In this review, the basic components of the ETC are
discussed and the process of electron transfer in each complex,
including the structure, composition and function of each complex
is reviewed. In addition, the ROS generation sites in the ETC are
summarized and the ROS regulation is mentioned. Moreover, proton
leak is emphatically introduced, including the structure, tissue
distribution, functions and regulatory factors of UCPs. The
diseases implicated in ROS or UCPs are simply summarized.
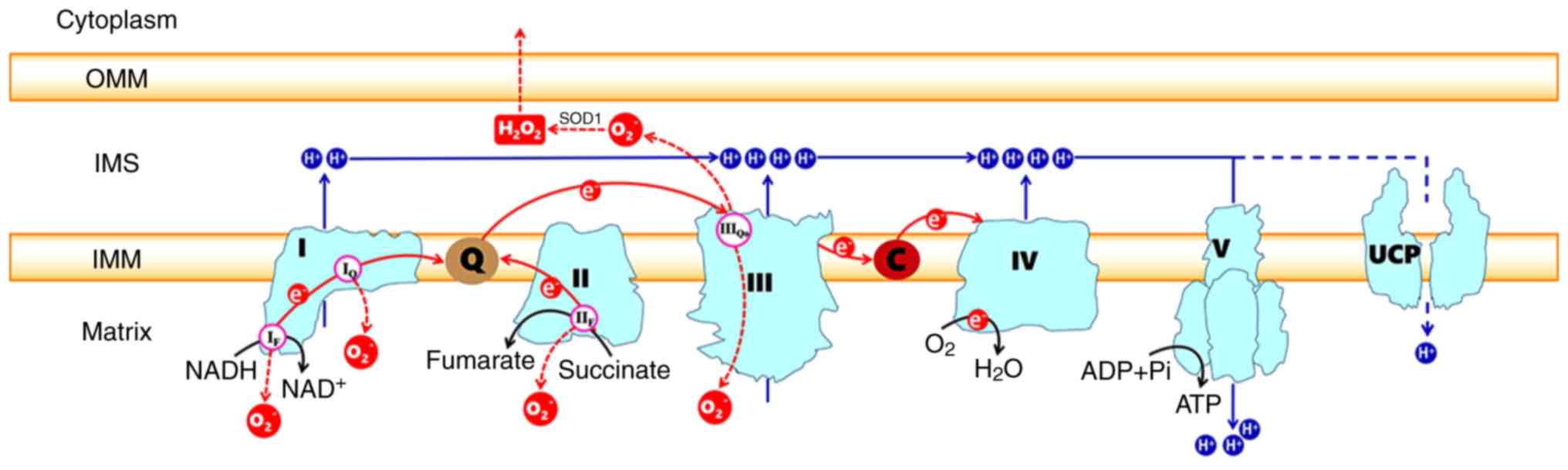
The ETC, which is composed of transmembrane protein
complexes (I-IV) and the freely mobile electron transfer carriers
ubiquinone and cytochrome c, exists in the folded inner membranes
called cristae (Fig. 1). The
complexes must be assembled into a specifically configured
supercomplex to function properly (2,3).
These assembled components together with
F1F0ATP synthase (namely, complex V) become
the basis of ATP production during oxidative phosphorylation
(OXPHOS). To better understand the whole process of how electron
transportation produces ATP via the ETC, it is necessary to know
the ultrastructure and function of the individual complexes.
CI, also called NADH-ubiquinone oxidoreductase, is
the largest multisubunit enzyme complex in the ETC. The key role of
CI is to transfer electrons from matrix NADH to ubiquinone, as the
name implies. A number of studies have reported the structure of
the bacterial mitochondrial CI using X-ray crystallography at a
nearly atomic resolution (4,5).
Mitochondria from the Bos taurus heart have been regarded as
the best model for human CI (6-9).
These studies demonstrate that the L-shaped eukaryotic CI contains
two domains: The membrane arm embedded in the inner membranes and
the matrix arm protruding into the matrix. The two domains are
mainly composed of 14 core subunits that are conserved from
bacterial CI and are the core of the enzymatic reaction. There are
45 clearly identified proteins that participate in the formation of
the core subunits. The matrix arm contains seven core subunits
(NDUFS1, NDUFS2, NDUFS3, NDUFS7, NDUFS8, NDUFV1 and NDUFV 2) that
contain the following cofac-tors: A flavin mononucleotide (FMN)
molecule; 7-9 FeS clusters [including the (2Fe-2S)N1b,
(4Fe-4S)N3, (4Fe-4S)N4,
(4Fe-4S)N5, (4Fe-4S)N6a/b and
(4Fe-4S)N2 clusters] (4,10);
and the final electron accepting iron-sulfur cluster (N2 cluster),
which was recently found to deliver electrons to ubiquinone binding
sites (11). The membrane arm
contains seven hydrophobic subunits (ND1-6 and ND4L), all of which
are encoded by the mitochondrial genome. In addition, a large
number of accessory subunits are arranged around the core subunits.
The assembly of these modules has been reviewed in detail elsewhere
(12). An FMN bound at the cusp
of the matrix arm could form FMNH2 by accepting a pair
of electrons derived from matrix NADH, which is primarily produced
by the tricarbox-ylic acid (Krebs) cycle that continuously occurs
in the matrix. These interactions also mean that electrons go into
the ETC and are then passed to ubiquinone via a chain of
iron-sulfur clusters arranged from low to high potential [the
transfer order was reported as FMN→N3→N1b→N4→N5→N6a→N6b→ N2
(4)]. The ubiquinone binding site
is located at the junction of the membrane arm and matrix arm, in
which ubiquinone (CoQ) is reduced to ubiquinol (QH2).
Then, the conformational changes of the N2 cluster induce the
formation of a proton translocation channel by the ND1, ND3, ND6
and ND4L subunits near the CoQ binding site (13). The energy released by the transfer
of a pair of electrons from NADH to CoQ in CI probably (not
definitively) induce the pumping of four protons from the matrix
into the intermembrane space (14-17). Several hypotheses exist in current
research: Ohnishi (18) proposed
a hypothesis that two protons are indirectly pumped out in a
conformation-coupled manner and that the other two protons are
directly pumped out by the induction of ubiquinone redox. Sazanov
and Hinchliffe (4) hypothesized
that three protons are indirectly pumped via three antiporter
homologs, and the final proton is shifted in an unclear way. In
addition, Tan et al (14)
speculated that the conformation changes and the density of water
molecules in the trans-membrane domain determine the proton
translocation in CI. However, how the energy transfers from the
redox reaction to proton translocation are still unknown.
CII, namely, succinate dehydrogenase, is a component
of the Krebs cycle as well as the ETC, serving as a link between
metabolism and OXPHOS (19,20). As a part of the Krebs cycle, CII
catalyzes the oxidation of succinate to fumarate. CII is another
entry point for electrons and donates them from succinate to CoQ
via FeS clusters, similar to CI. CII consists of four subunits
(20). A total of two of the
subunits, the membrane-anchor proteins CybL and CybS, are
hydrophobic, anchor the complex to the inner membrane, and contain
the CoQ binding site. The other two subunits are located on the
matrix side of the inner membrane and contain the binding site of
the substrate succinate, three FeS clusters [(2Fe-2S), (4Fe-4S) and
(3Fe-4S)], and a flavoprotein covalently bound to a FAD cofactor.
The assembly steps of the four subunits are detailed elsewhere
(21). FAD is reduced to
FADH2 after receiving electrons from succinate and then
transfers the electrons to FeS clusters. Then, CoQ is reduced to
QH2 after obtaining the electrons from the FeS cluster
(3Fe-4S) (22). Electron
transport in CII is not accompanied by the translocation of
protons.
CIV, also known as cytochrome c oxidase, transfers
electrons from cytochrome c to the terminal electron acceptor
O2 to generate H2O. Mammalian CIV consists of
13 different subunits containing four redox-active metal centers,
namely, CuA, heme a (Fea) and a binuclear
center composed of heme a3 (Fea3) and
CuB (29,30). Subunits I, II, III are encoded by
mitochondrial DNA and are the core subunits, while the 10
nuclear-coded subunits are the accessory subunits (31,32). Subunit I contains three of the
four cofactors, heme a and the binuclear center, which transfers
electrons from heme a to O2 (29). Subunits II and III are located on
both sides of subunit I and there are two CuA cofactors
on the side of the intermembrane space of subunit II. Subunit III
stabilizes the other two core proteins and is mainly involved in
proton pumping (33,34). The nuclear-coded subunits
participate in the modulation of physiological activity via the
allosteric ATP-mediated inhibition of CIV, which depends on the
ATP/ADP-ratio (35-39).
Cytochrome c, similar to CoQ, is a mobile electron
carrier that is loosely connected to the outer surface of the inner
mitochondrial membrane by electrostatic interactions, allowing it
to interact with the cytochrome c1 of CIII and to accept
electrons (39). The reduced
cytochrome c moves along the surface of the membrane and interacts
with subunit II of CIV by electrostatic interactions,
simultaneously transmitting electrons to the CuA site of
subunit II, and then the electrons are passed from heme a to the
binuclear center of subunit I (29,39), where the O2 is reduced
to H2O. A total of four electrons at a time from
cytochrome c are almost simultaneously transferred to bind
dioxygen; eight protons in total are removed from the matrix, of
which half are used to form the two water molecules and the other
four are pumped across the membrane into the IMS (40).
In conclusion, the entire composition of each
individual complex has been well described over the past century
and it is now widely accepted that these complexes must establish
interactions and form supercomplexes to perform their function. Due
to the application of cryo-electron microscopy, a greater
understanding of the high-resolution structure of these complexes
has been gained (45-47).
Mitochondria are a main source of cellular ROS.
Under physiological conditions, 0.2-2% of the electrons in the ETC
do not follow the normal transfer order but instead directly leak
out of the ETC and interact with oxygen to produce superoxide or
hydrogen peroxide (48,49). A total of 11 sites that produce
superoxide (O2−) and/or hydrogen peroxide
(H2O2) that are associated with substrate
oxidation and the ETC have currently been identified in mammalian
mitochondria (50). Sites
OF, PF, BF and AF are
in the 2-oxoacid dehydrogenase complexes, sites IF and
IQ are in CI, site IIIQo is in CIII, and
sites IIF, GQ, EF and
DQ are linked to the Q-dependent dehydrogenases in the
QH2/Q pool (50). The
occurrence of numerous diseases and hypoxia are closely related to
the increase of ROS production. CI and CIII, especially CI, are
considered to be the main sites of ROS production in mitochondria
(51,52).
