Introduction
Ulcerative colitis (UC) is a chronic inflammatory
bowel disease that occurs in the mucosa and submucosa of the large
intestine. The primary clinical manifestations are bloody diarrhea,
mucopurulent bloody stool and abdominal pain, fecal urgency, and
tenesmus, and in severe cases, weight loss and fever (1-3).
The overall incidence and prevalence of UC are 1.2/20.3 and 7.6/245
cases per 100,000 individuals per year, respectively (4,5).
Epidemiological studies have shown a bimodal distribution of age at
UC onset, with a peak incidence between 20-30 years, followed by
another peak at 50-80 years (6).
The pathogenesis of UC is complex, and environmental factors,
genetic susceptibility, intestinal microbiology, and a dysregulated
immune response are major risk factors (7,8). In
recent years, 5-aminosalicylic acid, corticosteroid drugs, Janus
kinase inhibitors, and anti-TNF-α antibodies have been widely
applied for the treatment of UC (9).
At present, the incidence of UC is high, and the
mortality rate is low. UC not only is incurable, but it is also a
challenge to manage. Patients suffer from intractable or
intolerable side effects of UC therapeutics following
administration, which leads to a high recurrence rate, and UC is
closely associated with colorectal cancer (CRC) (10-12).
The high incidence of UC in developed and developing countries has
evolved into a global burden, and several treatments for UC also
induce resistance to said treatments in patients. Thus, there is an
urgent need to identify novel drugs that will be effective in
achieving optimal disease control (13).
At present, several traditional Chinese formulas,
such as Shen-Ling-Bai-Zhu-San (14), Tongxie Yaofang (15), Pulsatilla decoction
(16), Huangkui Lianchang
decoction (17), and Gegen
Qinlian decoction (18),
amongst others, are used for the treatment of UC in China (19). Tongxie Yaofang consists of
four drugs Rhizoma Atractylodis Macrocephalae, Radix
Paeoniae Alba, Pericarpium Citri Reticulatae, and
Radix Saposhnikovia Divaricata (20) and exhibited efficacy for the
treatment of UC in our previously reported work (21). Paeoniflorin (PF) is the primary
bioactive component isolated from Radix Paeoniae Alba, and
Paeonia suffruticosa Andr. has been used in several other
traditional Chinese formulas for the treatment of inflammatory
bowel diseases, UC, and other gastric and splenic diseases for
>2,000 years (22,23).
Several studies have revealed that PF exhibits
anti-tumor, anti-inflammatory, anti-oxidative, and immunoregulatory
effects (24-26).
It has been shown that PF reduces TNF-α, IL-6, and IL-1β levels to
protect against inflammatory pain (27). However, the molecular mechanisms
underlying the protective effects of PF against dextran sulfate
sodium (DSS)-induced UC remain unclear.
Based on the advances in the fields of molecular
biology, systems biology, polypharmacology, and bioinformatics,
network pharmacology has been developed as a means to reveal the
mechanisms of drugs and is considered a cost-effective method of
drug development (28). In recent
years, network pharmacology has been applied to determine the
actions of active compounds and to analyze the mechanisms of action
of natural products in the treatment of several diseases (29).
To further analyze the therapeutic mechanism of
action of PF in UC, network pharmacology was used to predict the
targets and pathways by which PF exerted its effects in UC. Then,
the relevant pathways were selected for validation in vivo
to confirm the therapeutic mechanisms of PF in animal models.
Materials and methods
Screening of PF and UC prediction
targets
The Structure Data File (SDF) of PF (C23H28O11) was
downloaded from the PubChem database (PubChem CID: 442534;
https://pubchem.ncbi.nlm.nih.gov/) and
then imported into the PharmMapper database (http://www.lilab-ecust.cn/pharmmapper/) to obtain
prediction targets with the conformation generation was set to
‘generate conformers’ and the pharmacophore mapping was set to
‘druggable pharmacophore models (v2017, 16159)’. UC-related human
genes were collected from three databases; namely, the Therapeutic
Target Database (TTD) (http://bidd.nus.edu.sg/group/cjttd/), Online Mendelian
Inheritance in Man (OMIM) (https://www.omim.org/), and GeneCards (https://www.genecards.org/). These disease-target
network databases were used to verify whether the genes were
related to UC. The official gene symbols of the targets were
normalized in the UniProt (http://www.UniProt.org) database. The common terms
amongst the disease-related targets and drug component targets were
obtained using Venny 2.1.0 (http://bioinfo.cnb.csic.es/tools/venny/index.html).
Protein-protein interaction (PPI)
analyses
A PPI network was constructed using the Search Tool
for the Retrieval of Interacting Genes (STRING) database
(http://string-db.org/; version 10.5). The
selection criteria were ‘Homo sapiens’ for species and a
confidence score >0.4. Then, the PPI network was imported into
Cytoscape (30), and the plugin
cytoHubba was used to find the clusters (highly interconnected
regions) within the PPI network (31). The top-ranked genes were selected
as hub targets based on the degree levels. Four topological
features (degree, betweenness centrality, average shortest path
length and closeness centrality) were analyzed using Network
Analyzer (32). The major hub
network comprising putative major components and major targets was
extracted by defining nodes with degrees higher than the average
number of neighbors.
Gene ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) enrichment analyses
After obtaining the core target, clusterProfiler
(33) in R (34,35)
version 3.6.1 was used to perform the GO (36,37)
and KEGG (38) enrichment analyses
in the RStudio platform 2022.12.0-353. GO enrichment primarily
analyzes the biological processes (BPs), cellular composition (CC),
and molecular functions (MFs) of the common targets. KEGG pathway
enrichment (www.kegg.jp/kegg/kegg1.html) was used to identify the
critical pathways that were closely related to the treatment of UC
by PF.
Molecular docking simulation
The binding of the potential target and its
corresponding components were evaluated by molecular docking
simulation. Molecular docking simulations of potential targets and
their corresponding components were performed using AutoDock
version 4.2(39) and AutoDock Vina
software (Scripps Research Institute) (40) according to published methods. The
macromolecular protein target receptors were obtained from the
Research Collaboratory for Structural Bioinformatics Protein Data
Bank database (https://www.rcsb.org), and the 2D
structures of the small-molecule compound components were obtained
from the PubChem Database (https://pubchem.ncbi.nlm.nih.gov).
Chemicals and reagents
PF (CAS:23180-57-6) was obtained from Chengdu
Herbpuritfy Co., Ltd. and its identity was confirmed by
high-performance liquid chromatography with a purity of 99.71%.
ELISA kits for IL-6 (cat. no. EK206), IL-10 (cat. no. EK210), TNF-a
(cat. no. EK282), TGF-β (cat. no. EK981), and IL-1β (cat. no.
EK201B) were purchased from Multisciences (Lianke) Biotech, Co.,
Ltd. The primers for reverse transcription-quantitative (RT-q) PCR
and the Promega M-MLV vector were purchased from Shanghai GeneChem
Co., Ltd. All antibodies were purchased from Affinity Biosciences
Co., Ltd.
