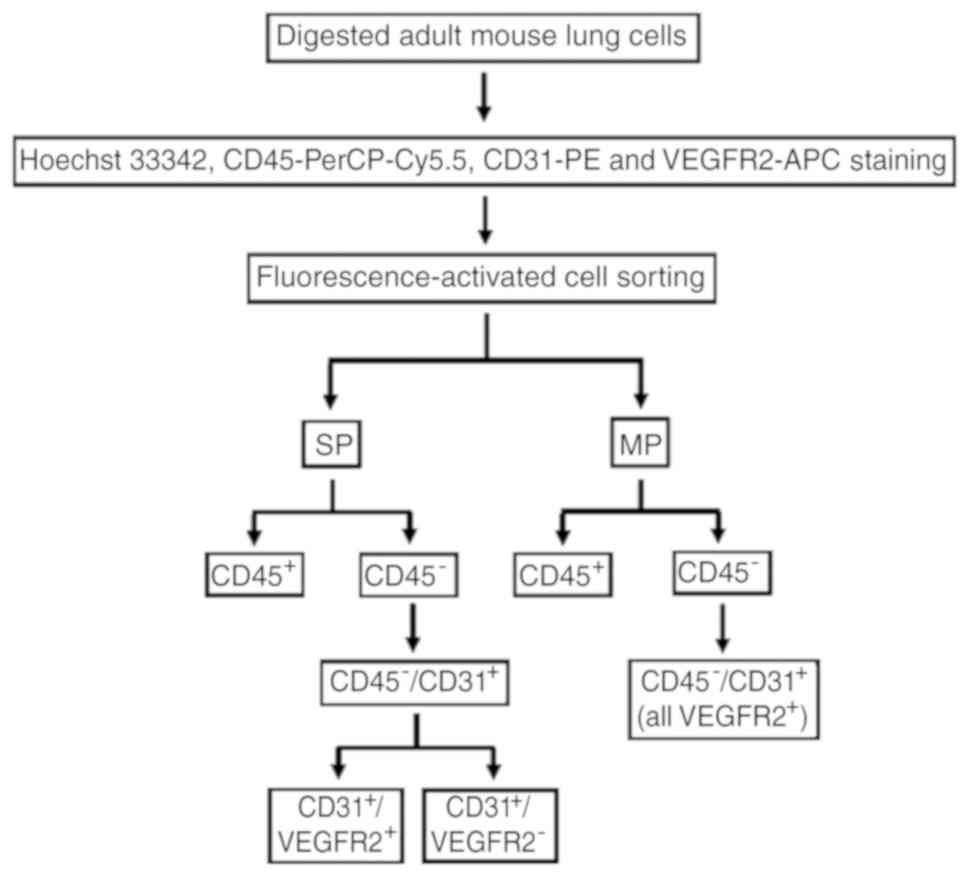
Introduction
Chronic pulmonary diseases, including pulmonary
hypertension, chronic obstructive pulmonary disease and
interstitial pulmonary fibrosis, constitute major contributors to
morbidity and mortality rates (1,2).
The use of stem cells for cell-based therapy in chronic pulmonary
diseases has been under investigation (3–8).
In previous years, different populations of lung stem- or
progenitor-cells have been reported to reside within the adult
mouse lung. They are thought to be involved in tissue maintenance
and repair, however, they may also contribute to pulmonary diseases
(4,5,8).
Using different markers and methods, these cells have been
variously described as: i) Airway clara cells (9), ii) alveolar type II cells (10), iii) lung c-kit positive cells
(11,12), and iv) lung side population (LSP)
cells (13–16). SP cells are identified by their
capacity to efflux Hoechst 33342 dye, allowing the weakly-labeled
SP cells to be distinguished from the intensely-labeled main
population (MP) of cells (17–19). This unique property is mediated
through the action of the p-glycoprotein multidrug/ATP-binding
cassette transporter protein, ATP-binding cassette super-family G
member 2 (ABCG2) on the surface of SP cells, which pumps Hoechst
dye out of the cell (20,21). SP cells have been found in
numerous adult mammalian tissues and organs, including bone marrow,
the heart, liver, skeletal muscle, brain, kidney and lung (14,19,21–23). These cells are rare and they are
enriched for stem/progenitor cell activity.
Several studies have suggested that LSP cells are
capable of differentiation into epithelial cells, endothelial
cells, smooth muscle cells and mature hematopoietic cells (13–16,24). LSP cells are known to be
heterogeneous (13,14,24,25). Based on their expression of CD45,
LSP cells can be separated into CD45+ and
CD45− subpopulations. CD45+ LSP cells possess
hematopoietic progenitor and stem cell activity, and are most
likely bone marrow-derived (14,24). By contrast, CD45− LSP
cells can be further divided into CD45−/CD31−
and CD45−/CD31+ subpopulations (15,24). The
CD45−/CD31− subpopulation has been shown to
comprise resident mesenchymal progenitor cells, capable of
differentiating in vitro into cartilage, bone and adipose
cells (15). However, the nature
and differentiation potential of CD45−/CD31+
LSP cells remain to be fully elucidated. Our previous study
demonstrated that CD45− LSP cells may be progenitors of
endothelial and smooth muscle cells in the embryonic mouse lung
(14,24). In addition, it has been suggested
that mouse heart and lung tissues, including blood vessel cells,
may share a novel population of progenitor cells (26). In the adult mouse heart, the stem
cell antigen-1 (SCA1)+/CD31+ subpopulation of
cardiac SP (CSP) cells have been reported to be endothelial
progenitors (27,28). Therefore, it was hypothesized that
CD45−/CD31+ LSP cells may serve as
progenitors of endothelial and smooth muscle cells in the adult
mouse lung. Previous studies have indicated that
CD45−/VEGFR2+ LSP cells may be endothelial
progenitor cells in the adult mouse lung. The association between
CD45−/CD31+ and
CD45−/VEGFR2+ LSP cells remains to be
elucidated. The present study aimed to characterize
CD45−/CD31+ LSP cells in the adult mouse
lung, to clarify their differentiation potential, and to examine
the association between these two types of LSP cells in
vitro.
Materials and methods
Animals
Female wild-type C57BL/6J mice (aged 8–10 weeks,
weighting 22–25 g) were purchased from Beijing HFK-Biotechnology
(Beijing, China). The mice were group-housed in
individually-ventilated cages under pathogen-free conditions. The
mice were maintained under a 12 h light/dark photoperiod, at 20±1°C
with 60±10% relative humidity and food and water were provided
ad libitum. The general health status of the animals was
monitored daily. Animal care, handling and all animal experiments
described herein were conducted strictly according to ethical
standards approved by the Animal Ethical Committee of Jinzhou
Medical University (Jinzhou, China; approval ID: LY2014D001).
CD45−/CD31+ LSP
cell isolation, fluorescence-activated cell sorting (FACS) and
analysis
The LSP cells were isolated as previously described
(14,15,28,29). Briefly, the mice were sacrificed
by isoflurane anesthesia followed by cervical dislocation. Prior to
lung extraction, animals were bled by transecting the abdominal
aorta. Subsequently, in an attempt to remove circulating blood
cells, perfusion of the pulmonary vasculature was performed using
ice-cold saline until the lungs were bleached white. The lung
extracts were then digested by finely mincing tissue with a razor
blade in the presence of 0.1% collagenase (Roche Diagnostics,
Indianapolis, IN, USA), 2.4 U/ml dispase (Roche Diagnostics), and
2.5 mM CaCl2 at 37°C for 1 h. Removal of nonspecific
debris was accomplished by sequential filtration through 70– and
40–µm filters, and the cells were resuspended at a density
of 1×106 cells/ml. Cell staining was performed by
incubating with Hoechst 33342 (5 µg/ml; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at 37°C for 90 min in the presence or
absence of verapamil (50 mM; Sigma-Aldrich; Merck KGaA) (21). The suspension was subsequently
incubated with one or more fluorescently-conjugated monoclonal rat
anti-mouse antibodies, including anti-CD45-PerCP-Cy5.5 (1:100; cat.
no. 550994; BD Biosciences, Franklin Lakes, NJ, USA), anti-CD31-PE
(1:100; cat. no. 553373; BD Biosciences), anti-VEGFR2-APC (1:100;
cat. no. 561993; BD Biosciences), anti-SCA1-FITC (1:100; cat. no.
557405; BD Biosciences) and anti-c-kit-FITC (1:100; cat. no.
561680; BD Biosciences). Isotype control antibodies were used in
parallel as negative controls and to establish gating parameters
for positively stained cells. All incubations were performed for 30
min at 4°C in the dark.
FACS was conducted using the MoFlo™ XDP flow
cytometer (Beckman Coulter, Inc., Brea, CA, USA) to isolate
CD45−/CD31+ LSP cells,
CD45−/CD31+/VEGFR2− LSP cells,
CD45−/CD31+/VEGFR2+ LSP cells,
CD45− lung MP cells and
CD45−/CD31+ LMP cells for all experiments.
The following six parameters were used to discriminate cells in the
experiments: Hoechst 33342 (red), Hoechst 33342 (blue), forward
scatter, side scatter, PerCP-Cy5.5-CD45, FITC-SCA1 and PE-CD31
reactivity (green) as previously described (14,27,30). This protocol is illustrated in
Fig. 1. The Hoechst dye was
excited at 350 nm. A 488-nm argon laser was employed for exciting
PerCP-Cy5.5, PE and FITC, and a 633-nm HeNe laser was used for APC.
Data analysis was performed using Summit™ V5.2 software (Beckman
Coulter, Inc.).