CIII produces small amounts of ROS, which could be
overlooked compared to the ROS production of CI (52). CIII transfers electrons through
the Q-cycle. In this process, ubisemiquinone (QH−) of
the Qo site carrying a single electron can move freely
in CIII, directly leaking the single electron to O2,
forming ROS through a nonenzymatic reaction (58,59). The formed ROS can be released into
both the matrix and IMS despite the location of the Qo
site on the IMS side of the inner mitochondrial membrane. Muller
et al (60) built two
models explaining how superoxide can reach the matrix. The
O2− released into the IMS can be converted to
the relatively more stable form of H2O2 by
superoxide dismutase (SOD) enzymes (Fig. 1). This permanent and stable
oxidant molecule, which freely disperses through the outer membrane
of mitochondria, acts as an intracellular signaling molecule,
physiologically functioning via the direct modification of amino
acids (61). However, supporting
evidence demonstrates that O2−can permeate
through the mitochondrial membrane into the cytosol through anion
channels (62). Treberg et
al (63) experiments in the
mitochondria of wild-type rat skeletal muscle proved that ~63% of
ROS are produced in the matrix. Antimycin A can specifically block
the Qi site of CIII, resulting in the stalling of
electrons on the QH- at site IIIQo, which
could react with O2 to generate ROS (64,65). As specific inhibitors of the
Qo site, stigmatellin and myxothiazol can block the
binding of QH2 to the Qo site, which also
blocks the transfer of electrons into CIII, thereby preventing the
production of ROS in CIII (64).
Previously, a chemical suppressor of site IIIQo electron
leak called S3QELs was screened and found to specifically suppress
the ROS formation at site IIIQo without affecting
electron transport or the redox states of other centers (66).
In the past, it was believed that ROS were
exclusively harmful to cells. However, recent studies have
demonstrated that ROS appear to be very important second messengers
that mediate different intracellular pathways (50,61,68). ROS act through the oxidative
modification of numerous types of proteins, particularly receptors,
kinases, phosphatases, caspases, ion channels and transcription
factors (68). The ROS produced
from CIII are necessary for HIF-1α stabilization and consequently,
for the proliferation of cells, including vascular smooth muscle
cells, endothelial cells and erythroid progenitors (69). There is ample evidence that ROS
are also involved in different protein kinase signaling cascades,
such as the protein kinase B (AKT), AMP-activate protein kinase
(AMPK) and mitogen-activated protein kinase kinase
kinase/mitogen-activated protein kinase 8 pathways, changing the
fate of cells between autophagy and apoptosis [(Table I and (70)]. Under hypoxic conditions, ROS
activate AMPK, which can upregulate cytoprotective autophagy by
inhibiting downstream mammalian target of rapamycin activity
(71). ROS have also been
demonstrated to regulate synaptic plasticity-related signalling
molecules, receptors and channels, including N-methyl-d-aspartate
receptor (72), Ca2+
channel (73,74), Ca2+ kinase II (CaMKII)
(75), extracellular
signal-regulated kinase (76) and
cyclic adenosine monophosphate (cAMP) response element binding
protein (CREB) (74,77). ROS are also necessary for
long-term potentiation, a phenomenon of synaptic plasticity widely
regarded as one of the main molecular mechanisms that form the
basis of learning and memory (77,78). Physiological levels of ROS can
promote the establishment of neuronal polarity and regulate
neuronal cytoskeletal organization and dynamics by regulating
intracellular Ca2+ release (79-81).
The amount of ROS generated as a result of a
stimulus determines whether ROS play beneficial or harmful roles,
which means different physiological or pathological pathways are
activated. A large amount of ROS cause lipid peroxidation, DNA
damage, protein oxidation, irreversible impairment of mitochondria,
insufficient ATP generation and, eventually, cell death (82). The ROS-mediated activation of
NHE-1 is implicated in cardiac hypertrophy (83). In addition, the activation of
CaMKII by ROS contributes to an increase in cardiomyocyte death and
the development of heart failure (84). ROS are involved in a number of
chronic inflammatory diseases, particularly atherogenesis, through
activating the NF-κB pathway (85). In addition, it is widely known
that the ROS burst during reperfusion plays a critical role in
ischemia-reperfusion injury (86). Table II summarizes the pathologies in
which ROS has been implicated.
OXPHOS is not completely coupled. Under routine
circumstances, a small number of protons do not pass through ATP
synthase and instead flow directly into the mitochondrial matrix
across the inner mitochondrial membrane, without the generation of
ATP, in a process known as proton leak. In the concept of
'respiratory state' proposed by Chance and Williams (16), mitochondrial respiration persists
in the absence of ADP (state 4) and reflects the oxygen consumption
of proton leak. The existence of proton leak can also be proven by
the collapse of the proton gradient (Δp) in the presence of the ATP
synthase inhibitor oligomycin in isolated mitochondria (137).
It was found that the proton leak of the inner
mitochondrial membrane demonstrated nonohmic conductivity (137). According to Ohm's Law (R=U/I),
the resistance of a conductor is fixed and the electric current
increases linearly with increasing voltage. However, the rate of
proton leak increases exponentially with increasing ΔΨ. That is,
the proton conductivity increases when ΔΨ is high. The existence of
nonohmic conductivity indicates that there is a bidirectional
self-regulatory mechanism between electron transport and proton
re-entry: Protons are pumped out of the matrix into the IMS driven
by the electron transport in CI, CIII, CIV, and ΔΨ is gradually
elevated. On one hand, the elevated ΔΨ inhibits the transfer of
electrons to further elevate ΔΨ, through which the inner membrane
is protected from electric shock and maintains suitable ΔΨ. On the
other hand, the exorbitant ΔΨ can cause the increase in proton leak
to decrease.
Proton leak consists of two parts: Basal proton
leak and inducible proton leak. Basal proton leakage is not
regulated and is related to the lipid bilayer of the inner
mitochondrial membrane and the adenine nucleotide translocase
(ANT). Induced proton leak is precisely regulated and can be
catalyzed or suppressed by uncoupling proteins (UCPs) and ANT.
Basal proton leak has an important relationship
with the basal metabolic rate (BMR) in mammals at rest. The lower
the BMR of a species, the weaker the basal proton conductance.
Studies have demonstrated that the extent of basal proton leak
among species has a phylogenetic relationship (138,139). Although the lipid bilayer can
significantly increase proton conductivity, only ~5% of basal
proton leak is mediated by lipid bilayers (140) and most of the basic proton leak
is correlated with ANT (141).
UCP1, which is abundant in brown adipose tissue (BAT), may also be
involved in basal proton leak (142), although there remains
controversy (143). In
particular, proton leak through ANT or UCP1 is independent of
protein activity, as proton leak still occurs in the presence of
the ANT inhibitor carboxyatractylate and the UCP1 inhibitor GDP
(141,144).
The majority of the induced proton leak is
catalyzed by UCPs. UCPs belong to the family of mitochondrial anion
carrier proteins, through which the protons can reflux into the
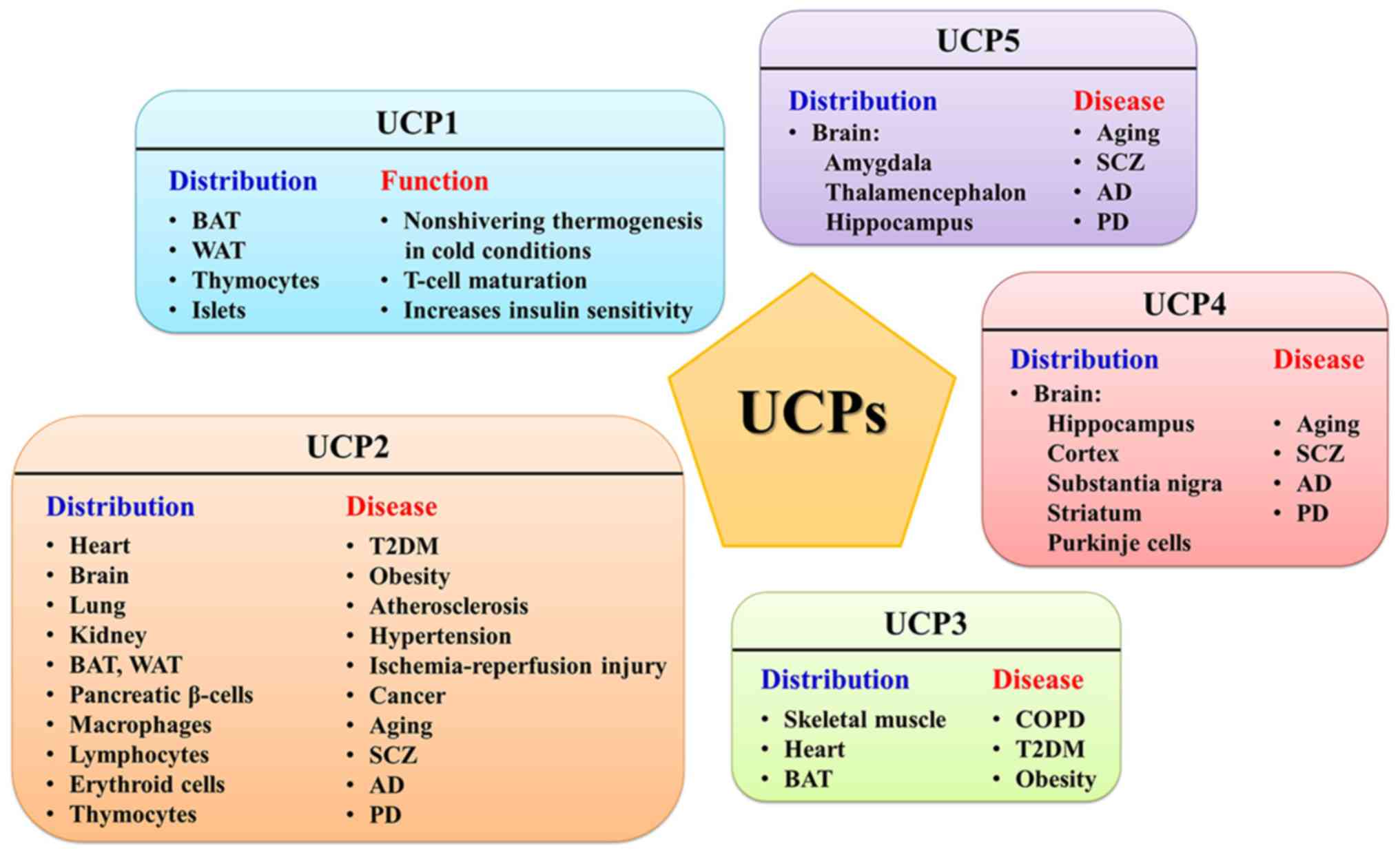
matrix. To date, five UCPs have been identified in mammals, UCP1,
UCP2, UCP3, UCP4 and UCP5, and all are present in the form of
dimers in the inner mitochondrial membrane (145). These UCPs have a purine
nucleotide binding site located on a projection in the IMS
(146). The purine nucleotides
(ATP, ADP, GTP and GDP) are inhibitors of UCP-mediated proton flux,
whereas ROS and fatty acids are activators (147,148). In addition to the role of
uncoupling, UCPs may also participate in other processes, such as
the regulation of calcium homeostasis, ion transportation or
synaptic plasticity (149,150).