Establishment of the UC mouse model
and dosing
A total of 27 specific pathogen-free male Balb/c
mice (6 weeks old, from Jinan Peng Yue Experimental Animal
Technology Co., Ltd.), weighing 18±2 g, were kept in the Animal
Laboratory of Jining Medical University. The animal experiments
were approved by the Animal Ethics Committee of Jining Medical
University (approval no. JNMC-2020-DW-ZXY-001). If weight loss
reached 15-20% of the original weight within 72 h, then the mice
were sacrificed by cervical dislocation. After 3 days of adaptive
feeding, mice were randomly divided into the following three groups
(n=9 per group): Control group, model group, and treatment group.
For DSS-induced colitis, 5% (w/v) DSS (Meilun Biotech Co., Ltd.)
was added to the drinking water in the model group and treatment
group for 7 days. After the establishment of the DSS-colitis model,
mice in the control and model groups were administered normal
saline, while mice in the treatment group were administered PF (20
mg/kg/day). All administration methods were by gavage, and the mice
were dosed continuously for 7 days. Mice from each group were
sacrificed by cervical dislocation after 14 days.
Disease activity index (DAI)
score
The DAI score in each group was taken during the
experimental period. The DAI score was calculated according to loss
of body weight, stool consistency and gross bleeding, as follows:
DAI score=(weight loss score + stool shape score + stool blood
score)/3 (41,42).
Hematoxylin and eosin staining
Colonic tissue was obtained from the mice and fixed
in 10% formalin for 24 h at room temperature and then dehydrated
using an increasing alcohol gradient, embedded in paraffin and
sectioned in 5-µm thick slices. Then, the sections were stained
with hematoxylin and eosin (H&E) for histopathological
examination.
ELISA
ELISA kits were used to detect the serum levels of
IL-10, TGF-β, IL-6, IL-1β, and TNF-α according to the
manufacturers' instructions. The optical density value was measured
at 450 nm using a microplate reader (Model 680; Bio-Rad
Laboratories, Inc.), and the concentration was calculated using a
standard curve.
RT-qPCR
Total RNA was isolated from the tissues using
TRIzol® Reagent (Shanghai Pufei Biotechnology Co., Ltd.)
according to the manufacturer's protocol. For RT-qPCR analysis,
first-strand cDNA synthesis from 1 µg total RNA was performed using
an M-MLV Reverse Transcriptase kit according to the manufacturer's
protocol (Promega Corporation). The resulting cDNA was analyzed by
qPCR using the SYBR Green PCR kit (Takara Bio, Inc.) and the 7500
Fast Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). β-actin was used as the loading control.
Comparative quantification was performed using the
2-ΔΔCq method (43). The primers sequences are listed in
Table I.
 | Table ISequences of the primers. |
Table I
Sequences of the primers.
| Gene | Sequence,
5'-3' |
|---|
| PI3K | |
|
Forward |
TCATTGACAGTAGGAGGAGGTT |
|
Reverse |
TATTTTCATTCCCCAGCCAC |
| AKT1 | |
|
Forward |
TGGACTCAAGAGGCAGGAAG |
|
Reverse |
GGTTCTCAGTAAGCGTGTGG |
| mTOR | |
|
Forward |
AGGTCATGCCCACATTCCTT |
|
Reverse |
AGCCACCACAATCTGCTCAA |
| BCL2 F | |
|
Forward |
ATGCCTTTGTGGAACTATATGGC |
|
Reverse |
GGTATGCACCCAGAGTGATGC |
| EGFR | |
|
Forward |
AACTCTTCGGGACACCCAATCA |
|
Reverse |
CAGCCTTCCGAGGAGCATAAA |
| Actin | |
|
Forward |
GCACCACACCTTCTACAATG |
|
Reverse |
GTGAGGGAGAGCATAGCC |
Western blot analysis
Total proteins were obtained from colon tissues
using RIPA lysate and then quantified using the BCA method. Equal
amounts of protein (50 µg/lane) were separated by 10% SDS-PAGE gel
using 100 V for 2 h and then subsequently transferred onto PVDF
membranes. The membranes were blocked with 5% non-fat milk for 2 h
at room temperature and washed and then incubated with primary
antibodies (1:500 dilution) against EGFR (cat. no. AF6043), p-PI3K
(cat. no. AF3242), p-mTOR (cat. no. AF3308), Bcl-2 (cat. no.
AF6139), p-AKT1 (cat. no. AF0016), PI3K (cat. no. AF6241), mTOR
(cat. no. AF6308), and AKT1 (cat. no. AF0836) overnight at 4˚C.
Then, the membranes were incubated with horseradish
peroxidase-labeled secondary antibody (cat. no. S0002, 1:5,000
dilution) for 2 h at room temperature after washing with
Tris-buffered saline containing Tween-20 (0.1%) three times.
Signals were visualized using enhanced chemiluminescence reagent
and analyzed using ImageJ (1.53 h; National Institutes of
Health).
Statistical analysis
Data are presented as the mean ± SD. An unpaired
Student's t-test was used to assess differences between the two
groups. A one-way ANOVA followed by a Dunnett's post hoc test was
used for comparisons between multiple groups. GraphPad Prism
version 8.0.0 (GraphPad Software, Inc.) was used for statistical
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
Screening of PF and UC prediction
targets
The chemical structure of PF was obtained from the
PubChem database. Then the SDF file of PF was imported into the
PharmMapper server (http://www.lilab-ecust.cn/pharmmapper/index.html).
A total of 288 potable targets were obtained in PharmMapper with
NormFit scores >0. Additionally, a total of 599 potential gene
targets were obtained from the OMIM, TTD, and GeneCards databases.
Common targets identified from both the active ingredient and
UC-related searches were considered potential targets. A total of
60 common potential genes were screened by Venny (Fig. 1).
Construction of the PPI network
The 60 common targets were submitted to STRING
(https://string-db.org/) to construct a PPI
network. Then, the PPI network was constructed and visualized using
Cytoscape. The network contained 60 nodes and 481 edges, and the
average node degree was 16 (Fig.
2A). Two key topological parameters (degree and betweenness
centrality) were used to characterize the most important nodes in
the network; higher quantitative values of the topological
parameters indicated greater importance of the node. A Cytoscape
plug-in, Network Analyzer, was used to identify the network
topology attributes to identify the core genes of the PPI network
(32). Based on this principle,
the top 20 nodes were screened out as key components, and the top
10 included the following: ALB (albumin, degree=51), AKT1 (AKT
serine/threonine kinase 1, degree=41), MMP9 (matrix
metallopeptidase 9, degree=40), CASP3 (degree=36), SRC (degree=36),
EGFR (degree=35), IGF1 (degree=33), MMP2 (degree=29), ESR1
(estrogen receptor 1, degree=28), IL-2 (degree=28) and MAPK14
(mitogen-activated protein kinase 14, degree=25) (Fig. 2B).
GO and KEGG enrichment analysis
GO and KEGG pathway enrichment analyses were
performed using the Metascape database and R for visualization of
the 60 common target genes. In terms of MFs, treatment of UC with
PF primarily involved endopeptidase activity, serine-type peptidase
activity, serine hydrolase activity, nuclear receptor activity,
transcription factor activity, and direct ligand-regulated
sequence-specific DNA binding (Fig.
