Cancer currently ranks as either the first or second
leading cause of mortality among people prior to the age of 70
years worldwide (1). Based on
global cancer statistics collected in 2020, the large burden of
cancer incidence and mortality is projected to grow, with a
predicted 22 million new cancer cases and 13 million cancer-related
deaths occurring annually by the year 2030 (2,3).
Cancer is increasing in prominence as an effort is being made to
increase the range of novel treatment approaches for cancer. The
conventional therapeutic methods for the treatment of cancer,
including chemotherapy and irradiation following conservative
surgery, are not always effective and are associated with
undesirable side-effects (4).
Hence, additional intensive strategies are required to improve the
chances of successful therapy.
The immune system in the tumor microenvironment is
considered to be a key target for cancer therapy (5,6).
Immune responses to tumor cells have been generally considered to
eliminate the vast majority of incipient cancer cells and
consequently, nascent tumors (5).
Cancer immunotherapy aims to boost the power of the host's immune
system to prevent, control and eliminate cancer (7). The use of tumor antigen-specific
T-cells in preclinical and clinical studies on multiple types of
cancer has the objective of inducing the activation of the
patient's immune system, directing it specifically toward
tumor-associated antigens (TAAs) (8). Due to their genetic instability,
TAAs are antigens that are aberrantly expressed to a greater degree
in tumors than in normal tissues (9). This feature contributes to the low
systemic toxicity if a TAA is targeted for treatment, and since
TAAs are common markers of cancer cells, they are attractive as
specific targets for a range of cancer types (7).
Clinical trials are currently being conducted on
TAAs to translate the discoveries of these new potential molecular
targets into clinical cancer treatments. Several good examples
include treatments for malignant pleural mesothelioma, pancreatic
and ovarian cancer that target mesothelin antigens (phase I,
ClinicalTrials.gov: NCT01355965 and ClinicalTrials.gov: NCT02159716), NKG2D-L antigen in
phase I/II and treatments for ovarian, colorectal, pancreatic,
bladder, triple-negative breast cancer (TNBC), acute myeloid
leukemia and multiple myeloma that target the NKG2D-L antigen
(phase I/II, ClinicalTrials.gov: NCT03018405). Several clinical
studies are using human epidermal growth factor receptor 2 (HER-2)
antigen in the treatment of sarcoma, including osteosarcomas, Ewing
sarcoma and soft tissue sarcomas, such as synovial sarcoma or
desmoplastic small round cell tumors, nonrhabdomyosarcoma and
rhabdomyosarcoma (ClinicalTrials.gov: NCT01935843), and in the treatment
of advanced sarcoma (ClinicalTrials.gov: NCT00902044), lung,
gastrointestinal tract, ovarian, biliary tract, pancreatic and
brain cancer (glioblastoma multiforme) (ClinicalTrials. gov:
NCT01109095 and ClinicalTrials.gov: NCT02349724). Moreover, the
treatment of New York esophageal squamous cell carcinoma 1
(NY-ESO-1; Wilms' tumor 1, WT1) is cited in an attempt to create
off-the-shelf dendritic cell vaccines that could target multiple
malignancies sharing the same antigen (NCT02387125).
Among the various known TAA options available for
immunotherapy to combat cancers is nucleolin (NCL), which was first
described as a nucleolar protein in Novikoff hepatoma cells and
Chinese Hamster Ovary cells (10,11). NCL is one of the most abundant
proteins in the nucleus and >90% of the total NCL levels in
cells are located there (12). It
is also found in the cytoplasm and on the cell membrane. Notably,
NCL on the cell surface can bind to various ligands to affect
numerous physiological functions, including ribosome biogenesis,
chromatin organization and stability, DNA and RNA metabolism,
cytokinesis, cell proliferation, angiogenesis, apoptosis
regulation, stress response and microRNA processing (13). The shuttling of NCL between the
nucleus, cytoplasm and plasmalemma is significantly higher in
cancer cells than in normal cells (14). Previous studies have demonstrated
the role of NCL in angiogenesis (15), epithelial-to-mesenchymal
transition and stemness (16,17). Increasing evidence from several
research articles and datasets associates an elevated NCL
expression with a poor prognosis of patients with various types of
cancer, including pediatric and adult ependymoma (18,19), hepatocellular carcinoma (20), non-small cell lung cancer
(21), esophageal squamous cell
carcinoma (22), B-cell lymphoma
(23), gastric cancer (24) and endometrial cancer (25). Although NCL is mainly present in
the nucleus, its functions in aggressive cancer progression have
been reported to be associated with its presence in the cancer cell
cytoplasm and membrane (26). The
nuclear, cytoplasmic and cell membrane expression of NCL can be
used as a prognostic marker and therapeutic target in various types
of cancer (26-28) (Table
I). A high cytoplasmic expression of NCL is associated with a
poor prognosis of patients with breast cancer, endometrial
carcinoma, gastric cancer and non-small cell lung cancer (24,25,29,30) whereas a high nuclear expression of
NCL is an independent good prognostic marker in gastric cancer,
endometrial carcinoma and pancreatic ductal adenocarcinoma
(17,24,25) (Table
I). However, the surface NCL has been linked to the poor
survival of patients with gastric cancer, rhabdomyosarcoma, breast
cancer, hepatocellular carcinoma, colon cancer and prostate cancer
(26,31-33) (Table
I). In agreement with the rational of a decrease in cell
surface NCL expression or activity, several in vitro and
in vivo studies showed that decreases in cell surface NCL
expression or activity inhibited the growth of cancer cells and
triggered apoptosis (34,35).
Epigenetic regulation is well known for its effect
on protein expression, which contributes to cancer initiation and
progression, including DNA methylation, histone modification,
nucleosome remodeling and RNA or microRNA-mediated targeting
molecules (36,37). The epigenetic regulators are
targetable and have become prognostic biomarkers for various types
of cancers including brain, breast, liver, ovary, prostate,
gastric, lung, colorectal cancers, melanoma and angiosarcoma
(36-39). NCL was also identified as an
epigenetic regulator of leukemogenesis (40). The overexpression of NCL has been
shown to promote the survival of leukemic cells by enhancing DNA
methyltransferase 1 expression and subsequently inducing DNA
hypermethylation followed by the epigenetic silencing of tumor
suppressor gene transcription (40). Therefore, targeting NCL as
epigenetic therapy may improve the survival of patients with
leukemia (40).