UCP1 is mainly expressed in BAT, which converts
stored energy in Δp into heat for thermogenesis (151). UCP1 can also be detected in the
beige adipocytes of white adipose tissue (WAT) during thermal
acclimation under specific conditions (152). The genetic deletion of UCP1
severely inhibits cold adaptive thermogenesis and diet-induced
adrenergic thermogenesis, and UCP1-null mice develop fatal
hypothermia upon cold exposure (153,154). Interestingly, WAT can exert
nonshivering thermogenesis with a UCP1-independent pathway
(155,156). UCP1 has also been found in
thymocytes and demonstrated to be involved in the maturation and
fate determination of developing T-cells (157-159). Sale et al (160) found that UCP1 is expressed in
islets and associates with the acute insulin response to
glucose.
UCP1-catalyzed proton leak could be activated by
long chain free fatty acids and inhibited by purine nucleotides
(161). Acute cold or
overfeeding stimuli induce the release of norepinephrine by
sympathetic nerves and then induce cAMP-responsive pathways through
β3-adrenergic receptors on brown adipocytes, which could further
promote the transcription of UCP1 and lipolysis for more free fatty
acids (162). There are
currently three models for the regulated mechanism of
UCP1-implicated proton leak (162-169).
UCP2 and UCP3, paralogues of UCP1, exhibit ~60%
sequence identity with UCP1 and ~70% identity with each other
(170,171). UCP2 is rather ubiquitous,
expressed in WAT, BAT, macrophages, erythroid cells, thymocytes and
pancreatic β-cells as well as heart, brain, lung, kidney and
lymphocytes (172-176). The UCP3 gene is mainly expressed
in skeletal muscle, BAT and heart (177,178) and has also been detected in the
thymus, spleen (179) and skin
cells (180). Studies with
UCP2/3-ablated mice have demonstrated that UCP2 and UCP3 are not
implicated in adaptive thermogenesis or the regulation of body
weight (170,181). However, the role of UCP2 and
UCP3 in inhibiting the production of ROS in mitochondria by
reducing ΔΨ is widely accepted (182). A strong correlation between ROS
production and mitochondrial membrane potential (ΔΨ) has already
been confirmed (183).
Experiments have demonstrated that ROS production is increased in
UCP2/3-ablated mice (184-186). ROS-induced lipid hydroperoxides
such as hydroxynonenal can activate UCP2/3-mediated proton leak,
but the mechanism remains uncertain (178). By reducing ΔΨ, the transfer of
electrons in the ETC can be accelerated and the likelihood of
electrons being directly transferred to O2 can be
minimized. Therefore, mild uncoupling is a feedback mechanism
adopted by the body to prevent excessive ROS in the mitochondria,
which was termed 'uncoupling to survive' (187). However, whether UCP1 is
implicated in the regulation of ROS in BAT is still controversial
(175,188,189). In addition to the function of
reducing the generation of ROS, UCP3 has been demonstrated to be
involved in exporting mitochondrial fatty acid anions to the
cytoplasm, thereby protecting the mitochondrial against lipid
peroxide-induced damage (190,191).
UCP4 and UCP5 (also called brain mitochondrial
carrier protein 1), which have 30% homology to UCP1 (192), are primarily expressed in the
central nervous system of mammals (193,194). Although UCP4 and UCP5 are more
widely distributed in the brain than UCP2, less is known about
their function. UCP4 was first detected in the brain, but it has
recently been found in adipocytes (195). In addition, UCP4 also plays a
predominant role in insect mitochondria (196). On the other hand, UCP5, which is
not limited to the brain, is also expressed in the testis, uterus,
kidney, lung, stomach, liver and heart (149). It has been demonstrated that
neuronal UCP4 and UCP5 share similar conformational and proton
transport activities with UCP1-UCP3 (149,197). Although UCP4 and UCP5 may play
an unconfirmed role in the neural system, their function for
reducing oxidative stress is clear (195,198). Hoang et al (149) speculated that UCP4 acts in a
neuroprotective role during early neuronal development, while UCP2
and UCP5 provided the protective function of restricting ROS
production in developed neurons and other tissues, based on the
observation that UCP2 and UCP5 displayed higher proton transport
rates than UCP4. Oxidative stress has been proven to be involved in
both neurodegenerative diseases and aging, so the UCP-dependent
reduction of ROS in the nervous system has the potential to be
neuroprotective in diseases such as Alzheimer's disease,
Parkinson's disease and amyotrophic lateral sclerosis (199,200). Certain evidence indicates that
protein-protein interactions exist between UCP4 and CII:
UCP4-overexpressing neuroblastoma cells increase ATP synthesis via
increasing the succinate-induced respiration mediated by CII
(201). Pfeiffer et al
(202) demonstrated that the
Caenorhabditis elegans UCP4 plays a novel role in the
regulation of CII by controlling succinate transport into
mitochondria. UCP4 was also deemed to regulate calcium homeostasis
in neuronal cells (203).
Regardless, as an inner mitochondrial membrane
protein, UCP1 mainly plays a role in the maintenance of body
temperature in a cold environment through non-shivering
thermogenesis and UCP2-5 can protect cells from oxidative stress by
reducing the mitochondrial membrane potential via mediating
uncoupling. Due to their wide distribution, UCPs have different
physiological significance in specific tissues. Therefore, abnormal
changes in UCPs in each tissue will lead to tissue-specific
diseases (Fig. 2).
The ubiquitous UCP2 is associated with a number of
metabolic diseases, such as diabetes, obesity, cardiovascular
disease and even cancer, which has created immense interest in
exploring the relationship between UCP2 and these diseases. Since
UCP2 can regulate fatty acid and lipid metabolism, a number of
studies have confirmed that UCP2 overexpression is associated with
diet-induced obesity (204-206). UCP2 is highly expressed in
pancreatic β-cells and has a negative regulatory effect on insulin
secretion. Robson-Doucette et al (207) in 2011 demonstrated that the
overexpression of UCP2 could reduce the secretion of
glucose-induced insulin and subsequently induce type II diabetes
(T2DM). Moreover, UCP2 knockout mice exhibited hyperinsulinemia and
hypo-glycaemia (208). Briefly,
glucose is metabolized through the ETC to increase ATP production,
which leads to the release of insulin and the production of ROS.
Chronic elevated glucose status can lead to the excessive
expression of UCP2 to reduce the overproduction of ROS, resulting
in reduced ATP production, reduced insulin secretion and,
eventually, progression to diabetes (209,210). Chronic inflammation, including
atherosclerosis, hypertension, diabetic vasculopathy and
ischemia-reperfusion injury, is accompanied by excessive ROS
production, which means that UCP2 can play a protective role in
these diseases by reducing oxidative stress. The signaling
pathways, such as nuclear factor (NF)-κB and p53, that can lead to
cellular senescence, inflammation, and irreversible
vasoconstriction can be inhibited by reducing ROS production. A
study showed that the protein levels of UCP2 were significantly
higher in human tumor tissues from the head and neck, skin,
prostate and pancreas (211).
Although the role of UCP2 in tumors is intuitive, the regulatory
effects of UCP2 on cellular glucose and lipid metabolism, as well
as the regulation of cellular oxidative stress, may be related to
the development of tumors. Several studies have confirmed that UCP2
overexpression provides protection for tumor cells and leads to
chemoresistance (212-214).
UCP3 has been verified to be associated with
exercise intolerance in chronic obstructive pulmonary disease
(COPD) patients. COPD patients have exhibited impairment in the
form of exercise intolerance, which was linked to increased levels
of intramuscular lipid peroxidation products (114,215). Given the fiber-type-specific
expression of UCP3, researchers have examined UCP3 levels in muscle
biopsies from COPD patients and found that UCP3 content was reduced
in COPD (216). It can be
speculated that low muscle UCP3 levels contribute to impaired
exercise tolerance in COPD patients based on the function of fatty
acid anion transportation. In addition, a number of studies have
demonstrated that the accumulation of lipid peroxide damage,
resulting from decreased UCP3 in skeletal muscle, leads to
excessive oxidative stress and is a crucial aspect in the
development and progression of obesity and T2DM (191,217,218).
UCP4, UCP5, together with UCP2, are expressed in
the nervous system and are implicated in several neurological
disorders, such as schizophrenia (SCZ), Alzheimer's disease (AD),
and Parkinson's disease (PD). Various studies link UCP2 with
neurodegeneration and aging (123,124,219). The three UCPs exert
neuroprotective effects mainly through the alleviation of oxidative
stress. The results from selected single nucleotide polymorphism
markers within the neuronal UCPs showed that UCP2 and UCP4 are
important in the genetic etiology of SCZ (219). Surprisingly, despite the
downregulation of UCP2 mRNA levels in SCZ patients, Gigante et
al (220) found that there
were no differences in UCP2 protein between patients and controls.
Future studies will be necessary to clarify whether the mechanism
of UCP2 is protective and opposes SCZ progression. Furthermore, in
the brains of AD patients, the expression levels of UCP2, 4, and 5
were significantly reduced, which limited the activation of
cytoprotective mechanisms to slow the progress of AD (124). UCP5 and, especially, UCP4 are
linked to PD. UCP4 is regulated by the oxidized DJ-1 and partially
via the NF-κB pathway and can protect against oxidative stress and
stabilize Ca2+ homeostasis in PD, as demonstrated by
Ramsden et al (148) in
2012. Drugs that induce neuronal UCP expression might represent
another effective strategy to ameliorate PD.
In conclusion, the ETC is the core component of
mitochondria. The OXPHOS process in the ETC, coupled with the
generation of ATP, also results in ROS production. As a
double-edged sword, ROS can play a role in signaling pathways, but
ROS overproduction can cause cellular damage. The ROS produced by
CIII is not only released into the matrix but also released into
the IMS. The ROS released into the IMS can be converted to
H2O2 in a reaction catalyzed by SOD1, and the
H2O2 may diffuse out of the mitochondria and
play an important role in physiological and pathological pathways.