3A). The BPs primarily involved regulation of the inflammatory
response, neutrophil-mediated immunity, response to reactive oxygen
species, response to lipopolysaccharide, and response to molecules
of bacterial origin (Fig. 3B). The
CC primarily involved vesicle lumen, secretory granule lumen,
cytoplasmic vesicle lumen, collagen-containing extracellular
matrix, and membrane raft (Fig.
3C).
The top 20 functionally enriched processes were
selected to plot a bubble diagram following KEGG enrichment
analysis (Fig. 3D). The primary
signaling pathways involved in the treatment of UC were identified,
and the top five pathways included proteoglycans in cancer, EGFR
tyrosine kinase inhibitor resistance, endocrine resistance,
prostate cancer, fluid shear stress, and atherosclerosis. The
IL-17, PI3K/AKT, and HIF-1 signaling pathways were also included in
the top 20 pathways. Among them, 12, 7, and 7 genes were enriched
in the PI3K/AKT (AKT1, BCL2L1, EGFR, FGFR2, IGF1, IGF1R, IL2, JAK2,
KDR, MET, NOS3, and SYK), IL-17 (CASP3, LCN2, MAPK14, MMP13, MMP3,
MMP9, S100A9), and HIF-1 (AKT1, EGFR, HMOX1, IGF1, IGF1R, NOS2,
NOS3) signaling pathways, respectively.
Molecular docking simulation
analysis
Docking analysis was conducted between PF and the
core genes related to the PI3K/AKT signaling pathway (Fig. 4). According to the molecular
docking results, docking of PF with AKT1 and EGFR proteins had a
good binding energy of -2.44 kcal/mole and -2.19 kcal/mole
respectively, and AKT1-PF had the highest binding affinity.
PF reduces DSS-induced experimental
colitis
The control group showed a steady weight increase.
After administration of DSS, a significant loss in the body weight
of the animal was observed, and this continued until the end of the
experiment in the PF group and model group compared with the
control group (P<0.01, Fig.
5A). Compared with the model group, the PF-treated mice showed
a significant decrease in DAI scores (P<0.01, Fig. 5B). The colonic tissue was observed
by light microscopy. The histomorphology of the colonic tissue of
mice in the control group appeared normal. In the model group, the
colonic tissue showed colonic mucosal injury with a decrease in the
number of glands and substantial inflammatory cell infiltration in
the mucosa and submucosa. In the PF treatment group, these
abnormalities were alleviated, the colonic mucosa was restored to
the levels of the control group, and inflammatory cell infiltration
and congestion were also reduced in the PF group (Fig. 5C-E).
PF treatment reduces inflammatory
cytokine release
The expression levels of IL-10 and TGF-β in the
model group were significantly decreased (P<0.01), while the
expression levels of IL-6, TNF-α, and IL-1β were significantly
increased compared with those in the control group (P<0.01). The
expression levels of IL-10 and TGF-β in the PF group were
significantly increased compared with those in the model group
(P<0.01). The expression levels of IL-6, TNF-α, and IL-1β were
significantly decreased in the PF group compared with the model
group (P<0.01, Fig. 6).
 | Figure 6Expression levels of (A) IL-10, (B)
IL-6, (C) TNF-α, (D) TGF-β, and (E) IL-1β in the serum. The
expression levels of IL-10 and TGF-β in the model group were
decreased, while the expression levels of IL-6, TNF-α, and IL-1β
were increased when compared with those in the control group
(F=37.470, 23.370, 58.343, 19.526, 11.615). The expression levels
of IL-10 and TGF-β in the PF group were increased compared with
those in the model group (F=37.470 and 23.370, respectively). The
expression levels of IL-6, TNF-α, and IL-1β were decreased in the
PF group compared with those in the model group (F=58.343, 19.526,
and 11.615, respectively). Data are presented as the mean ± SD.
**P<0.001 vs. control group; ##P<0.001
vs. model group. |
PF reduces DSS-induced inflammation
via regulation of the PI3K/AKT signaling pathway
The mRNA expression levels of PI3K, AKT1, mTOR,
BCL2, and EGFR were significantly increased in the model group
compared with those in the control group (all P<0.05). After
treatment with PF, the mRNA expression levels of PI3K, AKT1, mTOR,
BCL2, and EGFR were significantly decreased compared with those in
the model group (P<0.05, Fig.
7).
 | Figure 7PF downregulated the mRNA expression
levels of AKT1, BCL2, EGFR, mTOR, and PI3K in the colonic mucosa.
The mRNA expression levels of PI3K, AKT1, mTOR, BCL2, and EGFR were
significantly increased in the model group compared with those in
the control group (F=8.256, 19.157, 9.787, 10.327, and 15.592,
respectively; P=0.001, <0.0001, <0.0001, 0.003, and
<0.0001, respectively). The mRNA expression levels of PI3K,
AKT1, mTOR, BCL2, and EGFR in the PF group were significantly
decreased compared with those in the model group (P=0.002,
<0.0001, 0.007, <0.0001, and <0.0001, respectively). Data
are presented as the mean ± SD. *P<0.05 and
**P<0.01 vs. control group; ##P<0.01
vs. model group. PF, paeoniflorin. |
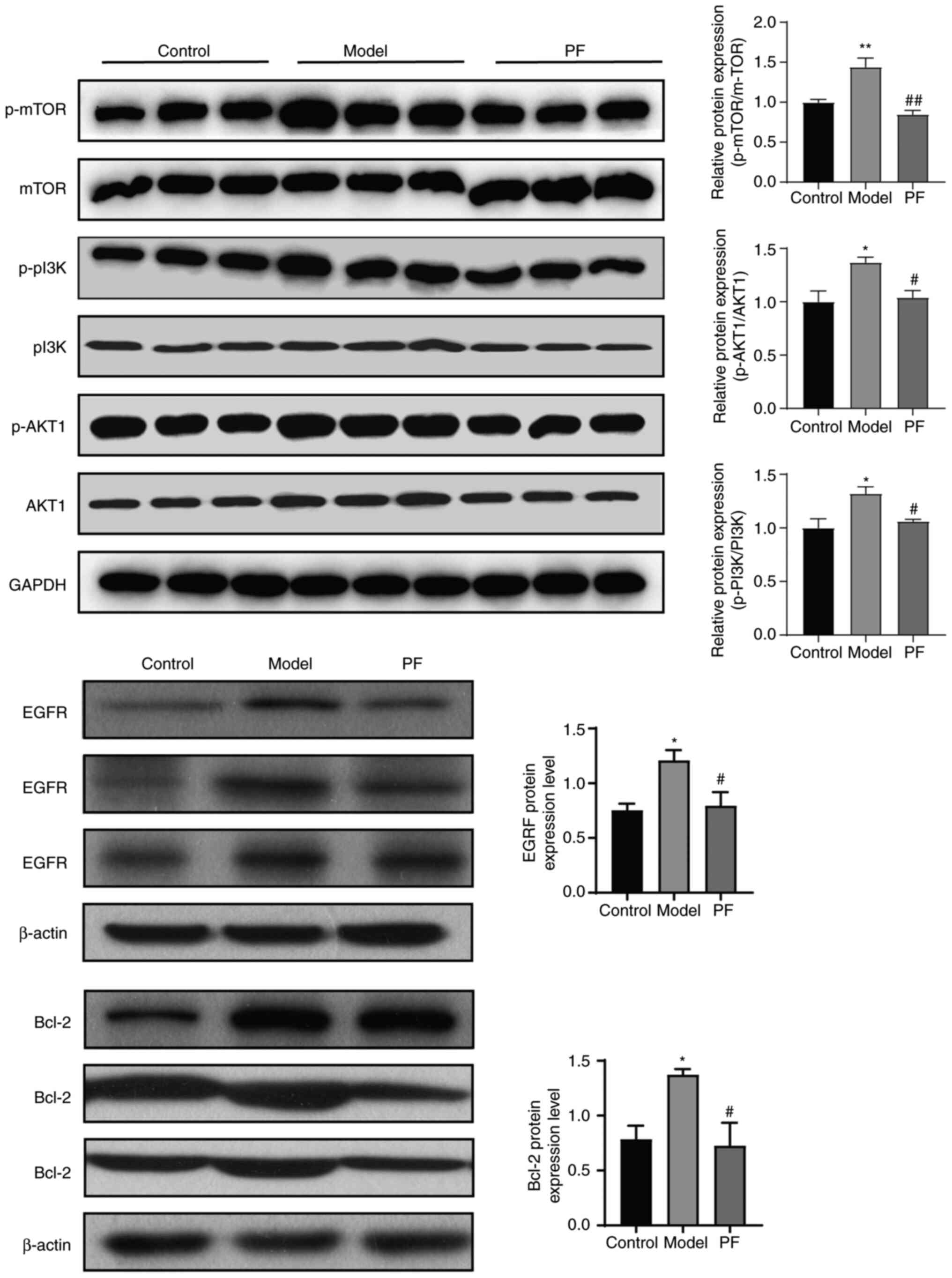
As shown in Fig. 8,
western blot results showed that there was a marked increase in the
protein expression levels of p-PI3K/PI3K, p-AKT1/AKT1, and
p-mTOR/mTOR in the colonic mucosa of the model group after the
administration of DSS in the drinking water (P<0.05). The
