Introduction
Nitric oxide (NO) generated and released by
endothelial NO synthase (eNOS) exerts multiple beneficial effects
on blood vessels and plays a critical role in maintaining
cardiovascular homeostasis (1).
The dysregulation of NO synthesis attributable to the abnormal
activity and/or the expression of eNOS is considered to be a major
contributor to the pathogenesis of cardiovascular diseases, such as
hypertension and atherosclerosis (2,3).
Although the expression of eNOS is regulated at multiple levels,
increasing evidence has suggested that post-transcriptional
regulation plays an important role in the control of eNOS
expression. It has been demonstrated that eNOS expression is mainly
regulated through the modification of eNOS mRNA expression through
the 3′-untranslated region (3′-UTR) of eNOS mRNA (4–8).
Diabetes is associated with an increased risk of
cardiovascular disease due to multiple pathophysiological links
between insulin resistance, obesity, hypertension and atherogenic
dyslipidemia (9–11). One common characteristic of these
conditions is endothelial dysfunction, caused by a relative
deficiency of NO (12). Insulin
has vasodilator effects, which depend on the production of NO in
the vascular endothelium (13,14). Key signaling molecules mediating
metabolic actions of insulin, such as the insulin receptor tyrosine
kinase, phosphatidylinositol-3 kinase (PI3K) and Akt, are also
necessary for insulin to stimulate the production of NO in human
vascular endothelial cells (15,16). Thus, a physiological role of
insulin may be to couple the regulation of hemodynamic homeostasis
with metabolic homeostasis (17).
Previous studies have reported a common polymorphism
in the insulin receptor substrate-1 (IRS-1) gene, in which a
Gly/Arg substitution occurs at codon 972 (Arg972)
(18,19). The presence of Arg972
IRS-1 is associated with an impaired IRS-1 ability to activate
PI3K, leading to the diminished activity of insulin (18,19). In the present study, we
investigated the association between Arg972 IRS-1 and
eNOS expression/activity in a relatively large sample of human
subjects and primary cultures of human endothelial cells.
Materials and methods
Subjects
Between November 2012 and October 2013, 832 subjects
randomly selected from a total of 2,672 Han Chinese subjects in the
database of the physical examination center at Guangdong Provincial
People’s Hospital, Guangzhou, China were recruited in the present
study. Blood samples were drawn from all subjects within 72 h of
enrollment. The characteristics of the subjects are presented in
Table I. The present study
conforms to the Declaration of Helsinki and was approved by the
Ethics Committee of Guangdong Provincial People’s Hospital (for the
collection of patient blood samples, approval reference number:
H40125; and for obtaining umbilical vein endothelial cells,
approval reference number: H40126). Written informed consent was
obtained from all subjects prior to enrollment.
 | Table ICharacteristics of the 832 study
subjects. |
Table I
Characteristics of the 832 study
subjects.
| Variables | Mean + SD or n
(%) | Range |
|---|
| Age (years) | 53.5±6.3 | 33–76 |
| Age group
(years) |
| 30–39 | 123 (14.8) | – |
| 40–49 | 189 (22.7) | – |
| 50–59 | 202 (24.3) | – |
| 60–69 | 172 (20.7) | – |
| 70–79 | 146 (17.5) | – |
| Male | 426 (51.2) | – |
| Smoking | 184 (22.1) | – |
| BMI
(kg/m2) | 26.3±4.5 | 17.9–31.4 |
| Fasting glucose
(mmol/l | 5.4±2.1 | 3.1–11.2 |
| Total cholesterol
(mmol/l) | 5.8±1.7 | 3.6–9.5 |
| LDL-cholesterol
(mmol/l) | 3.8±1.5 | 1.7–6.7 |
| HDL-cholesterol
(mmol/l) | 1.4±0.6 | 0.6–2.4 |
| Triglycerides
(mmol/l) | 2.3±1.3 | 0.9–6.4 |
| Systolic blood
pressure (mmHg) | 124.3±19.5 | 96–185 |
| Diastolic blood
pressure (mmHg) | 86.7±21.2 | 59–126 |
| Plasma eNOS
(ng/ml) | 38.9±12.5 | 18.2–70.8 |
| Plasma
nitrite/nitrate (μM) | 9.2±2.7 | 2.2–12.9 |
| Hyperlipidemia | 151 (18.1) | – |
| Hypertension | 225 (27.0) | – |
| Coronary heart
disease | 106 (12.7) | – |
| Type 2 diabetes
mellitus | 90 (10.8) | – |
| Kidney disease | 52 (6.3) | – |
| Arg972
IRS-1 genotype |
| GG | 639 (76.8) | – |
| GA | 176 (21.2) | – |
| AA | 17 (2.0) | – |
Plasmids and reagents
A fragment of human genomic DNA containing the
entire coding sequence of IRS-1 was cloned and ligated into the
pcDNA3.1 expression vector as described in a previous study
(20). The Arg972
IRS-1 expression vector was constructed as previously described
(21). SuperFect and HiPerFect
transfection reagents were purchased from Qiagen (Valencia, CA,
USA). TRIzol reagent and SYBR-Green Master Mix were purchased from
Invitrogen (Carlsbad, CA, USA) and PE Applied Biosystems (Foster
City, CA, USA), respectively. Anti-eNOS (49G3; #9586),
anti-phospho-eNOS (Ser1177; #9570) and anti-β-actin (8H10D10;
#3700) antibodies were purchased from Cell Signaling Technology
(Danvers, MA, USA). The eNOS human ELISA kit (#ab166866) was
purchased from Abcam (Cambridge, MA, USA). The nitrite/nitrate
colorimetric assay kit (#780001) was purchased from Cayman Chemical
Co. (Ann Arbor, MI, USA). The selective PI3K inhibitor, BKM120
(#sc-364437A), and anti-IRS-1 (C-20; #sc-559), anti-Akt (5C10;
#sc-81434), anti-phospho-Akt (Ser473; #sc-101629) and anti- von
Willebrand factor (vWF) (1.B.690; #sc-73268) antibodies were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Mature miR-155 mimic (5′-UUAAUGCUAAUC
GUGAUAGGGGUCCCUAUCACGAUUAGCAUUAAUU-3′) was purchased from Shanghai
GenePharma Co., Ltd. (Shanghai, China). Antagomir-155 (#410078-00;
miRCURY LNA™ microRNA power inhibitor) was purchased from Exiqon
(Woburn, MA, USA). Endothelial cell growth medium (#211-500) was
purchased from Cell Applications, Inc. (San Diego, CA, USA). G418,
insulin and all other chemicals of reagent grade were purchased
from Sigma (St. Louis, MO, USA).
Human umbilical vein endothelial cell
(HUVEC) culture
Umbilical cords were obtained from mothers
delivering at Guangdong Provincial People’s Hospital. Primary HUVEC
cultures were obtained as previously described (22). Following perfusion of the
umbilical cords with 0.1% collagenase at 37°C, the HUVECs were
grown on 0.2% gelatin-coated tissue culture plates in endothelial
cell growth medium supplemented with 20% FBS and endothelial cell
growth supplement (Cell Applications, Inc.). In all the
experiments, the cells were used between the third and the fifth
passages. Commercial HUVECs (#200K-05n) were purchased from Cell
Applications, Inc. for the stable transfection of IRS-1 and
Arg972 IRS-1. We confirmed by sequencing that the
commercial HUVECs expressed homozygous wild-type (WT) IRS-1 (GG
genotype).