Thus, NCL is considered to be a promising target for
anticancer immunotherapy. In the present review, the nature of NCL
and the NCL-based targeting strategies under development are
discussed. Special focus has been paid to recently developed
cell-based immunotherapy approaches.
NCL is a 100-110-kDa protein that is a
multifunctional phosphoprotein expressed in exponentially growing
eukaryotic cells. The biophysical and biochemical research of NCL
mainly results from its multidomain structure. The human NCL gene
consists of 14 exons and 13 introns on chromosome 2q12-qter
(41). Mammalian NCL consists of
707 amino acids and a predicted molecular mass of ~77 kDa (13,42). The biophysical and biochemical
research of NCL has disclosed a multidomain structure comprising
three structural domains: An N-terminal domain rich in acidic
regions and containing multiple phosphorylation sites; a central
domain containing four RNA-binding domains (RBDs); and a C-terminal
domain containing multiple glycine, arginine and phenylalanine
residues (27,43). The N-terminal domain contains
acidic stretches (rich in glutamic acid and aspartic acid) and its
length varies greatly between the different NCL-like proteins
(43). This domain participates
in functions during the cell cycle that result in a high degree of
phosphorylation of cell division control protein 2 homolog
(p34cdc2), casein kinase 2, protein kinase C and
cyclin-dependent kinase 1 (13,44-46). The involvement of the N-terminal
domain in several protein-protein interactions has also been shown,
including interactions with components of the pre-rRNA processing
complex, and interactions with chromatin and untranslated regions
controlling rDNA transcription (47-50). The central domain has four domains
with RNA-binding domains (RBDs) or RNA-recognition motifs, and
their modulatory effects have been the focus of numerous studies on
cancer research (13). These
domains are known for their RNA-binding specificity (43). One of the functions of NCL RBDs is
assisting in pre-RNA transcription where they interact with the
stem-loop structure of 18S and 28S ribosomal RNA (51), helping the pre-rRNA to fold
correctly (52). Moreover,
studies have demonstrated that the RBDs of NCL function in RNA
packaging, pre-mRNA splicing, poly-A tail synthesis, maturation,
mRNA stability and translational control (53). The C-terminal domain is rich in
arginine-glycine-glycine repeats interspersed with several aromatic
amino acids (43,54), and can interact with a number of
target mRNAs and proteins (55).
Fractionation studies have located >90% of the
NCL in the nucleolar portion of the cellular pool (12,27). The distribution of NCL between the
cytoplasm and membrane is difficult to estimate, as it is dependent
on the quality of the fractionation or sensitivity of the detection
techniques (56,57). However, NCL appears to be
ubiquitously distributed, being not only in the nucleus, but also
in the cytoplasm and cell membrane (22). The distribution of NCL between
cellular compartments, shuttling between the nucleus, cytoplasm and
membrane, plays a significant role in the diverse mechanisms of NCL
involvement in cancer (26,27). Several methylations and
phosphorylations of the NH2 terminus of the NCL protein
are required for the nuclear export of NCL to the cytoplasm
(58,59). Highly proliferative cells, such as
metabolically active cancer cells (60), have the common features of NCL
overexpression and localization in the cytoplasm and cell membrane
(27).
NCL on the cell surface serves as an anchor protein
that binds various molecules implicated in cell differentiation,
adhesion, trafficking, inflammation, angiogenesis and cancer
development (12,67-69). The mechanism whereby NCL is
translocated to the plasma membrane remains unclear. Accumulating
evidence validates the conclusion that the NCL, which is localized
on the surface of various types of cancer cells, but not on their
normal counterparts, is an effective strategic target for the
treatment of cancer (68). By
binding with Fas (70), or with
Ras via the C-terminal domain (71), NCL significantly promotes cancer
cell proliferation and inhibits apoptosis by decreasing the
expression of the pro-apoptotic BAX gene (72). Targeting cell-surface NCL might
trigger multiple inhibitory effects, depending on the cell type
(68,73). Taken together, to date, these
observations highlight a critical role for NCL in tumorigenesis and
tumor progression. However, potential methods with which to
