Introduction
Hepatocellular carcinoma (HCC) ranks as the fifth
most frequently occurring neoplasm and the second leading cause of
cancer-associated mortalities worldwide. A total of >745,000
fatalities occur as a result of HCC, along with 500,000 newly
diagnosed cases every year (1–3).
Current diagnostic standards may be inaccurate and result in missed
cases, and since early diagnosis is key for successful operations,
it is imperative to search for more efficient biomarkers to
increase the early diagnostic rate of HCC (4,5).
Cancer stem cells (CSCs) are the source of numerous solid tumor
types, including HCC, and are characterized by a variety of stem
cell markers (6–11). A previous study revealed that
cluster of differentiation (CD)90 (Thy-1) is a potential marker for
liver CSCs (12) and is expressed
in a variety of cells, including T-cells, thymocytes, neurons,
endothelial cells and fibroblasts. It is important in cell-to-cell
and cell-to-matrix interactions, apoptosis, adhesion, migration and
fibrosis (13). Previous studies
have demonstrated that CD90+, however not
CD90− cells, obtained from HCC cell lines, exhibit
tumorigenic and metastatic capacities (12,14,15). The human HCC cell line JHH-6
demonstrates increased proliferative capacity of CD90+
compared with CD90− cells (16). Conversely, CD90 functions as a
tumor suppressor in ovarian cancer (17). The specific function and mechanism
by which CD90+ cells contribute to HCC progression
remains to be elucidated.
Circular RNAs (circRNAs) are a novel type of
endogenous noncoding RNA characterized as stable, abundant, and
conserved, and usually exhibit tissue/developmental-stage specific
expression (18). Numerous
studies have revealed that circRNAs participate in the initiation
and development of multiple diseases, including Alzheimer's,
Parkinson's, atherosclerosis, and various cancers (19,20). CircRNAs bind to miRNAs, acting as
miRNA sponges, and thereby regulate gene expression (21). Hsa_circ_0004018 has been reported
to be involved in cancer-associated pathways in HCC via
interactions with miRNAs (22).
The circRNA that acts as a sponge for miR-7 is Cdr1as (ciRS-7), the
knockdown of which suppresses HCC cell viability and invasion
(23). This evidence suggests
that circRNAs are involved in HCC progression and may be promising
diagnostic or predictive biomarkers. However, the expression of
circRNAs in HCC has not extensively been studied and the mechanisms
of circRNAs in HCC remain to be elucidated.
The present study aimed to investigate the effect of
CD90 on the cell cycle and the migration, invasion, and sphere
formation abilities of HCC cells. Furthermore, differentially
expressed circRNAs between CD90+ and CD90−
HCC cells were identified via high throughput microarray assay.
Materials and methods
Ethics statement
The present study was approved by the Ethics
Committee of Sun Yat-Sen University (Guangzhou, China), and written
informed consent was obtained from all patients in Sun Yat-Sen
Memorial Hospital of Sun Yat-Sen University.
Specimen collection and cell culture
A total of eight pairs of HCC tumor tissues and
corresponding non-tumor tissues were collected from the Department
of Hepatopancreatobiliary Surgery, Sun Yat-Sen Memorial Hospital of
Sun Yat-Sen University. The present study included 8 patients with
HCC, 5 males and 3 females, aged 35–66 years with a mean age of
55.9±9.5 years. Exclusion criteria were the following: i) Patients
with moderate or tense ascites, ii) patients refusing liver biopsy,
iii) patients with previous treatment for HCC and iv) patients with
a history of severe trauma. None of the recruited participants
accepted adjunctive treatment prior to the surgery. All tissue
samples were immediately preserved at −80°C following washing with
sterile phosphate-buffered saline (PBS). HCC cell lines (LM3, Huh7,
MHCC 97L and SK-Hep-1) were purchased from Shanghai Cell Center
(Shanghai, China) and cultured in Dulbecco's modified Eagle's
medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal calf serum (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) and 1% penicillin-streptomycin G
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were incubated
at 37°C in a humidified atmosphere containing 5%
CO2.
Flow cytometry
The quantitation of CD90 expression in the cells
(Huh7, MHCC 97L and SK-Hep-1) was determined using flow cytometry.
A total of 2×105 cells were transferred into a
centrifuge tube and suspended in PBS buffer. Following
centrifugation at 1,000 × g for 5 min at 4°C, the cells were fixed
with fresh fixative (4% paraformaldehyde; Thermo Fisher Scientific
Inc.) at room temperature for 15 min, and then preincubated in 200
ml blocking solution (10% goat serum containing 0.05% Tween-20;
Thermo Fisher Scientific Inc.) for 1 h at room temperature. The
cells were incubated with fluorescein isothiocyanate
(FITC)-conjugated human anti-CD90 (catalog no. 85-11-0909-41; 1:100
dilution; eBiosciences; Thermo Fisher Scientific Inc.) for 1 h at
4°C, then the cells were washed in PBS and incubated with
FITC-conjugated goat anti-mouse IgG secondary antibody (catalog no.
ab6785; 1:1,000 dilution; Abcam, Cambridge, MA, USA). The standard
was established by isotype match control (FITC-conjugated mouse
IgG1 isotype control, catalog no. 85-11-4714-81; 1:100 dilution;
eBiosciences; Thermo Fisher Scientific Inc.) for 1 h at 4°C. A
total of 1×106 FITC-labeled cells were measured using BD
AccuriC6 Flow Cytometer with Cell Quest software version 5.1 (BD
Biosciences, Franklin Lakes, NJ, USA). Each experiment was
performed in triplicate. The cell cycle of SK-Hep-1
CD90+ and CD90− cells was additionally
determined with flow cytometry. Briefly, cells were resuspended in
PBS twice prior to fixation by dropwise addition into 95%
pre-cooled ethanol for 15 min at room temperature, and then
centrifuged at 3,000 × g for 10 min at 4°C, resuspended twice in
PBS, and stained with propidium iodide (PI) containing 50
µg/ml RNaseA (Takara Bio, Inc., Otsu, Japan) in the dark for
30 min at room temperature. The DNA content was analyzed using BD
AccuriC6 Flow Cytometer with Cell Quest software, version 5.1 (BD
Biosciences).
Cell separation via magnetic sorting
Separation of CD90+ cells from SK-Hep-1
cells was performed using MACS magnetic cell sorting (Miltenyi
Biotech GmbH, Bergisch Gladbach, Germany) according to the
manufacturer's protocol. SK-Hep-1 cells were incubated with 50
µl of FcR blocking agent (Miltenyi Biotech GmbH) and
CD90-biotin immunomagnetic beads (Miltenyi Biotech GmbH) for 10 min
at 4°C. Goat anti-mouse IgG magnetic beads (Miltenyi Biotech GmbH)
were used as the control. A MACS cell separation column (MS column)
was used to retain the CD90+ cells linked to the beads.
Unlabeled cells (SK-Hep-1 CD90− cells) were washed with
3×500 µl buffer (containing PBS, 0.5% BSA, 2 mM EDTA) and
collected. The MS column was then removed from the separator and
placed on another collection tube. A total of 1 ml buffer was
pipetted onto the MS column, and the labeled CD90+ cells
(SK-Hep-1 CD90+ cells) were flushed out by applying the
plunger supplied with the column. The labeled CD90+
cells were eventually obtained via centrifugation at 3,000 × g for
10 min at 4°C and resuspension in serum-free medium (Gibco; Thermo
Fisher Scientific, Inc.). The SK-Hep-1 CD90+ and
SK-Hep-1 CD90− cells were harvested and washed with PBS,
and the expression of CD90 in these cells was directly investigated
with flow cytometry. Following sorting, the SK-Hep-1
CD90+ cells indicated an ≥85% expression level of CD90
and the SK-Hep-1 CD90− cells revealed <5% expression
level of CD90.
