Introduction
Oxidative stress is one of the main causes of
neuronal injury in neurodegenerative disorders, involving different
signaling pathways (1). Evidence
declares that reactive oxygen species (ROS) or hydrogen peroxide
(H2O2), a main precursor of ROS, induces
neuronal cell death contributing to the pathogenesis in
neurodegenerative disorders such as Alzheimer's and Parkinson's
diseases and stroke (2). Neuronal
cells respond to H2O2 with cytotoxicity and
neurodegeneration, whereas antioxidants may abolish these effects
(3). Notably, mitochondria is the
major source of ROS and mitochondrial dysfunction has been
recognized as a critical contributor to the neurodegenera-tive
disorders (4). Furthermore, the
antioxidant defense system serves vital role in regulating
oxidative stress, and Chinese medicinal herbs provide rich
resources for developing natural antioxidants (5). Therefore, it is important to
elucidate antioxidants from natural products and herbal medicine
for treating neurodegenerative disorders.
Lithocarpus polystachyus Rehd. is distributed
as a wild plant in the mountain area in Southern China, especially
in Shiqian and Fenggang cities of the Guizhou Province with
plentiful natural resources (6).
It is also called ‘Sweet Tea’ (ST) and its leaves are used as a
medicinal tea and a folk herbal medicine to prevent or cure
multiple diseases for many years and exhibited no adverse drug
reactions or toxic effects (7).
Accumulating evidence demonstrates that ST has extensive
pharmacological activities, including anti-hypertensive,
anti-hypoglycemic, anti-diabetic and anti-obesity pharmacological
effects (7-9). However, little is known about
whether ST possesses protective effects on the oxidative
stress-induced neuronal cell injury.
AMP-activated protein kinase (AMPK) is known as a
vital intracellular serine/threonine protein kinase that regulates
celluar energy metabolism and responses to metabolic stress
(10). Additionally,
AMPK-activated peroxisome proliferator-activated receptor
coactivator-1α (PGC-1α) exerts crucial effect in the regulation of
mitochondrial biogenesis and oxidative stress (11). Sirtuins belong to class III
histone deacetylases family and their enzymatic activities are
dependent on nicotinamide adenine dinucleotide, and seven mammalian
homologues of sirtuins (Sirt1-7) have been identified (12). Among those sirtuins, Sirt3 has
been demonstrated to control mitochondrial oxidative signaling, and
serves as the downstream target of AMPK-PGC-1α signaling pathway
(13,14). Additionally, a previous study
reported that Sirt3 activation prevents neuronal mitochondrial
oxidative stress and damage in neurodegenerative disorders
(15). However, whether ST
regulates neuronal Sirt3 and its corresponding upstream and
downstream regulatory signaling pathways remain unclear.
Therefore, the present study was designed to
determine the protective effect of ST on SH-SY5Y cell injury
induced by H2O2, and to further elucidate its
underlying mechanisms.
Materials and methods
Materials
Dulbecco's modified Eagle's medium: Nutrient Mixture
F-12 (DMEM/F-12) (cat. no. 11320082), fetal bovine serum (FBS; cat.
no. 1618862) and a penicillin/streptomycin mixture (cat. no.
15070-063) were obtained from Gibco; Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). 3-(4,5-dimethyl thiazol-2-yl)-2,5-diphenyl
tetrazolium bromide (MTT; cat. no. M2128), 2′,7′-dichlorofluorescin
diacetate (DCFH-DA; cat. no. D6883), and rhodamine 123 (R8004) were
obtained from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany).
MitoSOX Red Mitochondrial Superoxide Indicator (M36008) and Mito
Tracker green probe (M7514) were obtained from Invitrogen; Thermo
Fisher Scientific, Inc. The lactate dehydrogenase (LDH; cat. no.
A020-2), malondialdehyde (MDA; cat. no. A003-1), superoxide
dismutase (SOD; cat. no. A001-3), reduced glutathione (GSH; cat.
no. A006-2) and glutathione peroxidase (GSH-Px; cat. no. A005)
assay kits were purchased by Nanjing Jiancheng Bioengineering
Institute (Nanjing, China). Anti-adenosine monophosphate-activated
protein kinase (AMP; cat. no. ab131512), anti-p-AMPK (cat. no.
ab23875), anti-peroxisome proliferator-activated receptor
coactivator-1α (PGC-1α; cat. no. ab54481), anti-Sirt3 (cat. no.
ab217319), anti-isocitrate dehydrogenase (IDH2; cat. no. ab84726),
anti-forkhead boxO3a (Foxo3a; cat. no. ab12162) and anti-SOD2 (cat.
no. ab13533) antibodies were obtained from purchased from Abcam
(Cambridge, UK). All other chemicals were of analytical grade.
Preparation of ST aqueous extracts
Fresh leaves of ST were obtained from Fenggang City
in Guizhou Province in summer and identified by Professor Jianwen
Yang (School of Pharmacy, Zunyi Medical University, Zunyi, China).
A voucher specimen was deposited at the herbarium of School of
Pharmacy, Zunyi Medical University (voucher number, 20151016ST).
From 1 kg dried ST, 100 g of dried aqueous extract was obtained
(yield 10%) and filtered. Subsequently, the filter liquor were
condensed by rotary vacuum evaporation and lyophilized. The final
powder was dissolved in double-distilled water, sterilized by
passing through 0.22 µm filters, and attenuated in DMEM/F-12
culture solution prior to employ at a final concentration of 25, 50
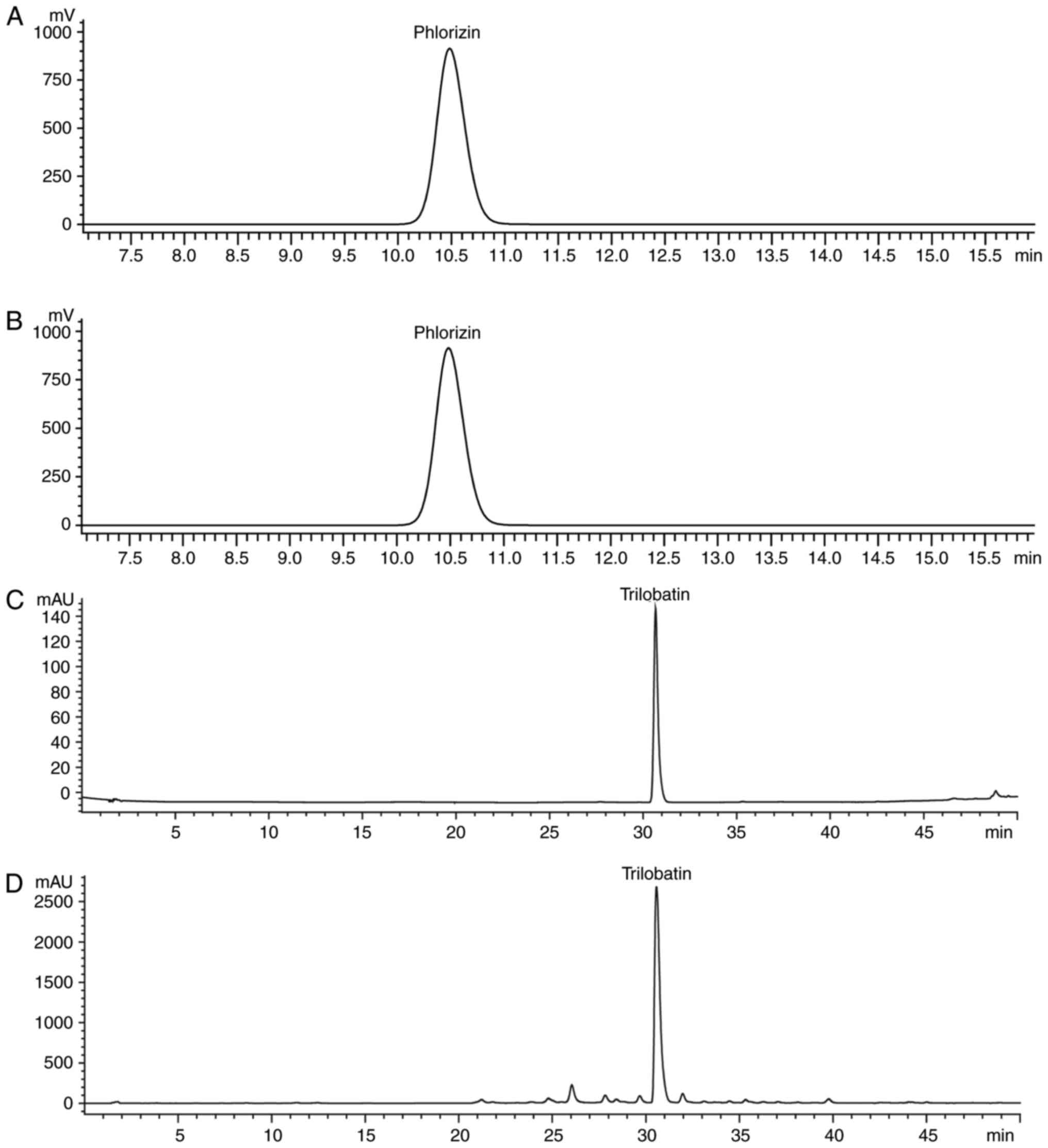
or 75 µg raw herb/ml. ST was standardized by the content of
phlorizin and trilobatin, which are the active components of ST
(16). Phlorizin and trilobatin
(Chengdu Push Bio-Technology Co., Ltd., Chengdu, China) were
determined by reverse-phase high-performance liquid chromatography
(HPLC; Agilent 1200 series; Agilent Technologies, Inc., Santa
Clara, CA, USA) with a UV detector, auto-sampler, column oven, and
multi-channel pump. Subsequently, the separation was executed by a