Therefore, the artificial regulation of ROS at the CIII site (site
IIIQo) may be of great significance. Although the
precise mechanism of ROS production is still unclear, the use of
specific ROS inhibitors to reduce the excessive production of ROS
under pathological conditions has ameliorated oxidative
stress-mediated diseases. UCP-mediated proton leak is a positive
feedback mechanism for the protection of cells against oxidative
stress due to the rapid production of ATP. The proper activation of
UCPs can reduce the production of cellular ROS without causing a
decrease in ATP; however, when the expression of UCPs becomes too
high or too low or the UCP genes are mutated, pathological effects
that are involved in various diseases can occur. UCP1 mediates the
inducible proton conductance that is responsible for non-shivering
thermogenesis in BAT, a critical response to prolonged cold
exposure. UCP2 is involved in a variety of diseases, such as
diabetes, obesity and cardiovascular disease. In addition, UCP2,
UCP4 and UCP5 play an important role in neuroprotection and are
associated with neurological diseases such as SCZ, AD and PD. Drugs
targeting UCP expression and activity might represent as an
effective strategy to ameliorate these diseases. However, the
detailed mechanisms of the role of UCPs and the regulation of UCP
expression under normal and stressful situations warrant further
exploration.
The present study was supported by the National
Natural Science Foundation of China (grant no. 81571844).
Not applicable.
ZBY and LZ conceived the review and analyzed the
relevant literature. RZZ and SJ collected and reviewed the
literature related to the topic of this manuscript and drafted the
main part of this manuscript. RZZ, LZ and ZBY critically revised
the manuscript. RZZ and SJ produced the figures. All authors read
and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
|
1
|
Mitchell P: Coupling of phosphorylation to
electron and hydrogen transfer by a chemiosmotic type of mechanism.
Nature. 191:144–148. 1961. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guo R, Zong S, Wu M, Gu J and Yang M:
Architecture of human mitochondrial respiratory megacomplex
I2III2IV2. Cell. 170:1247–1257.e1212. 2017. View Article : Google Scholar
|
|
3
|
Iwata S, Lee JW, Okada K, Lee JK, Iwata M,
Rasmussen B, Link TA, Ramaswamy S and Jap BK: Complete structure of
the 11-subunit bovine mitochondrial cytochrome bc1 complex.
Science. 281:64–71. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sazanov LA and Hinchliffe P: Structure of
the hydrophilic domain of respiratory complex I from thermus
thermophilus. Science. 311:1430–1436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Efremov RG and Sazanov LA: Structure of
the membrane domain of respiratory complex I. Nature. 476:414–420.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jones AJ, Blaza JN, Varghese F and Hirst
J: Respiratory complex I in Bos taurus and paracoccus denitrificans
pumps four protons across the membrane for every NADH Oxidized. J
Biol Chem. 292:4987–4995. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carroll J, Fearnley IM, Skehel JM, Shannon
RJ, Hirst J and Walker JE: Bovine complex I is a complex of 45
different subunits. J Biol Chem. 281:32724–32727. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vinothkumar KR, Zhu J and Hirst J:
Architecture of mammalian respiratory complex I. Nature. 515:80–84.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ohnishi ST, Shinzawa-Itoh K, Ohta K,
Yoshikawa S and Ohnishi T: New insights into the superoxide
generation sites in bovine heart NADH-ubiquinone oxidoreductase
(Complex I): The significance of protein-associated ubiquinone and
the dynamic shifting of generation sites between semiflavin and
semiquinone radicals. Biochim Biophys Acta. 1797:1901–1909. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gai Z, Matsuno A, Kato K, Kato S, Khan
MRI, Shimizu T, Yoshioka T, Kato Y, Kishimura H, Kanno G, et al:
Crystal structure of the 3.8-MDa respiratory supermolecule
hemocyanin at 3.0 A resolution. Structure. 23:2204–2212. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hunte C, Zickermann V and Brandt U:
Functional modules and structural basis of conformational coupling
in mitochondrial complex I. Science. 329:448–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Formosa LE, Dibley MG, Stroud DA and Ryan
MT: Building a complex complex: Assembly of mitochondrial
respiratory chain complex I. Semin Cell Dev Biol. 76:154–162. 2018.
View Article : Google Scholar
|
|
13
|
Berrisford JM and Sazanov LA: Structural
basis for the mechanism of respiratory complex I. J Biol Chem.
284:29773–29783. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tan P, Feng Z, Zhang L, Hou T and Li Y:
The mechanism of proton translocation in respiratory complex I from
molecular dynamics. J Recept Signal Transduct Res. 35:170–179.
2015. View Article : Google Scholar
|
|
15
|
Wikstrom M and Hummer G: Stoichiometry of
proton translocation by respiratory complex I and its mechanistic
implications. Proc Natl Acad Sci USA. 109:4431–4436. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chance B and Williams GR: Respiratory
enzymes in oxidative phosphorylation. IV. The respiratory chain. J
Biol Chem. 217:429–438. 1955.PubMed/NCBI
|
|
17
|
Stoner CD: Determination of the
P/2e-stoichiometries at the individual coupling sites in
mitochondrial oxidative phosphorylation. Evidence for maximum
values of 1.0, 0.5, and 1.0 at sites 1, 2, and 3. J Biol Chem.
262:10445–10453. 1987.PubMed/NCBI
|
|
18
|
Ohnishi T: Structural biology: Piston
drives a proton pump. Nature. 465:428–429. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cecchini G: Function and structure of
complex II of the respiratory chain. Annu Rev Biochem. 72:77–109.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D,
Bartlam M and Rao Z: Crystal structure of mitochondrial respiratory
membrane protein complex II. Cell. 121:1043–1057. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bezawork-Geleta A, Rohlena J, Dong L,
Pacak K and Neuzil J: Mitochondrial complex II: At the crossroads.
Trends Biochem Sci. 42:312–325. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iverson TM: Catalytic mechanisms of
complex II enzymes: A structural perspective. Biochim Biophys Acta.
1827:648–657. 2013. View Article : Google Scholar
|
|
23
|
Schagger H, Link TA, Engel WD and von
Jagow G: Isolation of the eleven protein subunits of the bc1
complex from beef heart. Methods Enzymol. 126:224–237. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang XH and Trumpower BL: Purification of
a three-subunit ubiquinol-cytochrome c oxidoreductase complex from
paracoccus denitrificans. J Biol Chem. 261:12282–12289.
1986.PubMed/NCBI
|
|
25
|
Gao X, Wen X, Esser L, Quinn B, Yu L, Yu
CA and Xia D: Structural basis for the quinone reduction in the bc1
complex: A comparative analysis of crystal structures of
mitochondrial cytochrome bc1 with bound substrate and inhibitors at
the Qi site. Biochemistry. 42:9067–9080. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mitchell P: Chemiosmotic coupling in
energy transduction: A logical development of biochemical
knowledge. J Bioenerg. 3:5–24. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mitchell P: Possible molecular mechanisms
of the protonmotive function of cytochrome systems. J Theor Biol.
62:327–367. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Trumpower BL: A concerted, alternating
sites mechanism of ubiquinol oxidation by the dimeric cytochrome
bc(1) complex. Biochim Biophys Acta. 1555:166–173. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kadenbach B and Hüttemann M: The subunit
composition and function of mammalian cytochrome c oxidase.
Mitochondrion. 24:64–76. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tsukihara T, Aoyama H, Yamashita E,
Tomizaki T, Yamaguchi H, Shinzawa-Itoh K, Nakashima R, Yaono R and
Yoshikawa S: The whole structure of the 13-subunit oxidized
cytochrome c oxidase at 2.8 A. Science. 272:1136–1144. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Konstantinov AA: Cytochrome c oxidase:
Intermediates of the catalytic cycle and their energy-coupled
interconversion. FEBS Lett. 586:630–639. 2012. View Article : Google Scholar
|
|
32
|
Sharma V and Wikstrom M: The role of the
K-channel and the active-site tyrosine in the catalytic mechanism
of cytochrome c oxidase. Biochim Biophys Acta. 1857:1111–1115.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Varanasi L and Hosler JP: Subunit
III-depleted cytochrome c oxidase provides insight into the process
of proton uptake by proteins. Biochim Biophys Acta. 1817:545–551.
2012. View Article : Google Scholar :
|
|
34
|
Alnajjar KS, Hosler J and Prochaska L:
Role of the N-terminus of subunit III in proton uptake in
cytochrome c oxidase of Rhodobacter sphaeroides. Biochemistry.
53:496–504. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Arnold S and Kadenbach B: Cell respiration
is controlled by ATP, an allosteric inhibitor of cytochrome-c
oxidase. Eur J Biochem. 249:350–354. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Arnold S and Kadenbach B: The
intramitochondrial ATP/ADP-ratio controls cytochrome c oxidase
activity allosteri-cally. FEBS Lett. 443:105–108. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arnold S, Goglia F and Kadenbach B:
3,5-Diiodothyronine binds to subunit Va of cytochrome-c oxidase and
abolishes the allosteric inhibition of respiration by ATP. Eur J
Biochem. 252:325–330. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Follmann K, Arnold S, Ferguson-Miller S
and Kadenbach B: Cytochrome c oxidase from eucaryotes but not from
procaryotes is allosterically inhibited by ATP. Biochem Mol Biol
Int. 45:1047–1055. 1998.PubMed/NCBI
|
|
39
|
Shimada S, Shinzawa-Itoh K, Baba J, Aoe S,
Shimada A, Yamashita E, Kang J, Tateno M, Yoshikawa S and Tsukihara
T: Complex structure of cytochrome c-cytochrome c oxidase reveals a
novel protein-protein interaction mode. EMBO J. 36:291–300. 2017.
View Article : Google Scholar
|
|
40
|
Wikstrom MK: Proton pump coupled to
cytochrome c oxidase in mitochondria. Nature. 266:271–273. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jonckheere AI, Smeitink JA and Rodenburg
RJ: Mitochondrial ATP synthase: Architecture, function and
pathology. J Inherit Metab Dis. 35:211–225. 2012. View Article : Google Scholar :
|
|
42
|
Dickson VK, Silvester JA, Fearnley IM,
Leslie AG and Walker JE: On the structure of the stator of the
mitochondrial ATP synthase. EMBO J. 25:2911–2918. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Watt IN, Montgomery MG, Runswick MJ,
Leslie AG and Walker JE: Bioenergetic cost of making an adenosine
triphosphate molecule in animal mitochondria. Proc Natl Acad Sci
USA. 107:16823–16827. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pecina P, Nůsková H, Karbanová V,
Kaplanová V, Mráček T and Houštěk J: Role of the mitochondrial ATP
synthase central stalk subunits γ and δ in the activity and
assembly of the mammalian enzyme. Biochim Biophys Acta Bioenerg.