modulate NCL for cancer immunotherapeutic purposes have been
challenging to implement.
The overexpression of surface NCL on cancer cells,
compared to normal cells, provides a very promising target for
cancer therapy, since targeting it does not alter NCL expression in
the nucleus and generates minimal toxicity (27). Among the various NCL-binding
ligands that have been tested thus far, the F3 peptide and the
AS1411 aptamer are those that have been explored the most
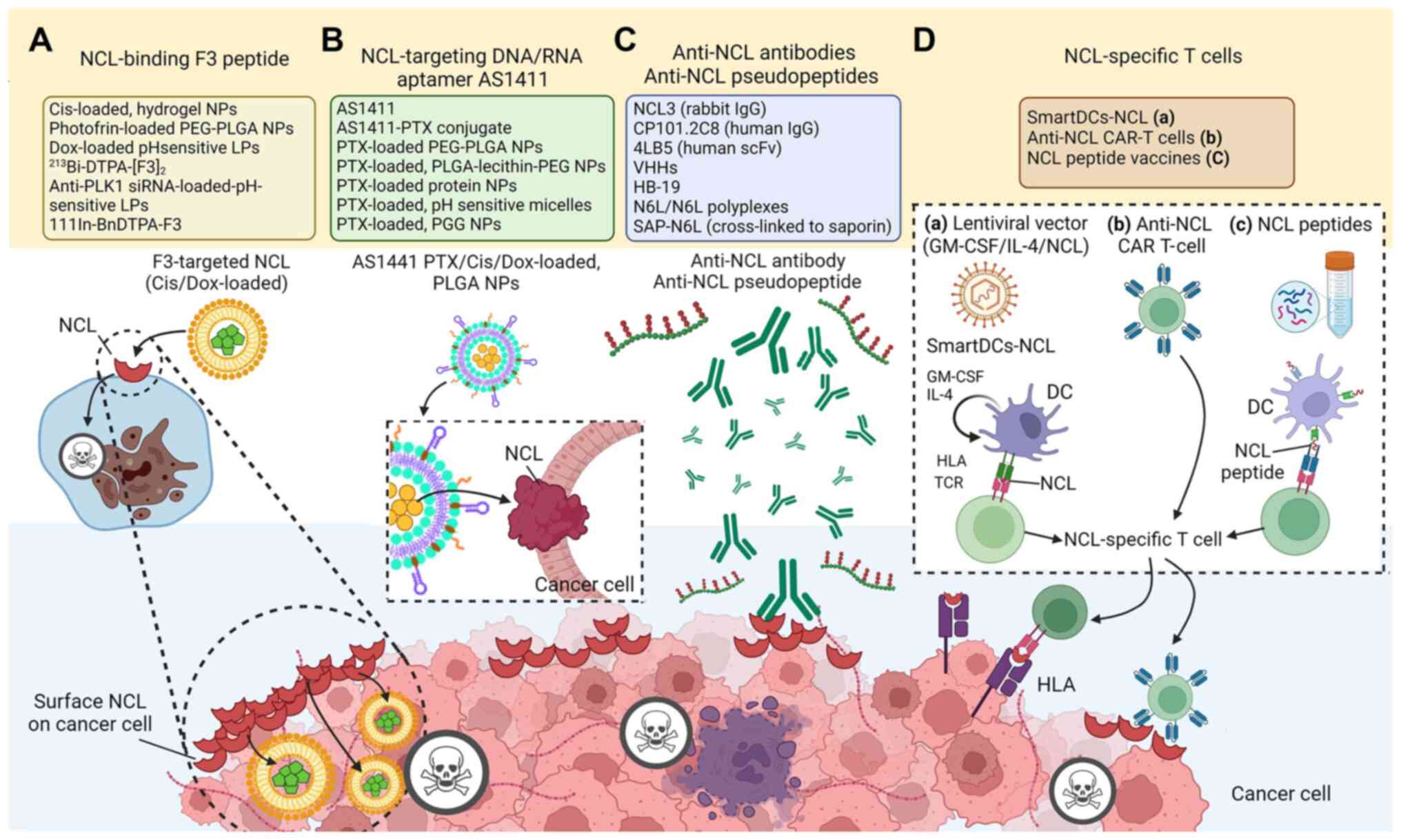
extensively (47).
The NCL-binding F3 peptide is a 31-amino acid
(KDEPQRRS ARLSAKPAPPKPEPKPKKAPAKK) (84) that was generated by a
phage-displayed cDNA library, and selected using ex vivo
screening on cell suspensions prepared from mouse bone marrow and
in vivo screening for tumor homing (85). The use of NCL-binding F3 peptides
has been encouraged for intracellular drug delivery (Table II and Fig. 1A). F3 homes to surface-expressed
NCL as a receptor in cancer cells, neovasculature and endothelial
cells (86). After the F3 peptide
binds at the N-terminal domain of NCL, it is internalized into the
targeted cell and translocated to the nucleus (85,86). The C-terminus of the F3 peptide
has been utilized as an effective carrier of nanoparticles (NPs)
for chemotherapeutic drugs, such as doxorubicin (87,88), cisplatin (Cis) (89), paclitaxel, the sphingolipid,
C6-ceramide (inhibitor of the PI3K pathway) (90), or photodynamic therapy agents
(91), and it has been chemically
conjugated to radiotherapeutics (92). A range of cancer models has been
utilized (77,87,93-116). Among notable findings, after
intravenous administration of an F3 peptide-targeted Cis-loaded
hydrogel NP there was decreased Cis-sensitive-derived or
Cis-resistant-derived ovarian cancer progression (89). F3 peptide has also been used for
the delivery of photodynamic therapeutic agents, dextran-coated
iron oxide NPs, siRNAs and oligonucleotides to various tumor cell
lines and tumor xenografts (117,118). In a previous study, results were
obtained from institutional pathology reports and reviewed by
experienced pathologists from the Portuguese Institute of Oncology,
Coimbra, Portugal, which confirmed that the uptake of F3-targeted
liposomes by cancer cells was associated only with the expression
of the NCL receptor (88).
Indeed, compared to treatments with phages and antibodies, the
small mass of F3 peptide (~5 kDa) permitted wider distribution in
cells (86). Moreover, an
anti-NCL single variable domain has been generated on the variable
domain of heavy-chain antibodies (VHH) (nanobodies) by grafting
10-amino acids of F3 peptide-derived NCL-binding sequences onto
complementarity-determining regions (CDR)1 or CDR3 of a parental
VHH to yield anti-NCL-CDR1 VHH and anti-NCL-CDR3 VHH. The two
different anti-NCL VHHs exhibited maximal binding and increased
cytotoxicity against an NCL-overexpressing breast cancer cell line
(119,120). In a previous study, the
treatment of Polo-like kinase 1 (PLK1)-overexpressing prostate
cancer and angiogenic endothelial cells with an F3-targeted
liposomal nanocarrier entrapping an anti-PLK1 siRNA significantly
decreased cell viability (121).
In preclinical models, targeted drug delivery into tumors has been
achieved by the conjugation of anticancer drugs with F3
tumor-homing peptides. An F3 dimer coupled to the
radiopharmaceutical DTPA chelated with 213Bi,
(213Bi-DTPA-[F3]2), has been tested for
binding affinity, bio-distribution, and anti-tumor activity in
vitro and in vivo in a preclinical model of peritoneal
carcinomatosis (122). In
another study, The 213Bi-DTPA-[F3]2 was
highly cytotoxic without severe side-effects in animals (122), significantly reduced clonogenic
survival in vitro, and delayed tumor growth in vivo
(84).
An aptamer is a short single-stranded nucleic acid
sequence, either DNA or RNA, that can specifically target cellular
and extracellular targets with high affinity (12). The aptamer AS1411, unmodified
guanosine (G)-rich oligonucleotide (5′-d GGT GGT GGT GGT TGT GGT
GGT GGT GG-3′), has a high affinity for NCL, binds to overexpressed
NCL on the cell surface, and is then internalized (123,124), without adverse effects on normal
cells (125-127). AS1411 functions as a molecular
decoy, blocking and shortening the half-life of
shuttle-NCL-regulated RNAs (128). The AS1411-based strategies have
been used to target NCL with a delivery system of NPs containing
agents such as siRNA, or small drugs (129,130) (Table II and Fig. 1B).