Western blotting
SK-Hep-1 cells were lysed in lysis buffer (Gibco;
Thermo Fisher Scientific, Inc.) and the concentration of total
protein was analyzed using a BCA Protein Assay Kit (Sangon Biotech
Co., Ltd, Shanghai, China). A total of 30 µg protein was
added into each lane of the 10% SDS-PAGE gel and 5% skim milk (BD
Biosciences) was used to block the polyvinylidene membranes for 10
min at room temperature. The membranes were then incubated with the
primary antibody anti-CD90/Thy1 (catalog no. ab133350; 1:100;
Abcam) at 4°C overnight. GAPDH was used as the internal control
(anti-GAPDH antibody; catalog no. ab9485; 1:100, Abcam). The next
day, the membranes were incubated with horseradish
peroxidase-conjugated anti-rabbit IgG secondary antibody for 1 h at
room temperature (1:500 dilution; catalog no. ab7090; Abcam). The
results were visualized using the enhanced chemiluminescence
substrate kit (GE Healthcare, Chicago, IL, USA). All images were
analyzed with ImageJ software, version 14.8 (National Institutes of
Health, Bethesda, MD, USA).
Cell viability
Cell viability was assessed with a 3-(4,5- dim
ethylthiazol-2-yl)-2,5-diphenyltrtrazolium bromide (MTT) assay. The
treated cells were seeded into a 96-well plate and cultured in
complete medium. When the cells adhered, 0.5 mg/ml MTT was added to
each well and incubated at 37°C for 4 h. The supernatant was then
carefully aspirated and 100 µl of dimethyl sulfoxide was
added. The absorbance was measured at a wavelength of 490 nm using
a microplate reader (Thermo Fisher Scientific, Inc.).
Migration and invasion assays
Cell migration and invasion assays were performed
using Transwell chambers with 8 µm pores (BD Biosciences)
according to the manufacturer's protocol. For migration assays,
SK-Hep-1 CD90+ (5×104 cells/well) and
SK-Hep-1 CD90− cells (5×104 cells/well) were
separately seeded into the upper compartment without Matrigel, for
invasion assay, the cells were seeded into the upper compartment
with Matrigel (Corning Inc., Corning, NY, USA) and incubated in
serum-free DMEM (Gibco; Thermo Fisher Scientific, Inc.) and the
lower compartment was filled with complete medium supplemented with
10% FBS. Following 24 h, migratory and invasive cells on the bottom
surface of the filters were fixed with 4% paraformaldehyde for 24 h
and stained with 0.1% crystal violet solution for 10 min at room
temperature. The cells in at least 10 randomly selected microscopic
fields were counted under a light microscope, at ×200 magnification
(Nikon Corp., Tokyo, Japan). Each experiment was performed in
triplicate.
Sphere formation assay
SK-Hep-1 CD90+ and SK-Hep-1
CD90− cells were trypsinized and cultured in DMEM-F12
(Sigma-Aldrich; Merck KGaA) that was supplemented with 20 ng/ml
EGF, 10 ng/ml FGF, 4 ng/ml insulin, and B27 (1:50) in Ultra-Low
Attachment 6-well plates (Corning Inc.). The cells were imaged
under a light microscope (magnification, ×200; Olympus Corp.,
Tokyo, Japan) following 20 days of incubation.
Microarray hybridization
Total RNA was treated with Rnase R (Epicentre
Illumina, Inc., San Diego, CA, USA) to remove linear RNA, and was
then amplified and transcribed into fluorescent cRNA using Quick
Amp Labeling kit (Takara Bio, Inc.) according to the manufacturer's
protocol. The cRNA was then purified with the RNeasy Mini Kit
(Qiagen GmbH, Hilden, Germany) and hybridized to the circRNA
expression microarray according to the manufacturer's protocols.
The microarray hybridization and the data collection were performed
by Forevergen Bio-tech Co. (Guangzhou, China). For the microarray
analysis, following filtering by flag signal, the raw data were
processed and normalized. Differentially expressed circRNA were
identified with P<0.05 and fold changes >2. In addition,
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses was
performed which provided more detailed information regarding the
pathways in which the target genes were involved.
Specialty confirmation and Sanger
sequencing
To verify the specialty of the PCR products of
hsa_circ_0067531 and hsa_circ_0057096, the cDNA and gDNA PCR
products that had been amplified by diverted primers were separated
on a 2% agarose gel with TE running buffer. Only a single band in
the gel was thought to be a specialty. The region was incised from
the whole gel and purified with SanPrep Column DNA Gel Extraction
Kit (Qiagen GmbH), and the PCR fragments were inserted into T
vector for Sanger sequencing (Tsingke Biotech Co., Ltd., Beijing,
China).
Quantitative polymerase chain reaction
(qPCR)
Total RNA was isolated using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) following
indicated treatments. qPCR assays were performed with SYBR Premix
Ex Taq (Takara Bio, Inc.), primers, and a cDNA template on the
Applied Biosystems 7500 Real-time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The primers used in the assay are
listed as follows: Forward, CGA ACC AAC TTC ACC AGC AA and reverse,
CTG ATG CCC TCA CAC TTG AC for CD90; forward, 5′-TCC AGA CCA GTA
CGT TCG AG-3′ and reverse, 5′-TCA CAC AGT TGA GAC AAG GGA T-3′ for
cir_0067531; forward, 5′-AGG CAA AGG AAG TCC ATC TCA T-3′ and
reverse, 5′-TCA ATC ACA CCC TGG GCC AT-3′ for circ_0057096 and
forward, 5′-CCG AGA ATG GGA AGC TTG TC-3′ and reverse, 5′-AAG CAC
CAA CAG AGG AGA A-3′ for GAPDH. PCR was conducted in a 50 µl
reaction system, including 1 µl forward and 1 µl
reverse primers, 0.5 µl 20× SYBR-Green I, 25 µl 2×PCR
buffer, 2 µl cDNA, and 20.5 µl DEPC. The qPCR program
was 94°C for 4 min, 30 cycles of 94°C for 20 sec, 60°C for 30 sec,
72°C for 30 sec, and 72°C for 5 min. Each individual sample was run
in triplicate and the expression level was quantified with the
comparative cycle threshold (Cq) method. Results were normalized to
GAPDH expression and RNA enrichments were calculated using the
2−ΔΔCq method (24).
Statistical analysis
Quantile normalization and subsequent data
processing were performed using the GeneSpring GX software, version
11.5.1 package (Agilent Technologies, Inc., Santa Clara, CA, USA).