Genotyping and determination of eNOS and
nitrite/nitrate levels
The Arg972 IRS-1 polymorphism was
identified by sequencing. Plasma eNOS and nitrite/nitrate levels
were respectively determined with an eNOS human ELISA kit (Abcam)
and a nitrite/nitrate colorimetric assay kit (Cayman Chemical)
according to the manufacturer’s instructions. Nitrite/nitrate
levels in the HUVEC culture media were normalized to the total
proteins of HUVECs.
Transfection
Plasmid constructs were transfected into the HUVECs
using SuperFect transfection reagent (Qiagen) according to the
manufacturer’s instructions. Pools of stable transfectants of IRS-1
and Arg972 IRS-1 were generated through selection with
G418 (800 μg/ml) as per the manufacturer’s instructions.
Mature miR-155 mimic or antagomir-155 was transfected into the
HUVECs using HiPerFect transfection reagent (Qiagen) according to
the manufacturer’s instructions.
Reverse transcription-quantitative
RT-qPCR
Total RNA from the HUVECs was prepared using TRIzol
reagent (Invitrogen) and reverse transcribed into cDNA using
SuperScript II reverse transcriptase (Invitrogen) or microRNA
TaqMan kit (Applied Biosystems, Foster City, CA, USA). Quantitative
PCR (qPCR) was carried out on an ABI PRISM 7700 Sequence Detection
System (Applied Biosystems), using the fluorescent dye SYBR-Green
Master Mix (PE Applied Biosystems) as per the manufacturer’s
instructions. The results were normalized to those of the
housekeeping gene β-actin in the same sample. The primers used were
as follows: eNOS, 5′-GTGGCTGTCTGCATGGACCT-3′ (forward) and
5′-CCACGATGGTGACTTTGGCT-3′ (reverse); β-actin,
5′-AGCCTCGCCTTTGCCGA-3′ (forward) and 5′-CTGGTGCCTGGGGCG-3′
(reverse). Stem-loop RT-PCR (TaqMan MicroRNA assays; P/N: 000448
for miR-125a, P/N: 000449 for miR-125b, P/N: 000521 for miR-218,
P/N: 000451 for miR-126, P/N: 002623 for miR-155, P/N: 000397 for
miR-21, P/N: 000524 for miR-221 and P/N: 000525 for miR-222;
Applied Biosystems) was used to quantify the miRNAs according to
the manufacturer’s instructions. Small unclear RNA U6 was used as
an internal control with the following primers: forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The reverse primer of U6 was used for reverse transcription. Each
experiment was repeated 3 times in duplicate.
Western blot analysis
Western blot analysis was carried out as previously
described (23). The HUVECs were
lysed in 250 μl of 2X SDS loading buffer (62.5 mm Tris-HCl,
pH 6.8, 2% SDS, 25% glycerol, 0.01% bromphenol blue and 5%
2-mercaptoethanol) and incubated at 95°C for 10 min. Equal amounts
of protein (100 μg) for each sample were separated by 8–15%
SDS-polyacrylamide gel and blotted onto polyvinylidene difluoride
microporous membranes (Millipore, Billerica, MA, USA). The
membranes were incubated for 1 h with a 1:1,000 dilution of primary
antibody, and then washed and revealed using secondary antibodies
with horseradish peroxidase conjugate (1:5,000, 1 h). Peroxidase
was revealed using a GE Healthcare ECL kit (GE Healthcare,
Shanghai, China).
Immunofluorescence staining
The cells were washed twice with 10 mmol/l sodium
phosphate (pH 7.4), 0.14 mol/l NaCl (PBS) and subsequently fixed
for 10 min at room temperature with methanol. After fixation, the
cells were washed twice with PBS and incubated for 1 h with
anti-vWF antibody (Santa Cruz Biotechnology) (1:500 dilution).
Subsequently, the coverslips were washed twice with PBS and
incubated for 1 h with FITC-conjugated seconday antibody (Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA), embedded
in mounting medium, and analyzed under a fluorescence microscope
(Axioplan 2; Carl Zeiss, Oberkochen, Germany).
IRS-1-associated PI3K activity assay
IRS-1-associated PI3K activity was determined as
previously described (24).
Briefly, 700 μg of total protein were immunoprecipitated
with anti-IRS-1 antibody (Santa Cruz Biotechnology), and kinase
activity was detected by the appearance of radiolabeled
32P-labeled phosphatidylinositol 3-phosphate
([32P]PI-3-P) following thin-layer chromatography (TLC)
as previously described (25).
Autoradiographic signals were quantified using NIH Image software
version 1.63.
Statistical analysis
Statistical analyses were carried out using SPSS
15.0 software (IBM, Chicago, IL, USA). All continuous variable
values are expressed as the means ± SD. Comparisons of the means
between 2 independent groups were performed using the Student’s
t-tests. A stepwise multi-linear regression model was used to
determine which variables explained unique variance in plasma eNOS
levels. All variables listed in Table
I were tested in the multi-linear regression model. Comparisons
of the means among multiple groups were performed using one-way
ANOVA followed by post hoc pairwise comparisons using Tukey’s
tests. Categorical variables were compared using χ2
tests. A value of p=0.05 was considered to indicate a statistically
significant difference in the present study.
Results
Association between Arg972
IRS-1 and plasma eNOS and nitrite/nitrate levels in human
subjects
The dysregulation of NO synthesis attributable to
the abnormal expression and activity of eNOS is considered to be a
major contributor to the pathogenesis of vascular diseases
(2,3). The results from one of our pilot
in vitro studies suggested that Arg972 IRS-1
exerts significant regulatory effects on eNOS expression in human
endothelial cells. In the present study, we first genotyped 832
randomly selected human subjects for the Arg972 IRS-1
polymorphism and determined their plasma eNOS and nitrite/nitrate
levels. As shown in Table I,
76.8% of the total subjects carried the homozygous WT IRS-1 (GG
genotype), 21.2% carried the heterozygous Arg972 IRS-1
(GA genotype) and only 2% carried the homozygous Arg972
IRS-1 (AA genotype). The A allele frequency was found to be 12.6%
in our study population. Multivariate regression analysis revealed
that after adjustment for all variables except for the plasma
nitrite/nitrate level (which highly correlated with the plasma eNOS
level) listed in Table I, the
copy number of the Arg972 IRS-1 A allele, age, body mass
index (BMI), smoking and blood fasting glucose levels were
independent predictors of the plasma eNOS levels, collectively
explaining 50.8% of the total variance (Table II). Analyses of the subject
characteristics by the Arg972 IRS-1 genotypes revealed
that the Arg972 IRS-1 heterozygous and homozygous
carriers had significantly higher blood pressure and a prevalence
of hyperlipidemia, coronary heart disease and type 2 diabetes
mellitus (T2DM), but lower levels of plasma eNOS and
nitrite/nitrate than the homozygous WT IRS-1 carriers (Table III). These findings indicate
that Arg972 IRS-1 is associated with decreased eNOS
expression and activity in the human body.
| Table IIMultivariate regression analysis of
independent predictors of plasma endothelial nitric oxide synthase
(eNOS) level. |
Table II
Multivariate regression analysis of
independent predictors of plasma endothelial nitric oxide synthase
(eNOS) level.