Phenomenex Luna end-capped C18 column (5 mm, 4.6x250 mm;
Phenomenex, CA, USA) at 25°C. The detector wavelength was set at
285 nm. The separated conditions of phlorizin were as follows:
Acetonitrile: Water (30:70, v/v) as mobile phase using a linear
gradient at a flow rate of 1.0 ml/min. The ST contained 1.19%
phlorizin (Fig. 1A and B). The
separated conditions of trilobatin were as follows: Acetic acid
water: Methanol (40:60, v/v) as mobile phase using a linear
gradient at a flow rate of 1.0 ml/min. The ST contained 18.54%
trilobatin (Fig. 1C and D).
Cell culture
The human neuroblastoma SH-SY5Y cells were purchased
from American Type Culture Collection (Manassas, VA, USA) and
maintained in DMEM/F-12 supplemented with 10% FBS and 0.1%
penicillin/streptomycin at 37°C in a humidified incubator
containing 5% CO2 and 95% air.
MTT assay
Effect of ST on SH-SY5Y cell viability were
determined using an MTT assay. In brief, SH-SY5Y cells were
cultured in 96-well plates, at a density of 2x104 cells
per well for 24 h at 37°C. Subsequently, the cells were pre-treated
with various concentrations (25, 50 and 75 µg/ml) of ST
aqueous extracts or NAC for 1 h and co-treatment with 200 µM
H2O2 for the further 24 h at 37°C. Following
the treatment, MTT solution was added to each well and the cells
were cultured for 4 h at 37°C. The dark-blue formazan crystals were
dissolved in DMSO, and the absorbance values were detected at 490
nm wavelength by microplate reader. In addition, according to our
primary study, 20 µM NAC was chosen to be a positive control
for antioxidant capacity that suppresses cell death for comparing
with the possible protective effect of ST (17).
LDH leakage assay
The SH-SY5Y cells (2x104 cells per well)
were pre-treated with or without ST as described above and the
supernatant was analyzed by a LDH detection kit, according to the
manufacturer's protocol. The leakage of LDH was determined at 490
nm wavelength using a microplate reader. At the same time, cellular
morphologic changes were observed from five random fields using a
phase contrast microscope (Olympus IX73; Olympus, Tokyo, Japan;
magnification, x200).
Evaluation of MDA and GSH content, GPx
and SOD activities
In brief, SH-SY5Y cells were treated as described
above. MDA and GSH content, SOD and GSH-Px activities were detected
by MDA, GSH, GSH-Px and SOD kits, respectively.
Measurement of intracellular and
mitochondrial ROS accumulation
Intracellular ROS production was measured using
DCFH-DA dye. Following SH-SY5Y cells were treated for 24 h as
described above, 20 µM DCFH-DA dye treated cells for 30 min
at 37°C in the dark. Subsequently, cells were washed with PBS, and
intracellular ROS generation was measured using a fluorescence
reader at excitation/emission wavelengths of 485/530 nm (Varioskan
Flash Multimode Reader; Thermo Fisher Scientific, Inc.).
Additionally, mitochondrial ROS was determined using MitoSOX Red, a
fluorescent indicator for mitochondrial superoxide (18). In brief, SH-SY5Y cells were
treated as described above and washed with balanced salt solution
(Gibco). MitoSOX Red dye was used at a concentration of 5 µM
for 20 min in the dark at 37°C. Subsequently, cells were washed
using PBS and labeled with Mito Tracker green probe (200 nM) and
stained for 20 min. Finally, cells were washed with PBS and
fluorescence was observed over the whole field of vision using
fluorescence microscopy (Olympus IX73; Olympus Corporation, Tokyo,
Japan; magnification, x200) with excitation/emission (510/580 nm)
filters. The mean fluorescence intensity (MFI) of MitoSOX Red from
five random fields was calculated using ImageJ 1.47i software, and
the MFI was applied as an index of the levels of mitochondrial
ROS.
Measurement of mitochondrial membrane
potential (MMP)
Measurement of MMP was detected using rhodamine 123
dye, a reliable indicator of MMP, as reported in previous studies
(19,20). Briefly, SH-SY5Y cells were treated
as described above. Subsequently, cells were stained using
rhodamine with 2 µg/ml 123 dye for 20 min at 37°C in the
dark, and cells were washed with PBS. Subsequently, fluorescence
was observed over the whole field of vision using fluorescence
microscopy (Olympus IX73; Olympus Corporation; magnification, x200)
with excitation/emission (485/595 nm) filters. The MFI of Rh123
from five random fields was calculated using ImageJ 1.47i software,
and the MFI was applied as an index of the levels of MMP.
NAD+/NADH measurement
The SH-SY5Y cells were treated as described above.
By the end of treatment, NAD+/NADH ratio were measured
using a NAD/NADH Quantification kit (cat. no. ab65348; Abcam)
according to the manufacture's protocol, and absorbance was
measured at 450 nm using the Varioskan Flash Multimode Reader.
Western blot analysis
Following pre-treatment with or without various
concentrations (25, 50 and 75 µg/ml) of ST on SH-SY5Y cells
for 1 h at 37°C prior to co-culture with 200 µM
H2O2. Following 24 h incubation, cells were
lysed in the lysis buffer (cat. no. 9803; Cell Signaling
Technology), and protein concentrations were quantified using the
BCA protein assay kit (cat. no. 7780; Cell Signaling Technology,
Inc.). Subsequently, samples containing 20 µg proteins were
subjected to 10% SDS-polyacrylamide gel, and transferred to a PVDF
membrane (cat. no. 1620177; Bio-Rad Laboratories). Following
blocking with 5% non-fat milk for 1 h at room temperature, the
membrane was incubated overnight at 4°C with the appropriate
primary antibodies: anti-p-AMPK (1:1,000), anti-AMPK (1:1,000),
anti-PGC-1α (1:1,000), anti-Sirt3 (1:1,000), anti-IDH2 (1:1,000),
anti-Foxo3a (1:1,000), anti-SOD2 (1:1,000). Subsequently, the
membranes were washed with TBS with Tween-20, and incubated with
appropriate HRP-conjugated goat anti-rabbit antibody (cat. no.