Other data were analyzed with the two-tail Student's t-test and
one-way analysis of variance followed by Tukey's test. SPSS
software, version 19.0 (IBM SPSS, Armonk, NY, USA). Each experiment
was repeated at least three times. All results were summarized and
are presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Distribution of CD90 cells in human HCC
cell lines
The expression levels of CD90 in the 4 different HCC
cell lines were determined by qPCR; results of which demonstrated
that the expression of CD90 was increased in the LM3, Huh7, and
MHCC 97L cell lines, and particularly in the SK-Hep-1 cell line
(Fig. 1A). The western blotting
results demonstrated that the protein level of CD90 in SK-Hep-1
CD90+ cells was significantly increased compared with
SK-Hep-1 and SK-Hep-1 CD90− cells. However, CD90
expression in SK-Hep-1 CD90− cells was significantly
decreased compared with SK-Hep-1 cells (Fig. 1B). In addition, the expression of
CD90 (blue line) and control IgG (red line) were verified in the
Huh7, MHCC 97L, and SK-Hep-1 cell lines by flow cytometry. It was
demonstrated that the relative expression level of CD90 in SK-Hep-1
cells (10.0%) was significantly increased compared with Huh7
(0.122%) and MHCC 97L (0.045%) cell lines (Fig. 1C). Consequently, the SK-Hep-1 cell
line was selected for the ensuing experiments. The CD90+
cells were sorted from the SK-Hep-1 cell line with an expression of
CD90 (88.1%) ≥85.0%, whereas CD90− cells were sorted
from the SK-Hep-1 cell line with an expression of CD90 (0.997%)
≤5.0% (Fig. 1C).
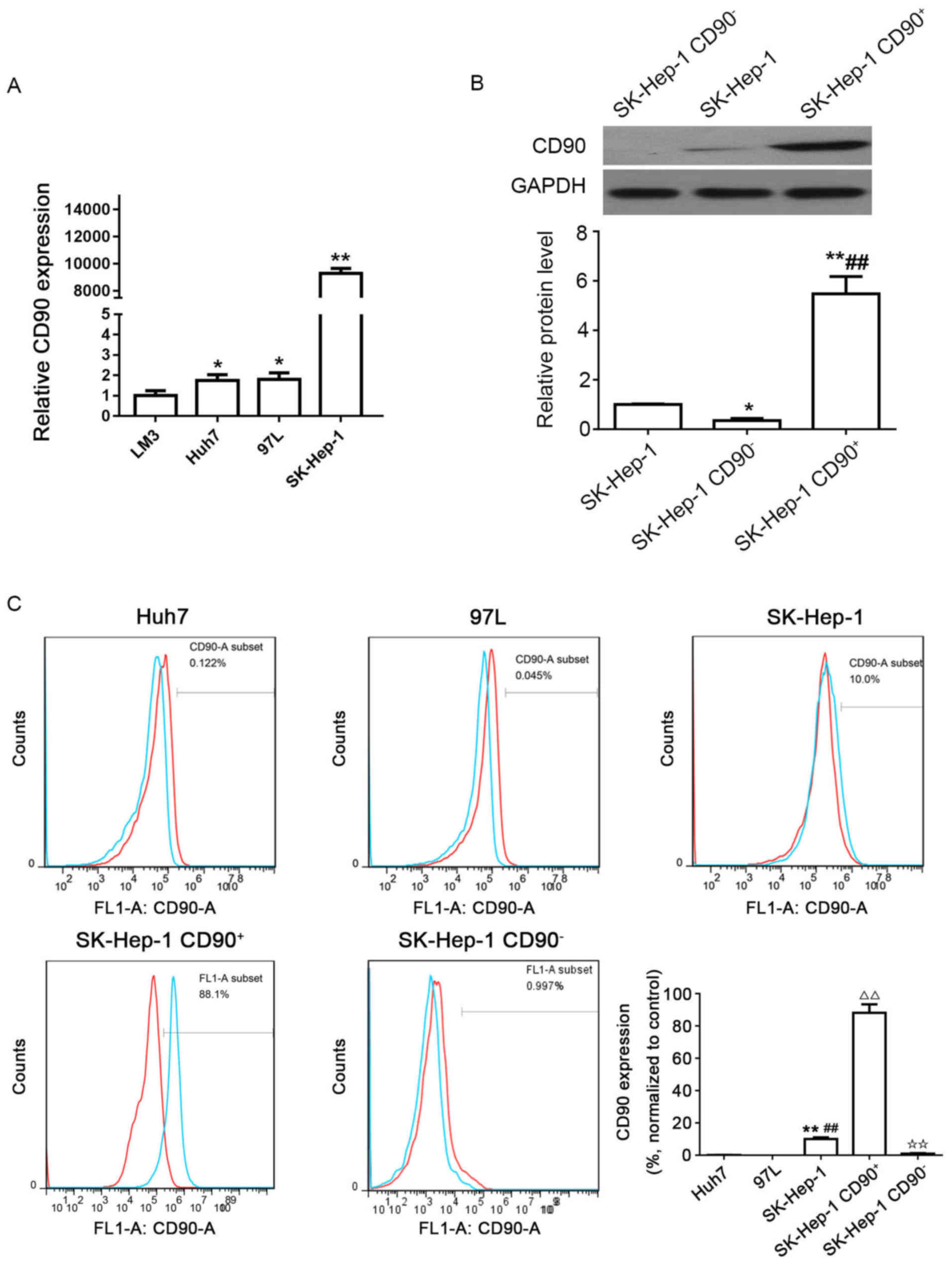 | Figure 1Distribution of CD90 cells in human
HCC cell lines. (A) Expression levels of CD90 in 4 different HCC
cell lines (LM3, Huh7, MHCC 97L and SK-Hep-1) were determined by
quantitative polymerase chain reaction. ✩P<0.05;
✩✩P<0.01 vs. LM3. (B) Protein expression levels of
CD90 in SK-Hep-1, SK-Hep-1 CD90−, and SK-Hep-1
CD90+ cells were determined by western blotting. GAPDH
was used as the internal control. ✩P<0.05;
✩✩P<0.01 vs. SK-Hep-1; ##P<0.01 vs.
SK-Hep-1 CD90−. (C) Expression levels of CD90 in 3
different HCC cell lines (Huh7, MHCC 97L, and SK-Hep-1), SK-Hep-1
CD90+ cells and SK-Hep-1 CD90− cells were
examined with flow cytometry. Each experiment was performed in
triplicate. IgG was used for control (red line). The blue line
reveals the expression of CD90. The SK-Hep-1 CD90+ cells
revealed an expression of 88.1% CD90, whereas CD90−
cells exhibited a CD90 expression of 0.997%. ✩✩P<0.01
vs. Huh7; ##P<0.01 vs. MHCC 97L;
△△P<0.01 vs. SK-Hep-1; ✩✩P<0.01 vs.