Primary cell cultures
The murine
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells were
cultured on fibronectin-coated 8-well chamber slides (10,000
cells/well; Nunc; Sigma-Aldrich; Merck KGaA) at 37°C under 5%
CO2 as previously described (24,28). To promote endothelial cell growth
and differentiation, the murine
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells were
cultured at 37°C with a defined endothelial differentiation medium
(EGM-2) from Cambrex (Baltimore, MA, USA) supplemented with 100
U/ml of penicillin (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and 100 µg/ml of streptomycin (Invitrogen;
Thermo Fisher Scientitic, Inc.). Following 14 days in culture, the
cells were collected for reverse-transcription quantitative
polymerase chain reaction (RT-qPCR) and immunostaining analyses,
respectively. To examine the association between
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells, the
CD45−/CD31+/VEGFR2− LSP cells were
cultured in the same conditions mentioned above for 2 days and then
collected for RT-qPCR analysis.
To promote smooth muscle growth and differentiation,
the CD45−/CD31+/VEGFR2− LSP cells
were cultured with a defined smooth muscle differentiation medium
(SmGm-2) from Cambrex supplemented with 100 U/ml of penicillin
(Invitrogen; Thermo Fisher Scientific, Inc.), and 100 µg/ml
of streptomycin (Invitrogen; Thermo Fisher Scientific, Inc.). The
culture medium was replaced every 2 days. Following 14 days in
culture, the cells were collected for RT-qPCR and immunostaining
analyses. To determine whether the
CD45−/CD31+/VEGFR2+ LSP cells were
able to differentiate into smooth muscle cells, they were cultured
in the medium described above for 14 days and were then harvested
for RT-qPCR analysis.
Cell line culture
The HepG2 human liver cancer cell line, obtained
from American Type Culture Collection (Manassas, VA, USA), was
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% GIBCO® fetal calf serum, 100 U/ml of
penicillin, and 100 µg/ml of streptomycin (all from
Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C in a 5%
CO2 incubator.
Cellular uptake of Dil-labeled acetylated
low-density lipoprotein (Dil-Ac-LDL)
The CD45−/CD31+ LSP cells at a
density of 7,000 cells/well were incubated with EGM-2 containing 10
mg/ml of Dil-Ac-LDL (Molecular Probes; Thermo Fisher Scientific,
Inc.) at 37°C for 8 h as previously described (28). Following washing with PBS, the
cells were fixed and nuclei were counterstained with
4-6-diamidino-2-phenylindol-dihydrochloride (DAPI). The slides were
examined using an Olympus-DP70 microscope and images were captured
with a digital camera (BX51; Olympus Corporation, Tokyo, Japan).
HepG2 cells were included to provide a negative control for Ac-LDL
uptake.
Methylcellulose assay (colony-forming
unit assay)
The CD45−/CD31+ LSP and LMP
cells (7,000 cells/ml) were cultured in Methocult GF M3534 medium
according to the manufacturer's protocol (StemCell Technologies,
Inc., Vancouver, Canada) as described previously (17,19,27,28). Cell colonies, consisting of >30
cells, were scored after 14 days in culture. The cell colonies were
fixed with 4% paraformaldehyde and stained with crystal violet
based on the manufacturer's protocol (Sigma-Aldrich; Merck KGaA).
Photomicrographs were obtained, from which the colonies were
counted using an Olympus-DP70 microscope and images were captured
with a digital camera (BX51). In order to determine whether the
cells retained their CD45−/CD31+ LSP
phenotype, Methoult medium was cut into sections and these were
incubated with DMEM at 37°C with agitation for 30 min. When the
Methocult medium had dissolved in the DMEM, the cells were
collected, labeled with the CD45-PerCP-Cy5.5 (1:100), CD31-PE
(1:100) and SCA1-FITC (1:100) antibodies, and analyzed by FACS.
RT-qPCR analysis
Total RNA was extracted from the freshly isolated
distinct LSP, LMP and cultured cells using an RNAeasy-Mini kit,
following the manufacturer's protocol (Qiagen GmbH, Hilden,
Germany). Complementary DNA was generated using an M-MLV MicroRNA
Reverse Transcription kit (Promega Corporation, Madison, WI, USA).
RT-qPCR analysis was performed as previously described using a
Rotor-gene 3000 machine (Corbett; Qiagen GmbH) with a SYBR-Green
qPCR-SuperMix-UDG kit (Thermo Fisher Scientific, Inc.) following
the manufacturer's protocol. Commercial primer sets were obtained
from Qiagen GmbH, as follows: ATP-binding cassette super-family G
member 2 (ABCG2; cat. no. QT00173138), CD133 (cat. no. QT01065162),
VEGFR2 (cat. no. QT00097020), von Willebrand factor (vWF; cat. no.
QT00116795), α-smooth muscle tropomyosin (αSMT; cat. no.
QT00137354), α-smooth muscle actin (αSMA; cat. no. QT0006746), CD44
(cat. no. QT00173404), myosin heavy chain (cat. no. QT01558130) and
β-actin (cat. no. QT01136772). qPCR was performed in a 20-µl
reaction system, which contained 10 µl SYBR-Green
qPCR-SuperMix-UDG (Thermo Fisher Scientific, Inc.), 2 µl
cDNA, 0.4 µl each primer and 7.2 µl RNase-free
H2O. The thermal cycler conditions were performed as
follows: Cycle 1 (95°C for 3 min), Cycle 2 (35 cycles of 95°C for
30 sec, 57°C for 30 sec, 72°C for 30 sec), Cycle 3 (72°C for 4
min), which was based on the manufacturer's protocol (31). All samples were amplified in
triplicate. Amplification data were analyzed with Corbett
Rotor-gene 3000 software (Qiagen GmbH) as described previously
(27,28). The relative mRNA expression levels
of the genes mentioned above were normalized against the expression
of housekeeper gene β-actin and calculated using the
2−ΔΔCq method as previously described (28,31,32). Gene expression data are presented
in relative units.
Immunofluorescence staining of isolated
and cultured cells
For analysis of the sorted cells, 1,000
CD45−/CD31+/VEGFR2− LSP cells,
CD45−/CD31+/VEGFR2+ LSP cells and
CD45− LMP cells were collected onto positively charged
slides by cyto-centrifugation at 60 × g for 5 min at room
temperature prior to air-drying. The slides were then fixed for 10
min in 4% paraformaldehyde/PBS, washed with 1X PBS, air dried and
stored at 4°C prior to staining. The cultured monolayer cells were
fixed and prepared for staining using the same procedure.
Immunofluorescence staining was performed as previously described
(14). Autofluorescence was first
quenched with 10 mM sodium borohydride for 30 min at room
temperature. Nonspecific binding was blocked by incubating with 1%
rabbit serum (cat. no. ab7356; Abcam, Cambridge, MA, USA) or goat
serum (cat. no. ab7487; Abcam). The primary antibodies used
included monoclonal rat anti-mouse ABCG2 (1:100; cat. no. ab24115;
Abcam), polyclonal rabbit anti-mouse vWF (1:100; cat. no. AB7356;
Chemicon, Temecula, CA, USA), monoclonal anti-mouse αSMA-Cy3
(1:100; cat. no. C-6198; Sigma-Aldrich; Merck KGaA) and polyclonal
rabbit anti-mouse αSMT (1:100; cat. no. T3651; Sigma-Aldrich; Merck
KGaA). Secondary antibodies included rabbit anti-rat IgG-FITC
(1:200; cat. no. ab6730-1; Abcam), goat anti-rabbit IgG-594 (1:200;
cat. no. A11012; Invitrogen; Thermo Fisher Scientific, Inc.) and
donkey anti-rabbit IgG-FITC (1:200; cat. no. A21206; Invitrogen;
Thermo Fisher Scientific, Inc.). All incubations of primary and
secondary antibodies were performed for 60 and 30 min at 4°C in the
dark, respectively. The nuclei were counterstained with DAPI.
Slides were examined using an Olympus-DP70 microscope and images
were captured with a digital camera (BX51).
Statistical analysis
Data are presented as the mean ± standard deviation.
Student's t-test (two-tailed unpaired) was used to analyze
experimental data containing two groups, and one-way analysis of
variance with the Bonferroni post hoc test was used to analyze
experimental data with ≥3 groups. Statistical analysis was
performed using SPSS version 16.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference. Each experiment was repeated three times to obtain the
means used for statistical analysis (n=3 experiments) as previously
described (28).
Results
LSP cell populations Identification,
isolation and gene expression
The adult mouse lungs were enzymatically digested
into single cell suspensions and subjected to FACS analysis
(Fig. 1). The LSP cells were
detected on density dot plots based on their capacity to efflux
Hoechst 33342 dye. Those cells that were weakly labeled for Hoechst
33342 dye (LSP cells) represented ~0.7% of the total cell
population (Fig. 2A). The LSP
population disappeared when the ABCG2 transporter was blocked with
verapamil (Fig. 2B). The LSP
cells were first separated into CD45+ (~54% of total LSP
cells) and CD45− (~46% of total LSP cells)
subpopulations as previously described (14) (Fig.
2C). Based on the expression of CD31, the CD45− LSP
cells were further divided into CD45−/CD31−
LSP cells (~39% of total CD45− LSP cells) and
CD45−/CD31+ LSP cells (~61%). According to
expression of VEGFR2, CD45−/CD31+ LSP cells
were further separated into
CD45−/CD31+/VEGFR2− (~29% of total
CD45− LSP cells) and
CD45−/CD31+/VEGFR2+ LSP cells
(~32%) (Fig. 2D). All
CD45−/CD31+ LSP cells (VEGFR2− and
VEGFR2+) expressed SCA1, but none expressed c-kit
(Fig. 2E and F). Using the same
surface markers, the LMP cells were separated into CD45+
and CD45− subpopulations, which were further divided
into CD45−/CD31−/VEGFR2− and
CD45−/CD31+/VEGFR2+ LMP cells.
