Introduction
Chronic renal failure (CRF) is considered the most
severe outcome of chronic kidney disease (CKD) and is defined by a
glomerular filtration rate (GFR) persistently below 15 ml/min/1.73
m2, and represents the end-stage of CKD requiring
treatments, such as dialysis or transplantation. Cardiovascular
disease and infection are two most frequent causes of death in
patients with CRF (1). There are
also other causes of death for such patients, which vary and these
include cancer, cachexia, death attributable to social factors and
other unknown causes (1–3). Kidney failure results in a decline
in renal function, as evidenced by neurohumoral and metabolic
abnormalities and the accumulation of damaging molecules, metabolic
acidosis, electrolyte abnormalities and volume overload. Large
observational databases have identified many hypothesis-generating
risk factors for mortality in CRF (1,4).
Despite some novel biomarkers which have been implicated in the
risk of mortality (5), their
effects on outcomes when used for therapeutic decisions have been
insufficiently identified.
Berberine (BBR), the major pharmacological
constituent of Coptis chinensis, is a type of isoquinoline
alkaloid used as a therapeutic agent in the treatment of cancer,
bacterial infections, diabetes, and cardiovascular and inflammatory
diseases (6–8). Furthermore, accumulating evidence
suggests that BBR can effectively inhibit cell proliferation and
induce apoptosis, and that it has antioxidant properties (9,10).
Therefore, the present study aimed to examine the effects of BBR on
the proliferation and apoptosis of mouse podocytes.
Tumor necrosis factor (TNF) receptor
(TNFR)-associated factors (TRAFs) were originally identified as
signal-transducing molecules for TNFR, but have also been linked to
downstream signaling via other receptors, such as interleukin
(IL)-1 receptor (11,12). To date, 7 members of the TRAF
family have been described. TRAF2, TRAF5 and TRAF6 have been
demonstrated to mediate the activation of nuclear factor-κB (NF-κB)
by interacting with the mitogen-activated protein kinase
(MAPK)/extracellular signal-regulated kinase (ERK1/2) pathway, the
TNF-β-activated kinase, or the atypical protein, thus being
implicated in the regulation of cell death and cellular responses
to stress (13,14). The TNF-α-induced activation of
NF-κB has been shown to be severely inhibited in murine embryonic
fibroblasts (MEFs) derived from TRAF5 knockout mice (15). Moreover, it has been reported that
the TRAF5-induced activation of NF-κB is involved in glioma cell
migration and invasion (16).
However, whether the TRAF5-induced NF-κB activation is involved in
proliferation and apoptosis remains unknown.
In the present study, we examined the effects of BBR
on mouse podocyte viability and apoptosis. We found that BBR
prevented the induction of cell apoptosis induced by TRAF5
overexpression in mouse podocytes by suppressing NF-κB activation.
Therefore, our results suggest that BBR plays an important role in
the proliferation and apoptosis of mouse podocytes, and thus TRAF5
may be a potential therapeutic target in CKD.
Materials and methods
Bioinformatics analysis
The array expression data of TRAF5 for 13 patients
with CKD and 12 healthy controls were downloaded from the NCBI Gene
Expression omnibus (GEO, http://www.ncbi.nlm.nih.gov/geo/) and are accessible
through GEO Series accession no. GSE48944 (17), following the approval of this
project by the consortium.
Patient samples
Serum samples were obtained from 30 patients with
CRF and 30 healthy controls admitted to Shuguang Hospital,
Shanghai, China. Ethical approval for the study was provided by the
independent ethics committee, Shuguang Hospital of Shanghai
university of Traditional Chinese Medicine. Written informed
consent was obtained from all participants in this study. None of
these patients had received radiotherapy or chemotherapy prior to
obtaining the samples.
Cell culture
Mouse podocytes were obtained from the the BeNa
Culture Collection (cat. no. BNCC100668; Beijing, China) and
cultured in RMPI-1640 supplemented with 10% fetal bovine serum, 10%
penicillin-streptomycin solution and 10 u/ml interferon-γ (IFN-γ),
and incubated in a humidified atmosphere at 33°C with 5%
CO2. Following culture for a period of time, the
podocytes were cultured in the above-mentioned medium without 10
u/ml IFN-γ and incubated in a humidified atmosphere at 37°C with 5%
CO2 for 10–14 days.
Induction of TRAF5 overexpression in
mouse podocytes
pLV-IRES-eGFP, psPAX2 and pMD2G were obtained from
Addgene (Cambridge, MA, USA). Commercial TRAF5 expression vectors
were obtained from Genewiz Biotechnology (Suzhou, China). The TRAF5
expression sequence was cloned into the pLV-IRES-eGFP lentiviral
vector. The blank pLV-IRES-eGFP lentiviral vector used as the
negative control (NC). 293T cells (ATCC, Manassas, VA, USA) were
seeded in 60 mm culture dishes, and after 24 h, they were
co-transfected with 2 μg of the plasmid vector, 1 μg
pLV-IRES-eGFP-TRAF5/ pLV-IRES-eGFP, 0.1 μg psPAX2 and 0.9
μg pMD2G using Lipofectamine 2000 (Invitrogen, Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
instructions. The recombinant lentivirus was collected 48 h after
transfection and used to infect the mouse podocytes.
Cell treatment
Following the induction of TRAF5 overexpres-sion in
mouse podocytes, the mouse podocytes were treated with various
concentrations of BBR (10, 30 and 90 μM; Sigma-Aldrich, St.
Louis, Mo, USA) and cell viability was measured by CCK-8 assay to
obtain the optimal concentration of BBR. To examine the effects of
BBR and NF-κB on cell viability, apoptosis and related protein
expression, the mouse podocytes were treated with 30 μM BBR
or 25 μg/ml of the NF-κB inhibitor, caffeic acid phenethyl
ester (CAPE; Selleck, Shanghai, China) for 0, 24, 48 and 72 h (for
cell viability assay) or for 48 h (for cell apoptosis assay). To
examine the effects of TNF-α, IL-6 and LPS on cell viability,
apoptosis and related protein expression, the mouse podocytes were
treated with 100 ng/ml TNF-α (PeproTech, Rocky Hill, NJ, USA), 100
u/ml IL-6 (PeproTech) or 100 ng/ml LPS (Sigma-Aldrich) for 0, 24,
48 and 72 h (for cell viability assay) or for 48 h (for cell
apoptosis assay) in the absence or presence of 30 μM BBR
treatment.
Reverse transcription and real-time
PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen, Life Technologies, Carlsbad, CA, USA) as previously
described (18). Complementary
DNA was synthesized using a cDNA synthesis kit (Thermo Fisher
Scientific Inc.). Real-time PCR was performed using a standard
SYBR-Green PCR kit (Takara Biotechnology Co., Ltd., Dalian, China)
and an ABI 7500 (Applied Biosystem Life Technologies, Foster City,
CA, USA) thermal cycler. The primers used were as follows: TRAF5
forward, 5′-CACTCCGTGCTTCACAAC-3′ and reverse,
5′-AAGGTGGTCCTGGAATCG-3′; glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) forward, 5′-ATCACTGCCACCCAGAAG-3′ and reverse,
5′-TCCACGACGGACACATTG-3′. GAPDH was used an internal control for
normalization. The gene expression was calculated using the
2−ΔΔCq method, as previously described (19).