The first successful transition of an aptamer into
clinical trials for cancer treatment was that of an NCL-targeting
DNA aptamer AS1411 (119,120).
In phase I clinical trials, three AS1411-based agents have been
assessed for safety and efficacy in advanced solid tumors
(ClinicalTrials.gov: NCT00881244) and
acute myeloid leukemia (AML) (ClinicalTrials.gov: NCT00512083). The first, tested in
NCT00881244, after it was demonstrated to significantly reduce
tumor growth in xenograft models of both renal and lung cancers
(131-133), demonstrated that 50% of 30
patients with advanced solid tumors had stable symptoms for 2-9
months after AS1411 administration without severe side-effects
(134).
During the development of clinical trials to further
assess the efficacy of the AS1411 aptamer regarding the overall
rate of response on a larger number of patients, the therapeutic
effects of other types of AS1411 aptamer-conjugated agents were
tested, including poly-lactic-co-glycolic acid (PLGA)-based NPs
(PLGA-NPs) and lipid polymers (144). Several preclinical models are
also being used to investigate the potential of the aptamer in
transport mechanisms for other anticancer drugs (Table II, Fig. 1B). For example, AS1411-conjugated
gold nanospheres have exhibited an enhanced cellular uptake, and
increased anti-proliferative and cytotoxic effects in both in
vitro and in vivo experiments (144). In lung cancer cells
overexpressing NCL, AS1411-gemcitabine-NPs have been shown to exert
an inhibitory effect on cell proliferation (145). Moreover, the AS1411-coated
polymeric nanosystem can function as a potential drug delivery
mechanism against various types of cancer, such as ovarian,
pancreatic and lung cancer (146). Targeting surface NCL
concurrently impairs the progression of several cancer cell types
in vitro and in vivo. AS1411 targets nuclear NCL,
leading to the re-localization of the protein arginine
methyltransferase 5-NCL complex from the nucleus to the cytoplasm
of prostate cancer, which may decrease malignant transformation
(147). Moreover, the
RNA-binding activity of cytoplasmic NCL to various mRNAs is
inhibited by AS1411 aptamer, resulting in the induction of tumor
cell death (148).
Active cancer immunotherapy uses the strategy of
deploying the immune system of cancer patients to destroy tumors
and prevent their recurrence (149). As proposed in Coley's toxins,
agents with potent immunostimulatory properties (adjuvants) have
achieved favorable responses in various types of cancer (150) and have significantly facilitated
the advent of novel immunotherapy technologies. In recent years,
cancer immunotherapy has made a significant breakthrough due to the
development of adoptive T-cell therapies and the use of monoclonal
antibodies that block the cytotoxic T-lymphocyte (CTL) antigen-4
(CTLA-4) and programmed cell death 1 (PD-1) immune checkpoints
(151). Another approach that is
often employed is the identification of TAA molecule(s) that are
either selectively expressed or overexpressed by tumors (152). As discussed herein, NCL may be
the most prominent eligible structure, which is stable on the cell
surface of proliferating cells (153); hence, it represents an
attractive target for immune-based anticancer agents.
Anti-NCL antibody production is the adaptive immune
system response to the detection of the antigen (47). The antibody-based targeting of NCL
has been previously explored (Table
II and Fig. 1C). Treatment
with anti-NCL3 antibodies has been shown to decrease the viability
of both angiogenic endothelial and melanoma cells, and this is
accompanied by the downregulation of BCL-2 expression (154). The same effect was observed in a
murine model bearing breast cancer, in which anti-NCL3 antibody
administration resulted in decreased tumor hypoxia (155). In another study using a leukemic
xenograft mouse model, the anti-NCL3 antibody reduced cell
viability without triggering immune responses (47,155), whereas CP101.2C8, which binds to
the RBDs of NCL, led to a 30% greater mouse survival rate compared
to the control IgG isotype group, and reduced the viability of
leukemic cells (155).
Furthermore, the generation of anti-NCL antibodies
has also been explored in the context of single-chain fragment
variable (scFv) and VHH antibodies. The 4LB5 antibody, an scFv
antibody that binds to the RNA-binding domain of cell-surface NCL,
was previously found to decrease cell viability, clonogenicity and
tumor growth in xenografts, while inducing the apoptosis of breast
cancer and hepatocellular carcinoma (31). Indeed, a decreased tumor growth
was observed in the in vivo model following the 4LB5
administration with no evidence of adverse effects (31). Novel anti-NCL VHHs have also been
developed using a strategy of grafting F3 peptide-derived
NCL-binding sequences onto a VHH CDR1 or CDR3 (111). These VHHs are more compact and
smaller (~15 kDa) than scFv, with a stable immunoglobulin fold
(156). The CDR3-grafted VHH
enabled a significant antibody-dependent cell-mediated cytotoxicity
effect against breast cancer cells with a 2-fold increase in cell
death (111).
HB-19 is a surface NCL-antagonistic pseudopeptide
with a pentavalent structure comprising the tripeptide
lysine-glycine-proline (with a reduced bond between the lysine and
proline residues) coupled to an 8-amino-acid template (15,33,157-180) (Fig. 1C). This pseudopeptide is
translocated to the cytoplasm, but not to the nucleus, leading to a
reduction in NCL levels in the cytoplasm and at the cell surface
(33,157,172-175). The binding of HB-19 to surface
NCL can form an irreversible complex that inhibits both tumor
progression and angiogenesis (171). HB-19 treatment has been shown to
suppress the tumor growth of established human breast cancer
xenografts and exhibits low toxicity to normal tissue (15). HB-19 has been shown to impair
angiogenesis, the formation of capillary-like structures, and blood
vessel branching in chick embryo chorioallantoic membranes
(15). Moreover, the colony
formation capacity of several cancer cell lines in soft agar was
decreased, which underscored the anticancer activities of HB-19
(15,167,169).