Notably, all CD45−/CD31+ LMP cells were
VEGFR2+ (Fig. 2G and
H). Based on their immune-phenotype,
CD45−/CD31+ LSP,
CD45−/CD31+/VEGFR2− LSP,
CD45−/CD31+/VEGFR2+ LSP,
CD45− LMP and CD45−/CD31+ LMP
cells were isolated by FACS (Fig.
2).
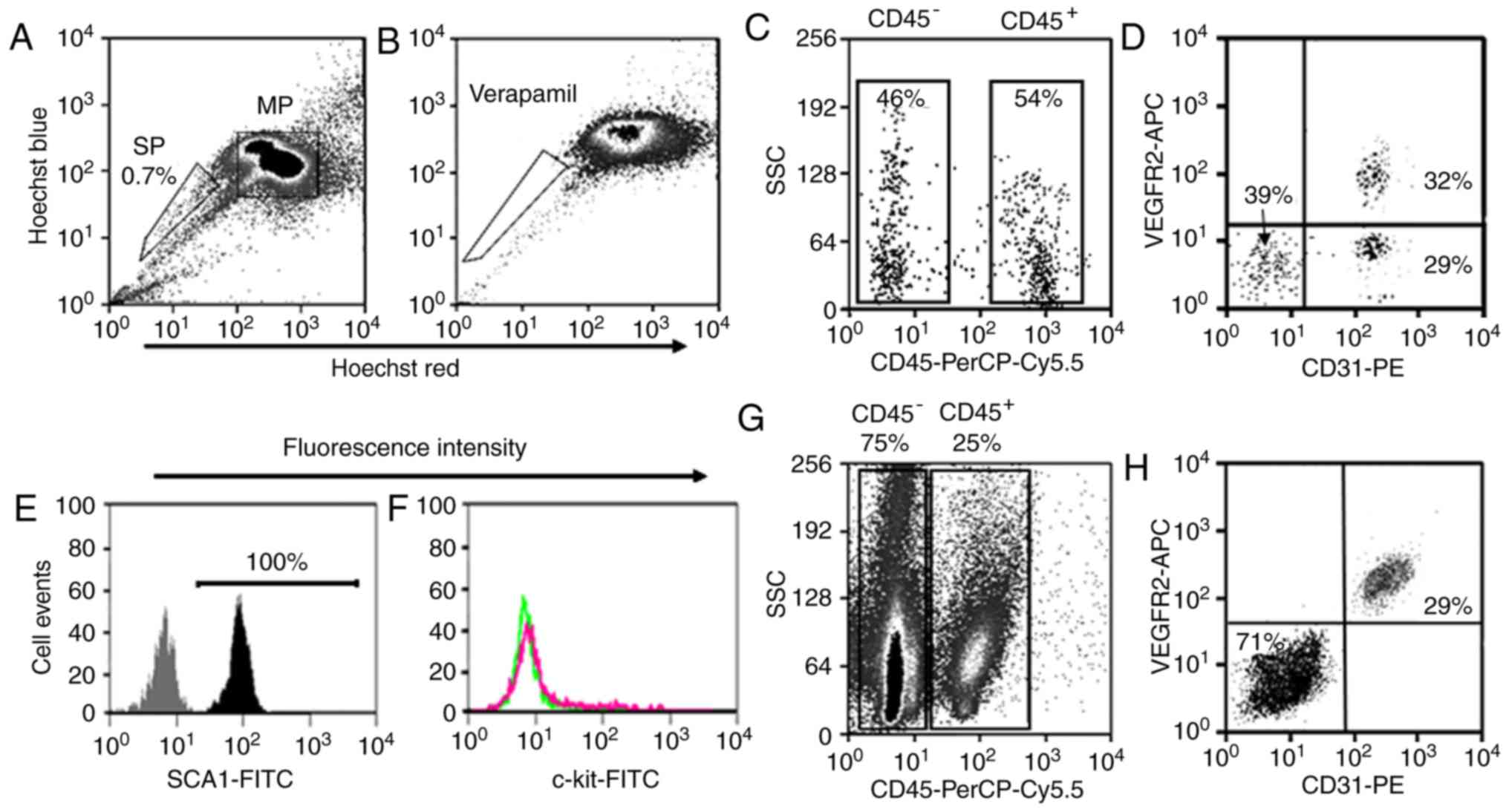 | Figure 2Analysis and isolation of LSP and LMP
cells from the adult mouse lung by fluorescence-activated cell
sorting. (A) Lung SP cells (outlined to lower left) representing
~0.7% of the total heart cell population and MP cells (outlined to
right) were identified based upon their intensity of Hoechst
staining. (B) SP cells disappeared when cells were labeled in the
presence of verapamil pretreatment, which inhibits the transporter
(outlined to lower left). (C) Based on the expression of CD45, LSP
cells were divided into CD45+ and CD45−
representing ~54 and 46% of the total LMP cells, respectively. (D)
According to expression of CD31 and VEGFR2, CD45− LSP
cells were further separated into
CD45−/CD31+/VEGFR2−,
CD45−/CD31+/VEGFR2+ and
CD45−/CD31−/VEGFR2−, representing
~29, 32 and 39% of the total LSP cells, respectively.
CD45−/CD31+ LSP cells (including
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells)
were stained with FITC-conjugated (E) SCA1 and (F) c-kit antibody
(black and red respectively) or its isotype control (gray and green
respectively), showing 100% positive for SCA1 and negative for
c-kit. (G) LMP cells were separated into CD45+ (25% of
total LMP) and CD45− (75% of total LMP) cells and (H)
further divided into
CD45−/CD31+/VEGFR2+ and
CD45−/CD31−/VEGFR2− LMP cells. SP,
side population; MP, main population; LSP, lung SP; LMP, lung MP;
VEGFR2, vascular endothelial growth factor receptor 2; SCA1, stem
cell antigen-1. |
In our previous study, it was found that
CD31+ CSP cells are endothelial progenitors of adult
mouse heart (27). Therefore, the
present study aimed to further characterize
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells by
examining the expression of genes specific for stem/progenitor
cells and endothelial cells, using RT-qPCR analysis. As expected,
the CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells
expressed high mRNA levels of the ABCG2 transporter, which is
responsible for the efflux of Hoechst 33342 dye (20). Both also expressed CD133, a marker
for endothelial progenitor cells (33). By contrast, the CD45−
LMP cells expressed neither ABCG2 nor CD133. VEGFR2, a marker of
endothelial cells, and vWF, a mature endothelial cell marker, were
just detectable in the
CD45−/CD31+/VEGFR2+ LSP cells but
not in the CD45−/CD31+/VEGFR2− LSP
cells (Fig. 3A). CD45−
LMP cells, serving as a positive control, expressed high levels of
VEGFR2 and vWF (Fig. 3A). The
CD45− LMP cells also expressed high levels of mRNAs
encoding αSMA, a marker of smooth muscle progenitors (24,34), αSMT, a marker of mature smooth
muscle cells (24,34) and CD44, a marker of mesenchymal
cells. By contrast, the expression of αSMA was low (but detectable)
in the CD45−/CD31+/VEGFR2− LSP
cells and completely undetectable in the
CD45−/CD31+/VEGFR2+ LSP cells,
indicating that
CD45−/CD31+/VEGFR2− LSP cells may
be smooth muscle progenitors, whereas
CD45−/CD31+/VEGFR2+ LSP cells are
not. In the CD45−/CD31+/VEGFR2−
and the CD45−/CD31+/VEGFR2+ LSP
cells, no expression of αSMT and CD44 was detectable (Fig. 3A), suggesting that neither the
CD45−/CD31+/VEGFR2− LSP cells nor
the CD45−/CD31+/VEGFR2+ LSP cells
are mature smooth muscle or mesenchymal cells.
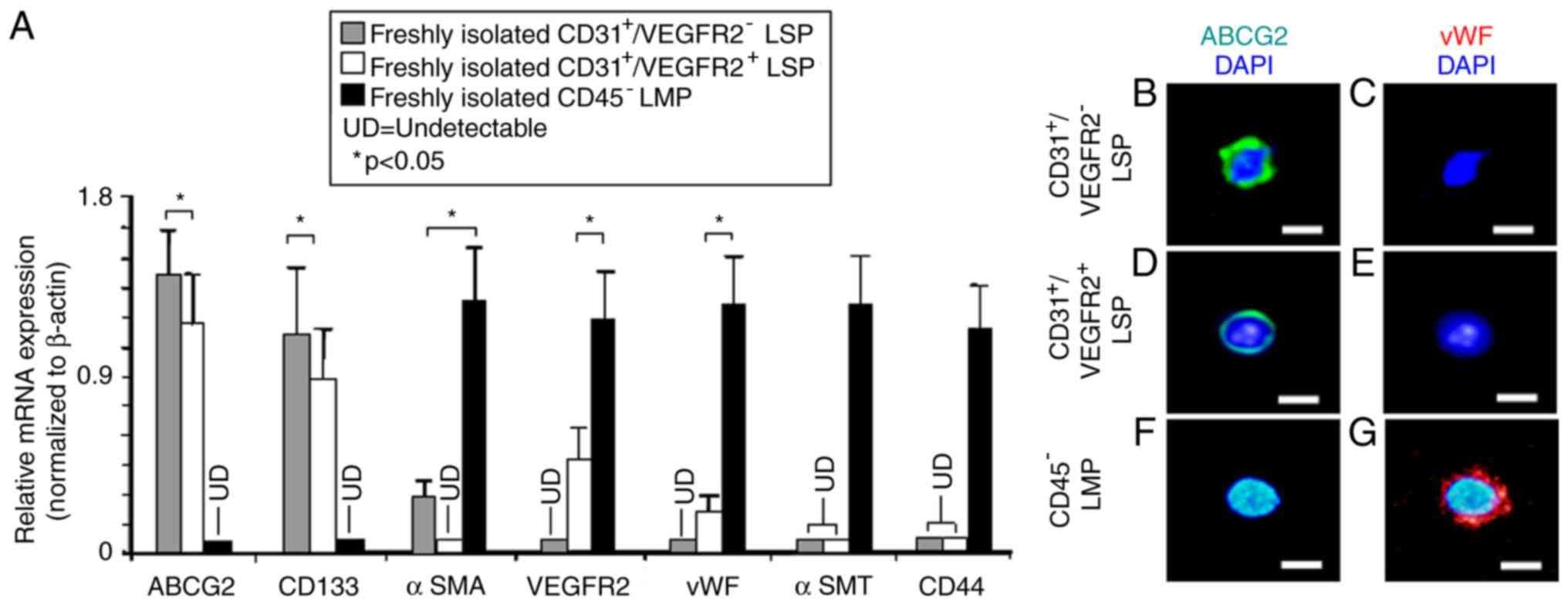 | Figure 3Gene expression in sorted LSP and LMP
cells. Expression of markers for stem cells (ABCG2), endothelial
progenitors (CD133), early smooth muscle (αSMA), endothelial cells
(VEGFR2), mature endothelial cells (vWF), mature smooth muscle
cells (αSMT) and mesenchymal cells (CD44) were compared among
sorted populations of cells. (A) Reverse transcription-quantitative
polymerase chain reaction analysis compared relative mRNA
expression levels in
CD45−/CD31+/VEGFR2− (grey bar) and
CD45−/CD31+/VEGFR2+ LSP (white
bar) cells. CD45− LMP cells (black bar) were used as a
control. For immunofluorescent labeling of cells, following
sorting, CD45−/CD31+/VEGFR2+ and
CD45−/CD31+/VEGFR2− LSP cells and
CD45− LMP cells were cytospun onto glass slides. Nuclei
(blue) were counterstained with DAPI.