Cell counting kit-8 (CCK-8) assay
Mouse podocytes infected with pLV-IRES-eGFP-TRAF5
(3×103/well) were plated in 96-well plates. Following
treatment as indicated for 0, 24, 48 and 72 h, 10% of CCK-8
solution (Dojindo Molecular Technologies, Kumamoto, Japan) diluted
in serum-free RMPI-1640 was mixed in each well for a further 1 h.
The optical density 450 nm value in each well was determined by a
micro-plate reader (SM600 Labsystem; Shanghai utrao Medical
Instrument Co., Ltd., Shanghai, China).
Apoptosis assay
Mouse podocytes infected with pLV-IRES-eGFP-TRAF5
infection (5×105/well) were plated in 6-well plates.
Follownig treatment as indicated for 48 h, the mouse podocytes were
collected and incubated with 195 μl Annexin V-fluorescein
isothiocyanate (FITC) and 5 μl propidium iodide (PI) for 15
min in the dark at 4°C, prior to analysis by flow cytometry (BD
Biosciences, Franklin Lakes, NJ, USA)
Western blot analysis
Mouse podocytes were seeded at a density of
5×105 cells/well in 6-well plates, cultured overnight
and then treated as indicated for 3 or 48 h. Total proteins were
isolated from the mouse podocytes and were subjected to 12%
glyceraldehyde 3-phosphate dehydrogenase (SDS-PAGE) and
electroblotted onto onto polyvinylidene fluoride membranes (Roche
Diagnostics, Mannheim, Germany). The membranes were first incubated
with rabbit polyclonal anti-Bax (1:300; Sc-493, Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-caspase-3 (1:500;
ab44976, Abcam, Cambridge, MA, USA) and anti-nephrin (1:500;
ab58968, Abcam) antibodies; rabbit monoclonal anti-TRAF5 (1:1,000;
ab137763, Abcam), anti-p-NF-κB p65 (1:1,000; #3033), anti-NF-κB p65
(1:1,000; #8242) (both from Cell Signaling Technology, Danvers, MA,
USA), anti-podocin (1:10,000; ab181143, Abcam) and anti-GAPDH
(1:1500; #5174; Cell Signaling Technology) antibodies; and mouse
monoclonal anti-Bcl-2 (1:400; ab117115, Abcam) antibody. The blots
were then incubated with goat anti-mouse or anti-rabbit secondary
antibody (1:1,000; A0208 and A0216, Beyotime Institute of
Biotechnology, Haimen, China) and visualized using enhanced
chemiluminescence (ECL; Thermo Fisher Scientific). GAPDH antibody
was used as an internal control. The blotting bands were quantified
with ImageJ software (National Institutes of Health, Bethesda, MD,
USA).
Statistical analysis
All data are expressed as the means ± SD and
representative of experiments were carried out in triplicate
analyzed with SPSS 19.0 software (SPSS Inc., Chicago, IL, USA). The
unpaired, two-tailed Student's t-test and ANOVA followed by Tukey's
post hoc test were used to analyze the significance of differences
between groups. Differences were considered significant if the
probability (P)-value was <0.05.
Results
TRAF5 levels in the peripheral blood of
patients with CKD and CRF
In order to examine the role of TRAF5 in kidney
disease, we first measured the levels of TRAF5 in the peripheral
blood of patients with CKD (n=13) and healthy controls (n=12) using
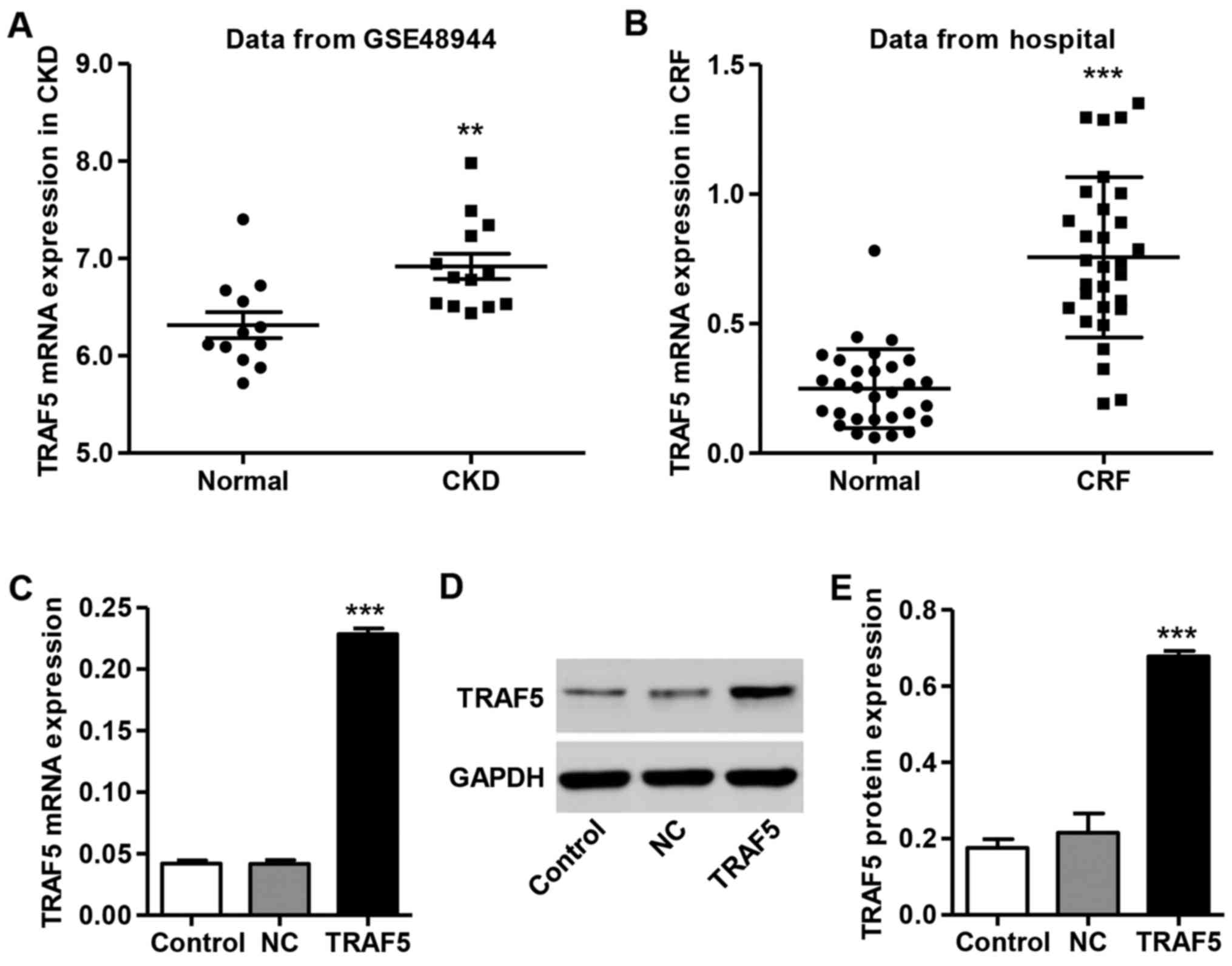
the data from the GSE48944 database. As shown in Fig. 1, TRAF5 mRNA expression was
significantly increased in patients with CKD compared with the
healthy controls. Furthermore, we also detected the levels of TRAF5
in the peripheral blood of patients with CRF (n=30) and healthy
controls (n=30) from the Shuguang Hospital database. As shown in
Fig. 1B, similar to the data from
the GSE48944 database, the mRNA expression of TRAF5 was
significantly increased in patients with CRF (average, 0.756 and
median, 0.726) compared with the healthy controls (average, 0.249
and median, 0.244).