| Dependent
variable | Independent
variable | Partial
r2 | Total
r2 | β co-efficient | P-value |
|---|
| Plasma eNOS
(ng/ml) | Copy number of
Arg972 IRS-1 A allele | 0.197 | 0.197 | −0.274 | 0.011 |
| Age | 0.140 | 0.337 | −0.228 | 0.015 |
| BMI | 0.082 | 0.419 | −0.202 | 0.016 |
| Smoking | 0.055 | 0.474 | −0.141 | 0.040 |
| Fasting
glucose | 0.034 | 0.508 | −0.095 | 0.047 |
| Table IIICharacteristics of study subjects by
Arg972 insulin receptor substrate-1 (IRS-1)
genotypes. |
Table III
Characteristics of study subjects by
Arg972 insulin receptor substrate-1 (IRS-1)
genotypes.
| Variables | Arg972
IRS-1 genotype
| P-value |
|---|
| GG (n=639) | GA (n=176) | AA (n=17) |
|---|
| Age (years) | 53.3±6.0 | 53.8±6.5 | 57.9±6.8 | 0.862 |
| Smoking
(n=184) | 136 (21.3) | 44 (25.0) | 4 (23.5) | 0.569 |
| BMI
(kg/m2) | 26.2±4.3 | 26.7±4.6 | 25.9±4.7 | 0.905 |
| Fasting glucose
(mmol/l) | 5.1±1.8 | 6.3±2.5 | 7.4±2.8 | 0.033a |
| Total cholesterol
(mmol/l) | 5.8±1.6 | 5.8±1.9 | 5.8±2.1 | 0.946 |
| LDL-cholesterol
(mmol/l) | 3.7±1.3 | 4.1±1.8 | 4.5±1.9 | 0.083 |
| HDL-cholesterol
(mmol/l) | 1.5±0.6 | 1.1±0.9 | 0.7±0.4 | 0.021a |
| Triglycerides
(mmol/l) | 2.0±1.0 | 3.2±1.6 | 4.3±1.9 | 0.007a |
| Systolic blood
pressure (mmHg) | 120.5±18.7 | 136.2±22.1 | 143.9±25.7 | 0.010a |
| Diastolic blood
pressure (mmHg) | 84.5±20.3 | 93.1±25.5 | 103.1±21.6 | 0.015a |
| Plasma eNOS
(ng/ml) | 41.2±10.4 | 32.7±15.8 | 27.0±16.9 | 0.019a |
| Plasma
nitrite/nitrate (μM) | 9.8±2.6 | 7.1±3.0 | 5.0±2.9 | 0.029a |
| Hyperlipidemia
(n=151) | 101 (15.8) | 39 (22.2) | 11 (64.7) | <0.001a |
| Hypertension
(n=225) | 170 (26.6) | 47 (26.7) | 9 (52.9) | 0.054 |
| Coronary heart
disease (n=106) | 67 (10.5) | 29 (16.5) | 10 (58.8) | <0.001a |
| Type 2 diabetes
mellitus (n=90) | 62 (9.7) | 25 (14.2) | 13 (76.5) | <0.001a |
| Kidney disease
(n=52) | 34 (5.3) | 12 (6.8) | 6 (35.3) | <0.001a |
eNOS expression/activity and eNOS
activation/phosphorylation in HUVECs expressing homozygous WT IRS-1
or heterozygous Arg972 IRS-1
To further examine the association between
Arg972 IRS-1 and eNOS expression/activity, we used HUVEC
cultures established from delivering mothers expressing homozygous
WT IRS-1 (GG genotype) or heterozygous Arg972 IRS-1 (GA
genotype). HUVEC cultures expressing homozygous Arg972
IRS-1 (AA genotype) were not included for the following reasons: i)
the low frequency of homozygous Arg972 IRS-1 in this
study (Han Chinese) population (Table
I); ii) heterozygous Arg972 IRS-1 carriers showed
significantly decreased plasma eNOS and nitrite/nitrate levels
compared to the homozygous WT IRS-1 carriers (Table III). Based on a two-sided
p=0.05, power=0.80 and an effect size=1.6 (based on data from our
pilot in vitro study), a sample size of 8 was calculated for
the comparison of eNOS expression between 2 HUVEC groups (26). As shown in Fig. 1A, the majority of the cultured
HUVECs (~90%) displayed typical endothelial morphology and
immunofluorescence staining for the endothelial cell marker, vWF.
Without insulin stimulation, the HUVECs showed no significant
differences in eNOS expression and nitrite/nitrate levels; we
examined different methods of insulin stimulation, and found that
treatment with 10−8 M of insulin for 24 h had the
strongest stimulatory effects on eNOS expression and
nitrite/nitrate levels in the HUVECs (data not shown). Thus, in all
subsequent experiments in the present study, the HUVECs were
pre-stimulated with 10−8 M of insulin for 24 h.
Following insulin stimulation, HUVECs expressing heterozygous
Arg972 IRS-1 (GA group) showed a 38% decrease in the
eNOS mRNA level compared with those expressing homozygous WT IRS-1
(GG group) (Fig. 1B). Western
blot analyses revealed that compared with the GG group of HUVECs,