CD45−/CD31+/VEGFR2− LSP cells were
immunopositive for (B) ABCG2 (green), but not (C) vWF (red).
Similarly, CD45−/CD31+/VEGFR2+ LSP
cells were positive for (D) ABCG2 (green), but not for (E) vWF
(red). By contrast, CD45− LMP cells were not
immunopositive for (F) ABCG2 (green) but were for (G) vWF (red).
Scale bar, 30 µm. Data are presented as the mean ± standard
deviation (n=3 experiments); *P<0.05; UD,
undetectable; LSP, lung side population; LMP, lung main population;
ABCG2, ATP-binding cassette super-family G member 2; VEGFR2,
vascular endothelial growth factor receptor 2; vWF, von Willebrand
factor; αSMT, α-smooth muscle tropomyosin; αSMA, α-smooth muscle
actin; DAPI, 4-6-diamidino-2-phenylindol-dihydrochloride. |
Subsequently, the
CD45−/CD31+/VEGFR2− LSP,
CD45−/CD31+/VEGFR2+ LSP and
CD45− LMP cells were collected onto duplicate glass
slides (1,000 cells/slide) and antibody probes were used to examine
the expression of proteins. The immunostaining confirmed that the
CD45−/CD31+/VEGFR2− LSP and
CD45−/CD31+/VEGFR2+ LSP cells
expressed the ABCG2 transporter protein, but not the vWF protein
(Fig. 3B-E). By contrast, the
CD45− LMP cells expressed vWF protein, but not the ABCG2
transporter (Fig. 3F and G).
Based on the mRNA analysis, the high expression of CD133, low
expression of αSMA, and absence of VEGFR2, vWF, αSMT and CD44,
suggested that CD45−/CD31+/VEGFR2−
LSP cells may be progenitors for endothelial and smooth muscle
cells, but not progenitors for mesenchymal cells. By contrast, the
high expression of CD133, low (detectable) expression of VEGFR2 and
vWF, and absence of αSMA, αSMT and CD44 indicate that
CD45−/CD31+/VEGFR2+ LSP cells may
be late-commitment endothelial progenitors. Taken together, these
results indicate that
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells may
be endothelial progenitor cells. The finding that
CD45−/CD31+ LSP cells, including
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells,
take up DiI-Ac-LDL (Fig. 4A-D)
supports this notion.
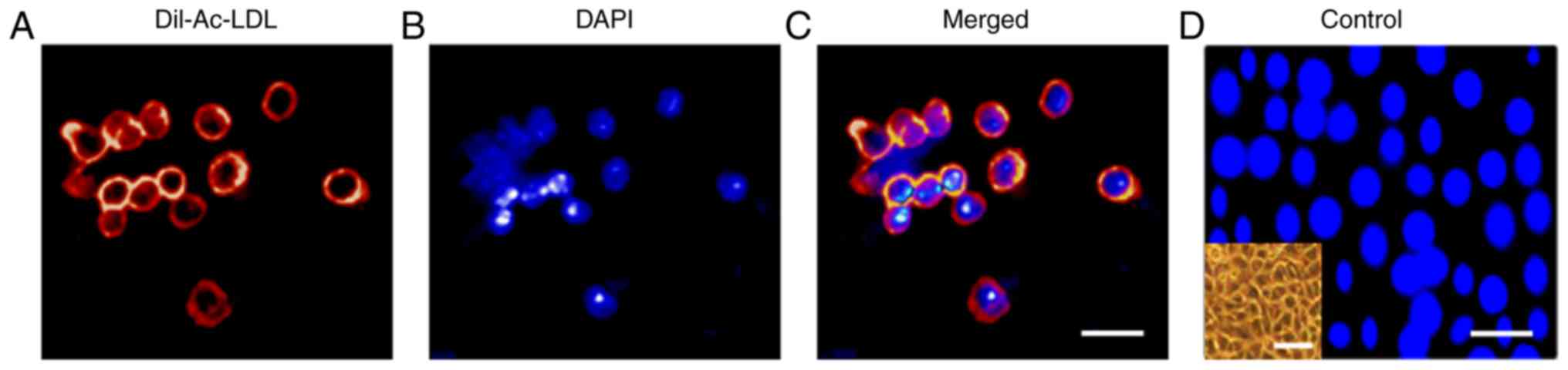 | Figure 4Representative photomicrograph field
showing Dil-Ac-LDL uptake by CD45−/CD31+ LSP
cells (including
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells).
(A) CD45−/CD31+ LSP cells demonstrating
cellular uptake of Dil-Ac-LDL. (B) DAPI fluorescence channel,
showing nuclei. (C) Merged fluorescence channels. Scale bar, 30
µm. (D) HepG2 cells, which did not take up Dil-Ac-LDL,
served as a negative control. Scale bar, 30 µm. The inset on
the lower left shows a phase contrast image of the cultured HepG2
cells (scale bar, 60 µm). VEGFR2, vascular endothelial
growth factor receptor 2; LSP, lung side population; Dil-Ac-LDL,
Dil-labeled acetylated low-density lipoprotein; DAPI,
4-6-diamidino-2-phenylindol-dihydrochloride. |
In vitro colony-formation by
CD45−/CD31+ LSP cells
Our previous study showed that CD31+ CSP
cells were able to form colonies in vitro (27). Therefore, the present study
examined the in vitro colony-forming potential of
CD45−/CD31+ LSP cells. LMP and LSP cells
freshly isolated from adult murine lungs were plated on
methylcellulose media. After 14 days in culture, the number of
colonies was counted. A typical colony formed by
CD45−/CD31+ LSP cells and a typical field of
CD45−/CD31+ LMP cells are shown in Fig. 5A and B, respectively. Compared
with the CD45−/CD31+ LMP cells, the
CD45−/CD31+ LSP cells produced more colonies
(Fig. 5C-E). FACS analysis of the
LSP cells that were subsequently isolated from the methylcellulose
media revealed surface expression of CD31 (100%) and SCA1 (100%),
but not CD45, indicating that the colony forming cells had retained
their phenotype following culture (Fig. 5F-H). These findings suggest that
CD45−/CD31+ LSP cells possess a substantially
greater potential for self-renewal in culture compared with LMP
cells.