Multivalent pseudopeptide NucAnt 6L (N6L) also
specifically binds to the surface NCL of cancer and endothelial
cells and displays antitumor activities (176). It was synthesized to improve the
biological actions of HB-19 by using a specific antagonist that
binds the RGG domain of the NCL C-terminal tail (170). The normalization of tumor
vessels and improved delivery and efficacy of chemotherapeutic
drugs were observed as a consequence of NCL inhibition by N6L
(17). The combination of N6L and
mTOR inhibitors was found to synergistically inhibit the
proliferation and viability of pancreatic cancer in both 2 and 3-D
preclinical models (179). In
another study, the pseudopeptide N6L cross-linked to saporin-S6
(SAP-N6L) induced the internalization of toxin to glioblastoma
cancer cells and enhanced the toxic activity 1,000-fold compared to
saporin alone in glioblastoma primary cells. The conjugated saporin
and N6L induced glioblastoma cell death at low nanomolar
concentrations and induced more apoptotic cell death than saporin
alone (177). Moreover, N6L
incorporated as polyplexes with nanoparticles (N6L-polyplexes)
exhibited enhanced anti-tumor activities against pancreatic cancer.
Gemcitabine treatment provides a standard of care for this cancer
type. However, in an in vivo model of pancreatic ductal
adenocarcinoma, the administration of N6L-polyplexes led to a
significantly greater reduction of tumor volume than the
administration of gemcitabine (175).
An active approach to obtaining NCL-specific
T-cells takes advantage of the potent antigen-presenting capacity
of dendritic cells (DCs) loaded with NCL antigen (181) (Table II and Fig. 1D-a). The self-differentiated
myeloid-derived APC reactive against tumors presenting NCL
(SmartDCs-NCL) are monocyte-derived DC transduced with lentivirus
harboring tricistronic complementary DNA sequences encoding
cytokines (granulocyte-macrophage-colony stimulating factor;
interleukin-4) and NCL antigen. A high copy number of integrated
lentiviral vectors per cell in the SmartDCs platform led to
~1.5-3.0-fold self-differentiation more than the control vector
(182), based on the endogenous
load on the major histocompatibility complex (MHC) class I complex
at the cell surface of APCs. DCs that acquire APC-like function
provide the majority of endogenous MHC class I ligands for
association with CD8+ CTLs (183). SmartDCs-NCL expressed all DCs
markers, including the downregulation of CD14 and the upregulation
of CD11c, CD40, CD80, CD83, CD86 and human leukocyte antigen
(HLA)-DR, as compared with monocytes. The T-cells established
following co-culture with SmartDCs-NCL comprised markedly increased
CD3+/CD8+/CD45RA−/CD62L−
effector memory T-cells (184),
when compared to those generated by conventionally produced DCs.
Furthermore, MHC class I-restricted stimulation of allogenic
T-cells by the NCL-specific T-cells was verified by higher levels
of interferon (IFN)-γ production. NCL-specific T-cells activated
with SmartDCs-NCL gave significantly increased percentages of
specific cell lysis at effector:target ratios of 1:1, 5:1, and 10:1
against NCLHigh MDA-MB-231 and HCC70 TNBC cells. No
killing activities of NCL-specific T-cells were detected against
normal mammary cells expressing no NCL (181).
Among the immunotherapeutic strategies that are
recurrently being conducted worldwide in preclinical studies or
clinical trials, the use of immune checkpoint inhibitors is
particularly prominent (185).
In a previous study, the combined treatment with
SmartDCs-NCL-activated NCL-specific T-cells and anti-programmed
cell death ligand 1 (PD-L1) peptide (CLQKTPKQC) (186) yielded significant killing of
NCLHigh/PD-L1High MDA-MB-231 and
NCLHigh/PD-L1High HCC70 TNBC cells.
NCL-specific T-cell-mediated TNBC cell killing was mediated through
both apoptotic and autophagic pathways. The higher lytic potential
was also revealed by exposing TNBC cells to an autophagy inducer,
the medicinal herb curcumin, which attenuated PD-L1 expression
(181). The degradation of PD-L1
via the autophagy pathway has been reported (187-189). Indeed, curcumin can suppress
PD-L1 production in cancer cells through p62-mediated autophagy.
This is the first small-scale study that opens the door for
potential clinical approaches targeting not only TNBC but also
NCLHigh/PD-L1High cancer cells with
NCL-specific T-cells in combination with a PD-L1 inhibitor or
autophagic stimulator (181).
Further intensive studies may lead to the incorporation of the
adoptive transfer of NCL-specific T-cells into the standard of care
protocol for clinical applications.
Currently, the identification of TAA and the
subsequent identification of T-cell epitopes render 'reverse
immunology' applicable to the treatment of malignant tumors
(191). This approach consists
in raising T-cells against specific peptides corresponding to
fragments of conventional proteins. New opportunities for cancer
prevention and treatment can arise from peptides that promise a
novel class of anticancer agents that can specifically target
cancer cells, while remaining non-toxic to normal tissues (192). The ability of some TAA peptides
to specifically recognize and bind to the membrane proteins of
cancer cells could overcome the issues associated with the high
molecular weight, low tissue diffusion, slow delivery and the poor
cellular uptake of therapeutic antibodies (193-195). Moreover, peptide-based therapies
are attractive over other treatment forms as peptides are easily
synthesized on a large scale, chemically stable, have a high in
vivo biocompatibility, are free of contaminating substances
such as bacterial pathogens and are devoid of oncogenic potential
(193,196). In breast cancer, HER-2/neu is
overexpressed in 15 to 30% of patients and is associated with a
poor prognosis (197). The E75
and GP2 HER-2/neu binding peptides were identified by their
antigen-specific T-cell responses and tested in phase 1 clinical
trial in patients with metastatic breast and ovarian cancer
(197,198). Moreover, the identification of
two novel 9-mer peptides, M1.1 and M1.2, with high binding affinity
to HLA-A*02, could lead to their application in vaccine therapies
for breast and ovarian cancer, based on the finding that >90% of
the cancer cells of patient overexpress human mucin 1 (MUC-1)
protein (199,200). More importantly, MUC1-specific
CTLs were induced in patients with breast and ovarian cancer after
vaccination with DC that had been pulsed with these peptides
(200). Very recently, in
silico predictive computational algorithms have revealed 9-mer
NCL epitope peptides with major HLA-A*02-binding motifs
(KMAPPPKEV15-23 and VLSNLSYSN488-496) in the
full-length NCL sequence (181)
(Fig. 1D-c). In in vitro
experiments using these 2 NCL peptides, high IFN-γ-producing
NCL-specific T-cells that kill NCL-positive cancer cells were
observed (unpublished data).