SK-Hep-1 CD90+. CD, cluster of differentiation; HCC,
hepatocellular carcinoma. |
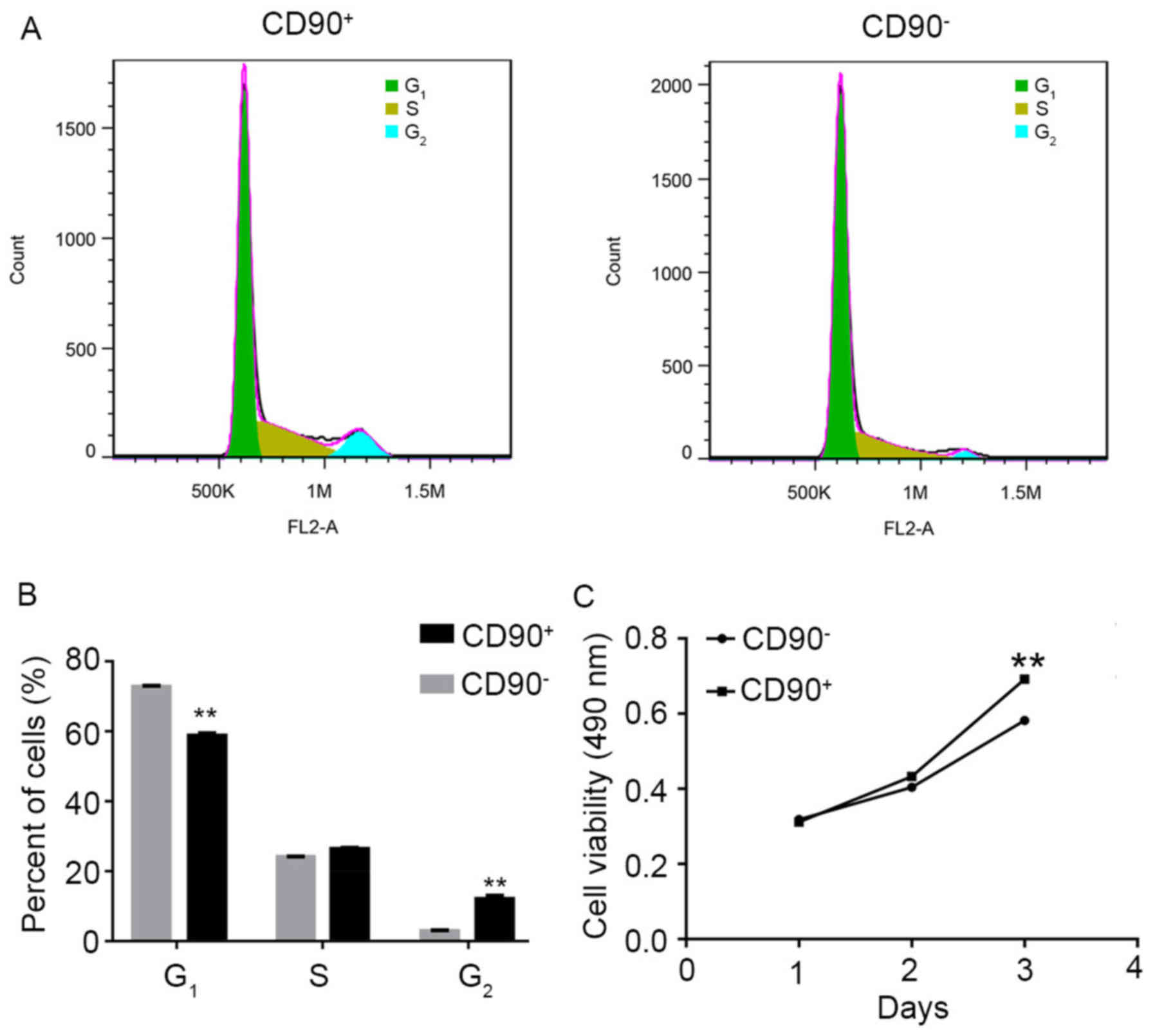
Effect of CD90 on the cell cycle and
viability of SK-Hep-1 cells
Flow cytometry analysis and MTT assay were performed
to investigate the effect of CD90 on the cell cycle and viability
in SK-Hep-1 cells. The raw FACScan data is presented in Fig. 2A. The numerical conversion of the
cell population in G1, S, and G2 phases is
presented in Fig. 2B. The results
indicated that the percentage of CD90+ cells was
significantly decreased in the G1 phase, compared with
CD90− cells. Conversely, the percentage of
CD90+ cells was significantly increased in the
G2 phase, compared with CD90− cells. These
results suggested that the G1 phase of CD90+
cells was shorter compared with CD90− cells. In
addition, the MTT assay results demonstrated that the cell
viability of CD90+ cells was significantly increased
compared with CD90− cells at day 3 (Fig. 2C).
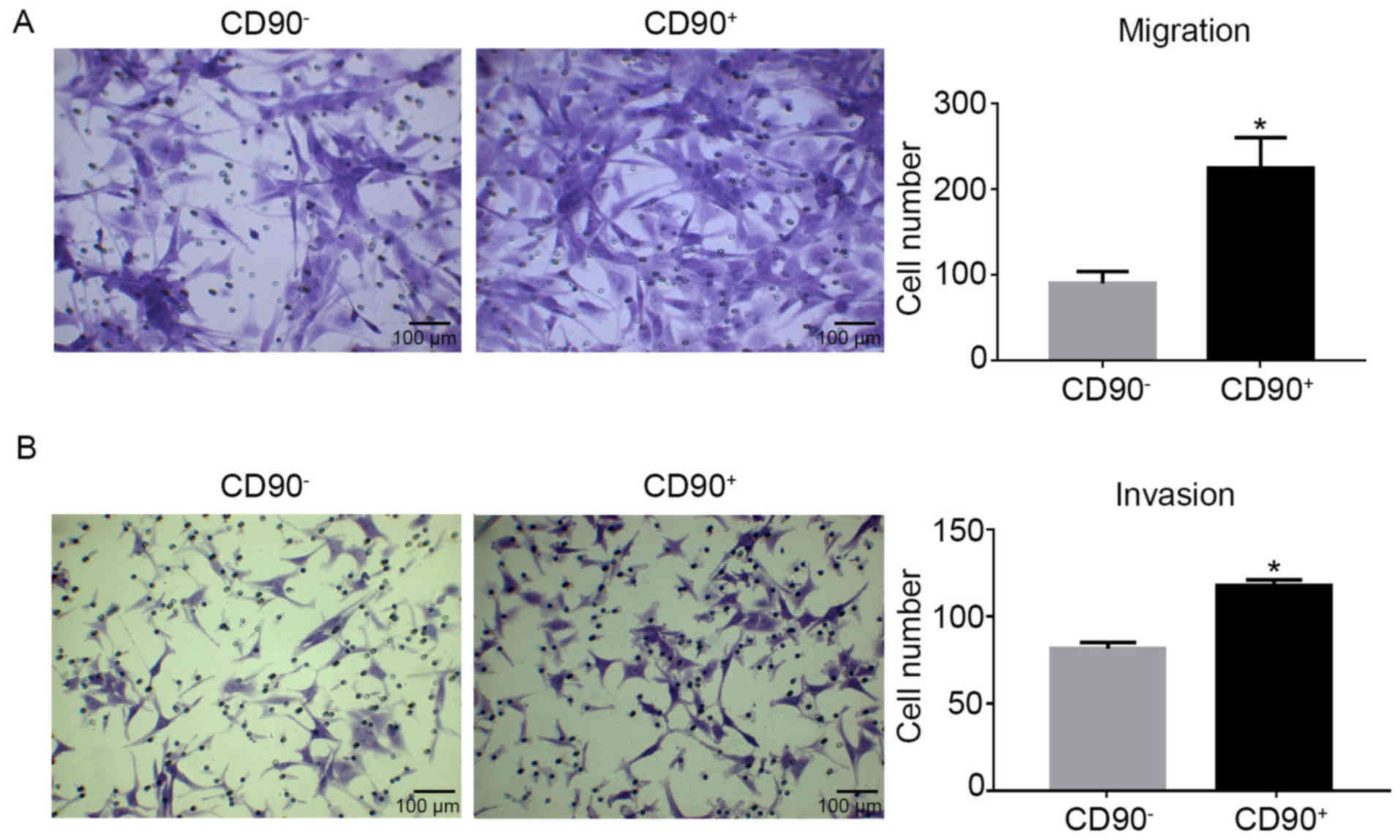
Migration and invasion abilities increase
in CD90+ cells
Transwell assays were used to determine the
migration and invasion abilities of CD90+ and
CD90− cells. The migration assay revealed that there
were significantly more CD90+ cells compared with
CD90− cells (Fig. 3A).