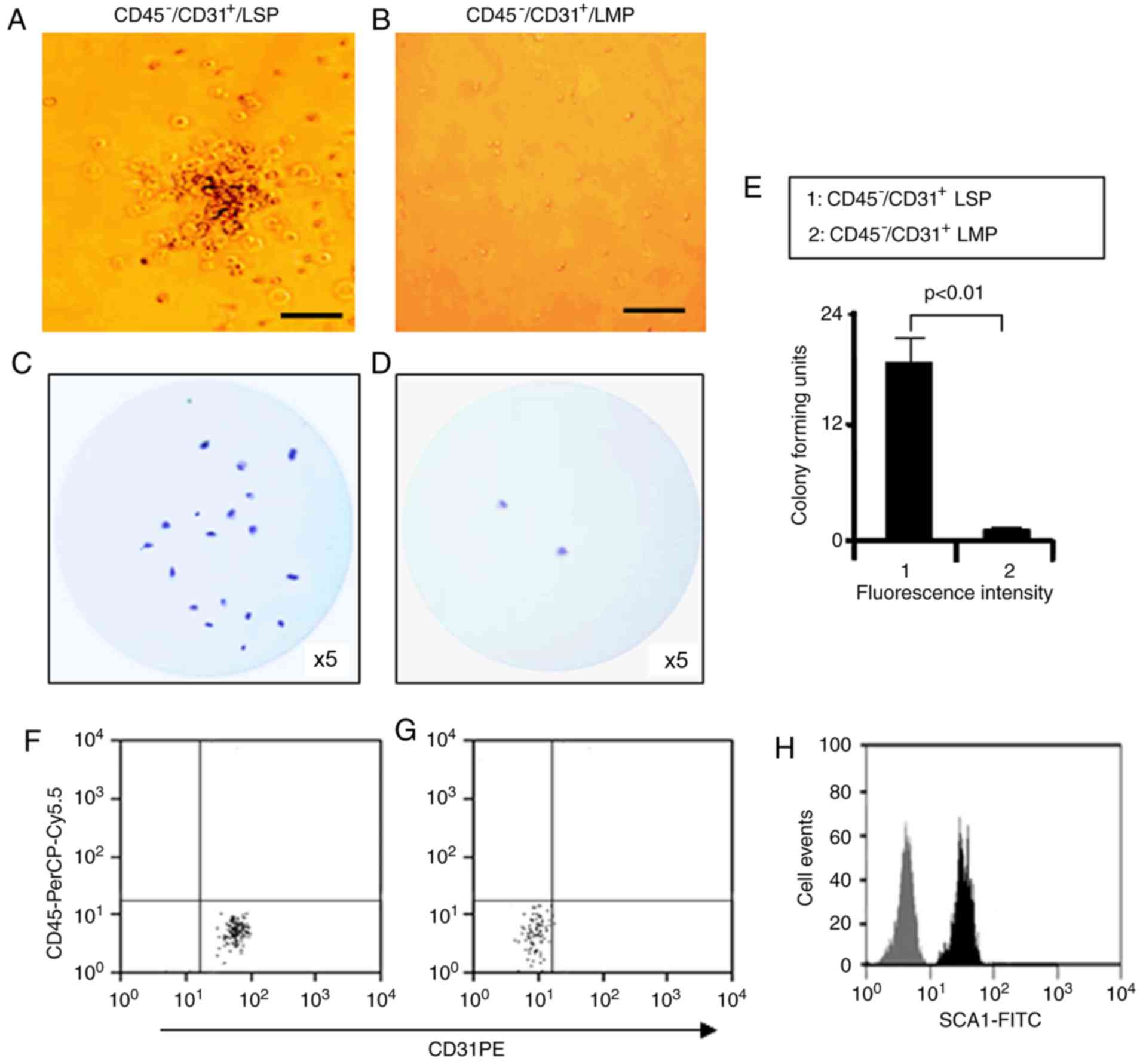 | Figure 5Colony formation by
CD45−/CD31+ LSP cells. (A) Representative
colony formed by CD45−/CD31+ LSP cells in
methylcellulose medium, visualized by phase contrast microscopy
(scale bar, 50 µm). (B) Representative image of a field of
CD45−/CD31+ LMP cells (scale bar, 50
µm). Representative, low magnification images of colony
formation by (C) CD45−/CD31+ LSP cells and
(D) LMP cells cultured in methylcellulose medium and stained with
crystal violet (×5 magnification). (E) Graph showing the ratio of
colonies formed by CD45−/CD31+ LSP cells,
compared to CD45−/CD31+ LMP cells
(P<0.01). (F) Colony-forming cells subsequently isolated from
methylcellulose media, labeled with CD45-PerCP-Cy5.5 and CD31-PE,
and analyzed by flow cytometry, with the results for the (G)
isotype control. (H) Colony-forming cells isolated from methocult
media labeled with SCA1-FITC (black) or its isotype antibody (grey)
and analyzed by flow cytometry. Data are presented as the mean ±
standard deviation (n=3 experiments). LSP, lung side population;
LMP, lung main population. |
In vitro differentiation of LSP
cells
To examine their differentiation potential,
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells were
separately isolated and cultured in conditions known to promote the
growth and differentiation of either endothelial or smooth muscle
cells. Following culture to promote endothelial differentiation for
14 days, the CD45−/CD31+/VEGFR2−
and CD45−/CD31+/VEGFR2+ LSP cells
each formed tube-like vascular networks (Fig. 6A and B), suggesting that they had
differentiated into endothelial, or endothelial-like, cells.
RT-qPCR analysis was then used to examine the expression of
stem/progenitor cell genes vs. endothelial cell-associated genes in
freshly-isolated and cultured
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells.
These were compared with freshly isolated
CD45−/CD31+ LMP cells, which represent mature
endothelial cells. ABCG2 and CD133 were expressed in the freshly
isolated CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells, but
not in the cultured cells nor in the freshly isolated
CD45−/CD31+ LMP cells (Fig. 6C). Expression levels of the
stem/progenitor markers ABCG2 and CD133 were significantly lower in
the freshly isolated
CD45−/CD31+/VEGFR2+ LSP cells
compared with those in the freshly isolated
CD45−/CD31+/VEGFR2− LSP cells. No
expression of endothelial cell markers VEGFR2 and vWF was detected
in the freshly isolated
CD45−/CD31+/VEGFR2− LSP cells, and
their expression was weak in the
CD45−/CD31+/VEGFR2+ LSP cells.
However, following culture, the
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells the
mRNA expression levels of VEGFR2 and vWF were high, at levels
indistinguishable from those in the freshly isolated
CD45−/CD31+ LMP cells (Fig. 6C). These results further support
the hypothesis that
CD45−/CD31+/VEGFR2− LSP cells and
CD45−/CD31+/VEGFR2+ LSP cells may
be endothelial progenitors, similar to our previous findings with
SP cells in the mouse heart (27).
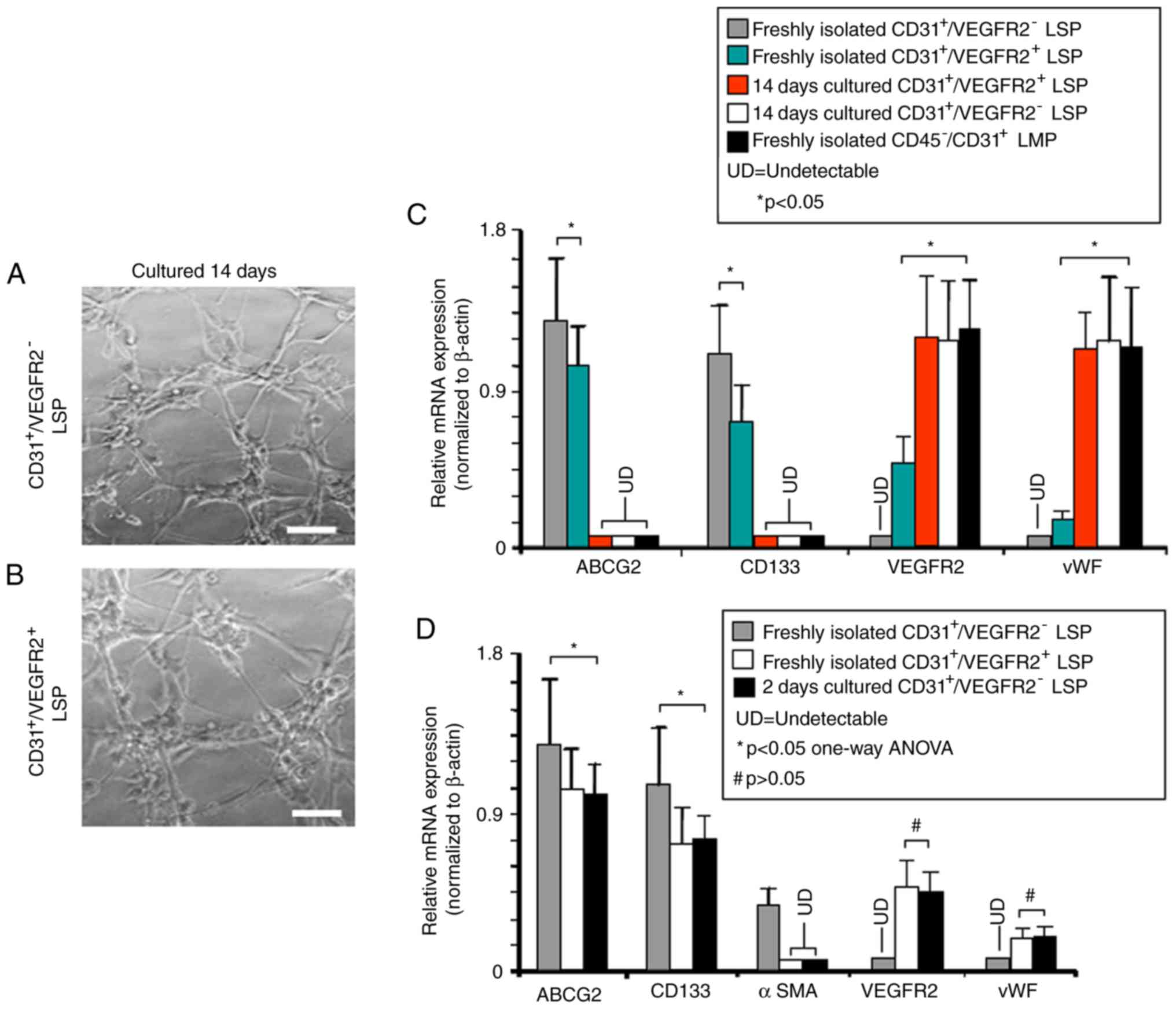 | Figure 6In vitro endothelial
differentiation by
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells.