NCL is an ubiquitous multifunctional protein with a
tripartite domain structure that includes an acidic histone-like
N-terminus, a central domain containing four RBDs, and an arginine
and glycine-rich C-terminus (27,43). It functions as a shuttle
phosphoprotein between nucleolus and cytoplasm in its diverse roles
in cell growth, proliferation, carcinogenesis, angiogenesis,
metastasis and cell death (13,43). Surface NCL is overexpressed in
cancer cells, which renders it an attractive target for cancer
treatment (12,26,67-69). A high level of cytoplasmic NCL, or
cytoplasmic NCL and total NCL together, can provide a potent
predictive marker for a shorter OS, whereas a high level of nuclear
NCL presents a predictive marker for a long OS and DFS of patients
(26). On the whole, this
evidence indicates that NCL not only provides a potential biomarker
for cancer diagnosis and prognosis, but also that anti-NCL
therapies could represent a novel opportunity for cancer
treatment.
A broad range of studies now supports this
potential for cancer immunotherapy. Targeting surface NCL impaired
the progression of several cancer cell types in vitro and
in vivo. Furthermore, a wide variety of ligands targeting
cell-surface NCL has been reported to block cancer cell growth and
progression, including aptamer AS1411 (140-143), F3 peptides (77,84,86-88,90,92-117), antibodies against NCL (31,47,111,154,155,201) and the multivalent pseudopeptides
(HB-19 and N6L) (15,33,157-180). The pace of clinical trials based
on this approach is accelerating. Four aptamers AS1411 have been
evaluated in clinical phase I/II including ClinicalTrials.gov: NCT00881244, NCT00512083,
NCT00740441 and NCT01034410 (140-143). Two anti-NCL antibodies have been
evaluated in clinical trials with the outcome of proliferation
reduction. The antagonistic pseudo peptide N6L showed good results
against solid tumors during a phase I/IIa clinical trial
[ClinicalTrials.gov: NCT01711398
(202)], and a phase II trial is
currently underway.
An important criterion for the selection of TAAs
for personalized immunization is that it should be expressed by
more than two-fold compared to para-tumor tissues (152). This property could allow for the
induction of antigen-specific T-cells for patients with relatively
low expression levels in normal tissues since immunization induced
anti-TAA-specific CD4+ and CD8+ T-cell
responses, which were enhanced by 0.1 to 10.0% (152). This highlights the potential of
using protein overexpression, which is associated with a favorable
OS for personalized TAA immunization. Sensitivity to NCL-specific
T-cells is dependent on the level of NCL expressed in cancer cells
(181). The in vitro data
showed that NCL-specific T-cells activated by the DC-based platform
decreased the viability of NCL-overexpressing breast cancer cells.
Notably, the effects of NCL-specific T-cells, apoptosis and
autophagic cell death, can be enhanced by an anti-PD-L1 peptide or
autophagy modulator. Accordingly, treatment with NCL-specific
T-cells is compatible with a combined approach, including immune
checkpoint inhibitors for effective cancer patient treatment
(181,190). However, at this time, the number
of experiments is too limited to draw firm conclusions. Large-scale
in vitro and clinical trials are required to validate the
safety and effectiveness of this approach.
It is widely accepted that NCL has a functional
role in cancer regulation. The present up-to-date review provides
data to support the notion of targeting NCL among the developing
anticancer approaches, including the use of anti-NCL peptides,
aptamers, anti-NCL antibodies/pseudopeptides and NCL-specific
T-cells. The NCL-specific T-cells generated by APCs or NCL peptides
illustrate the use of the platforms that open a window for using
immunotherapy targeted at NCL as another very promising approach in
NCL-positive cancers. Further studies involving a specific target
for each compartment of NCL are required in order to obtain a
better understanding of the mechanisms through which different
compartments of NCL contribute to the distinct prognoses of
patients with cancer. Importantly, precision NCL-based targeting
approaches with immunological potential and low toxicity need to be
tested in both preclinical and clinical research to extend the
value of NCL-targeting strategies.
Not applicable.
ST, PT, PTY and CT were involved in the
conceptualization of the study. ST, KA and CT were involved in the
writing and preparation of the original draft. CT critically
reviewed and finalized the manuscript. All authors have read and
approved the final manuscript. Data authentication is not
applicable.
In the section '3. Therapeutic strategies
exploiting NCL-based targeting', the in vitro experiments
mentioned (unpublished data) were performed using biological
samples of both healthy donors and patients with breast cancer.
These experiments were approved by Siriraj Institutional Review
Board (COA no. Si 580/2018). Informed consent was obtained from all
subjects involved in the study.
Not applicable.
The authors declare that they have no competing
interests.
The authors gratefully acknowledge Dr Jan O'Day
Davies, Mahidol University, Bangkok, Thailand, for the English
editing of the manuscript.
The present study was funded by the New Researcher Grant,
Mahidol University (grant no. R016420006), the Siriraj Research
Fund, Faculty of Medicine Siriraj Hospital, Mahidol University
(grant no. R016334002), a Research Grant, National Research Council
of Thailand, and Mid-Research Grant (N42A650343), National Research
Council of Thailand and Mahidol University (grant no.
R016541043).
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fidler MM, Bray F and Soerjomataram I: The
global cancer burden and human development: A review. Scand J
Public Health. 46:27–36. 2018. View Article : Google Scholar
|
|
3
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar
|
|
4
|
Gotwals P, Cameron S, Cipolletta D,
Cremasco V, Crystal A, Hewes B, Mueller B, Quaratino S,
Sabatos-Peyton C, Petruzzelli L, et al: Prospects for combining
targeted and conventional cancer therapy with immunotherapy. Nat
Rev Cancer. 17:286–301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Velcheti V and Schalper K: Basic overview
of current immunotherapy approaches in cancer. Am Soc Clin Oncol
Educ Book. 36:298–308. 2016. View Article : Google Scholar
|
|
7
|
Rodríguez Pérez Á, Campillo-Davo D, Van
Tendeloo V and Benitez-Ribas D: Cellular immunotherapy: A clinical
state-of-the-art of a new paradigm for cancer treatment. Clin
Transl Oncol. 22:1923–1937. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rosenberg SA and Restifo NP: Adoptive cell
transfer as personalized immunotherapy for human cancer. Science.
