacother.
83:221–228. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Kim J, Kim CS, Sohn E, Lee YM, Jo K and
Kim JS: Litsea japonica extract inhibits aldose reductase activity
and hyperglycemia-induced Lenticular Sorbitol accumulation in db/db
mice. Evid Based Complement Alternat Med.
2015(747830)2015.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kim J, Kim CS, Sohn E, Kim H, Jeong IH and
Jin SK: Lens epithelial cell apoptosis initiates diabetic
cataractogenesis in the Zucker diabetic fatty rat. Graefes Arch
Clin Exp Ophthalmol. 248:811–818. 2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhu L, Zhao K and Lou D: Apoptosis factors
of lens epithelial cells Responsible for cataractogenesis in
vitrectomized eyes with silicone oil tamponade. Med Sci Monit.
22:788–796. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kubota M, Shui YB, Liu M, Bai F, Huang AJ,
Ma N, Beebe DC and Siegfried CJ: Mitochondrial oxygen metabolism in
primary human lens epithelial cells: Association with age, diabetes
and glaucoma. Free Radic Biol Med. 97:513–519. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gong W, Zhu G, Li J and Yang X: LncRNA
MALAT1 promotes the apoptosis and oxidative stress of human lens
epithelial cells via p38MAPK pathway in diabetic cataract. Diabetes
Res Clin Pract. 144:314–321. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Nambu H, Kubo E, Takamura Y, Tsuzuki S,
Tamura M and Akagi Y: Attenuation of aldose reductase gene
suppresses high-glucose-induced apoptosis and oxidative stress in
rat lens epithelial cells. Diabetes Res Clin Pract. 82:18–24.
2008.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Barlow AD and Thomas DC: Autophagy in
diabetes: β-Cell dysfunction, insulin resistance, and
complications. DNA Cell Bio. 34:252–260. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Jegal KH, Ko HL, Sang MP, Byun SH, Kang
KW, Cho IJ and Kim SC: Eupatilin induces Sestrin2-dependent
autophagy to prevent oxidative stress. Apoptosis. 21:642–656.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Xie W, Zhang L, Jiao H, Guan L, Zha J, Li
X, Wu M, Wang Z, Han J and You H: Chaperone-mediated autophagy
prevents apoptosis by degrading BBC3/PUMA. Autophagy. 11:1623–1635.
2015.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Lim H, Lim YM, Kim KH, Jeon YE, Park K,
Kim J, Hwang HY, Lee DJ, Pagire H, Kwon HJ, et al: A novel
autophagy enhancer as a therapeutic agent against metabolic
syndrome and diabetes. Nat Commun. 9(1438)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lu Z, Rong C and Huang Y: MiR-30a inhibits
BECN1-mediated autophagy in diabetic cataract. Oncotarget.
8:77360–77368. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Öztürk E, Arslan AKK, Yerer MB and
Bishayee A: Resveratrol and diabetes: A critical review of clinical
studies. Biomed Pharmacother. 95:230–234. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rodrigo R, Miranda A and Vergara L:
Modulation of endogenous antioxidant system by wine polyphenols in
human disease. Clin Chim Acta. 412:410–424. 2011.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Vinod BS, Nair HH, Vijayakurup V, Shabna
A, Shah S, Krishna A, Pillai KS, Thankachan S and Anto RJ:
Resveratrol chemosensitizes HER-2-overexpressing breast cancer
cells to docetaxel chemoresistance by inhibiting docetaxel-mediated
activation of HER-2-Akt axis. Cell Death Discov.
1(15061)2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Guo H, Chen Y, Liao L and Wu W:
Resveratrol protects HUVECs from oxidized-LDL induced oxidative
damage by autophagy upregulation via the AMPK/SIRT1 pathway.
Cardiovasc Drugs Ther. 27:189–198. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang J, Li J, Cao N, Li Z, Han J and Li L:
Resveratrol, an activator of SIRT1, induces protective autophagy in
non-small-cell lung cancer via inhibiting Akt/mTOR and activating
p38-MAPK. OncoTargets Ther. 11:7777–7786. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wu Z, Yin X, Ji L, Gao YY, Pan YM, Lu Q
and Wang JY: Ginkgo biloba extract prevents against apoptosis
induced by high glucose in human lens epithelial cells. Acta
Pharmacol Sin. 29:1042–1050. 2008.PubMed/NCBI View Article : Google Scholar
|
|
26
|
He Y, She H, Zhang T, Xu H, Cheng L, Yepes
M, Zhao Y and Mao Z: p38 MAPK inhibits autophagy and promotes
microglial inflammatory responses by phosphorylating ULK1. J Cell
Biol. 217:315–328. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Fairhurst RM, Nayyar GM, Breman JG,
Hallett R, Vennerstrom JL, Duong S, Ringwald P, Wellems TE, Plowe
CV and Dondorp AM: Artemisinin-resistant malaria: Research
challenges, opportunities, and public health implications. Am J
Trop Med Hyg. 87:231–241. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Drenberg CD, Buaboonnam J, Orwick SJ, Hu
S, Li L, Fan Y, Shelat AA, Guy RK, Rubnitz J and Baker SD:
Evaluation of artemisinins for the treatment of acute myeloid
leukemia. Cancer Chemother Pharmacol. 77:1231–1243. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ping L, Zhang F, Qian S, Li X, Cui ZM,
Dang YJ and Tang QQ: Artemisinin derivatives prevent obesity by
inducing browning of WAT and enhancing BAT function. Cell Res.
26:1169–1172. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Kuršvietienė L, Stanevičienė I,
Mongirdienė A and Bernatonienė J: Multiplicity of effects and
health benefits of resveratrol. Medicina (Kaunas). 52:148–155.
2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gao L, Jauregui CE and Teng Y: Targeting
autophagy as a strategy for drug discovery and therapeutic
modulation. Future Med Chem. 9:335–345. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Marasco MR and Linnemann AK: β-cell
autophagy in diabetes pathogenesis. Endocrinology. 159:2127–2141.
2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Quan W, Lim YM and Lee MS: Role of
autophagy in diabetes and endoplasmic reticulum stress of
pancreatic β-cells. Exp Mol Med. 44:81–88. 2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mcleland CB, Rodriguez J and Stern ST:
Autophagy monitoring assay: Qualitative analysis of MAP LC3-I to II
conversion by immunoblot. Methods Mol Biol. 697(199)2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Pattingre S, Tassa A, Qu X, Garuti R,
Liang XH, Mizushima N, Packer M, Schneider MD and Levine B: Bcl-2
antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell.
122:927–939. 2005.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Komatsu M and Ichimura Y: Physiological
significance of selective degradation of p62 by autophagy. FEBS
Lett. 584:1374–1378. 2010.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Guo F, He XB, Li S and Le W: A central
role for phosphorylated p38α in linking proteasome
inhibition-induced apoptosis and autophagy. Mol Neurobiol. 54:1–13.
2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chung S, Yao H, Caito S, Hwang JW,
Arunachalam G and Rahman I: Regulation of SIRT1 in cellular
functions: Role of polyphenols. Arch Biochem Biophys. 501:79–90.
2010.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zeng K, Feng QG, Lin BT, Ma DH and Liu CM:
Effects of microRNA-211 on proliferation and apoptosis of lens
epithelial cells by targeting SIRT1 gene in diabetic cataract mice.
Bioscience Reports. 37(BSR20170695)2017.PubMed/NCBI View Article : Google Scholar
|