1859:374–381. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Guo R, Gu J, Zong S, Wu M and Yang M:
Structure and mechanism of mitochondrial electron transport chain.
Biomed J. 41:9–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fiedorczuk K, Letts JA, Degliesposti G,
Kaszuba K, Skehel M and Sazanov LA: Atomic structure of the entire
mammalian mitochondrial complex I. Nature. 538:406–410. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hahn A, Parey K, Bublitz M, Mills DJ,
Zickermann V, Vonck J, Kühlbrandt W and Meier T: Structure of a
complete ATP synthase dimer reveals the molecular basis of inner
mitochondrial membrane morphology. Mol Cell. 63:445–456. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Turrens JF: Mitochondrial formation of
reactive oxygen species. J Physiol. 552:335–344. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cadenas E and Davies KJ: Mitochondrial
free radical generation, oxidative stress, and aging. Free Radic
Biol Med. 29:222–230. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Brand MD: Mitochondrial generation of
superoxide and hydrogen peroxide as the source of mitochondrial
redox signaling. Free Radic Biol Med. 100:14–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kowaltowski AJ, de Souza-Pinto NC,
Castilho RF and Vercesi AE: Mitochondria and reactive oxygen
species. Free Radic Biol Med. 47:333–343. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Brand MD: The sites and topology of
mitochondrial superoxide production. Exp Gerontol. 45:466–472.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hernansanz-Agustin P, Ramos E, Navarro E,
Parada E, Sánchez-López N, Peláez-Aguado L, Cabrera-García JD,
Tello D, Buendia I, Marina A, et al: Mitochondrial complex I
deactivation is related to superoxide production in acute hypoxia.
Redox Biol. 12:1040–1051. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hoekstra AS and Bayley JP: The role of
complex II in disease. Biochim Biophys Acta. 1827:543–551. 2013.
View Article : Google Scholar
|
|
55
|
Cecchini G: Respiratory complex II: Role
in cellular physiology and disease. Biochim Biophys Acta.
1827:541–542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Quinlan CL, Orr AL, Perevoshchikova IV,
Treberg JR, Ackrell BA and Brand MD: Mitochondrial complex II can
generate reactive oxygen species at high rates in both the forward
and reverse reactions. J Biol Chem. 287:27255–27264. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ackrell BA, Kearney EB and Singer TP:
Mammalian succinate dehydrogenase. Methods Enzymol. 53:466–483.
1978. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Turrens JF, Alexandre A and Lehninger AL:
Ubisemiquinone is the electron donor for superoxide formation by
complex III of heart mitochondria. Arch Biochem Biophys.
237:408–414. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Richter C, Gogvadze V, Laffranchi R,
Schlapbach R, Schweizer M, Suter M, Walter P and Yaffee M: Oxidants
in mitochondria: From physiology to diseases. Biochim Biophys Acta.
1271:67–74. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Muller FL, Liu Y and Van Remmen H: Complex
III releases superoxide to both sides of the inner mitochondrial
membrane. J Biol Chem. 279:49064–49073. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
D'Autreaux B and Toledano MB: ROS as
signalling molecules: Mechanisms that generate specificity in ROS
homeostasis. Nat Rev Mol Cell Biol. 8:813–824. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bedard K and Krause KH: The NOX family of
ROS-generating NADPH oxidases: Physiology and pathophysiology.
Physiol Rev. 87:245–313. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Treberg JR, Quinlan CL and Brand MD:
Hydrogen peroxide efflux from muscle mitochondria underestimates
matrix superoxide production-a correction using glutathione
depletion. FEBS J. 277:2766–2778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Quinlan CL, Gerencser AA, Treberg JR and
Brand MD: The mechanism of superoxide production by the
antimycin-inhibited mitochondrial Q-cycle. J Biol Chem.
286:31361–31372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Erecinska M and Wilson DF: The effect of
antimycin A on cytochromes b561, b566, and their relationship to
ubiquinone and the iron-sulfer centers S-1 (+N-2) and S-3. Arch
Biochem Biophys. 174:143–157. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Orr AL, Vargas L, Turk CN, Baaten JE,
Matzen JT, Dardov VJ, Attle SJ, Li J, Quackenbush DC, Goncalves RL,
et al: Suppressors of superoxide production from mitochondrial
complex III. Nat Chem Biol. 11:834–836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Muramoto K, Ohta K, Shinzawa-Itoh K, Kanda
K, Taniguchi M, Nabekura H, Yamashita E, Tsukihara T and Yoshikawa
S: Bovine cytochrome c oxidase structures enable O2 reduction with
minimization of reactive oxygens and provide a proton-pumping gate.
Proc Natl Acad Sci USA. 107:7740–7745. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
De Giusti VC, Caldiz CI, Ennis IL, Perez
NG, Cingolani HE and Aiello EA: Mitochondrial reactive oxygen
species (ROS) as signaling molecules of intracellular pathways
triggered by the cardiac renin-angiotensin II-aldosterone system
(RAAS). Front Physiol. 4:1262013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Diebold L and Chandel NS: Mitochondrial
ROS regulation of proliferating cells. Free Radic Biol Med.
100:86–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kaminskyy VO and Zhivotovsky B: Free
radicals in cross talk between autophagy and apoptosis. Antioxid
Redox Signal. 21:86–102. 2014. View Article : Google Scholar
|
|
71
|
Emerling BM, Weinberg F, Snyder C, Burgess
Z, Mutlu GM, Viollet B, Budinger GR and Chandel NS: Hypoxic
activation of AMPK is dependent on mitochondrial ROS but
independent of an increase in AMP/ATP ratio. Free Radic Biol Med.
46:1386–1391. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Betzen C, White R, Zehendner CM,
Pietrowski E, Bender B, Luhmann HJ and Kuhlmann CR: Oxidative
stress upregulates the NMDA receptor on cerebrovascular
endothelium. Free Radic Biol Med. 47:1212–1220. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Huddleston AT, Tang W, Takeshima H,
Hamilton SL and Klann E: Superoxide-induced potentiation in the
hippocampus requires activation of ryanodine receptor type 3 and
ERK. J Neurophysiol. 99:1565–1571. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Hidalgo C and Arias-Cavieres A: Calcium,
reactive oxygen species, and synaptic plasticity. Physiology
(Bethesda). 31:201–215. 2016.
|
|
75
|
Shetty PK, Huang FL and Huang KP:
Ischemia-elicited oxidative modulation of
Ca2+/calmodulin-dependent protein kinase II. J Biol
Chem. 283:5389–5401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kemmerling U, Munoz P, Muller M, Sánchez
G, Aylwin ML, Klann E, Carrasco MA and Hidalgo C: Calcium release
by ryanodine receptors mediates hydrogen peroxide-induced
activation of ERK and CREB phosphorylation in N2a cells and
hippocampal neurons. Cell Calcium. 41:491–502. 2007. View Article : Google Scholar
|
|
77
|
Massaad CA and Klann E: Reactive oxygen
species in the regulation of synaptic plasticity and memory.
Antioxid Redox Signal. 14:2013–2054. 2011. View Article : Google Scholar :
|
|
78
|
Beckhauser TF, Francis-Oliveira J and De
Pasquale R: Reactive oxygen species: Physiological and
physiopathological effects on synaptic plasticity. J Exp Neurosci.
10(Suppl 1): S23–S48. 2016.
|
|
79
|
Gasperini RJ, Pavez M, Thompson AC,
Mitchell CB, Hardy H, Young KM, Chilton JK and Foa L: How does
calcium interact with the cytoskeleton to regulate growth cone
motility during axon pathfinding? Mol Cell Neurosci. 84:29–35.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Oswald MCW, Garnham N, Sweeney ST and
Landgraf M: Regulation of neuronal development and function by ROS.
FEBS Lett. 592:679–691. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hongpaisan J, Winters CA and Andrews SB:
Calcium-dependent mitochondrial superoxide modulates nuclear CREB
phosphorylation in hippocampal neurons. Mol Cell Neurosci.
24:1103–1115. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Orrenius S, Gogvadze V and Zhivotovsky B:
Mitochondrial oxidative stress: Implications for cell death. Annu
Rev Pharmacol Toxicol. 47:143–183. 2007. View Article : Google Scholar
|
|
83
|
Cingolani HE, Perez NG, Aiello EA, Ennis
IL, Garciarena CD, Villa-Abrille MC, Dulce RA, Caldiz CI, Yeves AM,
Correa MV, et al: Early signals after stretch leading to cardiac
hypertrophy. Key role of NHE-1. Front Biosci. 13:7096–7114. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Palomeque J, Rueda OV, Sapia L, Valverde
CA, Salas M, Petroff MV and Mattiazzi A: Angiotensin II-induced
oxidative stress resets the Ca2+ dependence of
Ca2+-calmodulin protein kinase II and promotes a death
pathway conserved across different species. Circ Res.
105:1204–1212. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang H and Patterson C: Roles of reactive
oxygen species in physiology and pathology. John Wiley & Sons,
Inc; Hoboken, NJ: pp. 379–392. 2015
|
|
86
|
Robin E, Guzy RD, Loor G, Iwase H, Waypa
GB, Marks JD, Hoek TL and Schumacker PT: Oxidant stress during
simulated ischemia primes cardiomyocytes for cell death during
reperfusion. J Biol Chem. 282:19133–19143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang L, Azad N, Kongkaneramit L, Chen F,
Lu Y, Jiang BH and Rojanasakul Y: The Fas death signaling pathway
connecting reactive oxygen species generation and FLICE inhibitory
protein down-regulation. J Immunol. 180:3072–3080. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Deng Y, Ren X, Yang L, Lin Y and Wu X: A
JNK-dependent pathway is required for TNFalpha-induced apoptosis.
Cell. 115:61–70. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Hattori K, Naguro I, Runchel C and Ichijo
H: The roles of ASK family proteins in stress responses and
diseases. Cell Commun Signal. 7:92009. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Sinha K, Das J, Pal PB and Sil PC:
Oxidative stress: The mitochondria-dependent and
mitochondria-independent pathways of apoptosis. Arch Toxicol.