Representative photomicrographs show vascular tube-like networks
formed by (A) CD45−/CD31+/VEGFR2−
and (B) CD45−/CD31+/VEGFR2+ LSP
cells after 2 weeks in culture under endothelial
differentiation-inducing conditions (scale bar, 50 µm). (C)
RT-qPCR analysis performed to compare relative expression of mRNAs
encoding ABCG2, CD133, VEGFR2 and vWF genes in freshly isolated
CD45−/CD31+/VEGFR2− LSP cells
(grey bars), CD45−/CD31+/VEGFR2+
LSP cells (green bars), in
CD45−/CD31+/VEGFR2+ LSP cells
after 14 days culture (red bar), and in
CD45−/CD31+/VEGFR2− LSP cells
after 14 days culture (white bar). Freshly isolated
CD45−/CD31+ LMP cells are included as a
control (black bar). (D) RT-qPCR analysis was performed to compare
the relative expression of mRNAs encoding ABCG2, CD133, αSMA,
VEGFR2 and vWF genes in freshly isolated
CD45−/CD31+/VEGFR2− LSP (grey
bars) CD45−/CD31+/VEGFR2+ LSP
(white bars), and in
CD45−/CD31+/VEGFR2− LSP after 2
days culture (black bars). Data are presented as the mean ±
standard deviation (n=3 experiments; *P<0.05;
#P>0.05; LSP, lung side population; LMP, lung main
population; UD, undetectable; ABCG2, ATP-binding cassette
super-family G member 2; VEGFR2, vascular endothelial growth factor
receptor 2; vWF, von Willebrand factor; αSMA, α-smooth muscle
actin. |
The detection of the expression of VEGFR2 and vWF,
and relatively low levels of ABCG2, CD133 suggested that
CD45−/CD31+/VEGFR2+ LSP cells may
be late-commitment endothelial progenitors. To investigate whether
CD45−/CD31+/VEGFR2+ LSP cells may
be derived from
CD45−/CD31+/VEGFR2− LSP cells,
freshly isolated
CD45−/CD31+/VEGFR2− LSP cells were
cultured for 2 days under conditions to promote endothelial
differentiation. During this culture, the
CD45−/CD31+/VEGFR2− LSP cells
showed significant reductions in the mRNA expression levels of
ABCG2 and CD133, with levels becoming similar to those found in
freshly isolated
CD45−/CD31+/VEGFR2+ LSP cells
(Fig. 6D). As expected, no
expression of endothelial markers VEGFR2 and vWF was detected in
the freshly isolated
CD45−/CD31+/VEGFR2− LSP cells.
However, following 2 days in culture, mRNA expression of VEGFR2 and
vWF was detected in
CD45−/CD31+/VEGFR2− LSP cells at
levels comparable with those found in freshly isolated
CD45−/CD31+/VEGFR2+ LSP cells
(Fig. 6D). The freshly isolated
CD45−/CD31+/VEGFR2− LSP cells
expressed the smooth muscle marker, αSMA, however, the expression
of αSMA disappeared following 2 days in culture. Therefore, the
CD45−/CD31+/VEGFR2− LSP cells came
to resemble CD45−/CD31+/VEGFR2+
cells in their pattern of gene expression following 2 days in
culture (Fig. 6D). This suggested
that the 2-day-cultured
CD45−/CD31+/VEGFR2− LSP cells and
freshly isolated
CD45−/CD31+/VEGFR2+ LSP cells had
lost their ability to differentiate into smooth muscle cells, which
was consistent with the hypothesis that
CD45−/CD31+/VEGFR2+ LSP cells are
late commitment endothelial progenitor cells. The gene expression
profile of CD45−/CD31+/VEGFR2− LSP
cells became almost identical to that of
CD45−/CD31+/VEGFR2+ LSP cells
following 2 days in culture, suggesting that
CD45−/CD31+/VEGFR2+ LSP cells may
be derived from
CD45−/CD31+/VEGFR2− LSP cells
(Fig. 6D).
The expression of αSMA by freshly isolated
CD45−/CD31+/VEGFR2− LSP cells
suggested that they may serve a role as smooth muscle progenitors.
To investigate this, the
CD45−/CD31+/VEGFR2− LSP cells were
cultured in medium previously shown to promote smooth muscle
differentiation (24). After 14
days in culture, many of the cells had developed the morphologic
characteristics of smooth muscle cells, being large, flat, and
stellate in appearance, and showing uniform immunostaining for αSMA
and αSMT (Fig. 7A-F). To extend
these findings, the gene expression profiles of freshly isolated
and cultured CD45−/CD31+/VEGFR2−
LSP cells were compared. The mRNA expression of ABCG2 and CD13 was
detected in the freshly isolated
CD45−/CD31+/VEGFR2− LSP cells,
however, the expression of these two genes was no longer detectable
following 14 days in smooth muscle medium (Fig. 7G). By contrast, the
CD45−/CD31+/VEGFR2− LSP cells
showed significantly upregulated expression levels of αSMA
following 14 days in smooth muscle media, resulting in levels
similar to those in the freshly isolated CD45− LMP cells
(Fig. 7G). Following 14 days of
culture in smooth muscle differentiation medium, the mature smooth
muscle markers, αSMT and myosin heavy chain (35) were expressed by the
14-day-cultured
CD45−/CD31+/VEGFR2− LSP cells and
freshly isolated CD45− LMP cells, but not by the freshly
isolated CD45−/CD31+/VEGFR2− LSP
cells (Fig. 7G). By contrast, the
CD45−/CD31+/VEGFR2+ LSP cells did
not express detectable mRNA levels of αSMA, αSMT or myosin heavy
chain either before or after 14 days in smooth muscle
differentiation medium (Fig. 7G).
Therefore, although
CD45−/CD31+/VEGFR2− LSP cells may
be smooth muscle progenitor cells, the
CD45−/CD31+/VEGFR2+ LSP cells
showed no such potential.
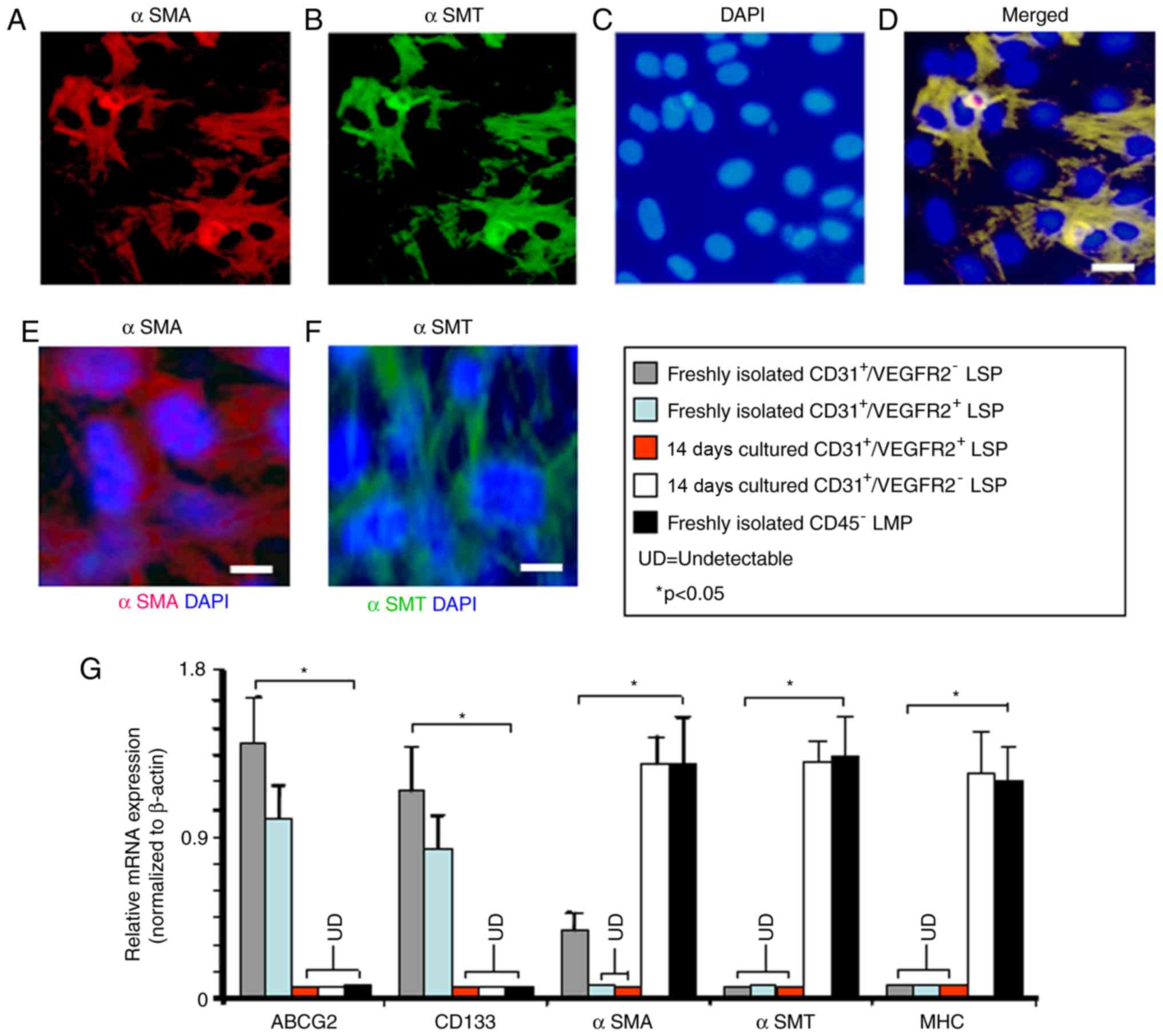 | Figure 7In vitro smooth muscle
differentiation potential of
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells.
Images (scale bar, 20 µm) show the representative
microscopic fields of
CD45−/CD31+/VEGFR2− LSP cells
double-labeled for αSMA and αSMT after 14 days in culture under
smooth muscle differentiation-inducing conditions; numerous cells
stained positively with (A) anti-αSMA antibody (red) and (B)
anti-αSMT antibody (green). (C) Nuclei were counterstained with
DAPI (blue). (D) Merge of the optical channels. Single
immunofluorescence labeling with (E) anti-αSMA (red) and (F)
anti-αSMT (green) is shown (scale bar, 10 µm). (G) Reverse
transcription-quantitative polymerase chain reaction analysis was
performed to compare gene expression by freshly isolated
CD45−/CD31+/VEGFR2− LSP cells
(grey bar), CD45−/CD31+/VEGFR2−
LSP cells after 14 days culture (white bar), freshly isolated
CD45−/CD31+/VEGFR2+ LSP cells
(light blue bar) and
CD45−/CD31+/VEGFR2− LSP cells
after 14 days culture (red bar). Freshly isolated CD45−
LMP cells are included as a control (black bar). Relative
expression levels of the ABCG2, CD133, αSMA, αSMT and MHC genes
were compared between the three types of cells. Data are presented
as the mean ± standard deviation (n=3 experiments).