A0208, Beyotime) and HRP-conjugated goat anti-rat antibody (cat.
no. ab97057; Abcam) for 1 h at room temperature under shaking. The
bonds of protein were developed with the ECL Western blot reagent
(cat. no. P0018, Beyotime), and the blots was visualized using
Davinch-Chemi™ imaging system and band optical intensity was
quantified using Quantity One 1-D analysis software v4.52 (BioRad
Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All data were confirmed by at least three
independent experiments and were expressed as the mean ± standard
deviation. Statistical analyses were performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA), and individual differences
were determined using one-way analysis of variance, followed by the
Fisher's least significant difference. P<0.05 was considered to
indicate a statistically significant difference.
Results
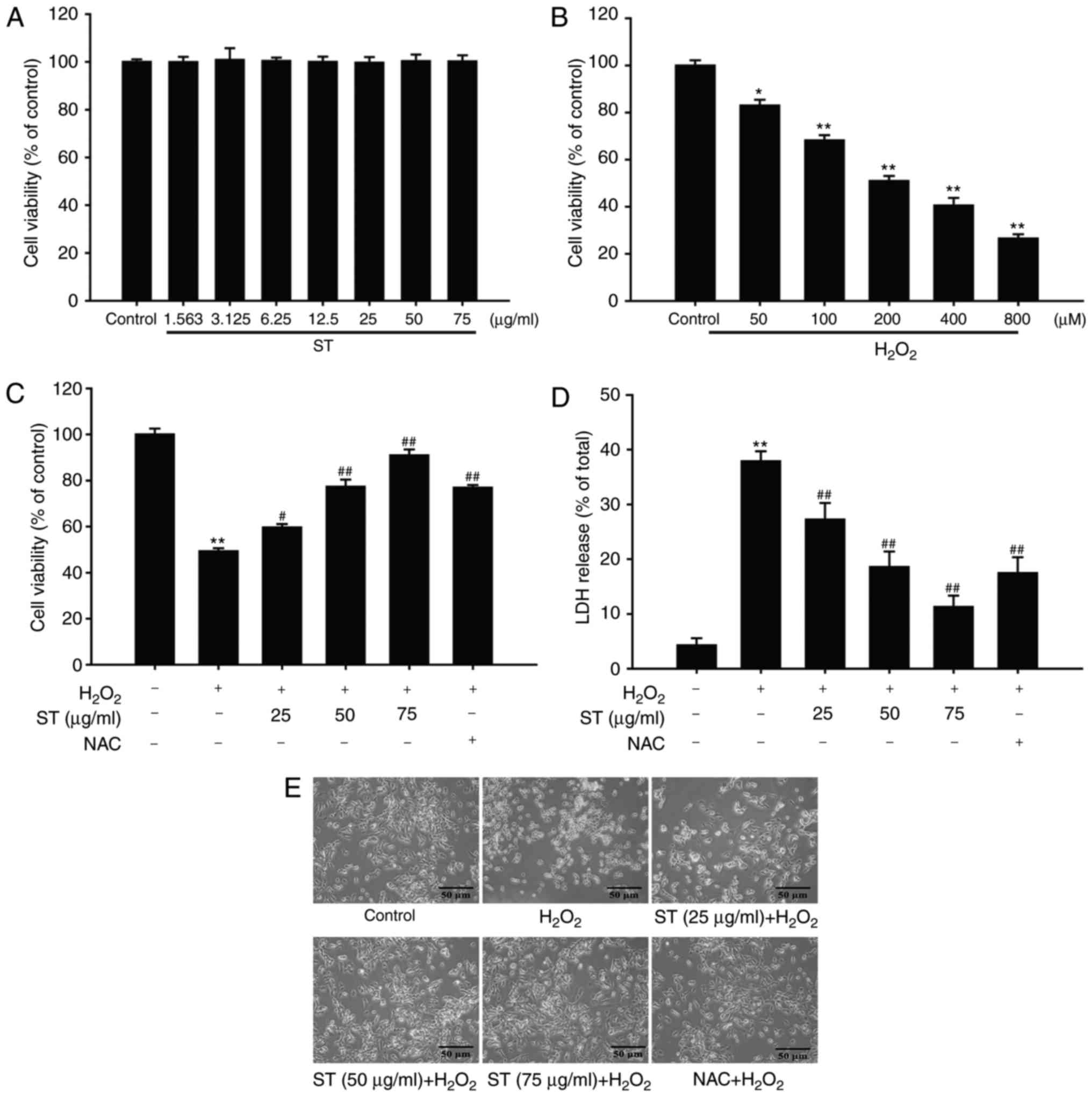
ST protects SH-SY5Y cells against
H2O2-induced cell injury
SH-SY5Y cells were treated with various
concentrations of ST (3.125, 6.25, 12.5, 25, 50, and 75
µg/ml) for 24 h, and cell viability was subsequently
detected using the MTT assay. Treatment with these concentrations
of ST alone did not affect cell viability and caused no cell
toxicity [F(7,16)=0.027, P=1.000; Fig. 2A]. Subsequently, the effect of
H2O2 (50-800 µM) on SH-SY5Y cells was
evaluated. The results indicated that H2O2
reduced cell viability in a concentration-dependent manner, and 200
µM H2O2 incubation with SH-SY5Y cells
for 24 h was confirmed to be an appropriate condition to taken as
an oxidative stress-induced injury in SH-SY5Y cellular model in
vitro with approximately 50% cell viability inhibition rate [F
(5,12)=200.045, P<0.001; Fig. 2B]. To investigate the protective
effect of ST against H2O2-induced SH-SY5Y
cell injury, SH-SY5Y cells were pre-treated with ST at different
concentrations (25, 50, and 75 µg/ml) for 1 h, the
co-treatment with 200 µM H2O2 for
another 24 h. The results demonstrated that cell viability
increased to 59.67, 77.33, 91.01 or 76.93% when cells were
pre-treated with ST (25, 50, and 75 µg/ml) or NAC,
respectively [F(5,12)=149.1724, P<0.001; Fig. 2C]. In parallel, 200 µM
H2O2 significantly increased LDH release to
37.88%. However, pre-treatment with ST or NAC, reduced the amount
of LDH release to 27.23, 18.55, 11.29 or 17.53% respectively
[F(5,12)=75.218, P<0.001; Fig. 2D]. At the meantime, this
protective effect of ST was also evidenced by morphologic
observations of SH-SY5Y cells. In the case of
H2O2-treated cells, most cells demonstrated
shrinkage and floatation. However, the SH-SY5Y cells were
pre-treated with ST or NAC displayed slightly morphological changes
(Fig. 2E). These findings
indicated that the protective effects of ST were equal or higher
than those exerted by an equivalent concentration of positive
control agent.