the GA group showed a 42% decrease in the eNOS protein level, and a
58% decrease in eNOS phosphorylation at serine 1177 (Ser1177),
which is required for the insulin-stimulated activation of eNOS
activity (Fig. 1C) (15). In addition, IRS-1-associated PI3K
activity and eNOS activity (measured by the nitrite/nitrate level
in the cell culture media) in the GA group decreased by
approximately 30 and 70%, respectively (Fig. 1D).
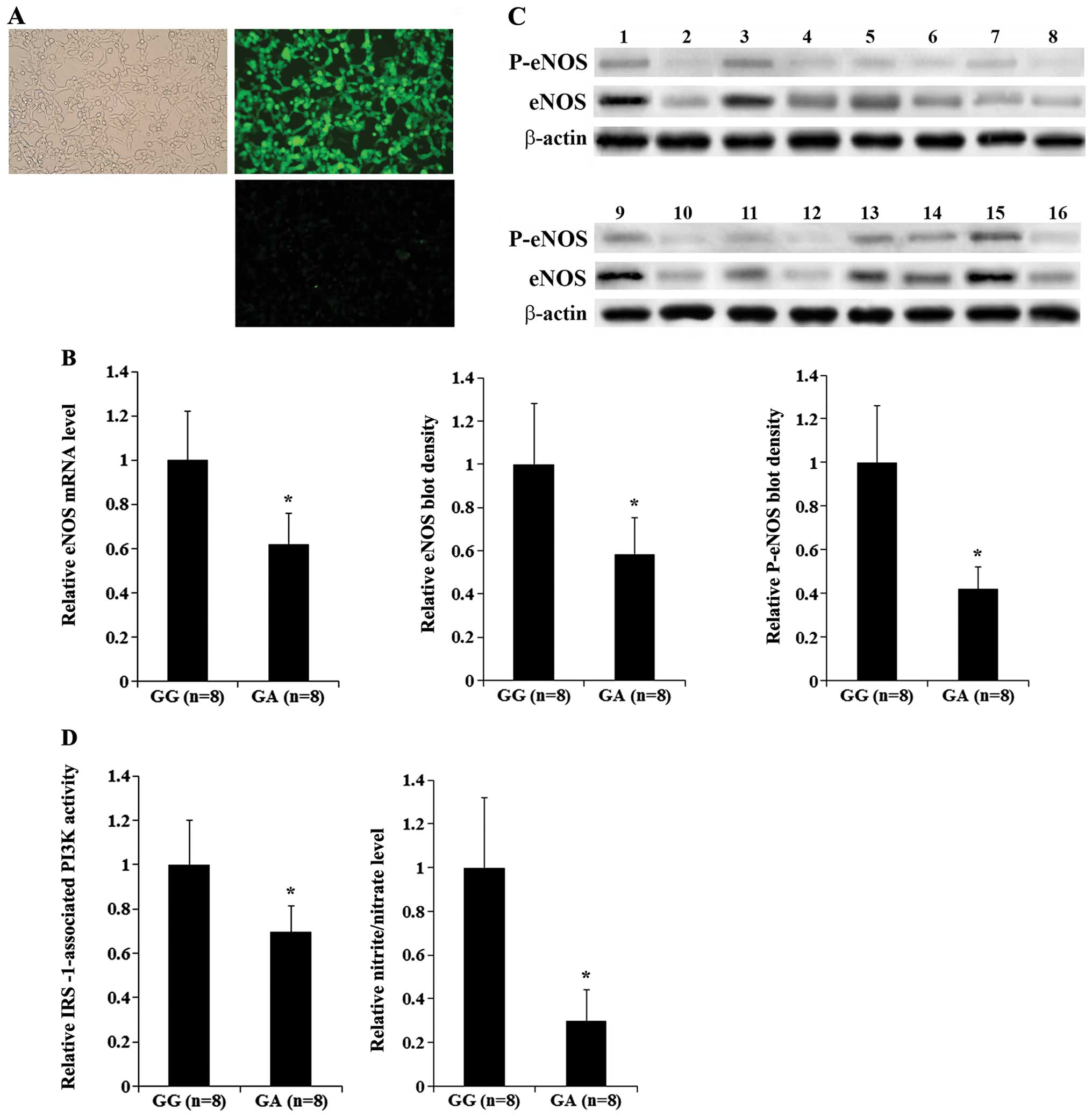 | Figure 1Endothelial nitric oxide synthase
(eNOS) expression and activity in human umbilical vein endothelial
cells (HUVECs) expressing homozygous wild-type (WT) insulin
receptor substrate-1 (IRS-1) or heterozygous Arg972
IRS-1. HUVECs were established from delivering mothers expressing
homozygous WT IRS-1 (GG group; n=8) or heterozygous
Arg972 IRS-1 (GA group; n=8) and stimulated with insulin
(10−8 M) for 24 h. (A) HUVEC cultures were stained for
von Willebrand factor (vWF), an endothelial cell marker. Phase
contrast Images of (left panel) and immunofluorescence staining for
vWF (upper right panel) of the same field of view are shown. A
negative staining control lacking the primary antibody for vWF is
included (lower right). (B) The eNOS mRNA level was determined by
RT-qPCR and shown as a fold change to that of the GG group
(designated as 1). (C) The eNOS protein level and eNOS
phosphorylation at serine 1177 (p-eNOS) were determined by western
blot analyses in the GG group (n=8, lanes 1, 3, 5, 7, 9, 11, 13 and
15) or the GA group (n=8, lanes 2, 4, 6, 8, 10, 12, 14 and 16).
β-actin was used as a loading control. Density of the eNOS blot was
normalized to that of β-actin to obtain a relative blot density.
The mean relative eNOS blot density of the GA group was expressed
as a fold change to that of the GG group (designated as 1). Density
of the p-eNOS blot was normalized to that of eNOS and β-actin to
obtain a relative blot density. The mean relative p-eNOS blot
density of the GA group was expressed as a fold change to that of
the GG group (designated as 1). (D) IRS-1-associated
phosphatidylinositol-3 kinase (PI3K) activities and nitrite/nitrate
levels in the GA group were expressed as fold changes to those of
the GG group (designated as 1). Data values are expressed as the
means + SD. *P<0.05 vs. GG. |
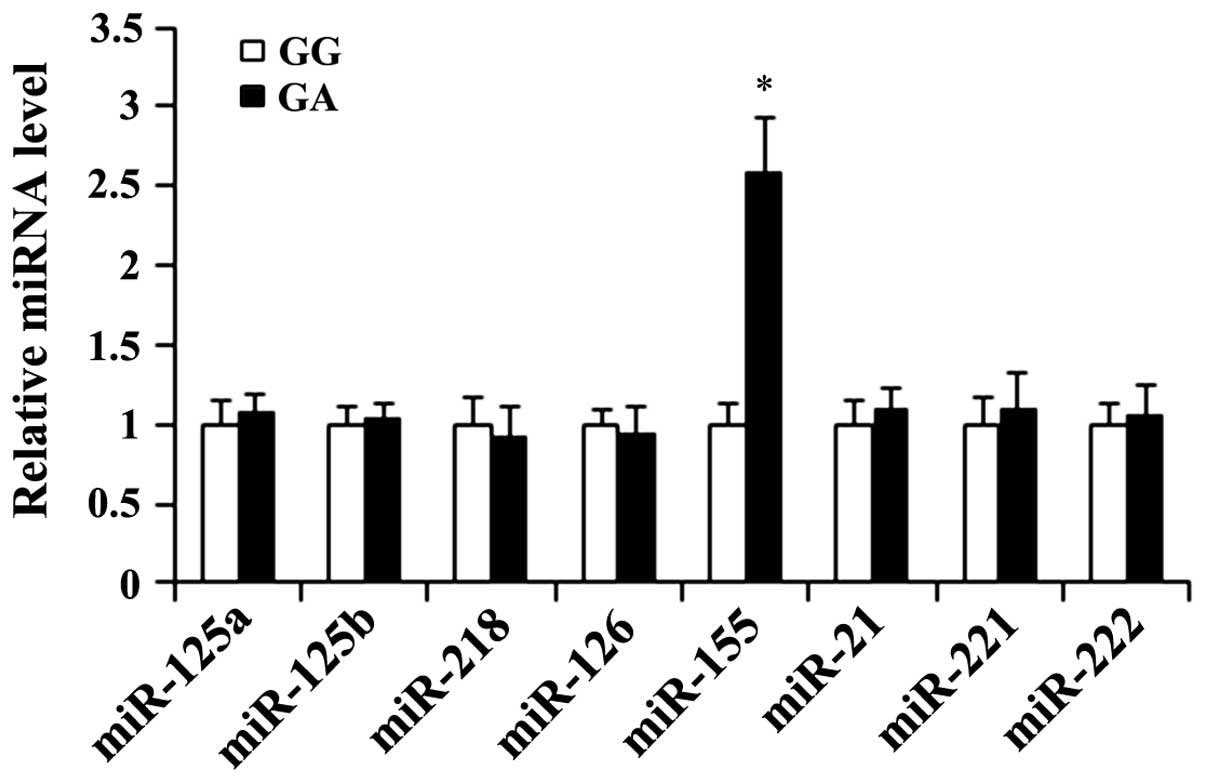
miRNA levels in HUVECs expressing
homozygous WT IRS-1 or heterozygous Arg972 IRS-1
A recent study demonstrated that eNOS expression can
be regulated by microRNAs (miRNAs or miRs) (27). Thus, in the HUVECs, we examined
the expression levels of miRNAs reportedly involved in regulating
vascular tone (miR-125a, miR-125b, miR-218, miR-126, miR-155 and
miR-21) (28) or eNOS expression
in endothelial cells (miR-155, miR-221 and miR-222) (27), particularly miR-155, which has
been shown to inhibit eNOS expression by directly targeting the
3′-untranslated region (3′-UTR) of eNOS mRNA (27). Following insulin stimulation, the
expression level of miR-155 in the GA (heterozygous
Arg972 IRS-1) group of HUVECs was increased
approximately 2.5-fold compared with that in the GG (homozygous WT
IRS-1) group of HUVECs, while there were no significant group
differences observed in the expression level of the other miRNAs
(Fig. 2).
eNOS expression/activity and eNOS
activation/phosphorylation in HUVECs overexpressing IRS-1 or
Arg972 IRS-1 in the presence or absence of miR-155 or
antagomir-155
All the above findings suggested that
Arg972 IRS-1 decreases eNOS expression and activity in
endothelial cells by upregulating miR-155. Thus, we examined eNOS
expression/activity and miR-155 expression levels in HUVECs stably
overexpressing IRS-1 or Arg972 IRS-1 in the presence or
absence of miR-155 or a functional inhibitor of miR-155
(antagomir-155). As shown in Fig.