*P<0.05; LSP, lung side population; LMP, lung main
population; UD, undetectable; ABCG2, ATP-binding cassette
super-family G member 2; VEGFR2, vascular endothelial growth factor
receptor 2; vWF, von Willebrand factor; αSMT, α-smooth muscle
tropomyosin; αSMA, α-smooth muscle actin; MHC, myosin heavy chain;
DAPI, 4-6-diamidino-2-phenylindol-dihydrochloride. |
Discussion
There have been several reports of the existence of
endogenous stem cell-like cell populations in the mouse lung
(14,25). In particular, the in vitro
differentiation of CD45−/CD31− LSP cells was
demonstrated by Summer et al (15). However, little is known about
CD45−/CD31+ LSP cells. The present study
provides new data showing that CD45−/CD31+
LSP cells can be divided into
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cell
subpopulations. To the best of our knowledge, this is the first
detailed in vitro investigation of the ability of
CD45−/CD31+ LSP cells from the adult mouse
lung to form cell colonies, differentiate into endothelial and
smooth muscle cells and vascularize. The results suggest that
CD45−/CD31+/VEGFR2+ LSP cells
differentiate into endothelial cells, whereas
CD45−/CD31+/VEGFR2− LSP cells can
differentiate into endothelial and smooth muscle cells.
The expression of CD31 in
CD45−/CD31+ LSP cells suggests that
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells may
be progenitors of lung endothelial cells. This was confirmed by
their gene expression profiles. The
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells
expressed ABCG2 and CD133 at high levels. The endothelial cell
marker vWF was undetectable in freshly isolated
CD45−/CD31+/VEGFR2− LSP cells. The
CD45−/CD31+/VEGFR2+ LSP cells
expressed relatively low mRNA levels of vWF, and no vWF protein was
detected. This phenotype is consistent with these SP cells being
endothelial stem/progenitor cells (27,36). Of note, the
CD45−/CD31+ LSP cells were capable of
DiI-Ac-LDL uptake, suggesting that they were endothelial
progenitors rather than hematopoietic progenitors. The expression
levels of ABCG2 and CD133 were significantly lower in the
CD45−/CD31+/VEGFR2+ LSP cells
compared with those in the
CD45−/CD31+/VEGFR2− LSP cells. In
addition, the CD45−/CD31+/VEGFR2−
LSP cells expressed αSMA, suggesting that these cells may serve as
progenitors for endothelial and smooth muscle cells. This
possibility is consistent with previous studies showing that
vascular smooth muscle cells are derived from endothelial
progenitor cells during vasculo-genesis (27,37). By contrast, the
CD45−/CD31+/VEGFR2+ LSP cells
expressed detectable levels of vWF and VEGFR2, but no αSMA,
indicating that these cells may be relative late commitment
endothelial progenitor cells.
The results of the present study showed that
CD45−/CD31+ LSP cells possessed a higher
colony-forming potential than CD45−/CD31+ LMP
cells. This finding is consistent with previous studies that
reported SP cells isolated from different tissues have higher
colony-forming capability than non-SP cells (19,27,38). A previous study showed that a
small number of cells isolated from the CD31+ population
from the adult mouse lung were endothelial progenitor cells
(39). This group of endothelial
progenitor cells may be CD45−/CD31+ LSP
cells. However, the data obtained in the present study do not rule
out the possibility that other populations of CD31+
cells function as endothelial progenitor cells.
In a previous study, Irwin et al (16) showed that
CD45−/VEGFR2+ LSP cells of the mouse lung
were able to differentiate into endothelial cells. However, whether
these cells expressed CD31 was unclear. The present study found
that it was possible to divide CD45−/CD31+
LSP cells into CD45−/CD31+/VEGFR2−
and CD45−/CD31+/VEGFR2+
sub-populations. Both the
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells were
capable of differentiation into mature endothelial cells, which
formed vascular tube-like structures in vitro. However, only
the CD45−/CD31+/VEGFR2− LSP cells
expressed αSMA and were capable of differentiation into mature
smooth muscle cells in vitro
(CD45−/CD31+/VEGFR2+ LSP cells did
not). When the CD45−/CD31+/VEGFR2−
LSP cells were cultured under endothelial differentiation
conditions for 2 days, the expression of αSMA was lost. They also
started to express VEGFR2 and vWF. The gene expression profiles of
those CD45−/CD31+/VEGFR2− LSP
cells that had been cultured under endothelial differentiation
conditions for 2 days became similar to that of
CD45−/CD31+/VEGFR2+ LSP cells
(Fig. 6C). Taken together, these
findings indicate that the
CD45−/CD31+/VEGFR2− LSP cells were
capable of differentiation into smooth muscle cells, endothelial
cells and CD45−/CD31+/VEGFR2+ LSP
cells in vitro. Therefore,
CD45−/CD31+/VEGFR2+ LSP cells may
be late commitment endothelial progenitors that are capable of
differentiating into endothelial cells, but not smooth muscle
cells, in vitro. However, the association between
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells
requires further elucidation. Several studies have shown that SP
cells from different tissues and organs have potential for
repairing damaged parts within a tissue or organ (40–42). The role of
CD45−/CD31+ LSP cells in repairing injured
lung tissue is to be the subject of a future investigation.
Our previous studies demonstrated that
CD31+/SCA1+ CSP cells served as endothelial
progenitor cells in the adult mouse heart (27,28). Of note,
SCA1+/CD31+ CSP cells share several features
with CD45−/CD31+ LSP cells
(CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2−), including
surface expression of SCA1, similarity in gene expression profiles,
capacity for colony formation and potential for endothelial
differentiation (27,28). Our findings support the hypothesis
that a novel population of progenitor cells may contribute to heart
and lung tissues, including their blood vessels (26). Our previous study also
demonstrated that vascular smooth muscle and airway smooth muscle
share a common progenitor cell in the lung. Further investigation
is required to determine whether
CD45−/CD31+/VEGFR2− LSP cells
differentiate into airway smooth muscle cells. Previous studies
have found that c-kit+ cells in the adult mouse lung may
serve as endothelial progenitor cells (11). However, the expression of c-kit
was undetectable in CD45−/CD31+ LSP cells.
Therefore, the association between lung c-kit cells and
CD45−/CD31+ LSP cells also requires further
investigation.
In conclusion, the present study isolated and
characterized CD45−/CD31+ SP cells from the
adult mouse lung. They were found to consist of
CD45−/CD31+/VEGFR2− and
CD45−/CD31+/VEGFR2+ LSP cells.
Based on their surface protein expression, their gene expression
profile, and capacity to form colonies and differentiate into
endothelial and smooth muscle cells in vitro, the results
suggest that CD45−/CD31+/VEGFR2−
LSP cells may serve as a source of endothelial and smooth muscle
progenitor cells in the adult mouse lung in vivo. The data
also suggest that
CD45−/CD31+/VEGFR2+ LSP cells may
be derived from
CD45−/CD31+/VEGFR2− LSP cells and
that this differentiation may involve a lineage commitment to
become endothelial progenitor cells. Understanding the function and
identifying ways to promote the proliferation and differentiation
of these populations of cells may offer potential for the
development of novel therapeutic strategies to enhance vessel
regeneration in lung injury and diseases.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81370619 to SXL).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
YX, PS and JYW performed experiments and wrote the
manuscript; ZZL and RLG performed experiments; XZW performed
analysis and interpretation of data; WDP analyzed data and edited
the manuscript; SXL designed and organized the entire research,
analyzed the data, and edited the manuscript. All the authors have
read and approved the final, submitted version of manuscript.
Ethics approval and consent to
participate
All animal experiments were conducted strictly
according to ethical standards approved by the Animal Ethical
Committee of Jinzhou Medical University (approval ID:
LY2014D001).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
SP
|
side population
|
|
LSP
|
lung side population
|
|
MP
|
main population
|
|
LMP
|
lung main population
|
|
VEGFR2
|
vascular endothelial growth factor
receptor 2
|
|
CSP
|
cardiac side population
|
|
FACS
|
fluorescence-activated cell
sorting
|
|
DMEM
|
Dulbecco's modified Eagle's
medium
|
|
Dil-Ac-LDL
|
Dil-labeled acetylated low-density
lipoprotein
|
|
DAPI
|
4-6-diamidino-2-phenylindol-dihydrochloride
|
|
vWF
|
von Willebrand factor
|
|
αSMT
|
α-smooth muscle tropomyosin
|
|
αSMA
|
α-smooth muscle actin
|
Acknowledgments
Not applicable.