Similarly, the invasion assay demonstrated the same results as the
migration assay (Fig. 3B).
Consequently, it was demonstrated that CD90+ cells
exhibited increased migration and invasion capabilities compared
with CD90− cells.
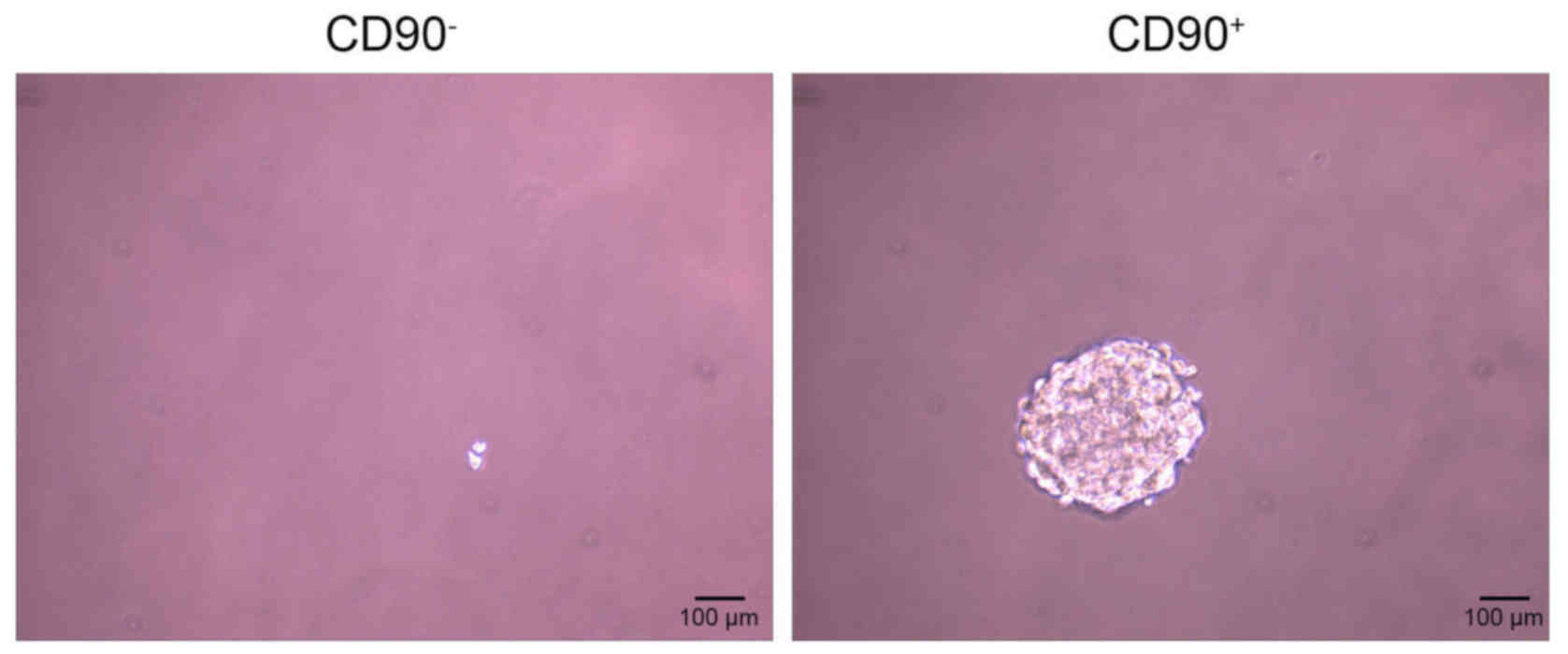
CD90+ induces sphere
formation
The sphere formation assay demonstrated that
CD90− cells did not form spheres following 20 days of
incubation, however the CD90+ cells did (Fig. 4).
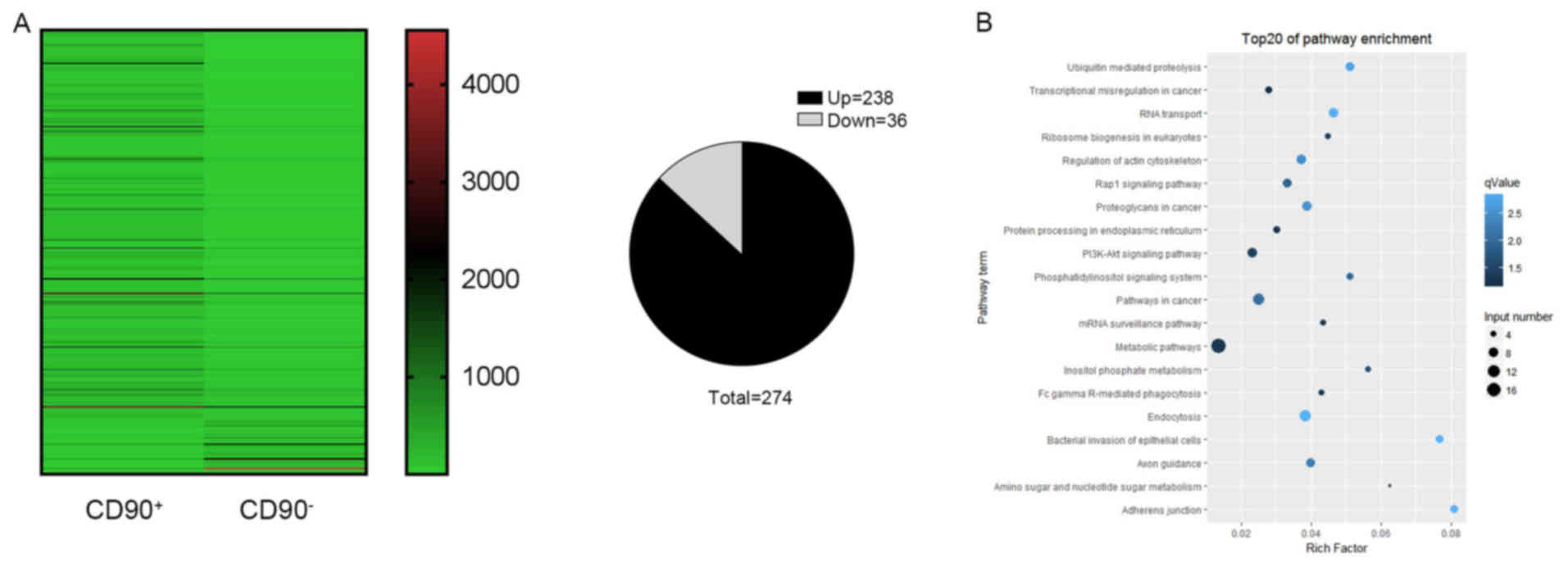
Identification of differentially
expressed circRNA profiles
A high throughput microarray assay was used to
identify the differing expressions of circRNAs in CD90+
and CD90− cells. In the present study, 274 dysregulated
circRNAs were identified, including 238 upregulated and 36
downregulated circRNAs (Fig. 5A).