3A, compared with the controls, the stable overexpression of
IRS-1 in the insulin-stimulated HUVECs decreased the miR-155
expression level by approximately 60%; this effect was reversed by
transfection with mature miR-155 mimic or by treatment with the
selective PI3K inhibitor, BKM120 (50 μM), for 60 min. On the
other hand, the stable overexpression of Arg972 IRS-1
increased the miR-155 expression level by 1.55-fold; this effect
was reversed by transfection with antagomir-155. Notably,
antagomir-155 decreased the miR-155 expression level, possibly by
inhibiting the quantitative RT-PCR reaction by competing with the
reverse transcription (RT) primer to bind miR-155 during reverse
transcription. As shown in Fig. 3B
and C, the overexpression of IRS-1 in the HUVECs increased eNOS
expression by approximately 1.5-fold; this effect was reversed by
transfection with mature miR-155 mimic or treatment with BKM120. On
the other hand, the overexpression of Arg972 IRS-1
decreased eNOS expression by approximately 50%, which was reversed
by transfection with antagomir-155. Transfection with mature
miR-155 mimic or treatment with BKM120 alone decreased eNOS
expression by approximately 80 and 70%, respectively, while
transfection with antagomir-155 alone increased eNOS expression by
over 1.6-fold. In addition, mature miR-155 mimic, antagomir-155 and
BKM120 showed no significant effect on IRS-1 expression in the
HUVECs (Fig. 3C), indicating that
their effects on eNOS expression are not mediated by altering the
expression of IRS-1. Taken together, these findings indicate that
Arg972 IRS-1 inhibits eNOS expression in endothelial
cells by upregulating miR-155. Of note, the overexpression of IRS-1
and Arg972 IRS-1 increased and decreased eNOS
phosphorylation at Ser1177 by 4.4-fold and 75%, respectively; these
effects were much more pronounced than their effects on eNOS
expression and were only partially reversed by miR-155 and
antagomir-155, respectively (Fig.
3C). By contrast, the effects of mature miR-155 mimic and
antagomir-155 on eNOS phosphorylation at Ser1177 were similar to
those on eNOS expression (Fig.
3C).
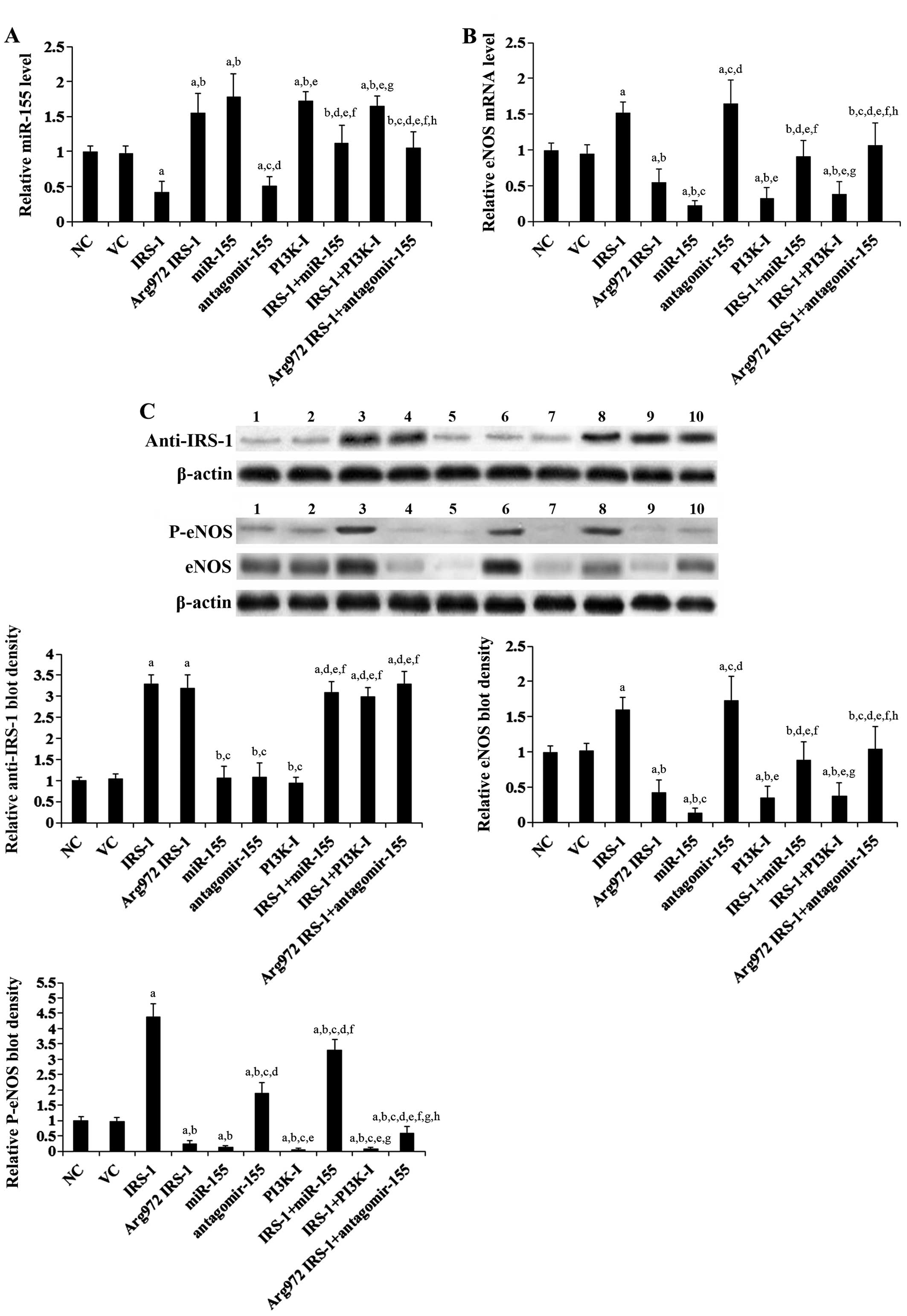 | Figure 3Endothelial nitric oxide synthase
(eNOS) and miR-155 levels in human umbilical vein endothelial cells
(HUVECs) overexpressing wild-type (WT) receptor substrate-1 (IRS-1)
or Arg972 IRS-1 in the presence or absence of the
overexpression or inhibition of miR-155. In HUVECs stimulated with
insulin (10−8 M) for 24 h, the expression of (A) miR-155
and (B) eNOS mRNA in normal control cells (NC), cells stably
transfected with empty pcDNA3 vector (VC), cells stably transfected
with IRS-1 with or without miR-155 treatment, cells stably
transfected with Arg972 IRS-1 with or without
antagomir-155 treatment, cells treated with selective
phosphatidylinositide 3-kinase (PI3K) inhibitor BKM120 (50
μM) (PI3K-I) for 60 min, and cells stably transfected with
IRS-1 plus BKM120 (50 μM) treatment was analyzed by RT-qPCR.