To further examine the effects of TRAF5 on kidney
function in vitro, mouse podocytes were infected with the
TRAF5 overexpressionvector pLV-IRES-eGFP-TRAF5. As shown in
Fig. 1C–E, the expression of
TRAF5 was markedly increased at both the mRNA and protein level in
the mouse podocytes infected with pLV-IRES-eGFP-TRAF5 compared with
the controls. However, the podocytes infected wth the blank
pLV-IRES-eGFP (NC) vector exhibited no changes in TRAF5
expression.
TRAF5 overexpression inhibits cell
viability and induces the apoptosis of mouse podocytes
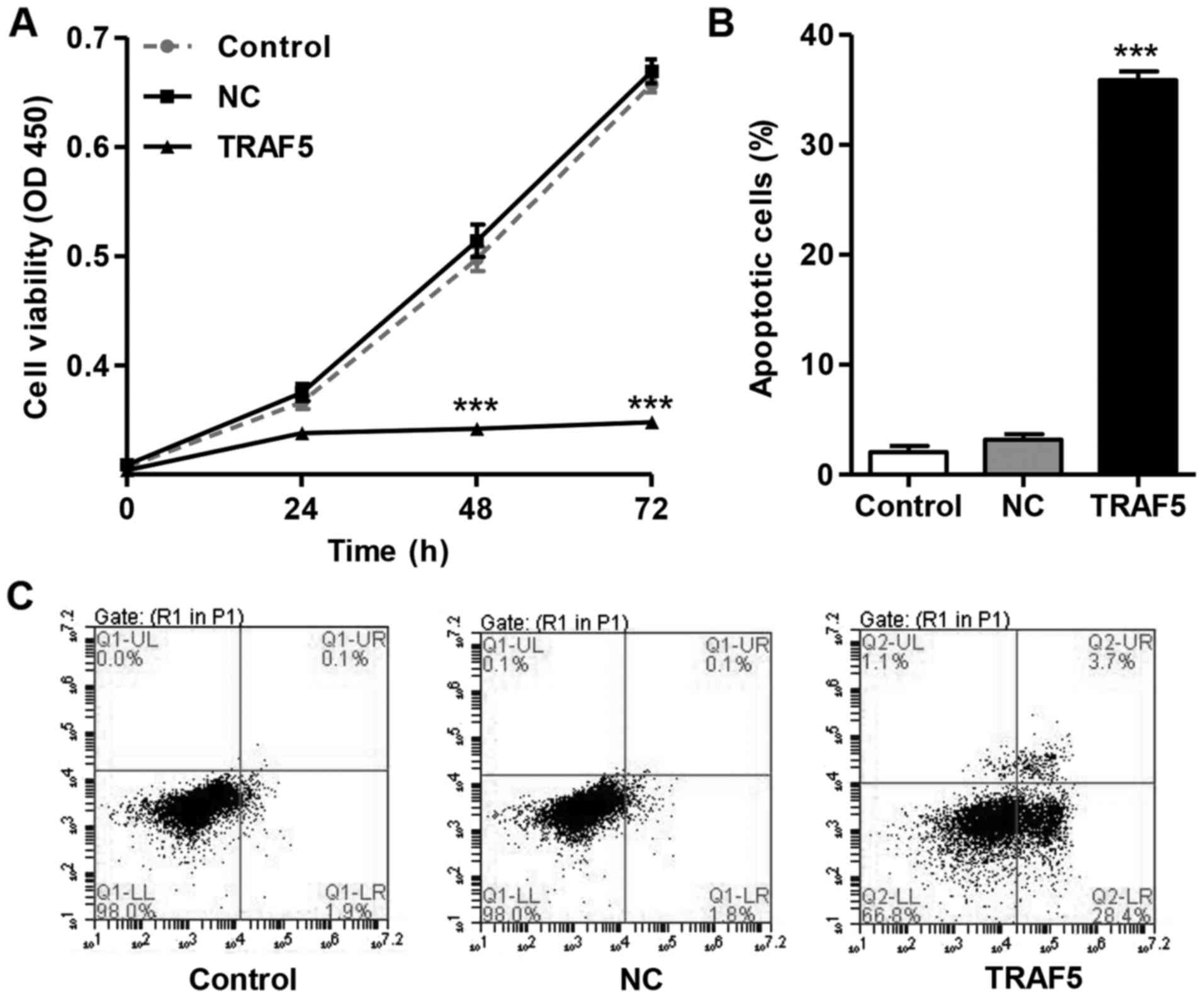
Following infection with pLV-IRES-eGFP-TRAF5, the
mouse podocytes exhibited a significant decrease in cell viability
in a time-dependent manner (Fig.
2A). After 72 h of incubation, the viability of the mouse
podocytes infected with the TRAF5 overexpression vector was
suppressed by 47.03±0.11% compared with the control group. However,
the mouse podocytes infected with the blank pLV-IRES-eGFP (NC)
vector exhibited no change in viability compared with the control.
Furthermore, we also investigated the role of TRAF5 in the
apoptosis of mouse podocytes. As shown in Fig. 2B and C, infection with the
pLV-IRES-eGFP-TRAF5 vector increased the apoptosis (35.9±0.8%) of
mouse podocytes compared with the control group (2.05±0.6%).
However, infection of the mouse podocytes with the blank
pLV-IRES-eGFP (NC) did not affect cell apoptosis compared with the
control. These results indicate that TRAF5 overexpression is
implicated in the inhibition of the viability and the apoptosis of
mouse podocytes.