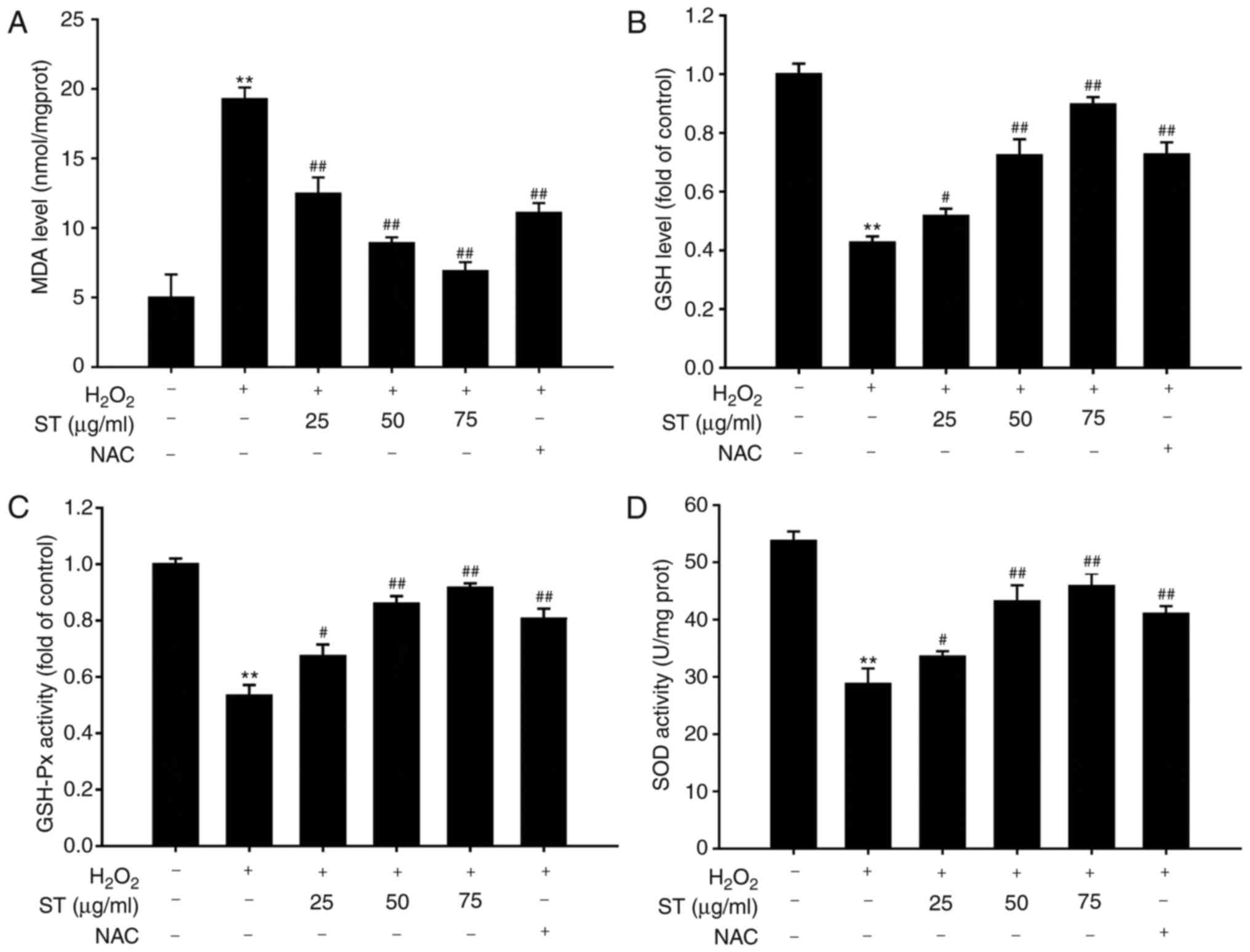
ST attenuated
H2O2-induced accumulation of MDA, enhanced
GSH content, GSH-Px, and SOD activities
The results indicated that
H2O2 caused a remarkable increase in MDA
content and decrease in GSH content, SOD and GSH-Px activities.
However, pre-treatment with ST (25, 50, and 75 µg/ml)
significantly reduced the increase in MDA content induced by
H2O2 [F(5,12)=77.225, P<0.001; Fig. 3A], while it significantly reversed
the decrease in GSH content, GSH-Px and SOD activities induced by
H2O2 treatment in a concentration-dependent
manner [F(5,12)=131.886, P<0.001; F(5,12)=57.409, P<0.001; F(5,12)=97.466, P<0.001, respectively;
Fig. 3B-D].
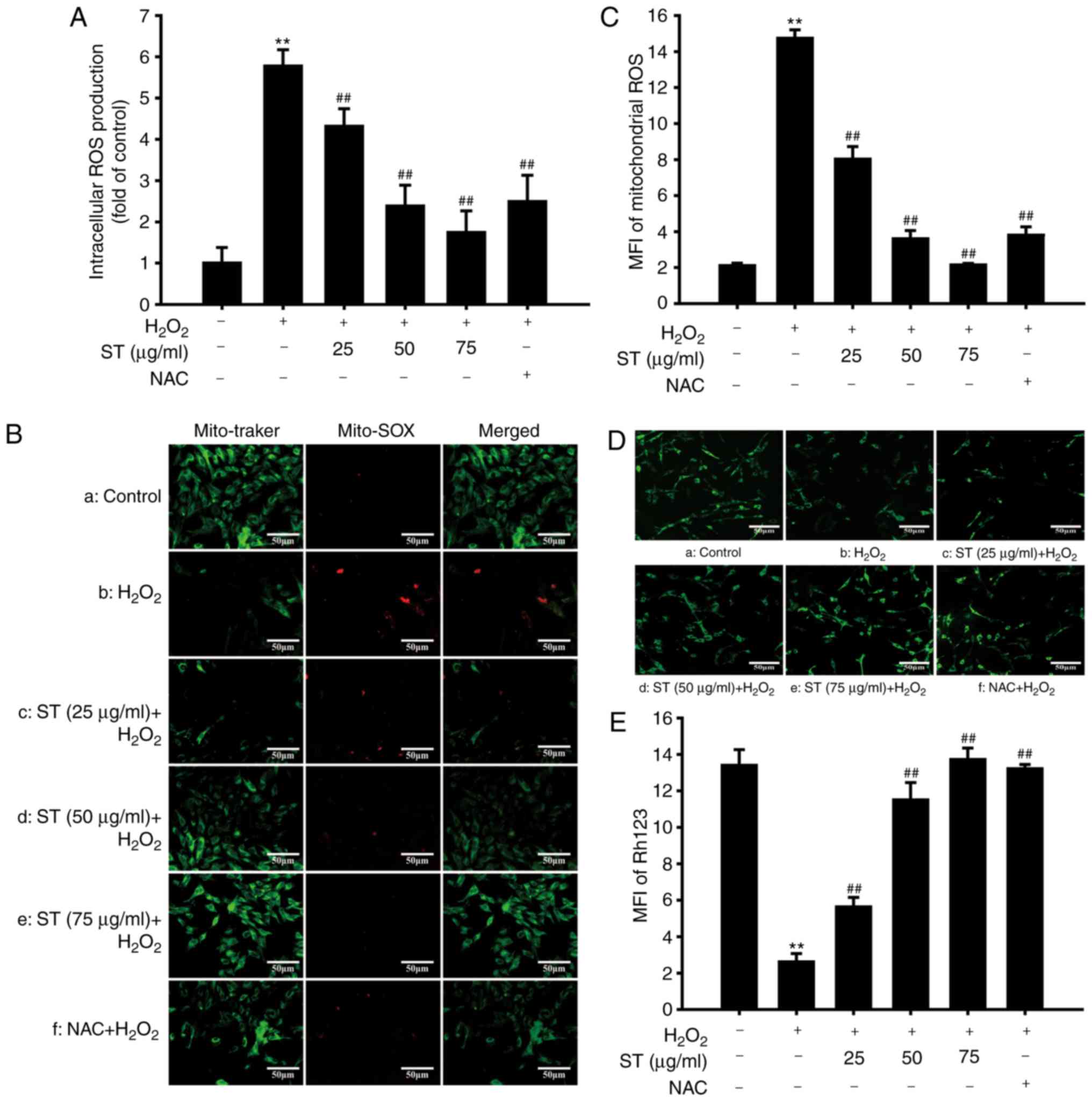
ST attenuated
H2O2-induced accumulation of intracellular
and mitochondrial ROS production, improved MMP
Effects of ST on intracellular, mitochondrial ROS
production, and MMP were evaluated. The results demonstrated that
following treatment with H2O2, the levels of
intracellular ROS increased markedly within 24 h. Pre-treatment
with ST (25, 50, and 75 µg/ml) suppressed
H2O2-induced excessive ROS production
[F(5,12)=154.603, P<0.001; Fig. 4A]. Furthermore, to determine the
level of oxidative stress in mitochondria during
H2O2 treatment, SH-SY5Y cells were stained
with MitoSOX, a cationic probe that distributes to the
mitochondrial matrix and specifically measures superoxide anion,
and co-localizes with Mito Tracker Green (21). Compared with the control,
H2O2 notably accelerated the accumulation of
mitochondrial ROS which was decreased by ST pre-treatment
[F(5,12)=382.644, P<0.001; Fig. 4B and C]. Furthermore, MMP
indicates the function of mitochondria (22). H2O2
decreased rhodamine 123 fluorescence intensity, while this decrease
in rhodamine 123 fluorescence was abrogated by pre-treatment with
ST or NAC [F(5,12)=156.453, P<0.001; Fig. 4D and E]. These results
demonstrated that ST effectively inhibited ROS levels and
attenuated reduction in MMP induced by H2O2
in SH-SY5Y cells.