The miR-155 and the eNOS mRNA levels were expressed as fold changes
to those of NC (designated as 1), respectively. (C) In HUVECs
stimulated with insulin (10−8 M) for 24 h, western blot
analyses were performed to examine IRS-1/Arg972 IRS-1,
eNOS and phosphorylated eNOS (at serine 1177) (P-eNOS) levels in
normal control cells (NC, lane 1), cells stably transfected with
empty pcDNA3 vector (VC, lane 2), cells stably transfected with
IRS-1 (lane 3), cells stably transfected with Arg972
IRS-1 (lane 4), cells transfected with miR-155 (lane 5), cells
transfected with antagomir-155 (lane 6), cells treated with BKM120
(50 μM) for 60 min (PI3K-I, lane 7), cells stably
transfected with IRS-1 plus miR-155 treatment (IRS-1 + miR-155,
lane 8), cells stably transfected with IRS-1 plus BKM120 (50
μM) treatment (IRS-1 + PI3K-I, lane 9) and cells stably
transfected with Arg972 IRS-1 plus antagomir-155
treatment (Arg972 IRS-1 + antagomir-155, lane 10).
β-actin was used as a loading control. Density of the anti-IRS-1
and the eNOS blots was respectively normalized to that of β-actin
to obtain a relative blot density, which was expressed as a fold
change to the relative anti-IRS-1 blot density and the relative
eNOS blot density of NC (designated as 1), respectively. Density of
the p-eNOS blot was normalized to that of eNOS and β-actin to
obtain a relative blot density, which was expressed as a fold
change to the relative P-eNOS blot density of NC (designated as 1).
Three independent experiments were performed for each western blot
analysis. Data values are expressed as the means + SD.
aP<0.05 vs. NC or VC; bP<0.05 vs.
IRS-1; cP<0.05 vs. Arg972 IRS-1;
dP<0.05 vs. miR-155; eP<0.05 vs. antagomir-155;
fP<0.05 vs. PI3K-I; gP<0.05 vs. IRS-1 +
miR-155; hP<0.05 vs. IRS-1 + PI3K-I. |
Arg972 IRS-1 is reportedly associated
with impaired insulin/IRS-1 signaling to activate the PI3K/Akt
pathway, which is required to activate eNOS activity by
phosphorylating eNOS at Ser1177 (15). Thus, we examined IRS-1-associated
PI3K activity, Akt activation/phosphorylation and eNOS activity
(measured by the nitrite/nitrate level in the cell culture media)
in the HUVECs stably overexpressing IRS-1 or Arg972
IRS-1 in the presence or absence of miR-155 or antagomir-155.
IRS-1-associated PI3K activity, Akt
activation/phosphorylation and eNOS activity in HUVECs
overexpressing IRS-1 or Arg972 IRS-1 in the presence or
absence of miR-155 or antagomir-155
As shown in Fig.
4A, compared with the controls, the stable overexpression of
IRS-1 in the insulin-stimulated HUVECs increased the
IRS-1-associated PI3K activity by 2-fold, which was abolished by
treatment with BKM120, but not by transfection with mature miR-155
mimic. On the other hand, the stable overexpression of
Arg972 IRS-1 decreased the IRS-1-associated PI3K
activity by approximately 40%, which was not significantly altered
by transfection antagomir-155. Mature miR-155 mimic and
antagomir-155 showed no significant effects on the IRS-1-associated
PI3K activity. A similar data trend was observed with Akt
phosphorylation at serine 473 (ser473) (Fig. 4B), which is required for the
activation of Akt by PI3K (29).
As shown in Fig. 4C, the
overexpression of IRS-1 and Arg972 IRS-1 increased and
decreased the nitrite/nitrate level by 3.9-fold and 70%,
respectively; these effects were much more pronounced than their
effects on eNOS expression (Fig.
3C) and were only partially reversed by transfection with
miR-155 and antagomir-155, respectively. By contrast, the effects
of mature miR-155 mimic and antagomir-155 on the nitrite/nitrate
level were similar to those on eNOS expression (Fig. 3C). Taken together, our findings
indicate that under insulin stimulation, Arg972 IRS-1
inhibits eNOS activity by two different means: i) by downregulating
eNOS expression through the upregulation of miR-155; and ii) by
inhibiting eNOS activation through the decrease in the
PI3K-mediated phosphorylation of eNOS at Ser1177.
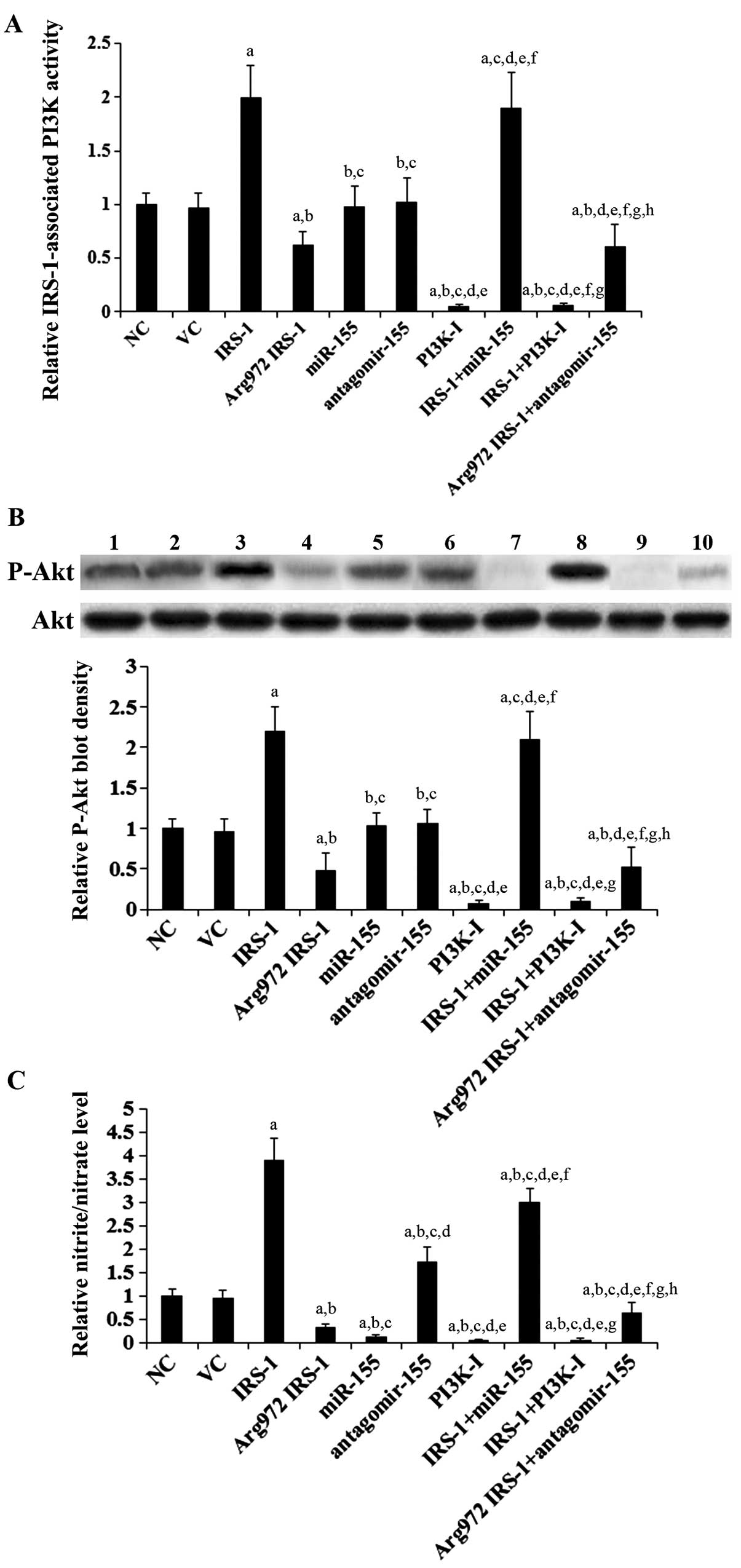 | Figure 4Insulin receptor substrate-1
(IRS-1)-associated phosphatidylinositide 3-kinase (PI3K) activity
and phosphorylated Akt (P-Akt) level in human umbilical vein
endothelial cells (HUVECs) overexpressing wild-type (WT) IRS-1 or
Arg972 IRS-1 in the presence or absence of the
overexpression or inhibition of miR-155. In HUVECs stimulated with
insulin (10−8 M) for 24 h, (A) IRS-1-associated PI3K
activity, (B) total Akt and p-Akt (at serine 473) levels and (C)
nitrite/nitrate levels in normal control cells (NC, lane 1), cells
stably transfected with empty pcDNA3 vector (VC, lane 2), cells
stably transfected with IRS-1 (lane 3), cells stably transfected
with Arg972 IRS-1 (lane 4), cells transfected with
miR-155 (lane 5), cells transfected with antagomir-155 (lane 6),
cells treated with selective PI3K inhibitor BKM120 (50 μM)
(PI3K-I, lane 7) for 60 min, cells stably transfected with IRS-1
plus miR-155 treatment (IRS-1 + miR-155, lane 8), cells stably
transfected with IRS-1 plus BKM120 (50 μM) treatment (IRS-1
+ PI3K-I, lane 9) and cells stably transfected with
Arg972 IRS-1 plus antagomir-155 treatment
(Arg972 IRS-1 + antagomir-155, lane 10) were determined.