BBR suppresses TRAF5 expression and
prevents the inhibition of cell viability induced by TRAF5
Considering the role of BBR in renal injury in
experimental rats (20), and as
the the mechanisms underlying the effects of BBR are not yet fully
understood, we wished to determine whether BBR also possesses a
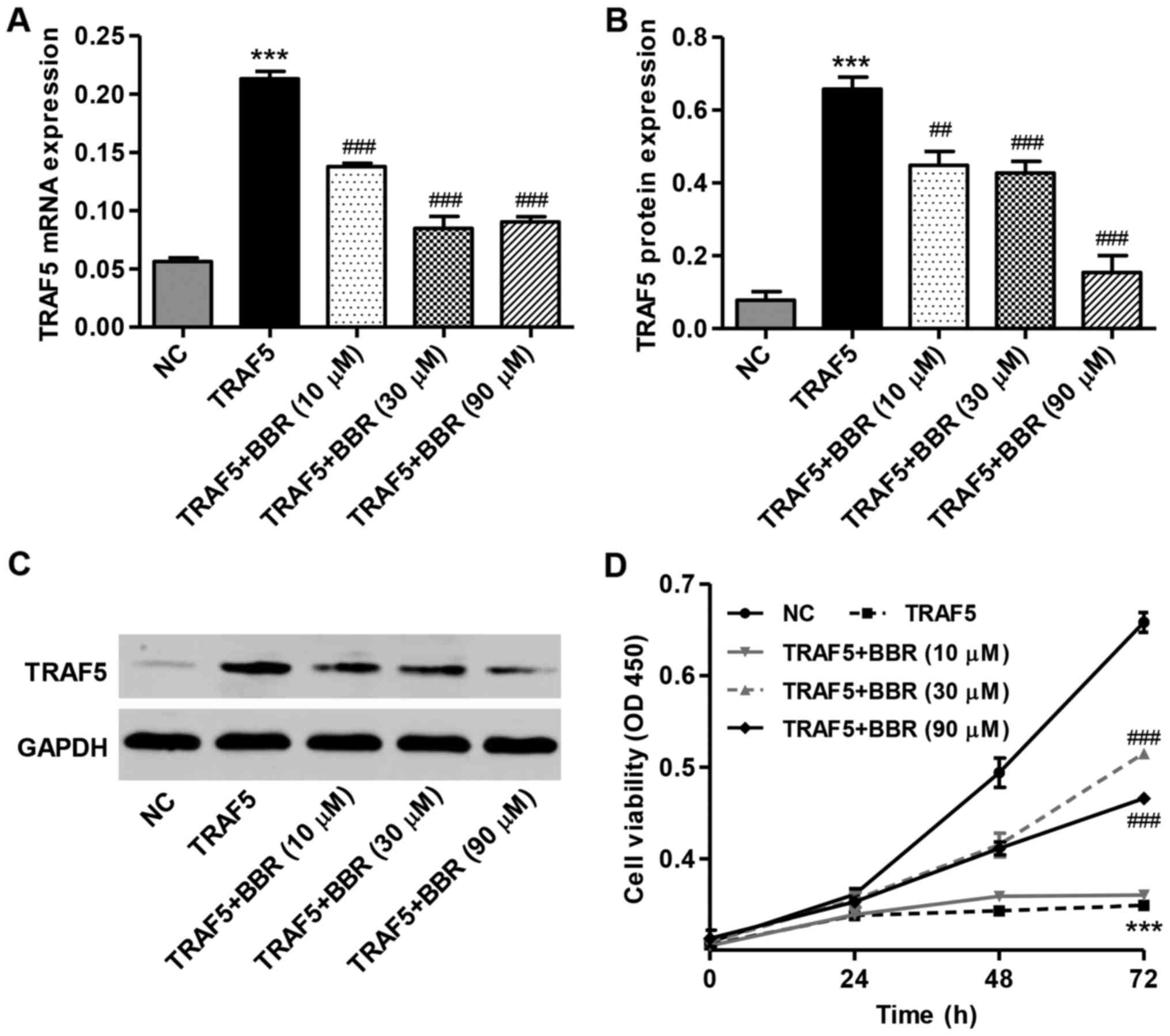
exerts an effect on TRAF5 overexpression in podocytes. As shown in
Fig. 3A, BBR (10, 30 and 90
μM) treatment significantly decreased the expression of
TRAF5 at both the mRNA (Fig. 3A)
and protein level (Fig. 3B and C)
compared with the mouse podocytes infected with the TRAF5
overexpression vector and not treated with BBR. Compared with the
podocytes infected with the TRAF5 overexpression vector and not
treated with BBR, treatment of the mouse podocytes with various
concentrations of BBR for 48 and 72 h increased cell viability in a
time-dependent manner (Fig. 3D).
After 72 h of incubation, the viability of the mouse podocytes
treated with with BBR (30 and 90 μM) was increased by
47.5±2.6 and 33.5±1.2%, respectively compared with the podocytes
infected with the TRAF5 overexpression vector and not treated with
BBR. These findings suggest that the downregulation of TRAF5 is
involved in the BBR-induced increase in podocyte viability.
BBR suppresses the NF-κB activation
induced by TRAF5 overexpression
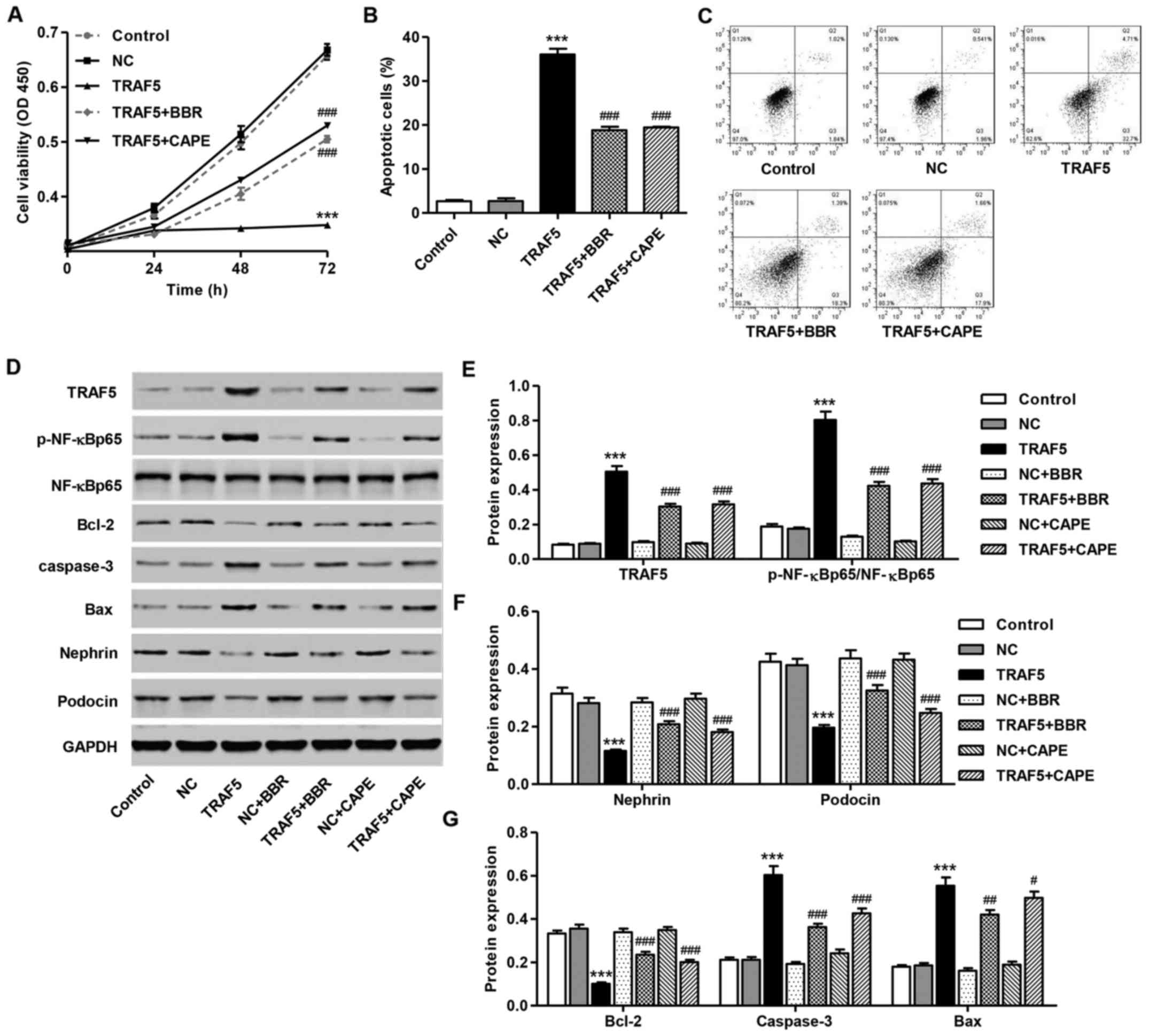
Following treatment with 30 μM BBR or 25
μg/ml CAPE for 72 h, the viability of the mouse podocytes
was significantly increased by 47.9±2.8 and 52.3±3.5%, respectively
compared with the untreated podocytes infected with the TRAF5
overexpression vector (Fig. 4A).