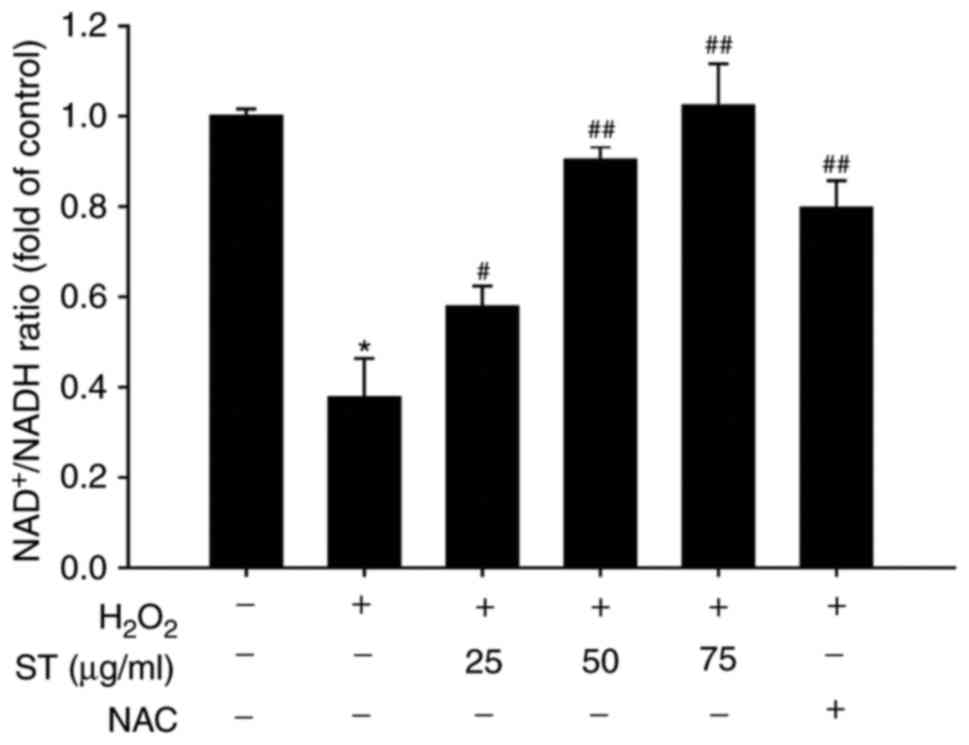
ST increased NAD+/NADH ratio,
AMPK, PGC-1α, Sirt3, IDH2, Foxo3a, SOD2 in SH-SY5Y cells
The results demonstrated that
H2O2 decreased the ratio of
NAD+/NADH, while pre-treated with ST increased the ratio
of NAD+/NADH in SH-SY5Y cells [F(5,12)=91.125, P<0.001; Fig. 5]. AMPK-PGC-1α signaling serves an
important role in regulating mitochondrial function and oxidative
stress and has been reported to serve as the downstream target of
AMPK-PGC-1α signaling (23).
Therefore, the effects of ST on the protein expressions of AMPK,
PGC-1α, Sirt3, IDH2, Foxo3a, SOD2 in
H2O2-induced SH-SY5Y cells evaluated, using
western blot analysis. The results demonstrated that 200 µM
H2O2 decreased the ratio of p-AMPK/AMPK and
the expression of PGC-1α and Sirt3. However, ST pre-treatment
prominently elevated the p-AMPK/AMPK ratio and the expression of
PGC-1α and Sirt3 [F(5,12)=29.286, P<0.001; F(5,12)=9.640, P<0.001; F(5, 12)=14.183,
P<0.001; Fig. 6A-D],
indicating that the AMPK-PGC-1α-Sirt3 axis was crucial for the
protective effects of ST on regulation of mitochondrial ROS
homeostasis in SH-SY5Y cells. Considering that Sirt3 is an
NAD+-dependent deacetylase, and IDH2, Foxo3a and SOD2
are major substrates of Sirt3. IDH2, Foxo3a and SOD2 expressions
were also investigated. The results showed that ST pre-treatment
notably increased IDH2, Foxo3a and SOD2 expression compared with
H2O2 treatment [F(5,12)=11.078, P<0.001; F(5,12)=20.023, P<0.001; F(5,12)=15.505, P<0.001; Fig. 6A and E-G], suggesting that ST
ameliorated H2O2 induced mitochondrial
dysfunction and reduced oxidative stress were mediated by the
activation of AMPK-PGC-1α-Sirt3 signaling.