expression levels of Bcl-2 and EGFR were also increased in the
model group, but the differences were not significant (P>0.05).
In the PF group, the protein levels of p-PI3K/PI3K, p-AKT1/AKT1,
and p-mTOR/mTOR, Bcl-2, and EGFR were decreased compared with those
in the model group (P<0.05). Taken together, these results
demonstrate that PF could protect against UC partially through
inhibition of the PI3K/AKT signaling pathway.
Discussion
UC, a type of inflammatory bowel disease, is
characterized by abdominal pain, diarrhea, rectal bleeding, and
mucopurulent bloody stools. The incidence and prevalence rates of
UC have significantly increased based on hospital-based studies and
register studies in China (44,45)
with a significant increasingly trend of UC prevalence from 2013 to
2016 of 24.2% reported by another study (46). Several factors are correlated with
the occurrence of UC including the gut microbiota, genetic factors,
antibiotics use (47,48). The primary treatment options for UC
are 5-aminosalicylic acid, corticosteroids, immunosuppressants, as
well as fecal microbiota transplantation, although they are limited
by suboptimal clinical efficacy and several side effects (48-50).
In recent years, certain natural compounds have received increasing
attention given their safety and efficacy, lack of side effects,
and wide-ranging pharmacological benefits, and have thus been used
for the treatment of a variety of diseases, including cancer,
diabetes, and UC (15,51-57).
PF is the primary active compound of Paeonia
suffruticosa, Paeonia suffruticosa, and Paeonia
lactiflora and has good, anti-tumor, anti-ulcer,
immunoregulatory, and neuroprotective effects (25,26).
In a network pharmacology and experimental validation study, Cao
et al (58) revealed that
PF can regulate inflammatory factors including IL-6 and IL-10
expression through the p38 MAPK signaling pathway in the treatment
of pancreatic cancer. Sun et al (59) found that PF downregulated IL-18,
TNF-α, and IL-1β expression levels in a rat model of diabetic foot
ulcers and was related to NLRP3/ASC/caspase-1 inflammasome
formation and NF-κB transcription. Several studies have revealed
that PF regulates the activation of immune cells, decreases the
release of inflammatory cytokines, and participates in the
regulation of the GPCR and PI3K/AKT/mTOR pathways (58-63).
PF has also been shown to regulate GPCR signaling by suppressing
the expression of β-arrestin 2 and upregulating cAMP-PKA signaling,
and this regulation was important for its therapeutic effect in
rheumatoid arthritis (60).
Another study found that PF could protect against cholestasis by
activating Nrf2 via a PI3K/Akt-dependent pathway (64). It was also shown that PF could
alleviate liver fibrosis and inhibit the activation of immune cells
by inhibiting HIF-1α expression partly through suppressing the mTOR
pathway (60).
However, the molecular mechanisms underlying the
protective effects of PF against DSS-induced UC remain unknown. To
determine the mechanism of action of PF in the treatment of UC,
network pharmacology was used to predict the target genes and
pathways of its action, followed by experimental validation.
The results showed that PF treatment and the
pathogenesis of UC shared 60 common target genes, and the top 10
targets were ALB, AKT1, MMP9, CASP3, SRC, EGFR, IGF1, MMP2, ESR1,
IL2 and MAPK14, which are ranked according to degree values. The
pathways identified included proteoglycans in cancer, EGFR tyrosine
kinase inhibitor resistance, endocrine resistance, prostate cancer,
fluid shear stress and atherosclerosis, IL-17, PI3K/AKT, and HIF-1
signaling pathways. AKT1, BCL2L1, EGFR, FGFR2, IGF1, IGF1R, IL2,
JAK2, KDR, MET, NOS3, and SYK were identified as genes that
participate in the PI3K/AKT signaling pathway.
The findings of the present study are consistent
with those of several previous studies. Multiple pathways have been
revealed to be involved in the pathogenesis and progression of UC
including the MAPK/AP-1, PPAR-α, PI3K/AKT, and NF-κB signaling
pathways (65,66). Studies have found that the PI3K/AKT
signaling pathway is related to the production of proinflammatory
cytokines, such as TNF-α, NF-κB, IL-6, and IL-17, and regulates
inflammation, apoptosis, and cell proliferation by modulating the
activity of downstream effector molecules, such as NF-κB, Bcl-2,
caspase 9, and mTOR (67,68). Once the PI3K/AKT signaling pathway
is activated by Akt, it can regulate the key cellular processes
involved in the immune-inflammatory response and induce NF-κB to
produce anti-apoptotic and pro-inflammatory effects (69).
The pathogenesis of UC may be associated with the
abnormal proliferation of colonic epithelial cells, enhanced
leukocyte infiltration into the colonic interstitium, elevated
mucosal T-cell activation, increased proinflammatory cytokine
production, and dysregulation of signal transduction pathways
(70-76).
Inflammation plays an important role in the progression of UC
(77). Anti-TNF agents have been
assessed for the treatment of UC in clinical settings and they
showed adequacy as a long-term maintenance therapy (78,79).
Researchers have found that IL-1β and IL-6 can promote an immune
response and that their expression levels are associated with
genetic susceptibility and steroid dependence in patients with UC
(80,81).