Treatment with BBR or CAPE treatment for 48 h also decreased the
apoptosis of the mouse podocytes by 47.8±3.9 and 46.1±2.5%,
respectively compared with the untreated podocytes infected with
the TRAF5 overexpression vector (Fig.
4B and C).
To clarify the effects of TRAF5 on NF-κB p65
activation in vitro, western blot analysis was performed. As
shown in Fig. 4D and E, the ratio
of p-NF-κB p65/NF-κB p65 was significantly decreased following
treatment with 30 μM BBR compared with the untreated
podocytes infected with the TRAF5 overexpression vector. Similarly,
CAPE (25 μg/ml), a potent and specific inhibitor of the
activation of NF-κB, mimicked the suppressive effects of BBR on
TRAF5 overexpression and NF-κB p65 activation in mouse podocytes.
These data demonstrate that the inactivation of NF-κB may
contribute to the BBR-induced protective effects on mouse
podocytes.
Effect of BBR and CAPE on protein
expression induced by TRAF5 overexpression
Following the infection of the mouse podocytes with
pLV-IRES-eGFP-TRAF5 for 48 h, the expression of nephrin and podocin
was significantly suppressed (Fig. 4D
and F), while the Bax/Bcl-2 ratio and caspase-3 levels were
significantly increased compared with the control group (Fig. 4D and G). However, treatment with
30 μM BBR or 25 μg/ml CAPE for 48 h suppressed the
effects induced by TRAF5 overexpression on these protein expression
levels in mouse podocytes (Fig.
4D–G).
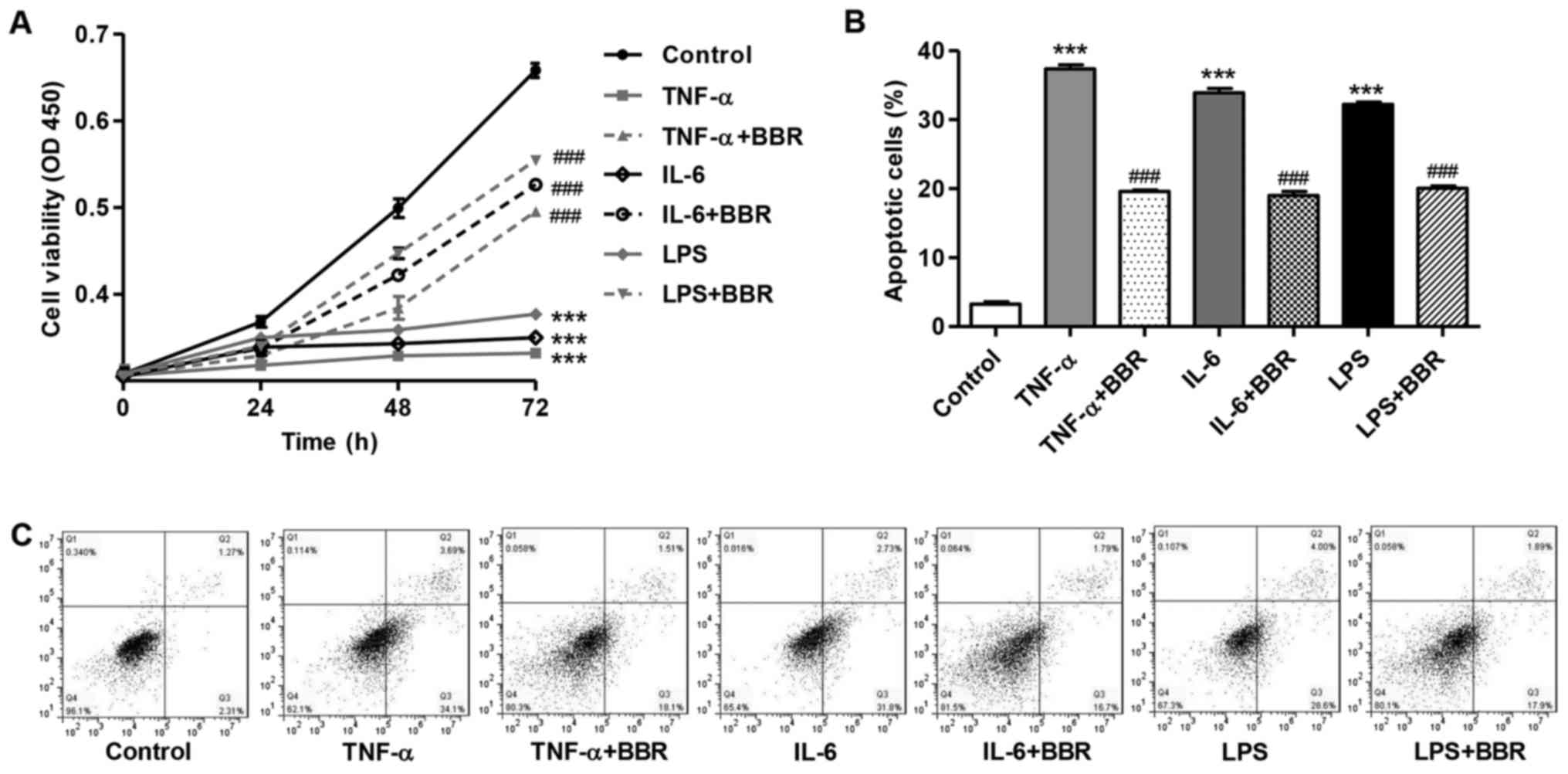
BBR prevents the inhibition of cell
viability and the apoptosis induced by TNF-α, IL-6 and LPS
Treatment with TNF-α (100 ng/ml), IL-6 (100 u/ml) or
LPS (100 ng/ml) for 72 h significantly decreased cell viability by
48.0±3.2, 46.8±2.7 and 45.1±2.9%, respectively compared with the
control group (Fig. 5A). However,
treatment with 30 μM BBR markedly prevented the inhibition
of cell viability induced by TNF-α, IL-6 or LPS in mouse podocytes.
Moreover, treatment with 30 μM BBR also suppressed the
apoptosis induced by TNF-α, IL-6 or LPS by 47.6±2.6, 43.9±3.3 and
37.7±1.9%, respectively compared with the cells exposed to TNF-α,
IL-6 or LPS (Fig. 5B and C).
These data suggest that BBR inhibits TNF-α-, IL-6- or LPS-induced
cytotoxicity in mouse podocytes.