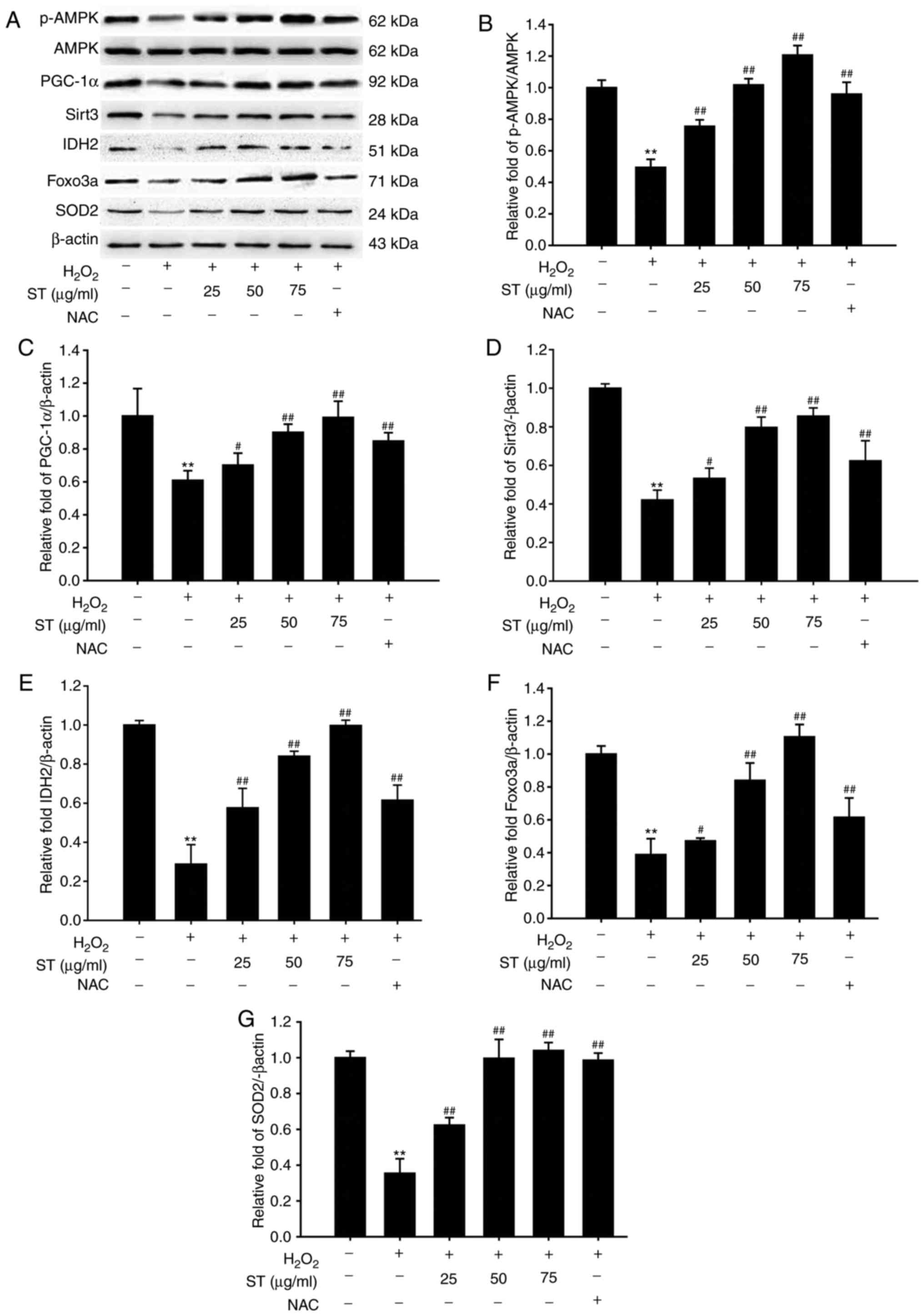 | Figure 6ST stimulates Sirt3 signaling pathway
in SH-SY5Y cells. (A) Western blot analysis of AMPK, p-AMPK,
PGC-1α, Sirt3, IDH2, Foxo3a, and SOD2 and quantification of (B)
p-AMPK/AMPK ratio; (C) PGC-1α; (D) Sirt3; (E) IDH2; (F) Foxo3a and
of (G) SOD2 protein expression levels. Data are shown as mean ±
standard error of the mean (n=3). *P<0.05,
**P<0.01 vs. untreated control cells;
#P<0.05, ##P<0.01 vs.
H2O2-treated cells. AMPK, adenosine
monophosphate-activated protein kinase; Foxo3a, forkhead box O3a;
IDH2, isocitrate dehydrogenase; p, phosho; PGC-1α, peroxisome
proliferator-activated receptor coactivator-1α; ST, sweet tea. |
Discussion
Excessive oxidative stress has been demonstrated to
serve a crucial role in neurodegenerative diseases (24). H2O2 not only
is the most steady ROS but also serves as extracellular and
intercellular messenger (25,26). Therefore, in the present study,
H2O2 was used as a toxic agent to simulate
oxidative stress-induced neuronal injury in vitro, thereby
leading to mitochondrial injury as described in our previous study
(27). The results demonstrated
that H2O2-induced neuronal cell injury as
evidenced by decreasing cell viability and increasing cell death,
combined with the repression of MMP and generation of excessive ROS
in the mitochondria, which were significantly reversed by
pre-treatment with ST. The present findings, for the first time to
the best of our knowledge, clarified that ST exerted the protective
effects on oxidative stress-induced neuronal cells injury.
Furthermore, emerging evidence demonstrates that
mitochondria are the major resource of ROS, and the mitochondrial
ROS free radical scavenging systems, including SOD2 and the
glutathione and thioredoxin systems, serves an key role in
regulating mitochondrial ROS (28). In the present study,
H2O2 decreased the enzymatic activities of
SOD2, GSH-Px and IDH2, while these effects were reversed by
pre-treatment of ST. Furthermore, MitoSOX Red, a cationic probe
that distributes to the mitochondrial matrix and specifically
detects superoxide, was used to determine the mitochondrial ROS as
described in our previous study (27). The results suggested that ST
scavenged excessive mitochondrial ROS via elevating ROS free
radical scavenging systems, thereby promoted GSH levels and
maintained NAD+/NADH ratio, which acts as an important
role in donating electrons when NADH converts into NAD+,
and eliminating excessive ROS (29). In addition, sirtuins are
NAD+-dependent enzymes, which include seven homologues
(Sirt1-7), and they deacetylate multiple substrates to execute
numerous physiological and pathophysiological functions (30). Sirt3 is known as an important
member of the sirtuin family, and it not only localizes in
mitochondria but also suppresses the excessive ROS production due
to deacetylating many mitochondrial proteins such as SOD2, IDH2,
which improve the glutathione antioxidant defense system (31). Furthermore, the present findings
indicated that H2O2 decreased Sirt3, Foxo3a
and SOD2 expressions, which was consistent with the theory that
oxidative stress downregulates Foxo3a and SOD2, which are the two
main substrates of Sirt3 (32,33). However, these effects were
abolished by pre-treatment with ST, suggesting that ST activated
Sirt3, thereby regulating Foxo3a directly and elevating SOD2
activity, which is mainly regulated by the mitochondrial sirtuin to
eliminate excessive ROS (34).
Additionally, growing evidence demonstrated that
Sirt3 serves as a downstream target of PGC-1α, which has been
demonstrated as the pivotal downstream target of AMPK, thereby
activating Foxo3a to reduce mitochondrial ROS production (35,36). It is also postulated that
decreased AMPK-PGC-1α signaling pathway may lead to downregulation
of Sirt3 expression (37). The
results in the current study demonstrated that
H2O2 decreased p-AMPK and PGC-1α expression,
which was consistent with the theory that oxidative stress inhibits
AMPK, and downregulates PGC-1α and Sirt3 expression. However,
pre-treatment with ST reversed these effects, suggesting that ST
preserved mitochondrial function and attenuated excessive ROS
through activating the phosphorylation of AMPK, thereby
upregulating PGC-1α and Sirt3. Therefore, it can be concluded that
the AMPK-PGC-1α-Sirt3 signaling pathway serves a crucial role in
regulating mitochondrial ROS homeostasis by ST pre-treatment in
oxidative stress-induced neuronal cell injury.
To conclude, the present findings indicated that ST
protects against oxidative stress-induced neuronal cell injury
in vitro through attenuating mitochondrial ROS, at least
partly, through activating Sirt3 signaling pathway (Fig. 7). These results provide valuable
insights for the role of ST, which may lead to its development as a
natural antioxidant to protect neurode-generative diseases, but
this needs to be further investigated.