PF has also been shown to have an inhibitory effect
on inflammation. A study by Tao et al (82) confirmed that the use of PF at 5-20
mg/kg significantly inhibited the levels of inflammatory mediators
such as NF-κB, TNF-α, IL-1β, and IL-6 and downregulated caspase-3
expression. Another study showed that Moutan Cortex Radicis,
a Chinese herb that contains PF as its primary compound, can
inhibit the activation of NF-κB and decrease IL-6 and TNF-α
expression levels in a DSS-induced colitis model (83). PF has also been shown to
significantly reduce the occurrence of liver fibrosis, inhibit the
inflammatory response caused by cholestasis primarily by
upregulating the PI3K/Akt/Nrf2 pathway (24), and inhibit the expression of HIF-1α
through the mTOR pathway (84).
PF is able to ameliorate the inflammatory degree of
DSS-induced colitis, primarily by inhibiting NF-κB pathway
activation and downregulating the expression of proinflammatory
factors; it also inhibits the reduction of eosinophil-related
chemokine gene expression and eosinophil infiltration and slightly
inhibits COX2 activity, as demonstrated by enzymatic assays
(85). PF can inhibit LPS-induced
inflammation in Caco-2 cell monolayers, which is correlated with
its inhibition of NF-κB activation and reduction of IL-6, iNOS,
TNF-α, and COX-2 expression (62).
Several studies have demonstrated that PF can suppress the
production of pro-inflammatory cytokines such as IL-1β, IL-6, and
TNF-α in inflammatory conditions and have demonstrated that this is
related to the NF-κB and JAK2/STAT3 pathways (84-88).
The total glucosides of peonies, which consist of PF, has been
found to improve the pathological state of a DSS-induced IBD animal
model and to downregulate the expression of TNF-α, IL-17A, IL-23,
and IFN-γ in colonic tissues to improve mucosal barrier function
(89).
The results of the present study showed that PF can
reduce the levels of IL-6, TNF-α, and IL-1β, which was consistent
with the findings of the abovementioned studies, indicating that PF
exerts its therapeutic effect by improving the pathological
condition of UC by inhibiting the inflammatory response.
The Bcl-2 family has been found to play an important
role in the modulation of apoptosis including DNA cleavage and
plasma membrane blebbing and also negatively regulates cell death
(90,91). Studies have shown that medical
treatment of UC may be accomplished via suppression of apoptosis by
modulating BAX and Bcl-2 levels and caspase activity (91). Gu et al (92) found that PF blocked the nuclear
translocation of NF-κB, downregulated the expression of COX-2 and
Bcl-2, and upregulated the expression of BAX by inhibiting the
MAPK/NF-κB pathway, thereby regulating intestinal immune
abnormalities, inhibiting inflammation and therefore reducing UC.
In addition, studies have suggested that the PI3K/AKT signaling
pathway may be involved in both inflammatory and cancerous
processes in UC and that activation of the EGFR/PI3K/AKT pathway
can increase the expression of vascular endothelial growth factor
by upregulating HIF-1α, which can lead to slower blood flow and
increased interstitial blood pressure, forming tumor vessels
(66,93,94).
The EGFR signaling pathway plays a critical role in
the regulation of colonic epithelial biology and the response to
injury and inflammation and is activated in colonic macrophages in
mice with DSS-induced colitis with UC (95). The present study found that PF can
decrease the expression levels of Bcl-2 and EGFR in a mouse model
of UC. The results revealed that the mechanism of action of PF was
related to EGFR and Bcl-2, which was consistent with the findings
of a previous study (96).
The results of the present study also showed that PF
could decrease the expression levels of p-AKT1, Bcl-2, p-mTOR, and
p-PI3K. The animal experiments demonstrated that PF treatment
improved the pathological structure of the animals, increased the
levels of IL-10 and TGF-β, and decreased the levels of IL-6, TNF-α,
and IL-1β. Taken together, it was confirmed that PF could protect
against UC through inhibition of the PI3K/AKT signaling pathway. In
other studies, wortmannin was used as a PI3K inhibitor and it was
found that it could relieve the clinical symptoms, alleviate the
inflammatory response, and decrease the secretion of TNF-α in the
intestinal mucosa of UC patients and in a mouse model of colitis
(66). This suggested that the
activation of PI3K resulted in damage to the intestinal mucosa by
increasing the secretion of pro-inflammatory cytokines. The
application of PI3K/Akt antagonists may be used to treat UC in the
future. However, here it was found that PF exerted its effects
through affecting multiple pathways, and the use of a PI3K/AKT
agonist or antagonist may lead to other side effects or affect
other biological processes. Thus, PI3K/Akt agonists and antagonists
were not included in the present study. The present study also
confirmed that the mechanism of PF in the treatment of UC was
associated with the regulation of the HIF-1 pathway, which is
worthy of future study.
In conclusion, 60 common genes were identified
between PF- and UC-related targets. The top 10 corresponding genes
included ALB, AKT1, MMP9, CASP3, SRC, EGFR, IGF1, MMP2, ESR1, IL2
and MAPK14. GO functional and KEGG pathway enrichment analyses
indicated that the molecular mechanisms of action of PF in UC were
closely related to important biochemical processes and signaling
pathways such as EGFR tyrosine kinase inhibitor resistance, the
IL-17 signaling pathway, the PI3K-Akt signaling pathway, and the
HIF-1 signaling pathway. PF treatment reduced colonic damage caused
by DSS and decreased IL-6, TNF-α, IL-1β, PI3K/Akt pathway-related
activity, and EGFR, PI3K, mTOR, and Bcl-2 mRNA and protein
expression levels. Thus, it was found that the PI3K/Akt pathway
played a critical role in PF-mediated treatment of UC.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by the National Natural Science
Foundation of China (grant no. 81603509), the Natural Science
Foundation of Shandong Province (grant nos. ZR2014HQ051 and
ZR2021QH129), and the Key Research and Development Plan of Jining
(grant no. 2021YXNS007).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
BY conceived and designed the study. SZ, QL and KN
designed the experiments. YQ, KN, BL and CZ acquired the data. SZ,
KN, YL, YZ, BL and BY analyzed and interpreted the data. KN and BY
wrote and revised the manuscript. BY, SZ and KN confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent participate
This study was approved by the Ethics Committee of
Jining Medical University (approval no. JNMC-2020-DW-ZXY-001).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Feuerstein JD, Moss AC and Farraye FA:
Ulcerative colitis. Mayo Clin Proc. 94:1357–1373. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Neurath MF and Leppkes M: Resolution of
ulcerative colitis. Semin Immunopathol. 41:747–756. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Krishna M, Britto S, Qian J, Ihekweazu F,
Rodriguez JR and Kellermayer R: Diagnostic delay and colectomy risk
in pediatric ulcerative colitis. J Pediatr Surg. 55:403–405.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Danese S and Fiocchi C: Ulcerative
colitis. N Engl J Med. 365:1713–1725. 2011.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Loftus EV Jr: Clinical epidemiology of
inflammatory bowel disease: Incidence, prevalence, and
environmental influences. Gastroenterology. 126:1504–1517.