The upregulated circRNAs were more frequently occurring compared
with downregulated circRNAs. In order to further understand the
biological functions of the differentially expressed circRNAs, KEGG
pathway analyses was performed which provided more detailed
information regarding the pathways in which the target genes were
involved. It was revealed that the key signaling pathways were the
metabolic pathway, pathways in cancer, and the phosphoinositide
3-kinase (P13K)-RAC-α serine/threonine-protein kinase (AKT)
signaling pathway (Fig. 5B).
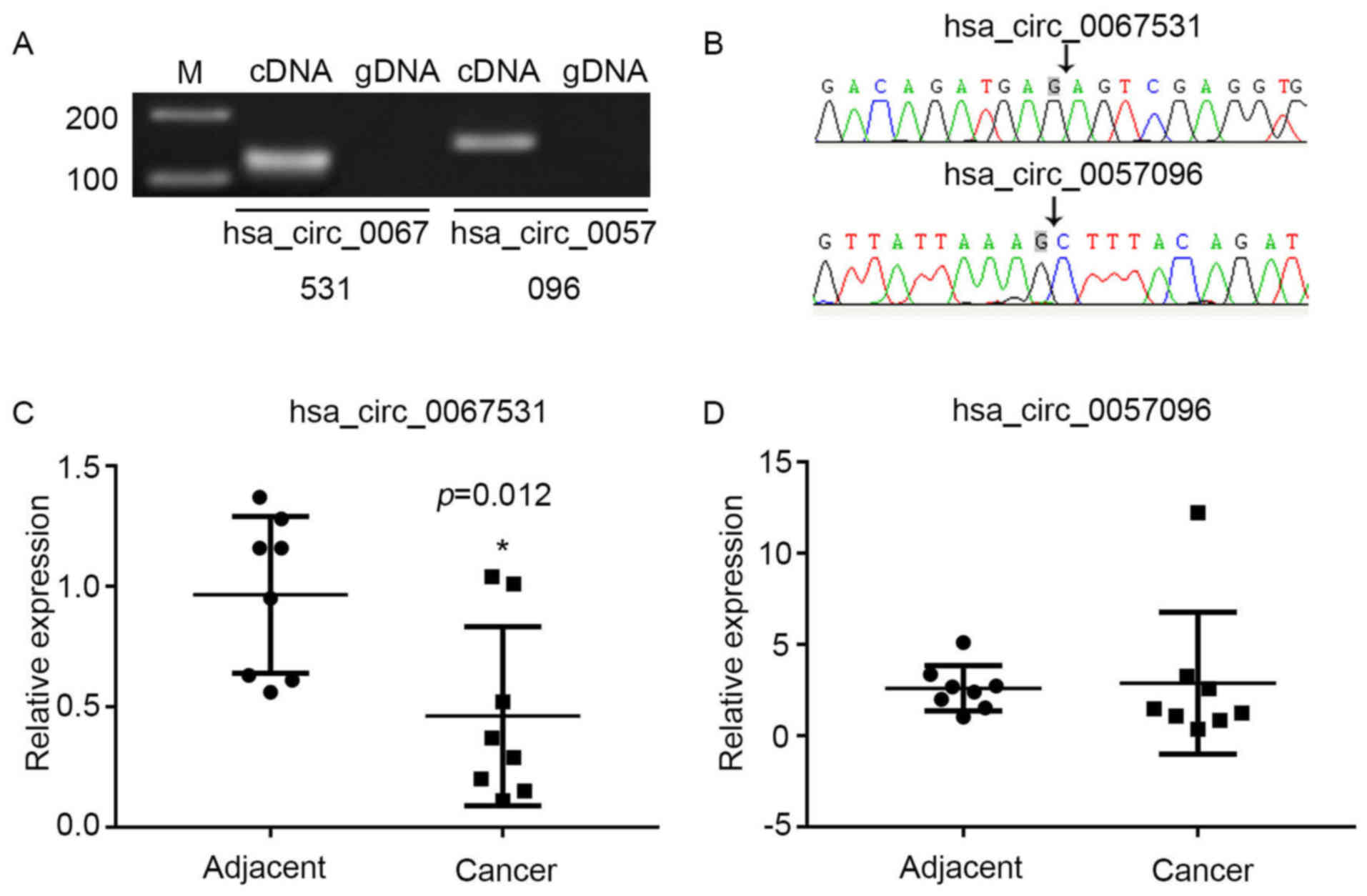
Characterization of hsa_circ_0067531 and
hsa_circ_0057096 in HCC
To verify the microarray data, two circRNAs with
differential expression were selected from the P13K-AKT signaling
pathway; hsa_circ_0067531 and hsa_ circ_0057096. The sequences of
these two circRNAs from pyruvate dehydrogenase kinase (PDK)1 and
phosphatidylino-sitol-4,5-bisphosphate 3-kinase catalytic subunit β
(PIK3CB) are listed in Table I.
Divergent primers amplify circRNAs in cDNA, however not genomic DNA
(gDNA) (Fig. 6A). The Sanger
sequencing verified head-to-tail splicing (Fig. 6B). The present study validated the
expression levels via qPCR in eight pairs of HCC and adjacent
normal tissues. It was demonstrated that the expression of
hsa_circ_0067531 was significantly decreased in the HCC tissues,
compared with the normal tissues (P=0.012) (Fig. 6C), however there was no
significant difference in hsa_circ_0057096 expression between
adjacent normal tissues and HCC tissues (Fig. 6D). Therefore, it was hypothesized
that hsa_circ_0067531 may be a promising target for HCC diagnosis
and therapy in the future.
 | Table ISequences of two differentiated
expression circular RNAs from PDK1 and PIK3CB. |
Table I
Sequences of two differentiated
expression circular RNAs from PDK1 and PIK3CB.
| circBASE ID | Gene | Sequence
(5′-3′) |
|---|
|
hsa_circ_0057096 | PDK1 C |
TTTACAGATACTGTGATACGGATCAGAAACCGACACAATGATGTCATTCCCACA
ATGGCCCAGGGTGTGATTGAATACAAGGAGAGCTTTGGGGTGGATCCTGTCACC
AGCCAGAATGTTCAGTACTTTTTGGATCGATTCTACATGAGTCGCATTTCAATTAG
AATGTTACTCAATCAGCACTCTTTATTGTTTGGTGGAAAAGGCAAAGGAAGTCCA
TCTCATCGAAAACACATTGGAAGCATAAATCCAAACTGCAAT GTACTTGAAGTTA
TTAAAG |
|
hsa_circ_0067531 | PIK3CB |
AGTCGAGGTGGAAAAAAGTTTCTTCCTGTATTGAAAGAAATCTTGGACAGGGATCCC
TTGTCTCAACTGTGTGAAAATGAAATGGATCTTATTTGGACTTTGCGACAAG
ACTGCCGAGAGATTTTCCCACAATCACTGCCAAAATTACTGCTGTCAATCAAGTGG
AATAAACTTGAGGATGTTGCTCAGCTTCAGGCGCTGCTTCAGATTTGGCCTAAAC
TGCCCCCCCGGGAGGCCCTAGAGCTTCTGGATTTCAACTATCCAGACCAGTACG
TTCGAGAATATGCTGTAGGCTGCCTGCGACAGATGAG |
Discussion
circRNAs have previously emerged as novel and
crucial layers of gene regulation and have demonstrated the
potential to be ideal biomarkers in the diagnosis of various
cancers (25,26). Certain circRNAs have already been
demonstrated to exhibit important roles in cancer, including
hsa_circ_002059 in gastric cancer, Cdr1as in hepatocellular
carcinoma, and cir-ITCH in esophageal squamous cancer (23,27,28). Previous studies have revealed that
circRNAs may be involved in the progression of cancer (20). Bachmayr-Heyda, et al
(29) identified that circRNAs
are significantly downregulated in colorectal cancer (CRC) tissues
compared with the normal colon mucosa. Li et al (28) demonstrated that hsa_circ_002059
expression is downregulated in gastric cancer and may be a
potential biomarker for its diagnosis (28). Huang et al reported that
the cir-ITCH expression is decreased in esophageal squamous cell
carcinoma (ESCC) and CRC, and may have an inhibitory effect on ESCC
and CRC (30). The mechanisms of
circRNAs in HCC remain to be elucidated.