348:62–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schuster M, Nechansky A and Kircheis R:
Cancer immunotherapy. Biotechnol. 1:138–147. 2006.
|
|
10
|
Bugler B, Caizergues-Ferrer M, Bouche G,
Bourbon H and Amalric F: Detection and localization of a class of
proteins immunologically related to a 100-kDa nucleolar protein.
Eur J Biochem. 128:475–480. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Orrick LR, Olson MO and Busch H:
Comparison of nucleolar proteins of normal rat liver and Novikoff
hepatoma ascites cells by two-dimensional polyacrylamide gel
electrophoresis. Proc Natl Acad Sci USA. 70:1316–1320. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z and Xu X: Roles of nucleolin: Focus
on cancer and anti-cancer therapy. Saudi Med J. 37:1312–1318. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jia W, Yao Z, Zhao J, Guan Q and Gao L:
New perspectives of physiological and pathological functions of
nucleolin (NCL). Life Sci. 186:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu W, Wang G, Sun X, Ye J, Wei F, Shi X
and Lv G: The involvement of cell surface nucleolin in the
initiation of CCR6 signaling in human hepatocellular carcinoma. Med
Oncol. 32:752015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Destouches D, El Khoury D, Hamma-Kourbali
Y, Krust B, Albanese P, Katsoris P, Guichard G, Briand JP, Courty J
and Hovanessian AG: Suppression of tumor growth and angiogenesis by
a specific antagonist of the cell-surface expressed nucleolin. PLoS
One. 3:e25182008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ugrinova I, Petrova M, Chalabi-Dchar M and
Bouvet P: Multifaceted nucleolin protein and its molecular partners
in oncogenesis. Adv Protein Chem Struct Biol. 111:133–164. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gilles ME, Maione F, Cossutta M,
Carpentier G, Caruana L, Di Maria S, Houppe C, Destouches D,
Shchors K, Prochasson C, et al: Nucleolin targeting impairs the
progression of pancreatic cancer and promotes the normalization of
tumor vasculature. Cancer Res. 76:7181–7193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ridley L, Rahman R, Brundler MA, Ellison
D, Lowe J, Robson K, Prebble E, Luckett I, Gilbertson RJ, Parkes S,
et al: Multifactorial analysis of predictors of outcome in
pediatric intracranial ependymoma. Neuro Oncol. 10:675–689. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Modena P, Buttarelli FR, Miceli R,
Piccinin E, Baldi C, Antonelli M, Morra I, Lauriola L, Di Rocco C,
Garrè ML, et al: Predictors of outcome in an AIEOP series of
childhood ependymomas: A multifactorial analysis. Neuro Oncol.
14:1346–1356. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo X, Xiong L, Yu L, Li R, Wang Z, Ren B,
Dong J, Li B and Wang D: Increased level of nucleolin confers to
aggressive tumor progression and poor prognosis in patients with
hepatocellular carcinoma after hepatectomy. Diagn Pathol.
9:1752014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang F, Wu Y, Tan H, Guo T, Zhang K, Li D
and Tong Z: Phosphorylation of nucleolin is indispensable to its
involvement in the proliferation and migration of non-small cell
lung cancer cells. Oncol Rep. 41:590–598. 2019.
|
|
22
|
Qi J, Li H, Liu N, Xing Y, Zhou G, Wu Y,
Liu Y, Chen W, Yue J, Han B, et al: The implications and mechanisms
of the extra-nuclear nucleolin in the esophageal squamous cell
carcinomas. Med Oncol. 32:452015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jain N, Zhu H, Khashab T, Ye Q, George B,
Mathur R, Singh RK, Berkova Z, Wise JF, Braun FK, et al: Targeting
nucleolin for better survival in diffuse large B-cell lymphoma.
Leukemia. 32:663–674. 2018. View Article : Google Scholar :
|
|
24
|
Qiu W, Zhou F, Zhang Q, Sun X, Shi X,
Liang Y, Wang X and Yue L: Overexpression of nucleolin and
different expression sites both related to the prognosis of gastric
cancer. APMIS. 121:919–925. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin Q, Ma X, Hu S, Li R, Wei X, Han B, Ma
Y, Liu P and Pang Y: Overexpression of nucleolin is a potential
prognostic marker in endometrial carcinoma. Cancer Manag Res.
13:1955–1965. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yangngam S, Prasopsiri J, Hatthakarnkul P,
Thongchot S, Thuwajit P, Yenchitsomanus PT, Edwards J and Thuwajit
C: Cellular localization of nucleolin determines the prognosis in
cancers: A meta-analysis. J Mol Med (Berl). 100:1145–1157. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Berger CM, Gaume X and Bouvet P: The roles
of nucleolin subcellular localization in cancer. Biochimie.
113:78–85. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yenchitsomanus PT, Vasuvattakul S, Kirdpon
S, Wasanawatana S, Susaengrat W, Sreethiphayawan S, Chuawatana D,
Mingkum S, Sawasdee N, Thuwajit P, et al: Autosomal recessive
distal renal tubular acidosis caused by G701D mutation of anion
exchanger 1 gene. Am J Kidney Dis. 40:21–29. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu JY, Lu S, Xu XY, Hu SL, Li B, Li WX and
Chang JY: Prognostic significance of nuclear or cytoplasmic
nucleolin expression in human non-small cell lung cancer and its
relationship with DNA-PKcs. Tumour Biol. 37:10349–10356. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Willmer T, Damerell V, Smyly S, Sims D, Du
Toit M, Ncube S, Sinkala M, Govender D, Sturrock E, Blackburn JM
and Prince S: Targeting the oncogenic TBX3: Nucleolin complex to
treat multiple sarcoma subtypes. Am J Cancer Res. 11:5680–5700.
2021.
|
|
31
|
Palmieri D, Richmond T, Piovan C, Sheetz
T, Zanesi N, Troise F, James C, Wernicke D, Nyei F, Gordon TJ, et
al: Human anti-nucleolin recombinant immunoagent for cancer
therapy. Proc Natl Acad Sci USA. 112:9418–9423. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dzhumashev D, Timpanaro A, Ali S, De
Micheli AJ, Mamchaoui K, Cascone I, Rössler J and Bernasconi M:
Quantum Dot-based screening identifies F3 peptide and reveals cell
surface nucleolin as a therapeutic target for rhabdomyosarcoma.