2004.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Langholz E, Munkholm P, Nielsen OH,
Kreiner S and Binder V: Incidence and prevalence of ulcerative
colitis in Copenhagen county from 1962 to 1987. Scand J
Gastroenterol. 26:1247–1256. 1991.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kaistha A and Levine J: Inflammatory bowel
disease: The classic gastrointestinal autoimmune disease. Curr
Probl Pediatr Adolesc Health Care. 44:328–334. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Miyamoto Y, Suyama K and Baba H: Recent
advances in targeting the EGFR signaling pathway for the treatment
of metastatic colorectal cancer. Int J Mol Sci.
18(752)2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Gisbert JP and Chaparro M: Acute severe
ulcerative colitis: State of the art treatment. Best Pract Res Clin
Gastroenterol. 32-33:59–69. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Duijvestein M, Battat R, Vande Casteele N,
D'Haens GR, Sandborn WJ, Khanna R, Jairath V and Feagan BG: Novel
therapies and treatment strategies for patients with inflammatory
bowel disease. Curr Treat Options Gastroenterol. 16:129–146.
2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Wang YN, Li J, Zheng WY, Wu D, Yang H, Li
Y, Lv H, Tan B, Shu HJ, Sun XY, et al: Clinical characteristics of
ulcerative colitis-related colorectal cancer in Chinese patients. J
Dig Dis. 18:684–690. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Bezzio C, Furfaro F, de Franchis R, Maconi
G, Asthana AK and Ardizzone S: Ulcerative colitis: Current
pharmacotherapy and future directions. Expert Opin Pharmacother.
15:1659–1670. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kobayashi T, Siegmund B, Le Berre C, Wei
SC, Ferrante M, Shen B, Bernstein CN, Danese S, Peyrin-Biroulet L
and Hibi T: Ulcerative colitis. Nat Rev Dis Primers.
6(74)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gao Q, Tian W, Yang H, Hu H, Zheng J, Yao
X, Hu B and Liu H: Shen-Ling-Bai-Zhu-San alleviates the imbalance
of intestinal homeostasis in dextran sodium sulfate-induced colitis
mice by regulating gut microbiota and inhibiting the NLRP3
inflammasome activation. J Ethnopharmacol.
319(117136)2024.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen K, Lou Y and Zhu Y: Tong Xie Yao
Fang: A classic Chinese medicine prescription with potential for
the treatment of ulcerative colitis. Evid Based Complement Alternat
Med. 2021(5548764)2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Niu C, Hu XL, Yuan ZW, Xiao Y, Ji P, Wei
YM and Hua YL: Pulsatilla decoction improves DSS-induced colitis
via modulation of fecal-bacteria-related short-chain fatty acids
and intestinal barrier integrity. J Ethnopharmacol.
300(115741)2023.PubMed/NCBI View Article : Google Scholar
|
|
17
|
He Z, Zhou Q, Wen K, Wu B, Sun X, Wang X
and Chen Y: Huangkui Lianchang decoction ameliorates DSS-induced
ulcerative colitis in mice by inhibiting the NF-kappaB signaling
pathway. Evid Based Complement Alternat Med.
2019(1040847)2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Li R, Chen Y, Shi M, Xu X, Zhao Y, Wu X
and Zhang Y: Gegen Qinlian decoction alleviates experimental
colitis via suppressing TLR4/NF-κB signaling and enhancing
antioxidant effect. Phytomedicine. 23:1012–1020. 2016.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ye Q, Hu Z, Yang M, Qin K and Zhou Y:
Effects and mechanisms of Chinese herbal medicine for ulcerative
colitis: Protocol for a systematic review and meta-analysis.
Medicine (Baltimore). 99(e19768)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang HY, Zeng HR, Wei HZ, Chu XY, Zhu HT,
Zhao B and Zhang Y: Tongxie-Yaofang formula regulated macrophage
polarization to ameliorate DSS-induced colitis via NF-κB/NLRP3
signaling pathway. Phytomedicine. 107(154455)2022.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhu Y, Li Q, Li H, Xie Y, Sun M, Zheng C
and Yu B: Effect of Tongxie Yaofang on the expression of
inflammatory factors in rats of stagnation of liver and spleen
deficiency syndrome. Lishizhen Med Mater Med Res. 29:1053–1057.
2018.
|
|
22
|
Zheng K, Jia J, Yan S, Shen H, Zhu P and
Yu J: Paeoniflorin ameliorates ulcerative colitis by modulating the
dendritic cell-mediated TH17/Treg balance. Inflammopharmacology.
28:1705–1716. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhao DD, Jiang LL, Li HY, Yan PF and Zhang
YL: Chemical components and pharmacological activities of terpene
natural products from the genus paeonia. Molecules.
21(1362)2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ma X, Zhang W, Jiang Y, Wen J, Wei S and
Zhao Y: Paeoniflorin, a natural product with multiple targets in
liver diseases-a mini review. Front Pharmacol.
11(531)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhai A, Zhang Z and Kong X: Paeoniflorin
alleviates H2O2-induced oxidative injury through down-regulation of
MicroRNA-135a in HT-22 cells. Neurochem Res. 44:2821–2831.
2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Xin Q, Yuan R, Shi W, Zhu Z, Wang Y and
Cong W: A review for the anti-inflammatory effects of paeoniflorin
in inflammatory disorders. Life Sci. 237(116925)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Wang Y, Zhou Y, Lin H, Chen H and Wang S:
Paeoniflorin inhibits the proliferation and metastasis of
ulcerative colitis-associated colon cancer by targeting EGFL7. J
Oncol. 2022(7498771)2022.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Luo TT, Lu Y, Yan SK, Xiao X, Rong XL and
Guo J: Network pharmacology in research of chinese medicine
formula: Methodology, application and prospective. Chin J Integr
Med. 26:72–80. 2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhou Z, Chen B, Chen S, Lin M, Chen Y, Jin
S, Chen W and Zhang Y: Applications of network pharmacology in
traditional chinese medicine research. Evid Based Complement
Alternat Med. 2020(1646905)2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT and
Lin CY: cytoHubba: Identifying hub objects and sub-networks from
complex interactome. BMC Syst Biol. 8 (Suppl 4)(S11)2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Assenov Y, Ramírez F, Schelhorn SE,
Lengauer T and Albrecht M: Computing topological parameters of
biological networks. Bioinformatics. 24:282–284. 2008.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
RStudio Team: RStudio: Integrated
development for R. RStudio, PBC., Boston MA, 2015.
|
|
35
|
Team RC: R: A language and Environment for
Statistical Computing. MSOR connections, pp1, 2014.
|
|
36
|
The Gene Ontology Consortium. The gene
ontology resource: 20 Years and still GOing strong. Nucleic Acids
Res. 47:D330–D338. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kanehisa M: Post-Genome Informatics.
Oxford University Press, Oxford, 2000.
|
|
39
|
Morris GM, Huey R, Lindstrom W, Sanner MF,
Belew RK, Goodsell DS and Olson AJ: AutoDock4 and AutoDockTools4:
Automated docking with selective receptor flexibility. J Comput
Chem. 30:2785–2791. 2009.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Trott O and Olson AJ: AutoDock Vina:
Improving the speed and accuracy of docking with a new scoring
function, efficient optimization, and multithreading. J Comput
Chem. 31:455–461. 2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wang K, Li YF, Lv Q, Li XM, Dai Y and Wei
FZ: Bergenin, acting as an agonist of PPARγ, ameliorates
experimental colitis in mice through improving expression of SIRT1,
and therefore inhibiting NF-κB-mediated macrophage activation.