The primary outcome of the present study was that
CD90 promoted cell migration, viability and sphere-forming
abilities in HCC, and the secondary outcome was that
hsa_circ_0067531 and the PI3K pathway may potentially be involved
in the aforementioned process. CD90 is a 25-37 kDa
glycophosphatidylinositolanchored protein that operates as an
important regulator of cell-to-cell and cell-to-matrix interactions
in cancer (13). CD90 expression
has been suggested to be associated with poor HCC prognosis
(31–33) and CD90+ CSCs, however
CD90− CSCs from HCC cell lines, tumor tissues, or
peripheral blood, have not been reported to exhibit tumorigenic and
metastatic capabilities (12,14,15). The present study first examined
the expression of CD90 in the 4 human HCC cell lines, and
demonstrated that CD90 expression was significantly upregulated in
the HCC cell line SK-Hep-1. In addition, the data demonstrated that
CD90+ cells isolated from the SK-Hep-1 cell line
exhibited increased viability, migration and invasive capabilities
compared with CD90− cells. The results were consistent
with previous research which suggests that CD90+ cells
isolated from HCC tumor tissues have the capacity to generate tumor
nodules in immunodeficient mice, whereas CD90− cells do
not (12,34,35).
circRNAs have previously been demonstrated to
exhibit important roles in HCC cancer, including Circular RNA MTO1,
Hsa_circ_0001649, and circZKSCAN1 (36–38). In the present study, 274
differentially expressed circRNAs (including upregulated and
downregulated genes) were identified in CD90+ HCC cells
compared with CD90− HCC cells via a high throughput
microarray assay. Furthermore, KEGG pathway analyses was performed
in order to further understand the biological functions of the
differentially expressed circRNA. The results demonstrated that the
key signaling pathways were the metabolic pathway, pathways in
cancer, and the PI3K-AKT pathway. Of the significantly enriched
pathways in the KEGG pathway analysis, PI3K signaling was of
interest as it exhibits an important role in HCC cell cycle
progression and viability (39,40). Deregulation of the PI3K/AKT
signaling pathway has previously been identified in HCC (41), and Rab31, a member of the Ras
superfamily, has been reported to have a role in tumor development
and progression (42). The
PI3K/AKT pathway was revealed to be involved in the Rab31
promotion of HCC progression (43). Previous research has revealed that
inhibiting the activation of the PI3K pathway blocks the
carcinogenesis and progression of HCC cells (44,45). The present study therefore
selected two differentially expressed circRNAs, shsa_circ_0067531
and hsa_circ_0057096, from PDK1 and PIK3CB, respectively. It was
demonstrated that hsa_circ_0067531 was markedly downregulated in
HCC tissues compared with adjacent normal tissues.
In conclusion, the results of the present study
suggested that CD90 may be used as a potential biomarker for HCC.
It was revealed that CD90 promoted cell migration, viability and
sphere-forming abilities of HCC. In addition, the expression of
hsa_circ_0067531 was significantly decreased in HCC compared with
adjacent normal tissues, and this suggested that hsa_circ_0067531
may be involved in the development of HCC, at least in part,
through the PI3K pathway. However, the present study does not
functionally verify how hsa_circ_0067531 affects HCC cell
biological functions, and did not verify the involvement of the
metabolic and cancer pathways in tumorigenesis of CD90+
HCC. The authors aim to investigate the mechanism underlying the
modulation of hsa_circ_0067531 on HCC stem cell properties, and
investigate the functional roles of the metabolic and cancer
pathways in HCC development in future studies.
Acknowledgments
The present study was supported by the Major
Projects on Collaborative Innovation of Industry, Guangzhou (grant
no. 201508020076). The authors gratefully acknowledge the
assistance of the Department of Hepatopancreatobiliary Surgery for
their help in collecting medical records. In addition, the authors
would like to thank all the participants of the study, without whom
the study would not have been possible.
References
|
1
|
Asia-Pacific Working Party on Prevention
of Hepatocellular Carcinoma: Prevention of hepatocellular carcinoma
in the Asia-Pacific region: Consensus statements. J Gastroenterol
Hepatol. 25:657–663. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Z, Zhang C, Lou C, Yan F, Mao Y, Hong X
and Zhang Y: Comparison of percutaneous cryosurgery and surgical
resection for the treatment of small hepatocellular carcinoma.
Oncol Lett. 6:239–245. 2013.PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bruix J, Reig M and Sherman M:
Evidence-based diagnosis, staging, and treatment of patients with
hepatocellular carcinoma. Gastroenterology. 150:835–853. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kinoshita A, Koike K and Nishino H:
Clinical features and prognosis of elderly patients with
hepatocellular carcinoma not indicated for surgical resection.
Geriatr Gerontol Int. 17:189–201. 2017. View Article : Google Scholar
|
|
6
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Collins AT, Berry PA, Hyde C, Stower MJ
and Maitland NJ: Prospective identification of tumorigenic prostate
cancer stem cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar
|
|
10
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar
|
|
11
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang ZF, Ho DW, Ng MN, Lau CK, Yu WC, Ngai
P, Chu PW, Lam CT, Poon RT and Fan ST: Significance of
CD90+ cancer stem cells in human liver cancer. Cancer
Cell. 13:153–166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rege TA and Hagood JS: Thy-1 as a
regulator of cell-cell and cell-matrix interactions in axon
regeneration, apoptosis, adhesion, migration, cancer, and fibrosis.
FASEB J. 20:1045–1054. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamashita T, Honda M, Nakamoto Y, Baba M,
Nio K, Hara Y, Zeng SS, Hayashi T, Kondo M, Takatori H, et al:
Discrete nature of EpCAM+ and CD90+ cancer
stem cells in human hepatocellular carcinoma. Hepatology.
57:1484–1497. 2013. View Article : Google Scholar
|
|
15
|
Yang ZF, Ngai P, Ho DW, Yu WC, Ng MN, Lau
CK, Li ML, Tam KH, Lam CT, Poon RT and Fan ST: Identification of
local and circulating cancer stem cells in human liver cancer.