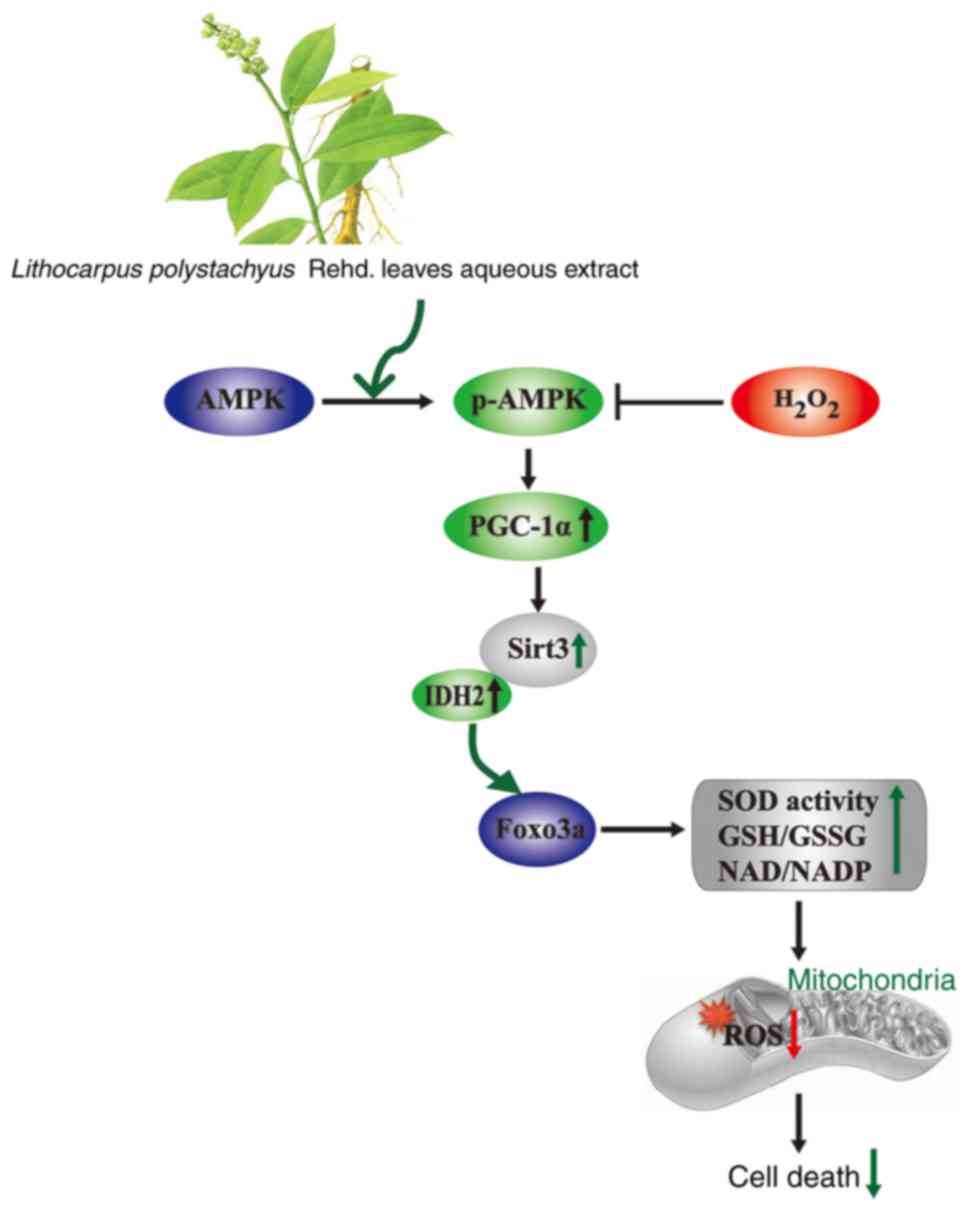 | Figure 7Model of a proposed mechanism of the
protective effect of on ST SH-SY5Y cells against
H2O2-induced SH-SY5Y cells injury through
activation of AMPK-PGC-1axα-Sirt3 axis. H2O2
is considered as the main precursor of ROS, and accumulation of
intracellular ROS cause mitochondrial dysfunction, downregulation
of AMPK, PGC-1α and Sirt3, leading to cell death. ST reduces
mitochondrial ROS production and cell death induced by
H2O2 via mitochondrial Sirt3 signaling
pathway. AMPK, adenosine monophosphate-activated protein kinase;
Foxo3a, forkhead box O3a; IDH2, isocitrate dehydrogenase; p,
phosho; PGC-1α, peroxisome proliferator-activated receptor
coactivator-1α; ST, sweet tea. |
Acknowledgements
Not applicable.
Funding
The present study was supported by Natural Science
Foundation of China (grant no. 81560666), Program for excellent
young talents of Zunyi Medical Uiverstity (grant no. 15zy-002).
Science and Technology Innovation Talent Team of Guizhou Province
(grant no. 20154023). The ‘hundred’ level of high-level innovative
talents in Guizhou Province (grant no. QKHRCPT 20165684), and
Program for Changjiang Scholars and Innovative Research Team in
University, China (grant no. IRT17R113).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
JG performed MTT assay, LDH leakage assay, MDA, GSH
content, GPx, SOD activities analysis, ROS production, MMP analysis
and was major contributor in writing the manuscript. ST aqueous
extracts experiment was conducted by YX. JZ performed the western
blot analysis. JS and QG designed the experimental approaches. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
Abbreviations:
|
ROS
|
reactive oxygen species
|
|
DCFH-DA
|
2′,7′-dichlorofluorescin diacetate
|
|
MDA
|
malondialdehyde
|
|
SOD
|
superoxide dismutase
|
|
GSH
|
reduced glutathione
|
|
GSH-Px
|
glutathione peroxidase
|
|
AMPK
|
adenosine monophosphate-activated
protein kinase
|
|
PGC-1α
|
peroxisome proliferator-activated
receptor coactivator-1α
|
|
Foxo3a
|
forkhead box O3a
|
References
|
1
|
Liu L, Zhang K, Sandoval H, Yamamoto S,
Jaiswal M, Sanz E, Li Z and Hui J: Glial lipid droplets and ROS
induced by mitochondrial defects promote neurodegeneration. Cell.
160:177–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang JX, Wang R, Xi J, Shen L, Zhu AY, Qi
Q, Wang QY, Zhang LJ, Wang FC, Lü HZ and Hu JG: Morroniside
protects SK-N-SH human neuroblastoma cells against
H2O2-induced damage. Int J Mol Med.
39:603–612. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanschmann EM, Godoy JR, Berndt C,
Hudemann C and Lillig CH: Thioredoxins, glutaredoxins, and
peroxire-doxins-molecular mechanisms and health significance: From
cofactors to antioxidants to redox signaling. Antioxid Redox
Signal. 19:1539–1605. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tseng AH, Shieh SS and Wang DL: SIRT3
deacetylates FOXO3 to protect mitochondria against oxidative
damage. Free Radic Biol Med. 63:222–234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Y, Wang Y, Li J, Hua L, Han B, Zhang
Y, Yang X, Zeng Z, Bai H, Yin H and Lou J: Effects of caffeic acid
on learning deficits in a model of Alzheimer's disease. Int J Mol
Med. 38:869–875. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang J, Huang Y, Li K, Chen Y, Vanegas D,
McLamore ES and Shen Y: Leaf extract from Lithocarpus polystachyus
Rehd promote glycogen synthesis in T2DM mice. PLoS One.
11:e01665572016. View Article : Google Scholar
|
|
7
|
Hou SZ, Chen SX, Huang S, Jiang DX, Zhou
CJ, Chen CQ, Liang YM and Lai XP: The hypoglycemic activity of
Lithocarpus polystachyus Rehd leaves in the experimental
hyperglycemic rats. J Ethnopharmacol. 138:142–149. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hou SZ, Xu SJ, Jiang DX, Chen SX, Wang LL,
Huang S and Lai XP: Effect of the flavonoid fraction of Lithocarpus
polys-tachyus Rehd on spontaneously hypertensive and normotensive
rats. J Ethnopharmacol. 143:441–447. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou CJ, Huang S, Liu JQ, Qiu SQ, Xie FY,
Song HP, Li YS, Hou SZ and Lai XP: Sweet tea leaves extract
improves leptin resistance in diet-induced obese rats. J
Ethnopharmacol. 145:386–392. 2013. View Article : Google Scholar
|
|
10
|
Gao XY, Wang SN, Yang XH, Lan WJ, Chen ZW,
Chen JK, Xie JH, Han YF, Pi RB and Yang XB: Gartanin protects
neurons against glutamate-induced cell death in HT22 cells:
Independence of Nrf-2 but involvement of HO-1 and AMPK. Neurochem
Res. 41:2267–2277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng L, Li B, Chen X, Su J, Wang H, Yu S
and Zheng Q: CTRP9 induces mitochondrial biogenesis and protects
high glucose-induced endothelial oxidative damage via
AdipoR1-SIRT1-PGC-1α activation. Biochem Biophys Res Commun.