Front Pharmacol. 8(981)2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Xie X, Liu P, Wu H, Li H, Tang Y, Chen X,
Xu C, Liu X and Dai G: miR-21 antagonist alleviates colitis and
angiogenesis via the PTEN/PI3K/AKT pathway in colitis mice induced
by TNBS. Ann Transl Med. 10(413)2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Wang Y and Ouyang Q: APDW 2004 Chinese IBD
working group. Ulcerative colitis in China: Retrospective analysis
of 3100 hospitalized patients. J Gastroenterol Hepatol.
22:1450–1455. 2007.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Jiang XL and Cui HF: An analysis of 10218
ulcerative colitis cases in China. World J Gastroenterol.
8:158–161. 2002.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Yang H, Zhou R, Bai X, Guo M, Ruan G, Wang
L and Qian J: Trend and geographic variation in incidence and
prevalence of inflammatory bowel disease in regions across China: A
nationwide employee study between 2013 and 2016. Front Med
(Lausanne). 9(900251)2022.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Han Z, Tan X, Sun J, Wang T, Yan G, Wang C
and Ma K: Systems pharmacology and transcriptomics reveal the
mechanisms of Sanhuang decoction enema in the treatment of
ulcerative colitis with additional Candida albicans infection. Chin
Med. 16(75)2021.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Ananthakrishnan AN, Bernstein CN,
Iliopoulos D, Macpherson A, Neurath MF, Ali RAR, Vavricka SR and
Fiocchi C: Environmental triggers in IBD: A review of progress and
evidence. Nat Rev Gastroenterol Hepatol. 15:39–49. 2018.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Burri E, Maillard MH, Schoepfer AM,
Seibold F, Van Assche G, Rivière P, Laharie D and Manz M: Swiss
IBDnet, an official working group of the Swiss Society of
Gastroenterology. Treatment algorithm for mild and
moderate-to-severe ulcerative colitis: An update. Digestion. 101
(Suppl 1):S2–S15. 2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Segal JP, LeBlanc JF and Hart AL:
Ulcerative colitis: An update. Clin Med (Lond). 21:135–139.
2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Hirten RP and Sands BE: New therapeutics
for ulcerative colitis. Annu Rev Med. 72:199–213. 2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Azab A, Nassar A and Azab AN:
Anti-inflammatory activity of natural products. Molecules.
21(1321)2016.PubMed/NCBI View Article : Google Scholar
|
|
53
|
An J, Chen B, Kang X, Zhang R, Guo Y, Zhao
J and Yang H: Neuroprotective effects of natural compounds on
LPS-induced inflammatory responses in microglia. Am J Transl Res.
12:2353–2378. 2020.PubMed/NCBI
|
|
54
|
Wei M, Li H, Li Q, Qiao Y, Ma Q, Xie R,
Wang R, Liu Y, Wei C, Li B, et al: Based on network pharmacology to
explore the molecular targets and mechanisms of Gegen Qinlian
decoction for the treatment of ulcerative colitis. Biomed Res Int.
2020(5217405)2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Bindman AB and Cox DF: Changes in health
care costs and mortality associated with transitional care
management services after a discharge among medicare beneficiaries.
JAMA Intern Med. 178:1165–1171. 2018.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Deng X, Xing X, Sun G, Xu X, Wu H, Li G
and Sun X: Guanxin danshen formulation protects against myocardial
ischemia reperfusion injury-induced left ventricular remodeling by
upregulating estrogen receptor β. Front Pharmacol.
8(777)2017.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Wei J, Guo F, Zhang M, Xian M, Wang T, Gao
J, Wu H, Song L, Zhang Y, Li D, et al: Signature-oriented
investigation of the efficacy of multicomponent drugs against heart
failure. FASEB J. 33:2187–2198. 2019.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Cao C, Zhao W, Chen X, Shen B, Wang T, Wu
C and Rong X: Deciphering the action mechanism of paeoniflorin in
suppressing pancreatic cancer: A network pharmacology study and
experimental validation. Front Pharmacol.
13(1032282)2022.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Sun X, Wang X, Zhao Z, Chen J, Li C and
Zhao G: Paeoniflorin inhibited nod-like receptor protein-3
inflammasome and NF-κB-mediated inflammatory reactions in diabetic
foot ulcer by inhibiting the chemokine receptor CXCR2. Drug Dev
Res. 82:404–411. 2021.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Zhang L and Wei W: Anti-inflammatory and
immunoregulatory effects of paeoniflorin and total glucosides of
paeony. Pharmacol Ther. 207(107452)2020.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Chang Y, Zhai L, Peng J, Wu H, Bian Z and
Xiao H: Phytochemicals as regulators of Th17/Treg balance in
inflammatory bowel diseases. Biomed Pharmacother.
141(111931)2021.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Wu XX, Huang XL, Chen RR, Li T, Ye HJ, Xie
W, Huang ZM and Cao GZ: Paeoniflorin prevents intestinal barrier
disruption and inhibits lipopolysaccharide (LPS)-induced
inflammation in Caco-2 cell monolayers. Inflammation. 42:2215–2225.
2019.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Chen M, Cao L, Luo Y, Feng X, Sun L, Wen M
and Peng S: Paeoniflorin protects against concanavalin A-induced
hepatitis in mice. Int Immunopharmacol. 24:42–49. 2015.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Chen Z, Ma X, Zhu Y, Zhao Y, Wang J, Li R,
Chen C, Wei S, Jiao W, Zhang Y, et al: Paeoniflorin ameliorates
ANIT-induced cholestasis by activating Nrf2 through an
PI3K/Akt-dependent pathway in rats. Phytother Res. 29:1768–1775.
2015.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Li N, Sun W, Zhou X, Gong H, Chen Y, Chen
D and Xiang F: Dihydroartemisinin protects against dextran sulfate
sodium-induced colitis in mice through inhibiting the PI3K/AKT and
NF-κB signaling pathways. Biomed Res Int.
2019(1415809)2019.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Huang XL, Xu J, Zhang XH, Qiu BY, Peng L,
Zhang M and Gan HT: PI3K/Akt signaling pathway is involved in the
pathogenesis of ulcerative colitis. Inflamm Res. 60:727–734.
2011.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Setia S, Nehru B and Sanyal SN:
Upregulation of MAPK/Erk and PI3K/Akt pathways in ulcerative
colitis-associated colon cancer. Biomed Pharmacother. 68:1023–1029.
2014.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Chen Q, Duan X, Fan H, Xu M, Tang Q, Zhang
L, Shou Z, Liu X, Zuo D, Yang J, et al: Oxymatrine protects against
DSS-induced colitis via inhibiting the PI3K/AKT signaling pathway.