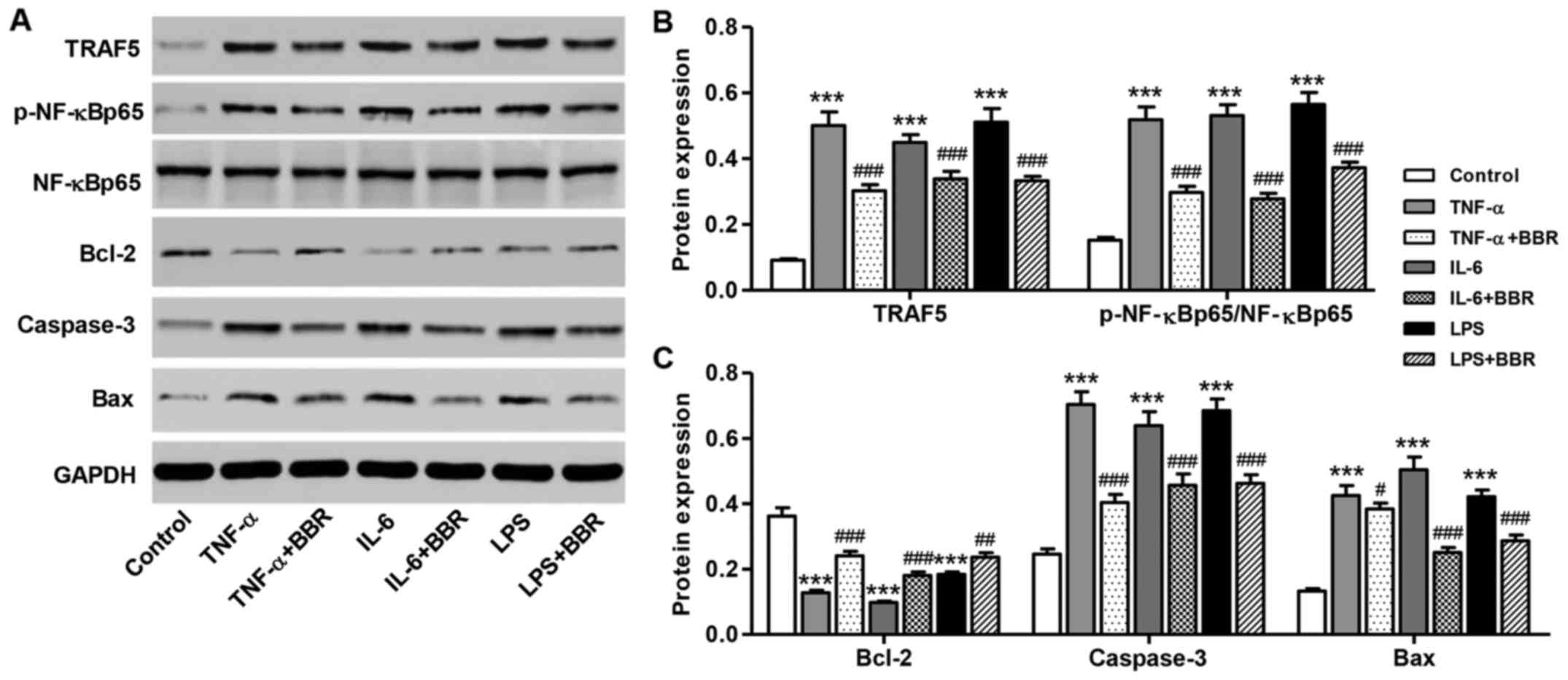
Effect of BBR on NF-κB activation and
protein expressions induced by TNF-α, IL-6 and LPS
As shown in Fig.
6, the ratio of p-NF-κB p65/NF-κB p65 and Bax/Bcl-2 and the
expression levels of TRAF5 and caspase-3 were significantly
increased by TNF-α, IL-6 or LPS treatment compared with the
controls. However, treatment with 30 μM BBR markedly
suppressed the effects of TNF-α, IL-6 or LPS on NF-κB activation
and on the expression levels of these proteins in mouse podocytes.
These results indicate that TRAF5 downregulation is implicated in
the protective effects of BBR against the effects of TNF-α, IL-6 or
LPS in mouse podocytes.
Discussion
This study reports the novel finding that TRAF5
expression was increased in the peripheral blood of patients with
CKD and CRF. In vitro experiments revealed that TRAF5
overexpression inhibited the viability and induced the apoptosis of
mouse podocytes. We further demonstrated that BBR inhibited the
negative effects of TRAF5 overexpression by suppressing NF-κB
activation in mouse podocytes. These data provide novel evidence
(at least to the best of our knowledge) that BBR protects mouse
podocytes from the suppressive effects of TRAF5 on viability and
the promoting effects of TRAF5 on apoptosis via the NF-κB signaling
pathway.
It has previously been reported that soluble TRAF5
levels are increased in plasma and peripheral blood mononuclear
cells from patients with Crohn's disease and ulcerative colitis
(21). Other studies have
implicated TRAF5 in carcinogenesis attributable to increased levels
in splenic marginal zone lymphoma (22) and Hodgkin-Reed-Sternberg cells
(23). Intriguingly, the
TRAF5/GAPDH mRNA ratios have been shown to be significantly
decreased in the blood of patients with chronic and acute coronary
heart disease, supporting the notion that TRAF5 represents a
protective marker in atherosclerosis (24). However, the expression of TRAF5 in
patients with CKD has not yet been elucidated. To examine the
hypothesis that TRAF5 may also be associated with clinical disease,
we performed bioinformatics analysis and a pilot clinical study in
patients with CKD and CRF. This study revealed increased peripheral
blood levels of TRAF5 in patients with CKD and CRF compared with
the healthy controls. As reported previously, TRAF5 knockout had no
effect on the viability of MEFs, while MEFs from double TRAF2 and
TRAF5 knockout mice exhibited significantly decreased cell
viability compared with wild-type and single TRAF2 or TRAF5
knockout mice, suggesting a critical role of TRAF2 rather than
TRAF5 in protection from cell death (15). However, as demonstrated in another
study, the elimination of TRAF5 expression significantly decreased
the migration and invasion of the glioma cells, and although the
underlying mechanisms were not elucidated, this may have bene due
to the inhibition of cell viability and apoptosis induction
(16). In the present study, to
examine whether the podocyte apoptosis was mediated via TRAF5
overexpression, we infected mouse podocyte with a TRAF5
overexpression vector. We found that TRAF5 overexpression
significantly induced mouse podocyte apoptosis.
TRAF5 was originally identified as an activator of
interleukin-induced NF-κB signal transduction via its TRAF domain.
NF-κB comprises a family of transcription factors involved in the
regulation of a wide variety of biological responses. It is
generally accepted that NF-κB activation is responsible for
apoptosis resistance (25,26).
However, there is evidence to support a pro-apoptotic role for
NF-κB. It has been speculated that NF-κB may have a dual function,
either as an inhibitor or an activator of apoptotic cell death,
depending on the levels of RelA and c-Rel (27). In this study, TRAF5 overexpression
in mouse podocytes led to NF-κB activation, accompanied by an
increased expression of caspase-3 and an increased Bax/Bcl-2 ratio.
Within cells, there is a machinery consisting of pro-apoptotic
genes (Bax) that promote apoptosis and anti-apoptotic genes (Bcl-2)
that function as suppressors of apoptosis, and the balance between
these genes may be a determinant of apoptosis or cell survival.