Hepatology. 47:919–928. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sukowati CH, Anfuso B, Torre G,
Francalanci P, Crocè LS and Tiribelli C: The expression of
CD90/Thy-1 in hepatocellular carcinoma: An in vivo and in vitro
study. PLoS One. 8:e768302013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen WC, Hsu HP, Li CY, Yang YJ, Hung YH,
Cho CY, Wang CY, Weng TY and Lai MD: Cancer stem cell marker CD90
inhibits ovarian cancer formation via β3 integrin. Int J Oncol.
49:1881–1889. 2016.PubMed/NCBI
|
|
18
|
Wang F, Nazarali AJ and Ji S: Circular
RNAs as potential biomarkers for cancer diagnosis and therapy. Am J
Cancer Res. 6:1167–1176. 2016.PubMed/NCBI
|
|
19
|
Li J, Yang J, Zhou P, Le Y, Zhou C, Wang
S, Xu D, Lin HK and Gong Z: Circular RNAs in cancer: Novel insights
into origins, properties, functions and implications. Am J Cancer
Res. 5:472–480. 2015.PubMed/NCBI
|
|
20
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kulcheski FR, Christoff AP and Margis R:
Circular RNAs are miRNA sponges and can be used as a new class of
biomarker. J Biotechnol. 238:42–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fu L, Yao T, Chen Q, Mo X, Hu Y and Guo J:
Screening differential circular RNA expression profiles reveals
hsa_circ_0004018 is associated with hepatocellular carcinoma.
Oncotarget. 8:58405–58416. 2017.PubMed/NCBI
|
|
23
|
Yu L, Gong X, Sun L, Zhou Q, Lu B and Zhu
L: The Circular RNA Cdr1as Act as an Oncogene in Hepatocellular
carcinoma through targeting miR-7 expression. PLoS One.
11:e01583472016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schmittgen TD: Real-time quantitative PCR.
Methods. 25:383–385. 2001. View Article : Google Scholar
|
|
25
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li F, Zhang L, Li W, Deng J, Zheng J, An
M, Lu J and Zhou Y: Circular RNA ITCH has inhibitory effect on ESCC
by suppressing the Wnt/β-catenin pathway. Oncotarget. 6:6001–6013.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li P, Chen S, Chen H, Mo X, Li T, Shao Y,
Xiao B and Guo J: Using circular RNA as a novel type of biomarker
in the screening of gastric cancer. Clin Chim Acta. 444:132–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bachmayr-Heyda A, Reiner AT, Auer K,
Sukhbaatar N, Aust S, Bachleitner-Hofmann T, Mesteri I, Grunt TW,
Zeillinger R and Pils D: Correlation of circular RNA abundance with
viability-exemplified with colorectal and ovarian cancer,
idiopathic lung fibrosis, and normal human tissues. Sci Rep.
5:80572015. View Article : Google Scholar
|
|
30
|
Huang G, Zhu H, Shi Y, Wu W, Cai H and
Chen X: cir-ITCH plays an inhibitory role in colorectal cancer by
regulating the Wnt/β-catenin pathway. PLoS One. 10:e01312252015.
View Article : Google Scholar
|
|
31
|
Lingala S, Cui YY, Chen X, Ruebner BH,
Qian XF, Zern MA and Wu J: Immunohistochemical staining of cancer
stem cell markers in hepatocellular carcinoma. Exp Mol Pathol.
89:27–35. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu JW, Chang JG, Yeh KT, Chen RM, Tsai JJ
and Hu RM: Overexpression of Thy1/CD90 in human hepatocellular
carcinoma is associated with HBV infection and poor prognosis. Acta
Histochem. 113:833–838. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu XH, Xu LB, Liu C, Zhang R and Wang J:
Clinicopathological characteristics of 20 cases of hepatocellular
carcinoma with bile duct tumor thrombi. Dig Dis Sci. 56:252–259.
2011. View Article : Google Scholar
|
|
34
|
Ho DW, Yang ZF, Yi K, Lam CT, Ng MN, Yu
WC, Lau J, Wan T, Wang X, Yan Z, et al: Gene expression profiling
of liver cancer stem cells by RNA-sequencing. PLoS One.
7:e371592012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang R, An LY, Miao QF, Li FM, Han Y, Wang
HX, Liu DP, Chen R and Tang SQ: Effective elimination of liver
cancer stem-like cells by CD90 antibody targeted thermosensitive
magnetoliposomes. Oncotarget. 7:35894–35916. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han D, Li J, Wang H, Su X, Hou J, Gu Y,
Qian C, Lin Y, Liu X, Huang M, et al: Circular RNA circMTO1 acts as
the sponge of miR-9 to suppress hepatocellular carcinoma
progression. Hepatology. 66:1151–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z,
Yang J, Fan J, Liu L and Qin W: Hsa_circ_0001649: A circular RNA
and potential novel biomarker for hepatocellular carcinoma. Cancer
Biomark. 16:161–169. 2016. View Article : Google Scholar
|
|
38
|
Yao Z, Luo J, Hu K, Lin J, Huang H, Wang
Q, Zhang P, Xiong Z, He C, Huang Z, et al: ZKSCAN1 gene and its
related circular RNA (circZKSCAN1) both inhibit hepatocellular
carcinoma cell growth, migration, and invasion but through
different signaling pathways. Mol Oncol. 11:422–437. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fulda S: Modulation of mitochondrial
apoptosis by PI3K inhibitors. Mitochondrion. 13:195–198. 2013.
View Article : Google Scholar
|
|
40
|
Fulda S: Synthetic lethality by
co-targeting mitochondrial apoptosis and PI3K/Akt/mTOR signaling.
Mitochondrion. 19:85–87. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gedaly R, Angulo P, Hundley J, Daily MF,
Chen C and Evers BM: PKI-587 and sorafenib targeting PI3K/AKT/mTOR
and Ras/Raf/MAPK pathways synergistically inhibit HCC cell
viability. J Surg Res. 176:542–548. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pan Y, Zhang Y, Chen L, Liu Y, Feng Y and
Yan J: The critical role of Rab31 in cell viability and apoptosis
in cancer progression. Mol Neurobiol. 53:4431–4437. 2016.
View Article : Google Scholar
|
|
43
|
Sui Y, Zheng X and Zhao D: Rab p31
promoted hepatocellular carcinoma (HCC) progression via inhibition
of cell apoptosis induced by PI3K/AKT/Bcl-2/BAX pathway. Tumor
Biol. 36:8661–8670. 2015. View Article : Google Scholar
|
|
44
|
Liang M, Liu J, Ji H, Chen M, Zhao Y, Li
S, Zhang X and Li J: A Aconitum coreanum polysaccharide fraction
induces apoptosis of hepatocellular carcinoma (HCC) cells via
pituitary tumor transforming gene 1 (PTTG1)-mediated suppression of
the P13K/Akt and activation of p38 MAPK signaling pathway and
displays antitumor activity in vivo. Tumour Biol. 36:7085–7091.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng J, Li C, Wu X, Liu M, Sun X, Yang Y,
Hao M, Sheng S, Sun Y, Zhang H, et al: Astrocyte elevated gene-1
(AEG-1) shRNA sensitizes Huaier polysaccharide (HP)-induced
anti-metastatic potency via inactivating downstream P13K/Akt
pathway as well as augmenting cell-mediated immune response. Tumour
Biol. 35:4219–4224. 2014. View Article : Google Scholar : PubMed/NCBI
|