In (B) western blot analyses, the density of the p-Akt blot was
normalized to that of total Akt to obtain a relative blot density,
which was expressed as a fold change to the relative p-Akt blot
density of NC (designated as 1). Three independent experiments were
performed for each western blot analysis. (A) IRS-1-associated PI3K
activity and (C) nitrite/nitrate levels were expressed as fold
changes to those of NC (designated as 1), respectively. Data values
are expressed as the means + SD. aP<0.05 vs. NC or
VC; bP<0.05 vs. IRS-1; cP<0.05 vs.
Arg972 IRS-1; dP<0.05 vs. miR-155;
eP<0.05 vs. antagomir-155; fP<0.05 vs.
PI3K-I; gP<0.05 vs. IRS-1 + miR-155;
hP<0.05 vs. IRS-1 + PI3K-I. |
Discussion
The dysregulation of NO synthesis attributable to
the abnormal expression/activity of eNOS is considered to be a
major feature of insulin-resistant states (30,31), as well as an essential contributor
to the pathogenesis of cardiovascular diseases (2,3).
Arg972 IRS-1 is reportedly associated with impaired
IRS-1 ability to activate PI3K, leading to insulin resistance
(18,19). In the present study, to the best
of our knowledge, we provide the first evidence that
Arg972 IRS-1 inhibits eNOS expression in human
endothelial cells by upregulating miR-155.
Multivariate regression analysis using data from 832
human subjects revealed that Arg972 IRS-1 was an
independent predictor of the human plasma eNOS level after
adjustment for multiple potential confounders, which was in
agreement with our results that heterozygous and homozygous
Arg972 IRS-1 carriers had significantly lower levels of
plasma eNOS and nitrite/nitrate than homozygous WT IRS-1 carriers.
This was corroborated by our findings that HUVECs derived from
delivering mothers expressing heterozygous Arg972 IRS-1
had significantly lower expression levels of eNOS and
nitrite/nitrate than those expressing homozygous WT IRS-1.
A recent study demonstrated that miR-155 inhibits
eNOS expression in human endothelial cells by directly targeting
the eNOS mRNA (27). In the
present study, the overexpression of IRS-1 and Arg972
IRS-1, respectively, upregulated and downregulated miR-155 in human
endothelial cells by altering PI3K signaling. This provides a
mechanistic explanation for the enhanced and inhibited eNOS
expression induced by the overexpression of IRS-1 and
Arg972 IRS-1, respectively. Since miR-155 and its
inhibitor, antagomir-155, respectively, reversed the effects of the
overexpression of IRS-1 and Arg972 IRS-1 on eNOS
expression, it can be concluded that Arg972 IRS-1
inhibits eNOS expression in endothelial cells by upregulating
miR-155. Although our findings indicate that Arg972
IRS-1 upregulates miR-155 in endothelial cells by impairing PI3K
signaling, the mechanisms through which PI3K signaling alters the
miR-155 level in endothelial cells remain unclear. We aim to
elaborate on this issue in our future studies.
We noted that compared with their effects on eNOS
expression, the overexpression of IRS-1 and Arg972 IRS-1
exerted much more pronounced effects on eNOS activity, which were
only partially reversed by miR-155 and antagomir-155, respectively.
This may be explained by the fact that Arg972 IRS-1
inhibited eNOS activity both by downregulating eNOS expression
through the upregulation of miR-155, and by inhibiting eNOS
activation through the impairment of the PI3K signaling-mediated
phosphorylation of eNOS (Ser1177), which is required to activate
eNOS (15).
Federici et al (22) reported that Arg972
IRS-1 inhibited eNOS expression in insulin-stimulated endothelial
cells in vitro. In the present study, we uncovered a
mechanism responsible for this phenomenon by demonstrating that
Arg972 IRS-1 inhibits eNOS expression through the
upregulation of miR-155 in endothelial cells. Moreover, our in
vivo data from a relatively large sample of human subjects
support the in vitro inhibitory effects of Arg972
IRS-1 on eNOS expression; this increases the generalizability of
our findings. In addition, our findings that Arg972
IRS-1 both downregulates eNOS expression and inhibits PI3K-mediated
eNOS activation/phosphorylation account for the marked inhibitory
effects of Arg972 IRS-1 on eNOS activity in
vitro. In view of the close association of eNOS activity/NO
production with cardiovascular diseases, such as hypertension and
atherosclerosis (2,3), our in vitro findings were
also in line with the data on human subjects showing that
heterozygous and homozygous Arg972 IRS-1 carriers had
significantly higher blood pressure and a prevalence of coronary
heart disease than homozygous WT IRS-1 carriers. Thus, the present
study provides both in vivo and in vitro evidence
supporting an important pathophysiological role for
Arg972 IRS-1 in cardiovascular diseases.
In conclusion, our in vivo data demonstrate
that Arg972 IRS-1 is associated with decreased plasma
eNOS and nitrite/nitrate levels in human subjects. Our in
vitro data demonstrate that Arg972 IRS-1 inhibits
eNOS expression in human endothelial cells by upregulating miR-155
through the impairment of PI3K signaling; additionally,
Arg972 IRS-1 exerts marked inhibitory effects on eNOS
activity by inhibiting both eNOS expression and eNOS
activation/phosphorylation through the impairment of PI3K
signaling. The present study provides new insight into the
pathophysiological role of Arg972 IRS-1 in
cardiovascular diseases.
Acknowledgments
The present study was supported by the Guangdong
Provincial Natural Science Foundation (grant nos. 12B423 and
13B175), Guangdong, P.R. China.