87:1157–1180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Shen C, Cai GQ, Peng JP and Chen XD:
Autophagy protects chondrocytes from glucocorticoids-induced
apoptosis via ROS/Akt/FOXO3 signaling. Osteoarthritis Cartilage.
23:2279–2287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Shah SZA, Zhao D, Hussain T, Sabir N,
Mangi MH and Yang L: p62-Keap1-NRF2-ARE pathway: A contentious
player for selective targeting of autophagy, oxidative stress and
mitochondrial dysfunction in prion diseases. Front Mol Neurosci.
11:3102018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wu H, Huang S, Chen Z, Liu W, Zhou X and
Zhang D: Hypoxia-induced autophagy contributes to the invasion of
salivary adenoid cystic carcinoma through the HIF-1α/BNIP3
signaling pathway. Mol Med Rep. 12:6467–6474. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Bensaad K, Cheung EC and Vousden KH:
Modulation of intracellular ROS levels by TIGAR controls autophagy.
EMBO J. 28:3015–3026. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Gurusamy N and Das DK: Autophagy, redox
signaling, and ventricular remodeling. Antioxid Redox Signal.
11:1975–1988. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Li R, Zhou P, Guo Y, Lee JS and Zhou B:
Tris (1, 3-dichloro-2-propyl) phosphate induces apoptosis and
autophagy in SH-SY5Y cells: Involvement of ROS-mediated
AMPK/mTOR/ULK1 pathways. Food Chem Toxicol. 100:183–196. 2017.
View Article : Google Scholar
|
|
98
|
Vanden Berghe T, Linkermann A,
Jouan-Lanhouet S, Walczak H and Vandenabeele P: Regulated necrosis:
The expanding network of non-apoptotic cell death pathways. Nat Rev
Mol Cell Biol. 15:135–147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Schenk B and Fulda S: Reactive oxygen
species regulate Smac mimetic/TNFα-induced necroptotic signaling
and cell death. Oncogene. 34:5796–5806. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Morgan MJ and Liu ZG: Reactive oxygen
species in TNFalpha-induced signaling and cell death. Mol Cells.
30:1–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ying Y, Kim J, Westphal SN, Long KE and
Padanilam BJ: Targeted deletion of p53 in the proximal tubule
prevents ischemic renal injury. J Am Soc Nephrol. 25:2707–2716.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Heid ME, Keyel PA, Kamga C, Shiva S,
Watkins SC and Salter RD: Mitochondrial reactive oxygen species
induces NLRP3-dependent lysosomal damage and inflammasome
activation. J Immunol. 191:5230–5238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zhou R, Tardivel A, Thorens B, Choi I and
Tschopp J: Thioredoxin-interacting protein links oxidative stress
to inflammasome activation. Nat Immunol. 11:136–140. 2010.
View Article : Google Scholar
|
|
104
|
Scholl FA, Dumesic PA, Barragan DI, Harada
K, Bissonauth V, Charron J and Khavari PA: Mek1/2 MAPK kinases are
essential for mammalian development, homeostasis, and raf-induced
hyperplasia. Dev Cell. 12:615–629. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Hulsmans M, Van Dooren E and Holvoet P:
Mitochondrial reactive oxygen species and risk of atherosclerosis.
Curr Atheroscler Rep. 14:264–276. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Kattoor AJ, Pothineni NVK, Palagiri D and
Mehta JL: Oxidative stress in atherosclerosis. Curr Atheroscler
Rep. 19:422017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Montezano AC, Dulak-Lis M, Tsiropoulou S,
Harvey A, Briones AM and Touyz RM: Oxidative stress and human
hypertension: Vascular mechanisms, biomarkers, and novel therapies.
Can J Cardiol. 31:631–641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Sanderson TH, Reynolds CA, Kumar R,
Przyklenk K and Huttemann M: Molecular mechanisms of
ischemia-reperfusion injury in brain: pivotal role of the
mitochondrial membrane potential in reactive oxygen species
generation. Mol Neurobiol. 47:9–23. 2013. View Article : Google Scholar :
|
|
109
|
Chouchani ET, Pell VR, James AM, Work LM,
Saeb-Parsy K, Frezza C, Krieg T and Murphy MP: A unifying mechanism
for mitochondrial superoxide production during ischemia-reperfusion
injury. Cell Metab. 23:254–263. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lorenzo O, Ramirez E, Picatoste B, Egido J
and Tunon J: Alteration of energy substrates and ROS production in
diabetic cardiomyopathy. Mediators Inflamm. 2013:4619672013.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zhao MX, Zhou B, Ling L, Xiong XQ, Zhang
F, Chen Q, Li YH, Kang YM and Zhu GQ: Salusin-beta contributes to
oxidative stress and inflammation in diabetic cardiomyopathy. Cell
Death Dis. 8:e26902017. View Article : Google Scholar
|
|
112
|
Jaitovich A and Jourd'Heuil D: A brief
overview of nitric oxide and reactive oxygen species signaling in
hypoxia-induced pulmonary hypertension. Adv Exp Med Biol.
967:71–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Fulton DJR, Li X, Bordan Z, Haigh S,
Bentley A, Chen F and Barman SA: Reactive oxygen and nitrogen
species in the development of pulmonary hypertension. Antioxidants
(Basel). 6:E542017. View Article : Google Scholar
|
|
114
|
Allaire J, Maltais F, LeBlanc P, Simard
PM, Whittom F, Doyon JF, Simard C and Jobin J: Lipofuscin
accumulation in the vastus lateralis muscle in patients with
chronic obstructive pulmonary disease. Muscle Nerve. 25:383–389.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Dekhuijzen PN, Aben KK, Dekker I, Aarts
LP, Wielders PL, van Herwaarden CL and Bast A: Increased exhalation
of hydrogen peroxide in patients with stable and unstable chronic
obstructive pulmonary disease. Am J Respir Crit Care Med.
154:813–816. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Montuschi P, Collins JV, Ciabattoni G,
Lazzeri N, Corradi M, Kharitonov SA and Barnes PJ: Exhaled
8-isoprostane as an in vivo biomarker of lung oxidative stress in
patients with COPD and healthy smokers. Am J Respir Crit Care Med.
162:1175–1177. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Sabharwal SS and Schumacker PT:
Mitochondrial ROS in cancer: Initiators, amplifiers or an achilles'
heel? Nat Rev Cancer. 14:709–721. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Sosa V, Moline T, Somoza R, Paciucci R,
Kondoh H and Lleonart ME: Oxidative stress and cancer: An overview.
Ageing Res Rev. 12:376–390. 2013. View Article : Google Scholar
|
|
119
|
Ahmad W, Ijaz B, Shabbiri K, Ahmed F and
Rehman S: Oxidative toxicity in diabetes and Alzheimer's disease:
Mechanisms behind ROS/RNS generation. J Biomed Sci. 24:762017.
View Article : Google Scholar
|
|
120
|
Rehman K and Akash MSH: Mechanism of
generation of oxidative stress and pathophysiology of type 2
diabetes mellitus: How are they interlinked? J Cell Biochem.
118:3577–3585. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Spahis S, Delvin E, Borys JM and Levy E:
Oxidative stress as a critical factor in nonalcoholic fatty liver
disease pathogenesis. Antioxid Redox Signal. 26:519–541. 2017.
View Article : Google Scholar
|
|
122
|
Rolo AP, Teodoro JS and Palmeira CM: Role
of oxidative stress in the pathogenesis of nonalcoholic
steatohepatitis. Free Radic Biol Med. 52:59–69. 2012. View Article : Google Scholar
|
|
123
|
de la Monte SM and Wands JR: Molecular
indices of oxidative stress and mitochondrial dysfunction occur
early and oftenprogresswith severity of Alzheimer's disease. J
Alzheimers Dis. 9:167–181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Dias V, Junn E and Mouradian MM: The role
of oxidative stress in Parkinson's disease. J Parkinsons Dis.
3:461–491. 2013.PubMed/NCBI
|
|
125
|
Nagano T, Mizuno M, Morita K and Nawa H:
Pathological implications of oxidative stress in patients and
animal models with schizophrenia: The role of epidermal growth
factor receptor signaling. Curr Top Behav Neurosci. 29:429–446.
2016. View Article : Google Scholar
|
|
126
|
Mahadik SP, Pillai A, Joshi S and Foster
A: Prevention of oxidative stress-mediated neuropathology and
improved clinical outcome by adjunctive use of a combination of
antioxidants and omega-3 fatty acids in schizophrenia. Int Rev
Psychiatry. 18:119–131. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Kwon DN, Park WJ, Choi YJ, Gurunathan S
and Kim JH: Oxidative stress and ROS metabolism via down-regulation
of sirtuin 3 expression in Cmahnull mice affect hearing loss. Aging
(Albany NY). 7:579–594. 2015. View Article : Google Scholar
|
|
128
|
Kamogashira T, Fujimoto C and Yamasoba T:
Reactive oxygen species, apoptosis, and mitochondrial dysfunction
in hearing loss. Biomed Res Int. 2015:6172072015. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Marazita MC, Dugour A, Marquioni-Ramella
MD, Figueroa JM and Suburo AM: Oxidative stress-induced premature
senescence dysregulates VEGF and CFH expression in retinal pigment
epithelial cells: Implications for age-related macular
degeneration. Redox Biol. 7:78–87. 2016. View Article : Google Scholar :
|
|
130
|
Nita M and Grzybowski A: The role of the
reactive oxygen species and oxidative stress in the pathomechanism
of the age-related ocular diseases and other pathologies of the
anterior and posterior eye segments in adults. Oxid Med Cell
Longev. 2016:31647342016. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Ghosh S, Sulistyoningrum DC, Glier MB,
Verchere CB and Devlin AM: Altered glutathione homeostasis in heart
augments cardiac lipotoxicity associated with diet-induced obesity
in mice. J Biol Chem. 286:42483–42493. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Mandas A, Iorio EL, Congiu MG, Balestrieri
C, Mereu A, Cau D, Dessì S and Curreli N: Oxidative imbalance in
HIV-1 infected patients treated with antiretroviral therapy. J
Biomed Biotechnol. 2009:7495752009. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Shin DH, Martinez SS, Parsons M, Jayaweera
DT, Campa A and Baum MK: Relationship of oxidative stress with HIV
disease progression in HIV/HCV Co-infected and HIV mono-infected
adults in miami. Int J Biosci Biochem Bioinforma. 2:217–223.