Int Immunopharmacol. 53:149–157. 2017.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Wei J and Feng J: Signaling pathways
associated with inflammatory bowel disease. Recent Pat Inflamm
Allergy Drug Discov. 4:105–117. 2010.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Vetuschi A, Latella G, Sferra R, Caprilli
R and Gaudio E: Increased proliferation and apoptosis of colonic
epithelial cells in dextran sulfate sodium-induced colitis in rats.
Dig Dis Sci. 47:1447–1457. 2002.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Laukoetter MG, Nava P, Lee WY, Severson
EA, Capaldo CT, Babbin BA, Williams IR, Koval M, Peatman E,
Campbell JA, et al: JAM-A regulates permeability and inflammation
in the intestine in vivo. J Exp Med. 204:3067–3076. 2007.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Panés J and Granger DN:
Leukocyte-endothelial cell interactions: Implications for the
pathogenesis and treatment of gastrointestinal disease. Dig Dis.
12:232–241. 1994.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Sturm A, de Souza HS and Fiocchi C:
Mucosal T cell proliferation and apoptosis in inflammatory bowel
disease. Current Drug Targets. 9:381–387. 2008.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Brown SJ and Mayer L: The immune response
in inflammatory bowel disease. Am J Gastroenterol. 102:2058–2069.
2007.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Bisping G, Lügering N, Lütke-Brintrup S,
Pauels HG, Schürmann G, Domschke W and Kucharzik T: Patients with
inflammatory bowel disease (IBD) reveal increased induction
capacity of intracellular interferon-gamma (IFN-gamma) in
peripheral CD8+ lymphocytes co-cultured with intestinal epithelial
cells. Clin Exp Immunol. 123:15–22. 2001.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Atreya I, Atreya R and Neurath MF:
NF-kappaB in inflammatory bowel disease. J Intern Med. 263:591–596.
2008.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Patil DT, Moss AC and Odze RD: Role of
histologic inflammation in the natural history of ulcerative
colitis. Gastrointest Endosc Clin N Am. 26:629–640. 2016.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Jentzer A, Veyrard P, Roblin X,
Saint-Sardos P, Rochereau N, Paul S, Bourlet T, Pozzetto B and
Pillet S: Cytomegalovirus and inflammatory bowel diseases (IBD)
with a special focus on the link with ulcerative colitis (UC).
Microorganisms. 8(1078)2020.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Ben-Horin S, Kopylov U and Chowers Y:
Optimizing anti-TNF treatments in inflammatory bowel disease.
Autoimmun Rev. 13:24–30. 2014.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Yamamoto-Furusho JK, Santiago-Hernández
JJ, Pérez-Hernández N, Ramírez-Fuentes S, Fragoso JM and
Vargas-Alarcón G: Interleukin 1 β (IL-1B) and IL-1 antagonist
receptor (IL-1RN) gene polymorphisms are associated with the
genetic susceptibility and steroid dependence in patients with
ulcerative colitis. J Clin Gastroenterol. 45:531–535.
2011.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Bernardo D, Vallejo-Díez S, Mann ER,
Al-Hassi HO, Martínez-Abad B, Montalvillo E, Tee CT, Murugananthan
AU, Núñez H, Peake ST, et al: IL-6 promotes immune responses in
human ulcerative colitis and induces a skin-homing phenotype in the
dendritic cells and Tcells they stimulate. Eur J Immunol.
42:1337–1353. 2012.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Tao YE, Wen Z, Song Y and Wang H:
Paeoniflorin attenuates hepatic ischemia/reperfusion injury via
anti-oxidative, anti-inflammatory and anti-apoptotic pathways. Exp
Ther Med. 11:263–268. 2016.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Chen TF, Hsu JT, Wu KC, Hsiao CF, Lin JA,
Cheng YH, Liu YH, Lee DY, Chang HH, Cho DY and Hsu JL: A systematic
identification of anti-inflammatory active components derived from
Mu Dan Pi and their applications in inflammatory bowel disease. Sci
Rep. 10(17238)2020.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Zhao Y, Ma X, Wang J, Zhu Y, Li R, Wang J,
He X, Shan L, Wang R, Wang L, et al: Paeoniflorin alleviates liver
fibrosis by inhibiting HIF-1α through mTOR-dependent pathway.
Fitoterapia. 99:318–327. 2014.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Li J, Ren S, Li M, Bi J, Yang G and Li E:
Paeoniflorin protects against dextran sulfate sodium (DSS)-induced
colitis in mice through inhibition of inflammation and eosinophil
infiltration. Int Immunopharmacol. 97(107667)2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Hu B, Xu G, Zhang X, Xu L, Zhou H, Ma Z,
Shen X, Zhu J and Shen R: Paeoniflorin attenuates inflammatory pain
by inhibiting microglial activation and Akt-NF-κB signaling in the
central nervous system. Cell Physiol Biochem. 47:842–850.
2018.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Wang D, Liu L, Li S and Wang C: Effects of
paeoniflorin on neurobehavior, oxidative stress, brain insulin
signaling, and synaptic alterations in intracerebroventricular
streptozotocin-induced cognitive impairment in mice. Physiol Behav.
191:12–20. 2018.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Wang G and Cheng N: Paeoniflorin inhibits
mast cell-mediated allergic inflammation in allergic rhinitis. J
Cell Biochem. 119:8636–8642. 2018.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Cao XY, Ni JH, Wang X, Feng GZ, Li HD, Bao
WL, Wang YR, You KY, Weng HB and Shen XY: Total glucosides of
paeony restores intestinal barrier function through inhibiting
Lyn/Snail signaling pathway in colitis mice. Phytomedicine.
87(153590)2021.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Sun J, Zhang H, Guan L, Zhou H and Sun M:
Alpha-lipoic acid attenuates trinitrobenzene sulfonic acid-induced
ulcerative colitis in mice. Int J Clin Exp Med. 8:358–367.
2015.PubMed/NCBI
|
|
92
|
Gu P, Zhu L, Liu Y, Zhang L, Liu J and
Shen H: Protective effects of paeoniflorin on TNBS-induced
ulcerative colitis through inhibiting NF-kappaB pathway and
apoptosis in mice. Int Immunopharmacol. 50:152–160. 2017.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Lee G, Goretsky T, Managlia E, Dirisina R,
Singh AP, Brown JB, May R, Yang GY, Ragheb JW, Evers BM, et al:
Phosphoinositide 3-kinase signaling mediates beta-catenin
activation in intestinal epithelial stem and progenitor cells in
colitis. Gastroenterology. 139:869–881.e1-e9. 2010.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Karar J and Maity A: PI3K/AKT/mTOR pathway
in angiogenesis. Front Mol Neurosci. 4(51)2011.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Lu N, Wang L, Cao H, Liu L, Van Kaer L,
Washington MK, Rosen MJ, Dubé PE, Wilson KT, Ren X, et al:
Activation of the epidermal growth factor receptor in macrophages
regulates cytokine production and experimental colitis. J Immunol.
192:1013–1023. 2014.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Yao J, Cao X, Zhang R, Li YX, Xu ZL, Zhang
DG, Wang LS and Wang JY: Protective effect of baicalin against
experimental colitis via suppression of oxidant stress and
apoptosis. Pharmacogn Mag. 12:225–234. 2016.PubMed/NCBI View Article : Google Scholar
|