References
|
1
|
Schulz R, Rassaf T, Massion PB, Kelm M and
Balligand JL: Recent advances in the understanding of the role of
nitric oxide in cardiovascular homeostasis. Pharmacol Ther.
108:225–256. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Balligand JL, Feron O and Dessy C: eNOS
activation by physical forces: from short-term regulation of
contraction to chronic remodeling of cardiovascular tissues.
Physiol Rev. 89:481–534. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Förstermann U and Münzel T: Endothelial
nitric oxide synthase in vascular disease: from marvel to menace.
Circulation. 113:1708–1714. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Searles CD: Transcriptional and
posttranscriptional regulation of endothelial nitric oxide synthase
expression. Am J Physiol Cell Physiol. 291:C803–C816. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alonso J, Sánchez de Miguel L, Montón M,
Casado S and López-Farré A: Endothelial cytosolic proteins bind to
the 3′ untranslated region of endothelial nitric oxide synthase
mRNA: regulation by tumor necrosis factor alpha. Mol Cell Biol.
17:5719–5726. 1997.PubMed/NCBI
|
|
6
|
Lai PF, Mohamed F, Monge JC and Stewart
DJ: Downregulation of eNOS mRNA expression by TNFalpha:
identification and functional characterization of RNA-protein
interactions in the 3′ UTR. Cardiovasc Res. 59:160–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Laufs U and Liao JK: Post-transcriptional
regulation of endothelial nitric oxide synthase mRNA stability by
Rho GTPase. J Biol Chem. 273:24266–24271. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yan G, You B, Chen SP, Liao JK and Sun J:
Tumor necrosis factor-alpha downregulates endothelial nitric oxide
synthase mRNA stability via translation elongation factor 1-alpha
1. Circ Res. 103:591–597. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Alberti KG and Zimmet PZ: Definition,
diagnosis and classification of diabetes mellitus and its
complications. Diabet Med. 15:539–553. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang PL: eNOS, metabolic syndrome and
cardiovascular disease. Trends Endocrinol Metab. 20:295–302. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang PL: A comprehensive definition for
metabolic syndrome. Dis Models Mech. 2:231–237. 2009. View Article : Google Scholar
|
|
12
|
Hink U, Li H, Mollnau H, Oelze M, Matheis
E, Hartmann M, Skatchkov M, Thaiss F, Stahl RA, Warnholtz A, et al:
Mechanisms underlying endothelial dysfunction in diabetes mellitus.
Circ Res. 88:E14–E22. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Steinberg HO, Brechtel G, Johnson A,
Fineberg N and Baron AD: Insulin-mediated skeletal muscle
vasodilation is nitric oxide dependent. A novel action of insulin
to increase nitric oxide release. J Clin Invest. 94:1172–1179.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Scherrer U, Randin D, Vollenweider P,
Vollenweider L and Nicod P: Nitric oxide release accounts for
insulin’s vascular effects in humans. J Clin Invest. 94:2511–2515.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Montagnani M, Chen H, Barr VA and Quon MJ:
Insulin-stimulated activation of eNOS is independent of
Ca2+ but requires phosphorylation by Akt at
Ser1179. J Biol Chem. 276:30392–30398. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zeng G, Nystrom FH, Ravichandran LV, Cong
L, Kirby M, Mostowski H and Quon MJ: Roles for insulin receptor,
PI3-kinase, and Akt in insulin-signaling pathways related to
production of nitric oxide in human vascular endothelial cells.
Circulation. 101:1539–1545. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Montagnani M and Quon MJ: Insulin action
in vascular endothelium: potential mechanisms linking insulin
resistance with hypertension. Diabetes Obes Metab. 2:285–292. 2000.
View Article : Google Scholar
|
|
18
|
Fallucca F, Dalfrà MG, Sciullo E, Masin M,
Buongiorno AM, Napoli A, Fedele D and Lapolla A: Polymorphisms of
insulin receptor substrate 1 and beta3-adrenergic receptor genes in
gestational diabetes and normal pregnancy. Metabolism.
55:1451–1456. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hribal ML, Federici M, Porzio O, Lauro D,
Borboni P, Accili D, Lauro R and Sesti G: The Gly→Arg972
amino acid polymorphism in insulin receptor substrate-1 affects
glucose metabolism in skeletal muscle cells. J Clin Endocrinol
Metab. 85:2004–2013. 2000.PubMed/NCBI
|
|
20
|
Porzio O, Federici M, Hribal ML, Lauro D,
Accili D, Lauro R, Borboni P and Sesti G: The
Gly972>Arg amino acid polymorphism in IRS-1 impairs
insulin secretion in pancreatic beta cells. J Clin Invest.
104:357–364. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang C, Lin Z, Zhou Y, Fang M, Sun S,
Jiang W, Dong H, Lv B, Lan H, Chen M, et al: Arg972
insulin receptor substrate-1 is associated with elevated plasma
endothelin-1 level in hypertensives. J Hypertens. 30:1751–1757.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Federici M, Pandolfi A, De Filippis EA,
Pellegrini G, Menghini R, Lauro D, Cardellini M, Romano M, Sesti G,
Lauro R, et al: G972R IRS-1 variant impairs insulin regulation of
endothelial nitric oxide synthase in cultured human endothelial
cells. Circulation. 109:399–405. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu W, Zhou X, Yu F, Hu J and Hu W:
Arg972 Insulin receptor substrate-1 is associated with
decreased serum angiotensin-converting enzyme 2 levels in acute
myocardial infarction patients: in vivo and in vitro evidence.
Cardiovasc Diabetol. 12:1512013. View Article : Google Scholar
|
|
24
|
Nagoshi T, Matsui T, Aoyama T, Leri A,
Anversa P, Li L, Ogawa W, del Monte F, Gwathmey JK, Grazette L, et
al: PI3K rescues the detrimental effects of chronic Akt activation
in the heart during ischemia/reperfusion injury. J Clin Invest.
115:2128–2138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matsui T, Li L, del Monte F, Fukui Y,
Franke TF, Hajjar RJ and Rosenzweig A: Adenoviral gene transfer of
activated phosphatidylinositol 3′-kinaseand Akt inhibits apoptosis
of hypoxic cardiomyocytes in vitro. Circulation. 100:373–2379.
1999. View Article : Google Scholar
|
|
26
|
Polit DF and Sherman RE: Statistical power
in nursing research. Nurs Res. 39:365–369. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun HX, Zeng DY, Li RT, Pang RP, Yang H,
Hu YL, Zhang Q, Jiang Y, Huang LY, Tang YB, et al: Essential role
of microRNA-155 in regulating endothelium-dependent vasorelaxation
by targeting endothelial nitric oxide synthase. Hypertension.
60:1407–1414. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chamorro-Jorganes A, Araldi E and Suarez
Y: MicroRNAs as pharmacological targets in endothelial cell
function and dysfunction. Pharmacol Res. 75:15–27. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen L, Monti S, Juszczynski P, Ouyang J,
Chapuy B, Neuberg D, Doench JG, Bogusz AM, Habermann TM, Dogan A,
et al: SYK inhibition modulates distinct PI3K/Akt-dependent
survival pathways and cholesterol biosynthesis in diffuse large B
cell lymphomas. Cancer Cell. 23:826–838. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Calles-Escandon J and Cipolla M: Diabetes
and endothelial dysfunction: a clinical perspective. Endocr Rev.
22:36–52. 2001. View Article : Google Scholar : PubMed/NCBI
|