2012.PubMed/NCBI
|
|
134
|
Haycock JW, MacNeil S, Jones P, Harris JB
and Mantle D: Oxidative damage to muscle protein in Duchenne
muscular dystrophy. Neuroreport. 8:357–361. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Austin L, de Niese M, McGregor A, Arthur
H, Gurusinghe A and Gould MK: Potential oxyradical damage and
energy status in individual muscle fibres from degenerating muscle
diseases. Neuromuscul Disord. 2:27–33. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Lee G, Kim HJ and Kim HM: RhoA-JNK
regulates the E-cadherin junctions of human gingival epithelial
cells. J Dent Res. 95:284–291. 2016. View Article : Google Scholar
|
|
137
|
Nobes CD, Brown GC, Olive PN and Brand MD:
Nonohmic proton conductance of the mitochondrial inner membrane in
hepatocytes. J Biol Chem. 265:12903–12909. 1990.PubMed/NCBI
|
|
138
|
Brand MD, Turner N, Ocloo A, Else PL and
Hulbert AJ: Proton conductance and fatty acyl composition of liver
mitochondria correlates with body mass in birds. Biochem J.
376:741–748. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Porter RK and Brand MD: Body mass
dependence of H+ leak in mitochondria and its relevance to
metabolic rate. Nature. 362:628–630. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Brookes PS, Rolfe DF and Brand MD: The
proton permeability of liposomes made from mitochondrial inner
membrane phospholipids: Comparison with isolated mitochondria. J
Membr Biol. 155:167–174. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Brand MD, Pakay JL, Ocloo A, Kokoszka J,
Wallace DC, Brookes PS and Cornwall EJ: The basal proton
conductance of mitochondria depends on adenine nucleotide
translocase content. Biochem J. 392:353–362. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Parker N, Crichton PG, Vidal-Puig AJ and
Brand MD: Uncoupling protein-1 (UCP1) contributes to the basal
proton conductance of brown adipose tissue mitochondria. J Bioenerg
Biomembr. 41:335–342. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Shabalina IG, Ost M, Petrovic N, Vrbacky
M, Nedergaard J and Cannon B: Uncoupling protein-1 is not leaky.
Biochim Biophys Acta. 1797:773–784. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Roussel D, Harding M, Runswick MJ, Walker
JE and Brand MD: Does any yeast mitochondrial carrier have a native
uncoupling protein function? J Bioenerg Biomembr. 34:165–176. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Krauss S, Zhang CY and Lowell BB: The
mitochondrial uncoupling-protein homologues. Nat Rev Mol Cell Biol.
6:248–261. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Huang SG and Klingenberg M: Fluorescent
nucleotide derivatives as specific probes for the uncoupling
protein: Thermodynamics and kinetics of binding and the control by
pH. Biochemistry. 34:349–360. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Xia C, Liu JZ and Xu Y: Effects of GDP on
the activity and expression of mitochondrial uncoupling proteins in
rat brain in vitro. Sheng Li Xue Bao. 60:492–496. 2008.In Chinese.
PubMed/NCBI
|
|
148
|
Ramsden DB, Ho PW, Ho JW, Liu HF, So DH,
Tse HM, Chan KH and Ho SL: Human neuronal uncoupling proteins 4 and
5 (UCP4 and UCP5): Structural properties, regulation, and
physiological role in protection against oxidative stress and
mitochondrial dysfunction. Brain Behav. 2:468–478. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Hoang T, Smith MD and Jelokhani-Niaraki M:
Toward understanding the mechanism of ion transport activity of
neuronal uncoupling proteins UCP2, UCP4, and UCP5. Biochemistry.
51:4004–4014. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Andrews ZB, Diano S and Horvath TL:
Mitochondrial uncoupling proteins in the cns: In support of
function and survival. Nat Rev Neurosci. 6:829–840. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Nicholls DG and Locke RM: Thermogenic
mechanisms in brown fat. Physiol Rev. 64:1–64. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Shabalina IG, Petrovic N, de Jong JM,
Kalinovich AV, Cannon B and Nedergaard J: UCP1 in brite/beige
adipose tissue mitochondria is functionally thermogenic. Cell Rep.
5:1196–1203. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Golozoubova V, Hohtola E, Matthias A,
Jacobsson A, Cannon B and Nedergaard J: Only UCP1 can mediate
adaptive nonshivering thermogenesis in the cold. FASEB J.
15:2048–2050. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Enerback S, Jacobsson A, Simpson EM,
Guerra C, Yamashita H, Harper ME and Kozak LP: Mice lacking
mitochondrial uncoupling protein are cold-sensitive but not obese.
Nature. 387:90–94. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Divakaruni AS and Brand MD: The regulation
and physiology of mitochondrial proton leak. Physiology (Bethesda).
26:192–205. 2011.
|
|
156
|
Granneman JG, Burnazi M, Zhu Z and Schwamb
LA: White adipose tissue contributes to UCP1-independent
thermogenesis. Am J Physiol Endocrinol Metab. 285:E1230–E1236.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Adams AE, Kelly OM and Porter RK: Absence
of mitochondrial uncoupling protein 1 affects apoptosis in
thymocytes, thymocyte/T-cell profile and peripheral T-cell number.
Biochim Biophys Acta. 1797:807–816. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Adams AE, Hanrahan O, Nolan DN, Voorheis
HP, Fallon P and Porter RK: Images of mitochondrial UCP 1 in mouse
thymocytes using confocal microscopy. Biochim Biophys Acta.
1777:115–117. 2008. View Article : Google Scholar
|
|
159
|
Adams AE, Carroll AM, Fallon PG and Porter
RK: Mitochondrial uncoupling protein 1 expression in thymocytes.
Biochim Biophys Acta. 1777:772–776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Sale MM, Hsu FC, Palmer ND, Gordon CJ,
Keene KL, Borgerink HM, Sharma AJ, Bergman RN, Taylor KD, Saad MF
and Norris JM: The uncoupling protein 1 gene, UCP1, is expressed in
mammalian islet cells and associated with acute insulin response to
glucose in african american families from the IRAS family study.
BMC Endocr Disord. 7:12007. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Shabalina IG, Jacobsson A, Cannon B and
Nedergaard J: Native UCP1 displays simple competitive kinetics
between the regulators purine nucleotides and fatty acids. J Biol
Chem. 279:38236–38248. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Breen EP, Gouin SG, Murphy AF, Haines LR,
Jackson AM, Pearson TW, Murphy PV and Porter RK: On the mechanism
of mitochondrial uncoupling protein 1 function. J Biol Chem.
281:2114–2119. 2006. View Article : Google Scholar
|
|
163
|
Rial E, Aguirregoitia E, Jimenez-Jimenez J
and Ledesma A: Alkylsulfonates activate the uncoupling protein
UCP1: Implications for the transport mechanism. Biochim Biophys
Acta. 1608:122–130. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Klingenberg M and Huang SG: Structure and
function of the uncoupling protein from brown adipose tissue.
Biochim Biophys Acta. 1415:271–296. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Garlid KD, Jaburek M and Jezek P: The
mechanism of proton transport mediated by mitochondrial uncoupling
proteins. FEBS Lett. 438:10–14. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Garlid KD, Orosz DE, Modriansky M,
Vassanelli S and Jezek P: On the mechanism of fatty acid-induced
proton transport by mitochondrial uncoupling protein. J Biol Chem.
271:2615–2620. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Winkler E and Klingenberg M: Effect of
fatty acids on H+ transport activity of the reconstituted
uncoupling protein. J Biol Chem. 269:2508–2515. 1994.PubMed/NCBI
|
|
168
|
Skulachev VP: Fatty acid circuit as a
physiological mechanism of uncoupling of oxidative phosphorylation.
FEBS Lett. 294:158–162. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Klingenberg M and Winkler E: The
reconstituted isolated uncoupling protein is a membrane potential
driven H+ translocator. EMBO J. 4:3087–3092. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Azzu V and Brand MD: The on-off switches
of the mitochondrial uncoupling proteins. Trends Biochem Sci.
35:298–307. 2010. View Article : Google Scholar
|
|
171
|
Ricquier D and Bouillaud F: The uncoupling
protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP. Biochem J.
345(Pt 2): 161–179. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Friederich M, Fasching A, Hansell P,
Nordquist L and Palm F: Diabetes-induced up-regulation of
uncoupling protein-2 results in increased mitochondrial uncoupling
in kidney proximal tubular cells. Biochim Biophys Acta.
1777:935–940. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Flachs P, Sponarova J, Kopecky P, Horvath
O, Sediva A, Nibbelink M, Casteilla L, Medrikova D, Neckar J, Kolar
F and Kopecky J: Mitochondrial uncoupling protein 2 gene transcript
levels are elevated in maturating erythroid cells. FEBS Lett.
581:1093–1097. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Affourtit C and Brand MD: On the role of
uncoupling protein-2 in pancreatic beta cells. Biochim Biophys
Acta. 1777:973–979. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Cadenas S: Mitochondrial uncoupling, ROS
generation and cardioprotection. Biochim Biophys Acta Bioenerg.
1859:940–950. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Teshima Y, Akao M, Jones SP and Marban E:
Uncoupling protein-2 overexpression inhibits mitochondrial death
pathway in cardiomyocytes. Circ Res. 93:192–200. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Vidal-Puig A, Solanes G, Grujic D, Flier
JS and Lowell BB: UCP3: An uncoupling protein homologue expressed
preferentially and abundantly in skeletal muscle and brown adipose
tissue. Biochem Biophys Res Commun. 235:79–82. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Aguirre E and Cadenas S: GDP and
carboxyatractylate inhibit 4-hydroxynonenal-activated proton
conductance to differing degrees in mitochondria from skeletal
muscle and heart. Biochim Biophys Acta. 1797:1716–1726. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Kelly OM and Porter RK: Absence of
mitochondrial uncoupling protein 3: Effect on thymus and spleen in
the fed and fasted mice. Biochim Biophys Acta. 1807:1064–1074.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Mori S, Yoshizuka N, Takizawa M, Takema Y,
Murase T, Tokimitsu I and Saito M: Expression of uncoupling
proteins in human skin and skin-derived cells. J Invest Dermatol.
128:1894–1900. 2008. View Article : Google Scholar
|
|
181
|
Harper ME and Himms-Hagen J: Mitochondrial
efficiency: Lessons learned from transgenic mice. Biochim Biophys
Acta. 1504:159–172. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Negre-Salvayre A, Hirtz C, Carrera G,
Cazenave R, Troly M, Salvayre R, Pénicaud L and Casteilla L: A role
for uncoupling protein-2 as a regulator of mitochondrial hydrogen
peroxide generation. FASEB J. 11:809–815. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Suski JM, Lebiedzinska M, Bonora M, Pinton
P, Duszynski J and Wieckowski MR: Relation between mitochondrial
membrane potential and ROS formation. Methods Mol Biol.