Despite extensive studies in either field, there is only limited
information on the role of Bcl-2 and Bax in CKD. In vivo,
Bcl-2 and Bax proteins have not been detected in the kidneys during
ischemia (28), whereas the
overexpression of Bcl-2 can suppress the apoptosis of renal tubule
cells induced by hypoxia/reoxygenation (29). Moreover, an increase in the
Bax/Bcl-2 ratio by hypoxia/reoxygenation or ischemia/reperfusion
injury triggers Bax translocation to the mitochondria and
cytochrome c release to cytoplasm, and enhances
caspase-3-mediated renal tubular apoptosis (30). Podocyte damage is a common feature
in glomerular diseases with ultrastructural changes, with the
reduced expression of components of the slit diaphragm, such as
nephrin and podocin (31). The
levels of nephrin and podocin have been shown to be significantly
reduced in lupus nephritis, with these effects beginning from the
earlier stages and becoming more pronounced at advanced
histological forms (32). Our
data reported that the expression of nephrin and podocin was
significantly decreased in mouse podocytes overexpressing
TRAF5.
NF-κB is activated by inflammatory cytokines and
cellular stresses, including TNF, IL-1, LPS, UV or γ-irradiation.
Thus far, TRAF5-deficient mice do not show substantial defects in
TNF-α-induced NF-κB activation, suggesting that TRAF5 plays a
redundant role in TNF-α-induced NF-κB activation (15). By contrast, TRAF5 acts downstream
of ILs, including IL-1β and IL-6, and plays a key role in
IL-1β/IL-6-mediated NF-κB activation during glioma migration and
invasion (16). In the present
study, we observed that the exposure of mouse podocytes to TNF-α,
IL-6 and LPS significant decreased cell viability and induced
apoptosis. More importantly, the activation of NF-κB and the
increased Bax/Bcl-2 ratio and caspase-3 expression were also
observed in the podocytes exposed to TNF-α, IL-6 and LPS. These
findings suggest that TRAF5 induces podocyte injury, to a certain
extent, through NF-κB activation induced by TNF-α, IL-6 and
LPS.
Emerging evidence has indicated that BBR has
multiple beneficial effects in the treatment of diabetes and
cardiovascular diseases (33,34). However, the protective effects of
BBR and its molecular mechanisms of action in CKD and chronic
kidney injury remain to be determined. BBR has been shown to
attenuate renal injury in diabetic C57BL/6 mice through the
suppression of the SphK-S1P signaling pathway (35). Additionally, BBR has been shown to
exert protective effects in the presence of high glucose related to
the inhibition of glucose-induced apoptosis that in turn
upregulates the expression of nephrin and podocin (36). In agreement with the previous
study, our results demonstrated that BBR increases cell viability
and inhibits apoptosis by upregulating the expression of nephrin
and podocin, and downregulating the expression of caspase-3 and the
Bax/Bcl-2 ratio. The NF-κB inhibitor, CAPE, mimicked the protective
effecs of BBR. Moreover, the NF-κB activation induced by TRAF5
overexpression and exposure to TNF-α, IL-6 or LPS was significantly
inhibited by BBR, which is in line with the findings of a previous
study that BBR ameliorates intrarenal inflammation and
tubulointerstitial injury, at least in part, through the
suppression of the NF-κB signaling pathway (37).
In conclusion, in this study, we demonstrate that
TRAF5 is overexpressed in CRF and inhibits the viability and
induces the apoptosis of mouse podocytes. Treatment of
TRAF5-overexpressing mouse podocytes with BBR suppressed the
inhibition of viability, prevented apoptosis, decreased the Bax/
Bcl-2 and caspase-3 expression, and increased the expression of
nephrin and podocin. Such effects appear to be mediated by the
inhibition of NF-κB activation. Thus, BBR may play an important
role in delaying the progression of chronic kidney injury by
preserving renal structure and function in patients with CRF.
Acknowledgments
This study was supported by the Shanghai Three-Year
Project of Traditional Chinese Medicine (nos. ZY3-JSFC-2-1026 and
ZY3-CCCX-2-1003).
References
|
1
|
Ortiz A, Covic A, Fliser D, Fouque D,
Goldsmith D, Kanbay M, Mallamaci F, Massy ZA, Rossignol P,
Vanholder R, et al Board of the EURECA-m Working Group of ERA-EDTA:
Epidemiology, contributors to, and clinical trials of mortality
risk in chronic kidney failure. Lancet. 383:1831–1843. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stengel B: Chronic kidney disease and
cancer: A troubling connection. J Nephrol. 23:253–262.
2010.PubMed/NCBI
|
|
3
|
Hsu CY, Ordoñez JD, Chertow GM, Fan D,
McCulloch CE and Go AS: The risk of acute renal failure in patients
with chronic kidney disease. Kidney Int. 74:101–107. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalantar-Zadeh K, Block G, Humphreys MH
and Kopple JD: Reverse epidemiology of cardiovascular risk factors
in maintenance dialysis patients. Kidney Int. 63:793–808. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ortiz A, Massy ZA, Fliser D, Lindholm B,
Wiecek A, Martínez-Castelao A, Covic A, Goldsmith D, Süleymanlar G,
London GM, et al: Clinical usefulness of novel prognostic
biomarkers in patients on hemodialysis. Nat Rev Nephrol. 8:141–150.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shidfar F, Ebrahimi SS, Hosseini S,
Heydari I, Shidfar S and Hajhassani G: The effects of Berberis
vulgaris fruit extract on serum lipoproteins, apoB, apoA-I,
homocysteine, glycemic control and total antioxidant capacity in
type 2 diabetic patients. Iran J Pharm Res. 11:643–652.
2012.PubMed/NCBI
|
|
7
|
Wang Y, Campbell T, Perry B, Beaurepaire C
and Qin L: Hypoglycemic and insulin-sensitizing effects of
berberine in high-fat diet- and streptozotocin-induced diabetic
rats. Metabolism. 60:298–305. 2011. View Article : Google Scholar
|
|
8
|
Zha W, Liang G, Xiao J, Studer EJ, Hylemon
PB, Pandak WM Jr, Wang G, Li X and Zhou H: Berberine inhibits HIV
protease inhibitor-induced inflammatory response by modulating ER
stress signaling pathways in murine macrophages. PLoS One.
5:e90692010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Katiyar SK, Meeran SM, Katiyar N and
Akhtar S: p53 cooperates berberine-induced growth inhibition and
apoptosis of non-small cell human lung cancer cells in vitro and
tumor xenograft growth in vivo. Mol Carcinog. 48:24–37. 2009.
View Article : Google Scholar
|
|
10
|
Jiang Q, Liu P, Wu X, Liu W, Shen X, Lan
T, Xu S, Peng J, Xie X and Huang H: Berberine attenuates
lipopolysaccharide-induced extracelluar matrix accumulation and
inflammation in rat mesangial cells: Involvement of NF-κB signaling
pathway. Mol Cell Endocrinol. 331:34–40. 2011. View Article : Google Scholar
|
|
11
|
Ichikawa H, Takada Y, Murakami A and
Aggarwal BB: Identification of a novel blocker of IκBα kinase that
enhances cellular apoptosis and inhibits cellular invasion through
suppression of NF-κB-regulated gene products. J Immunol.
174:7383–7392. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jackson-Bernitsas DG, Ichikawa H, Takada
Y, Myers JN, Lin XL, Darnay BG, Chaturvedi MM and Aggarwal BB:
Evidence that TNF-TNFR1-TRADD-TRAF2-RIP-TAK1-IKK pathway mediates
constitutive NF-kappaB activation and proliferation in human head
and neck squamous cell carcinoma. Oncogene. 26:1385–1397. 2007.