References
|
1
|
Schatz M, Kazzi AA, Brenner B, Camargo CA
Jr, Corbridge T, Krishnan JA, Nowak R and Rachelefsky G: Joint task
force report: Supplemental recommendations for the management and
follow-up of asthma exacerbations. Introduction. J Allergy Clin
Immunol. 124(2 Suppl): S1–S4. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mosier JM, Hypes C, Joshi R, Whitmore S,
Parthasarathy S and Cairns CB: Ventilator strategies and rescue
therapies for management of acute respiratory failure in the
emergency department. Ann Emerg Med. 66:529–541. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yan Z, Kui Z and Ping Z: Reviews and
prospectives of signaling pathway analysis in idiopathic pulmonary
fibrosis. Autoimmun Rev. 13:1020–1025. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cao Z, Lis R, Ginsberg M, Chavez D, Shido
K, Rabbany SY, Fong GH, Sakmar TP, Rafii S and Ding BS: Targeting
of the pulmonary capillary vascular niche promotes lung alveolar
repair and ameliorates fibrosis. Nat Med. 22:154–162. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wong SP, Rowley JE, Redpath AN, Tilman JD,
Fellous TG and Johnson JR: Pericytes, mesenchymal stem cells and
their contributions to tissue repair. Pharmacol Ther. 151:107–120.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marriott S, Baskir RS, Gaskill C, Menon S,
Carrier EJ, Williams J, Talati M, Helm K, Alford CE, Kropski JA, et
al: ABCG2pos lung mesenchymal stem cells are a novel pericyte
subpopulation that contributes to fibrotic remodeling. Am J Physiol
Cell Physiol. 307:C684–C698. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hung C, Linn G, Chow YH, Kobayashi A,
Mittelsteadt K, Altemeier WA, Gharib SA, Schnapp LM and Duffield
JS: Role of lung pericytes and resident fibroblasts in the
pathogenesis of pulmonary fibrosis. Am J Respir Crit Care Med.
188:820–830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kramann R, Schneider RK, DiRocco DP,
Machado F, Fleig S, Bondzie PA, Henderson JM, Ebert BL and
Humphreys BD: Perivascular Gli1+ progenitors are key
contributors to injury-induced organ fibrosis. Cell Stem Cell.
16:51–66. 2015. View Article : Google Scholar
|
|
9
|
Hong KU, Reynolds SD, Giangreco A, Hurley
CM and Stripp BR: Clara cell secretory protein-expressing cells of
the airway neuroepithelial body microenvironment include a
label-retaining subset and are critical for epithelial renewal
after progenitor cell depletion. Am J Respir Cell Mol Biol.
24:671–681. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barkauskas CE, Cronce MJ, Rackley CR,
Bowie EJ, Keene DR, Stripp BR, Randell SH, Noble PW and Hogan BL:
Type 2 alveolar cells are stem cells in adult lung. J Clin Invest.
123:3025–3036. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Q, Huang X, Zhang H, Tian X, He L,
Yang R, Yan Y, Wang QD, Gillich A and Zhou B: c-kit(+) cells adopt
vascular endothelial but not epithelial cell fates during lung
maintenance and repair. Nat Med. 21:866–868. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kajstura J, Rota M, Hall SR, Hosoda T,
D'Amario D, Sanada F, Zheng H, Ogorek B, Rondon-Clavo C,
Ferreira-Martins J, et al: Evidence for human lung stem cells. N
Engl J Med. 364:1795–1806. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Giangreco A, Shen H, Reynolds SD and
Stripp BR: Molecular phenotype of airway side population cells. Am
J Physiol Lung Cell Mol Physiol. 286:L624–L630. 2004. View Article : Google Scholar
|
|
14
|
Liang SX, Summer R, Sun X and Fine A: Gene
expression profiling and localization of Hoechst-effluxing
CD45− and CD45+ cells in the embryonic mouse
lung. Physiol Genomics. 23:172–181. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Summer R, Fitzsimmons K, Dwyer D, Murphy J
and Fine A: Isolation of an adult mouse lung mesenchymal progenitor
cell population. Am J Respir Cell Mol Biol. 37:152–159. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Irwin D, Helm K, Campbell N, Imamura M,
Fagan K, Harral J, Carr M, Young KA, Klemm D, Gebb S, et al:
Neonatal lung side population cells demonstrate endothelial
potential and are altered in response to hyperoxia-induced lung
simplification. Am J Physiol Lung Cell Mol Physiol. 293:L941–L951.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hierlihy AM, Seale P, Lobe CG, Rudnicki MA
and Megeney LA: The post-natal heart contains a myocardial stem
cell population. FEBS Lett. 530:239–243. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martin CM, Meeson AP, Robertson SM, Hawke
TJ, Richardson JA, Bates S, Goetsch SC, Gallardo TD and Garry DJ:
Persistent expression of the ATP-binding cassette transporter,
Abcg2, identifies cardiac SP cells in the developing and adult
heart. Dev Biol. 265:262–275. 2004. View Article : Google Scholar
|
|
19
|
Pfister O, Mouquet F, Jain M, Summer R,
Helmes M, Fine A, Colucci WS and Liao R: CD31− but Not
CD31+ cardiac side population cells exhibit functional
cardiomyogenic differentiation. Circ Res. 97:52–61. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou S, Schuetz JD, Bunting KD, Colapietro
AM, Sampath J, Morris JJ, Lagutina I, Grosveld GC, Osawa M,
Nakauchi H, et al: The ABC transporter Bcrp1/ABCG2 is expressed in
a wide variety of stem cells and is a molecular determinant of the
side-population phenotype. Nat Med. 7:1028–1034. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goodell MA, Brose K, Paradis G, Conner AS
and Mulligan RC: Isolation and functional properties of murine
hematopoietic stem cells that are replicating in vivo. J Exp Med.
183:1797–1806. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Asakura A and Rudnicki MA: Side population
cells from diverse adult tissues are capable of in vitro
hematopoietic differentiation. Exp Hematol. 30:1339–1345. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang SX and Phillips WD: Migration of
resident cardiac stem cells in myocardial infarction. Anat Rec.
296:184–191. 2013. View Article : Google Scholar
|
|
24
|
Summer R, Kotton DN, Liang S, Fitzsimmons
K, Sun X and Fine A: Embryonic lung side population cells are
hematopoietic and vascular precursors. Am J Respir Cell Mol Biol.
33:32–40. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Summer R, Kotton DN, Sun X, Ma B,
Fitzsimmons K and Fine A: Side population cells and Bcrp1
expression in lung. Am J Physiol Lung Cell Mol Physiol.
285:L97–L104. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peng T, Tian Y, Boogerd CJ, Lu MM, Kadzik
RS, Stewart KM, Evans SM and Morrisey EE: Coordination of heart and
lung co-development by a multipotent cardiopulmonary progenitor.
Nature. 500:589–592. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang SX, Khachigian LM, Ahmadi Z, Yang M,
Liu S and Chong BH: In vitro and in vivo proliferation,
differentiation and migration of cardiac endothelial progenitor
cells (SCA1+/CD31+ side-population cells). J
Thromb Haemost. 9:1628–1637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang XZ, Gao RL, Sun P, Liu S, Xu Y, Liang
DZ, Yin LM, Phillips WD and Liang SX: Proliferation,
differentiation and migration of SCA1−/CD31−
cardiac side population cells in vitro and in vivo. Int J Cardiol.
227:378–386. 2017. View Article : Google Scholar
|
|
29
|
Liang SX, Tan TY, Gaudry L and Chong B:
Differentiation and migration of Sca1+/CD31−
cardiac side population cells in a murine myocardial ischemic
model. Int J Cardiol. 138:40–49. 2010. View Article : Google Scholar
|
|
30
|
Sun P, Wu G, Gao RL, Liu S, Phillips WD
and Liang SX: Rifampicin-dependent antibodies target glycoprotein
IIb/IIIa and cause clearance of human platelets in NOD/SCID mice.
Br J Haematol. 172:137–140. 2016. View Article : Google Scholar
|
|
31
|
Qiagen: QuantiTect Primer Assay Handbook.
https://www.qiagen.com/sg/resources/resourcedetail?id=882a8baa-29df-4182-a2a5-17083f4dbe11&lang=en.
Accessed September 18, 2013.
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Gehling UM, Ergun S, Schumacher U, Wagener
C, Pantel K, Otte M, Schuch G, Schafhausen P, Mende T, Kilic N, et
al: In vitro differentiation of endothelial cells from
AC133-positive progenitor cells. Blood. 95:3106–3112.
2000.PubMed/NCBI
|
|
34
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Low RB and White SL: Lung smooth muscle
differentiation. Int J Biochem Cell Biol. 30:869–883. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Choi K, Kennedy M, Kazarov A,
Papadimitriou JC and Keller G: A common precursor for hematopoietic
and endothelial cells. Development. 125:725–732. 1998.PubMed/NCBI
|
|
37
|
Ema M, Faloon P, Zhang WJ, Hirashima M,
Reid T, Stanford WL, Orkin S, Choi K and Rossant J: Combinatorial
effects of Flk1 and Tal1 on vascular and hematopoietic development
in the mouse. Genes Dev. 17:380–393. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hirschmann-Jax C, Foster AE, Wulf GG,
Nuchtern JG, Jax TW, Gobel U, Goodell MA and Brenner MK: A distinct
'side population' of cells with high drug efflux capacity in human
tumor cells. Proc Natl Acad Sci USA. 101:14228–14233. 2004.
View Article : Google Scholar
|
|
39
|
Schniedermann J, Rennecke M, Buttler K,
Richter G, Stadtler AM, Norgall S, Badar M, Barleon B, May T,
Wilting J, et al: Mouse lung contains endothelial progenitors with
high capacity to form blood and lymphatic vessels. BMC Cell Biol.
11:1471–2121. 2010. View Article : Google Scholar
|
|
40
|
Ge G, Zhang H, Li R and Liu H: The
function of SDF-1CXCR4 axis in sp cells-mediated protective role
for renal ischemia/reper-fusion injury by SHH/GLI1-ABCG2 Pathway.
Shock. 47:251–259. 2017. View Article : Google Scholar
|
|
41
|
He DN, Qin H, Liao L, Li N, Zhu WM, Yu BJ,
Wu X, Zhao RC and Li JS: Small intestinal organoid-derived SP cells
contribute to repair of irradiation-induced skin injury. Stem Cells
Dev. 14:285–291. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ooka H, Kanda S, Okazaki H, Suzuki H,
Mishima K, Saito I, Yagi M, Tomoda K and Nishiyama T:
Characterization of side population (SP) cells in murine cochlear
nucleus. Acta Otolaryngol. 132:693–701. 2012. View Article : Google Scholar : PubMed/NCBI
|