810:183–205. 2012. View Article : Google Scholar
|
|
184
|
Pi J, Bai Y, Daniel KW, Liu D, Lyght O,
Edelstein D, Brownlee M, Corkey BE and Collins S: Persistent
oxidative stress due to absence of uncoupling protein 2 associated
with impaired pancreatic beta-cell function. Endocrinology.
150:3040–3048. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Vidal-Puig AJ, Grujic D, Zhang CY, Hagen
T, Boss O, Ido Y, Szczepanik A, Wade J, Mootha V, Cortright R, et
al: Energy metabolism in uncoupling protein 3 gene knockout mice. J
Biol Chem. 275:16258–16266. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Brand MD, Pamplona R, Portero-Otin M,
Requena JR, Roebuck SJ, Buckingham JA, Clapham JC and Cadenas S:
Oxidative damage and phospholipid fatty acyl composition in
skeletal muscle mitochondria from mice underexpressing or
overexpressing uncoupling protein 3. Biochem J. 368:597–603. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Brand MD: Uncoupling to survive? The role
of mitochondrial inefficiency in ageing. Exp Gerontol. 35:811–820.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Dlaskova A, Clarke KJ and Porter RK: The
role of UCP 1 in production of reactive oxygen species by
mitochondria isolated from brown adipose tissue. Biochim Biophys
Acta. 1797:1470–1476. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Shabalina IG, Vrbacky M, Pecinova A,
Kalinovich AV, Drahota Z, Houštěk J, Mráček T, Cannon B and
Nedergaard J: ROS production in brown adipose tissue mitochondria:
The question of UCP1-dependence. Biochim Biophys Acta.
1837:2017–2030. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Skulachev VP: Anion carriers in fatty
acid-mediated physiological uncoupling. J Bioenerg Biomembr.
31:431–445. 1999. View Article : Google Scholar
|
|
191
|
Schrauwen P and Hesselink MK: The role of
uncoupling protein 3 in fatty acid metabolism: Protection against
lipotoxicity? Proc Nutr Soc. 63:287–292. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Bouillaud F, Couplan E, Pecqueur C and
Ricquier D: Homologues of the uncoupling protein from brown adipose
tissue (UCP1): UCP2, UCP3, BMCP1 and UCP4. Biochim Biophys Acta.
1504:107–119. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Mao W, Yu XX, Zhong A, Li W, Brush J,
Sherwood SW, Adams SH and Pan G: UCP4, a novel brain-specific
mitochondrial protein that reduces membrane potential in mammalian
cells. FEBS Lett. 443:326–330. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Sanchis D, Fleury C, Chomiki N, Goubern M,
Huang Q, Neverova M, Grégoire F, Easlick J, Raimbault S,
Lévi-Meyrueis C, et al: BMCP1, a novel mitochondrial carrier with
high expression in the central nervous system of humans and
rodents, and respiration uncoupling activity in recombinant yeast.
J Biol Chem. 273:34611–34615. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Zhang M, Wang B, Ni YH, Liu F, Fei L, Pan
XQ, Guo M, Chen RH and Guo XR: Overexpression of uncoupling protein
4 promotes proliferation and inhibits apoptosis and differentiation
of preadipocytes. Life Sci. 79:1428–1435. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Slocinska M, Barylski J and Jarmuszkiewicz
W: Uncoupling proteins of invertebrates: A review. IUBMB Life.
68:691–699. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Ivanova MV, Hoang T, McSorley FR, Krnac G,
Smith MD and Jelokhani-Niaraki M: A comparative study on
conformation and ligand binding of the neuronal uncoupling
proteins. Biochemistry. 49:512–521. 2010. View Article : Google Scholar
|
|
198
|
Kwok KH, Ho PW, Chu AC, Ho JW, Liu HF, Yiu
DC, Chan KH, Kung MH, Ramsden DB and Ho SL: Mitochondrial UCP5 is
neuroprotective by preserving mitochondrial membrane potential, ATP
levels, and reducing oxidative stress in MPP+ and dopamine
toxicity. Free Radic Biol Med. 49:1023–1035. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Emerit J, Edeas M and Bricaire F:
Neurodegenerative diseases and oxidative stress. Biomed
Pharmacother. 58:39–46. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Kim-Han JS and Dugan LL: Mitochondrial
uncoupling proteins in the central nervous system. Antioxid Redox
Signal. 7:1173–1181. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Ho PW, Ho JW, Tse HM, So DH, Yiu DC, Liu
HF, Chan KH, Kung MH, Ramsden DB and Ho SL: Uncoupling protein-4
(UCP4) increases ATP supply by interacting with mitochondrial
complex II in neuroblastoma cells. PLoS One. 7:e328102012.
View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Pfeiffer M, Kayzer EB, Yang X, Abramson E,
Kenaston MA, Lago CU, Lo HH, Sedensky MM, Lunceford A, Clarke CF,
et al: Caenorhabditis elegans UCP4 protein controls complex
II-mediated oxidative phosphorylation through succinate transport.
J Biol Chem. 286:37712–37720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Chan SL, Liu D, Kyriazis GA, Bagsiyao P,
Ouyang X and Mattson MP: Mitochondrial uncoupling protein-4
regulates calcium homeostasis and sensitivity to store
depletion-induced apoptosis in neural cells. J Biol Chem.
281:37391–37403. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Walder K, Norman RA, Hanson RL, Schrauwen
P, Neverova M, Jenkinson CP, Easlick J, Warden CH, Pecqueur C,
Raimbault S, et al: Association between uncoupling protein
polymorphisms (UCP2UCP3) and energy metabolism/obesity in pima
indians. Hum Mol Genet. 7:1431–1435. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Pheiffer C, Jacobs C, Patel O, Ghoor S,
Muller C and Louw J: Expression of UCP2 in wistar rats varies
according to age and the severity of obesity. J Physiol Biochem.
72:25–32. 2016. View Article : Google Scholar
|
|
206
|
Millet L, Vidal H, Andreelli F, Larrouy D,
Riou JP, Ricquier D, Laville M and Langin D: Increased uncoupling
protein-2 and -3 mRNA expression during fasting in obese and lean
humans. J Clin Invest. 100:2665–2670. 1997. View Article : Google Scholar
|
|
207
|
Robson-Doucette CA, Sultan S, Allister EM,
Wikstrom JD, Koshkin V, Bhattacharjee A, Prentice KJ, Sereda SB,
Shirihai OS and Wheeler MB: Beta-cell uncoupling protein 2
regulates reactive oxygen species production, which influences both
insulin and glucagon secretion. Diabetes. 60:2710–2719. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Gonzalez-Barroso MM, Giurgea I, Bouillaud
F, Anedda A, Bellanné-Chantelot C, Hubert L, de Keyzer Y, de Lonlay
P and Ricquier D: Mutations in UCP2 in congenital hyperinsulinism
reveal a role for regulation of insulin secretion. PLoS One.
3:e38502008. View Article : Google Scholar : PubMed/NCBI
|
|
209
|
Lameloise N, Muzzin P, Prentki M and
Assimacopoulos-Jeannet F: Uncoupling protein 2: A possible link
between fatty acid excess and impaired glucose-induced insulin
secretion? Diabetes. 50:803–809. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Patane G, Anello M, Piro S, Vigneri R,
Purrello F and Rabuazzo AM: Role of ATP production and uncoupling
protein-2 in the insulin secretory defect induced by chronic
exposure to high glucose or free fatty acids and effects of
peroxisome proliferator-activated receptor-gamma inhibition.
Diabetes. 51:2749–2756. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Li W, Nichols K, Nathan CA and Zhao Y:
Mitochondrial uncoupling protein 2 is up-regulated in human head
and neck, skin, pancreatic, and prostate tumors. Cancer Biomark.
13:377–383. 2013. View Article : Google Scholar
|
|
212
|
Derdak Z, Mark NM, Beldi G, Robson SC,
Wands JR and Baffy G: The mitochondrial uncoupling protein-2
promotes chemoresistance in cancer cells. Cancer Res. 68:2813–2819.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Pons DG, Nadal-Serrano M, Torrens-Mas M,
Valle A, Oliver J and Roca P: UCP2 inhibition sensitizes breast
cancer cells to therapeutic agents by increasing oxidative stress.
Free Radic Biol Med. 86:67–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Kawanishi M, Fukuda T, Shimomura M, Inoue
Y, Wada T, Tasaka R, Yasui T and Sumi T: Expression of UCP2 is
associated with sensitivity to platinum-based chemotherapy for
ovarian serous carcinoma. Oncol Lett. 15:9923–9928. 2018.PubMed/NCBI
|
|
215
|
Franssen FM, Wouters EF, Baarends EM,
Akkermans MA and Schols AM: Arm mechanical efficiency and arm
exercise capacity are relatively preserved in chronic obstructive
pulmonary disease. Med Sci Sports Exerc. 34:1570–1576. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Gosker HR, Schrauwen P, Broekhuizen R,
Hesselink MK, Moonen-Kornips E, Ward KA, Franssen FM, Wouters EF
and Schols AM: Exercise training restores uncoupling protein-3
content in limb muscles of patients with chronic obstructive
pulmonary disease. Am J Physiol. Endocrinol Metab. 290:E976–E981.
2006.
|
|
217
|
Patti ME and Corvera S: The role of
mitochondria in the pathogenesis of type 2 diabetes. Endocr Rev.
31:364–395. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Schrauwen P, Hesselink MK, Blaak EE,
Borghouts LB, Schaart G, Saris WH and Keizer HA: Uncoupling protein
3 content is decreased in skeletal muscle of patients with type 2
diabetes. Diabetes. 50:2870–2873. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Yasuno K, Ando S, Misumi S, Makino S,
Kulski JK, Muratake T, Kaneko N, Amagane H, Someya T, Inoko H, et
al: Synergistic association of mitochondrial uncoupling protein
(UCP) genes with schizophrenia. Am J Med Genet B Neuropsychiatr
Genet. 144B:250–253. 2007. View Article : Google Scholar
|
|
220
|
Gigante AD, Andreazza AC, Lafer B, Yatham
LN, Beasley CL and Young LT: Decreased mRNA expression of
uncoupling protein 2, a mitochondrial proton transporter, in
post-mortem prefrontal cortex from patients with bipolar disorder
and schizophrenia. Neurosci Lett. 505:47–51. 2011. View Article : Google Scholar : PubMed/NCBI
|