View Article : Google Scholar
|
|
13
|
Korchnak AC, Zhan Y, Aguilar MT and Chadee
DN: Cytokine-induced activation of mixed lineage kinase 3 requires
TRAF2 and TRAF6. Cell Signal. 21:1620–1625. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Khan KA, Abbas W, Varin A, Kumar A, Di
Martino V, Dichamp I and Herbein G: HIV-1 Nef interacts with HCV
Core, recruits TRAF2, TRAF5 and TRAF6, and stimulates HIV-1
replication in macrophages. J Innate Immun. 5:639–656. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tada K, Okazaki T, Sakon S, Kobarai T,
Kurosawa K, Yamaoka S, Hashimoto H, Mak TW, Yagita H, Okumura K, et
al: Critical roles of TRAF2 and TRAF5 in tumor necrosis
factor-induced NF-κB activation and protection from cell death. J
Biol Chem. 276:36530–36534. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao T, Cheng C, Ji Y, Xu G, Zhang J, Zhang
L and Shen A: Numbl inhibits glioma cell migration and invasion by
suppressing TRAF5-mediated NF-κB activation. Mol Biol Cell.
23:2635–2644. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ko YA, Mohtat D, Suzuki M, Park AS,
Izquierdo MC, Han SY, Kang HM, Si H, Hostetter T, Pullman JM, et
al: Cytosine methylation changes in enhancer regions of core
pro-fibrotic genes characterize kidney fibrosis development. Genome
Biol. 14:R1082013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang LN, Wang Y, Lu Y, Yin ZF, Zhang YH,
Aslanidi GV, Srivastava A, Ling CQ and Ling C: Pristimerin enhances
recombinant adeno-associated virus vector-mediated transgene
expression in human cell lines in vitro and murine hepatocytes in
vivo. J Integr Med. 12:20–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saha SK, Roy S and Khuda-Bukhsh AR:
Ultra-highly diluted plant extracts of Hydrastis canadensis and
Marsdenia condurango induce epigenetic modifications and alter gene
expression profiles in HeLa cells in vitro. J Integr Med.
13:400–411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu D, Wen W, Qi CL, Zhao RX, Lü JH, Zhong
CY and Chen YY: Ameliorative effect of berberine on renal damage in
rats with diabetes induced by high-fat diet and streptozotocin.
Phytomedicine. 19:712–718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen J, Qiao YQ, Ran ZH and Wang TR:
upregulation and pre-activation of TRAF3 and TRAF5 in inflammatory
bowel disease. Int J Med Sci. 10:156–163. 2013. View Article : Google Scholar :
|
|
22
|
Ruiz-Ballesteros E, Mollejo M, Rodriguez
A, Camacho FI, Algara P, Martinez N, Pollán M, Sanchez-Aguilera A,
Menarguez J, Campo E, et al: Splenic marginal zone lymphoma:
Proposal of new diagnostic and prognostic markers identified after
tissue and cDNA microarray analysis. Blood. 106:1831–1838. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Horie R and Watanabe T, Ito K, Morisita Y,
Watanabe M, Ishida T, Higashihara M, Kadin M and Watanabe T:
Cytoplasmic aggregation of TRAF2 and TRAF5 proteins in the
Hodgkin-Reed-Sternberg cells. Am J Pathol. 160:1647–1654. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Missiou A, Rudolf P, Stachon P, Wolf D,
Varo N, Aichele P, Colberg C, Hoppe N, Ernst S, Münkel C, et al:
TRAF5 deficiency accelerates atherogenesis in mice by increasing
inflammatory cell recruitment and foam cell formation. Circ Res.
107:757–766. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Su VY-F and Yang K-Y: Mesenchymal stem
cell-conditioned medium induces neutrophils apoptosis via
inhibition of NF-κB pathway and increases endogenous pulmonary stem
cells in endotoxin-induced acute lung injury. Eur Respir J.
46(Suppl 59): OA35202015.
|
|
26
|
Arora R, Yates C, Gary BD, McClellan S,
Tan M, Xi Y, Reed E, Piazza GA, Owen LB and Dean-Colomb W:
Panepoxydone targets NF-κB and FOXM1 to inhibit proliferation,
induce apoptosis and reverse epithelial to mesenchymal transition
in breast cancer. PLoS One. 9:e983702014. View Article : Google Scholar
|
|
27
|
Chen F, Wang M, O'Connor JP, He M,
Tripathi T and Harrison LE: Phosphorylation of PPARgamma via active
ERK1/2 leads to its physical association with p65 and inhibition of
NF-kappabeta. J Cell Biochem. 90:732–744. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eschwege P1, Paradis V, Conti M, Loric S,
Dumas F, Berteau P, Ahmed M, Droupy S, Charpentier B, Legrand A, et
al: Bcl-2 and Bax expression on rat ischemic kidney. Transplant
Proc. 30:2861–2862. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saikumar P, Dong Z, Patel Y, Hall K,
Hopfer U, Weinberg JM and Venkatachalam MA: Role of hypoxia-induced
Bax translocation and cytochrome c release in reoxygenation injury.
Oncogene. 17:3401–3415. 1998. View Article : Google Scholar
|
|
30
|
Chien CT, Chang TC, Tsai CY, Shyue SK and
Lai MK: Adenovirus-mediated bcl-2 gene transfer inhibits renal
ischemia/reperfusion induced tubular oxidative stress and
apoptosis. Am J Transplant. 5:1194–1203. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ikezumi Y, Suzuki T, Karasawa T, Kawachi
H, Nikolic-Paterson DJ and Uchiyama M: Activated macrophages
downregulate podocyte nephrin and podocin expression via
stress-activated protein kinases. Biochem Biophys Res Commun.
376:706–711. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Perysinaki GS, Moysiadis DK, Bertsias G,
Giannopoulou I, Kyriacou K, Nakopoulou L, Boumpas DT and Daphnis E:
Podocyte main slit diaphragm proteins, nephrin and podocin, are
affected at early stages of lupus nephritis and correlate with
disease histology. Lupus. 20:781–791. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee YS, Kim WS, Kim KH, Yoon MJ, Cho HJ,
Shen Y, Ye JM, Lee CH, Oh WK, Kim CT, et al: Berberine, a natural
plant product, activates AMP-activated protein kinase with
beneficial metabolic effects in diabetic and insulin-resistant
states. Diabetes. 55:2256–2264. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Derosa G, Maffioli P and Cicero AF:
Berberine on metabolic and cardiovascular risk factors: An analysis
from preclinical evidences to clinical trials. Expert Opin Biol
Ther. 12:1113–1124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lan T, Shen X, Liu P, Liu W, Xu S, Xie X,
Jiang Q, Li W and Huang H: Berberine ameliorates renal injury in
diabetic C57BL/6 mice: Involvement of suppression of SphK-S1P
signaling pathway. Arch Biochem Biophys. 502:112–120. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen L, Guo W, Zhang S, Lu W, Liao S and
Li Y: Berberine prevents high glucose-induced cell viability
inhibition and apoptosis in podocytes. Int J Clin Exp Med.
9:5942–5950. 2016.
|
|
37
|
Wan X, Chen X, Liu L, Zhao Y, Huang WJ,
Zhang Q, Miao GG, Chen W, Xie HG and Cao CC: Berberine ameliorates
chronic kidney injury caused by atherosclerotic renovascular
disease through the suppression of NFκB signaling pathway in rats.
PLoS One. 8:e597942013. View Article : Google Scholar
|