477:685–691. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kincaid B and Bossy-Wetzel E: Forever
young: SIRT3 a shield against mitochondrial meltdown, aging, and
neurodegeneration. Front Aging Neurosci. 5:482013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guan Y, Cui ZJ, Sun B, Han LP, Li CJ and
Chen LM: Celastrol attenuates oxidative stress in the skeletal
muscle of diabetic rats by regulating the AMPK-PGC1α-SIRT3
signaling pathway. Int J Mol Med. 37:1229–1238. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Q, Li L, Li CY, Pei Z, Zhou M and Li
N: SIRT3 protects cells from hypoxia via PGC-1α- and
MnSOD-dependent pathways. Neuroscience. 286:109–121. 2015.
View Article : Google Scholar
|
|
15
|
Rangarajan P, Karthikeyan A, Lu J, Ling EA
and Dheen ST: Sirtuin 3 regulates Foxo3a-mediated antioxidant
pathway in microglia. Neuroscience. 311:398–414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong H, Ning Z, Yu L, Li L, Lin L and
Huang J: Preparative separation and identification of the flavonoid
phlorhizin from the crude extract of Lithocarpus polystachyus Rehd.
Molecules. 12:552–562. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen TY, Chi KH, Wang JS, Chien CL and Lin
WW: Reactive oxygen species are involved in FasL-induced
caspase-independent cell death and inflammatory responses. Free
Radic Biol Med. 46:643–655. 2009. View Article : Google Scholar
|
|
18
|
Ishii T, Takanashi Y, Sugita K, Miyazawa
M, Yanagihara R, Yasuda K, Onouchi H, Kawabe N, Nakata M, Yamamoto
Y, et al: Endogenous reactive oxygen species cause astrocyte
defects and neuronal dysfunctions in the hippocampus: A new model
for aging brain. Aging Cell. 16:39–51. 2017. View Article : Google Scholar :
|
|
19
|
Gao JM, Li R, Zhang L, Jia LL, Ying XX,
Dou DQ, Li JC and Li HB: Cuscuta chinensis seeds water extraction
protecting murine osteoblastic MC3T3-E1 cells against tertiary
butyl hydroperoxide induced injury. J Ethnopharmacol. 148:587–595.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang YY, Sun XT, Li ZX, Chen WY, Wang X,
Liang ML, Shi H, Yang ZS and Zeng WT: Protective effect of
angiotensin-(1-7) against hyperglycaemia-induced injury in H9c2
cardiomyoblast cells via the PI3KAkt signaling pathway. Int J Mol
Med. 41:1283–1292. 2018.
|
|
21
|
Roelofs BA, Ge SX, Studlack PE and Polster
BM: Low micromolar concentrations of the superoxide probe MitoSOX
uncouple neural mitochondria and inhibit complex IV. Free Radic
Biol Med. 86:250–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee Y, Heo G, Lee KM, Kim AH, Chung KW, Im
E, Chung HY and Lee J: Neuroprotective effects of 2,4-dinitrophenol
in an acute model of Parkinson's disease. Brain Res. 1663:184–193.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hong YA, Lim JH, Kim MY, Kim Y, Park HS,
Kim HW, Choi BS, Chang YS, Kim HW, Kim TY and Park CW:
Extracellular superoxide dismutase attenuates renal oxidative
stress through the activation of adenosine monophosphate-activated
protein kinase in diabetic nephropathy. Antioxid Redox Signal.
28:1543–1561. 2018. View Article : Google Scholar
|
|
24
|
Gao C, Chang P, Yang L, Wang Y, Zhu S,
Shan H, Zhang M and Tao L: Neuroprotective effects of hydrogen
sulfide on sodium azide-induced oxidative stress in PC12 cells. Int
J Mol Med. 41:242–250. 2018.
|
|
25
|
Di Domenico F, Barone E, Perluigi M and
Butterfield DA: The triangle of death in Alzheimer's disease brain:
The aberrant cross-talk among energy metabolism, mammalian target
of rapamycin signaling, and protein homeostasis revealed by redox
proteomics. Antioxid Redox Signal. 26:364–387. 2017. View Article : Google Scholar
|
|
26
|
Wang X, Dong W, Yuan B, Yang Y, Yang D,
Lin X, Chen C and Zhang W: Vitamin E confers cytoprotective effects
on cardiomyocytes under conditions of heat stress by increasing the
expression of metallothionein. Int J Mol Med. 37:1429–1436. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gao J, Deng Y, Yin C, Liu Y, Zhang W, Shi
J and Gong Q: Icariside II, a novel phosphodiesterase 5 inhibitor,
protects against H2O2-induced PC12 cells
death by inhibiting mitochondria-mediated autophagy. J Cell Mol
Med. 21:375–386. 2017. View Article : Google Scholar
|
|
28
|
Zhong J, Xu C, Gabbay-Benziv R, Lin X and
Yang P: Superoxide dismutase 2 overexpression alleviates maternal
diabetes-induced neural tube defects, restores mitochondrial
function and suppresses cellular stress in diabetic embryopathy.
Free Radic Biol Med. 96:234–244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rottenberg H and Hoek JB: The path from
mitochondrial ROS to aging runs through the mitochondrial
permeability transition pore. Aging Cell. 16:943–955. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang JY, Deng YN, Zhang M, Su H and Qu
QM: SIRT3 acts as a neuroprotective agent in rotenone-induced
Parkinson cell model. Neurochem Res. 41:1761–1773. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yin J, Han P, Tang Z, Liu Q and Shi J:
Sirtuin 3 mediates neuro-protection of ketones against ischemic
stroke. J Cereb Blood Flow Metab. 35:1783–1789. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou X, Chen M, Zeng X, Yang J, Deng H, Yi
L and Mi MT: Resveratrol regulates mitochondrial reactive oxygen
species homeostasis through Sirt3 signaling pathway in human
vascular endothelial cells. Cell Death Dis. 5:e15762014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guo Q, Li S, Xie Y, Zhang Q, Liu M, Xu Z,
Sun H and Yang Y: The NAD+-dependent deacetylase,
Bifidobacterium longum Sir2 in response to oxidative stress by
deacetylating SigH (sigmaH) and FOXO3a in Bifidobacterium longum
and HEK293T cell respectively. Free Radic Biol Med. 108:929–939.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo Y, Li Z, Shi C, Li J, Yao M and Chen
X: Trichostatin A attenuates oxidative stress-mediated myocardial
injury through the FoxO3a signaling pathway. Int J Mol Med.
40:999–1008. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu L, Gong B, Duan W, Fan C, Zhang J, Li
Z, Xue X, Xu Y, Meng D, Li B, et al: Melatonin ameliorates
myocardial ischemia/reperfusion injury in type 1 diabetic rats by
preserving mitochondrial function: Role of AMPK-PGC-1α-SIRT3
signaling. Sci Rep. 7:413372017. View Article : Google Scholar
|
|
36
|
Dai SH, Chen T, Wang YH, Zhu J, Luo P, Rao
W, Yang YF, Fei Z and Jiang XF: Sirt3 attenuates hydrogen
peroxide-induced oxidative stress through the preservation of
mitochondrial function in HT22 cells. Int J Mol Med. 34:1159–1168.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Karnewar S, Neeli PK, Panuganti D,
Kotagiri S, Mallappa S, Jain N, Jerald MK and Kotamraju S:
Metformin regulates mitochondrial biogenesis and senescence through
AMPK mediated H3K79 methylation: Relevance in age-associated
vascular dysfunction. Biochim Biophys Acta. 1864:1115–1128. 2018.
View Article : Google Scholar
|