Cancers (Basel). 14:50482022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fujiki H, Watanabe T and Suganuma M:
Cell-surface nucleolin acts as a central mediator for carcinogenic,
anti-carcinogenic, and disease-related ligands. J Cancer Res Clin
Oncol. 140:689–699. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meng GZ, Xiao SJ, Zeng SE and Li YQ:
Downregulation of cell-surface-expressed nucleolin inhibits the
growth of hepatocellular carcinoma cells in vitro. Zhonghua Zhong
Liu Za Zhi. 33:23–27. 2011.In Chinese. PubMed/NCBI
|
|
35
|
D'Avino C, Palmieri D, Braddom A, Zanesi
N, James C, Cole S, Salvatore F, Croce CM and De Lorenzo C: A novel
fully human anti-NCL immunoRNase for triple-negative breast cancer
therapy. Oncotarget. 7:87016–87030. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dawson MA and Kouzarides T: Cancer
epigenetics: From mechanism to therapy. Cell. 150:12–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nebbioso A, Tambaro FP, Dell'Aversana C
and Altucci L: Cancer epigenetics: Moving forward. PLoS Genet.
14:e10073622018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gougousis S, Petanidis S, Poutoglidis A,
Tsetsos N, Vrochidis P, Skoumpas I, Argyriou N, Katopodi T and
Domvri K: Epigenetic editing and tumor-dependent immunosuppressive
signaling in head and neck malignancies. Oncol Lett. 23:1962022.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Toh TB, Lim JJ and Chow EKH: Epigenetics
in cancer stem cells. Mol Cancer. 16:292017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shen N, Yan F, Pang J, Wu LC, Al-Kali A,
Litzw MR and Liu S: A nucleolin-DNMT1 regulatory axis in acute
myeloid leukemogenesis. Oncotarget. 5:5494–5509. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tuteja R and Tuteja N: Nucleolin: A
multifunctional major nucleolar phosphoprotein. Crit Rev Biochem
Mol Biol. 33:407–436. 1998. View Article : Google Scholar
|
|
42
|
Mamrack MD, Olson MO and Busch H: Amino
acid sequence and sites of phosphorylation in a highly acidic
region of nucleolar nonhistone protein C23. Biochemistry.
18:3381–3386. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ginisty H, Sicard H, Roger B and Bouvet P:
Structure and functions of nucleolin. J Cell Sci. 112:761–772.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Peter M, Nakagawa J, Dorée M, Labbé JC and
Nigg EA: Identification of major nucleolar proteins as candidate
mitotic substrates of cdc2 kinase. Cell. 60:791–801. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Caizergues-Ferrer M, Belenguer P, Lapeyre
B, Amalric F, Wallace MO and Olson MO: Phosphorylation of nucleolin
by a nucleolar type NII protein kinase. Biochemistry. 26:7876–7883.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Belenguer P, Caizergues-Ferrer M, Labbé
JC, Doree M and Amalric F: Mitosis-specific phosphorylation of
nucleolin by p34cdc2 protein kinase. Mol Cell Biol. 10:3607–3618.
1990.PubMed/NCBI
|
|
47
|
Romano S, Fonseca N, Simões S, Gonçalves J
and Moreira JN: Nucleolin-based targeting strategies for cancer
therapy: From targeted drug delivery to cytotoxic ligands. Drug
Discov Today. 24:1985–2001. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ghisolfi-Nieto L, Joseph G,
Puvion-Dutilleul F, Amalric F and Bouvet P: Nucleolin is a
sequence-specific RNA-binding protein: Characterization of targets
on pre-ribosomal RNA. J Mol Biol. 260:34–53. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cong R, Das S, Ugrinova I, Kumar S,
Mongelard F, Wong J and Bouvet P: Interaction of nucleolin with
ribosomal RNA genes and its role in RNA polymerase I transcription.
Nucleic Acids Res. 40:9441–9454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Roger B, Moisand A, Amalric F and Bouvet
P: Nucleolin provides a link between RNA polymerase I transcription
and pre-ribosome assembly. Chromosoma. 111:399–407. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Serin G, Joseph G, Ghisolfi L, Bauzan M,
Erard M, Amalric F and Bouvet P: Two RNA-binding domains determine
the RNA-binding specificity of nucleolin. J Biol Chem.
272:13109–13116. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Allain FH, Bouvet P, Dieckmann T and
Feigon J: Molecular basis of sequence-specific recognition of
pre-ribosomal RNA by nucleolin. EMBO J. 19:6870–6881. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ishikawa F, Matunis MJ, Dreyfuss G and
Cech TR: Nuclear proteins that bind the pre-mRNA 3′ splice site
sequence r(UUAG/G) and the human telomeric DNA sequence d(TTAGGG)n.
Mol Cell Biol. 13:4301–4310. 1993.PubMed/NCBI
|
|
54
|
Lapeyre B, Amalric F, Ghaffari SH, Rao SV,
Dumbar TS and Olson MO: Protein and cDNA sequence of a
glycine-rich, dimethylarginine-containing region located near the
carboxyl-terminal end of nucleolin (C23 and 100 kDa). J Biol Chem.
261:9167–9173. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ghisolfi L, Joseph G, Amalric F and Erard
M: The glycine-rich domain of nucleolin has an unusual
supersecondary structure responsible for its
RNA-helix-destabilizing properties. J Biol Chem. 267:2955–2959.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gaume X, Tassin AM, Ugrinova I, Mongelard
F, Monier K and Bouvet P: Centrosomal nucleolin is required for
microtubule network organization. Cell Cycle. 14:902–919. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Scherl A, Couté Y, Déon C, Callé A,
Kindbeiter K, Sanchez JC, Greco A, Hochstrasser D and Diaz JJ:
Functional proteomic analysis of human nucleolus. Mol Biol Cell.
13:4100–4109. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schwab MS and Dreyer C: Protein
phosphorylation sites regulate the function of the bipartite NLS of
nucleolin. Eur J Cell Biol. 73:287–297. 1997.PubMed/NCBI
|
|
59
|
Shen EC, Henry MF, Weiss VH, Valentini SR,
Silver PA and Lee MS: Arginine methylation facilitates the nuclear
export of hnRNP proteins. Genes Dev. 12:679–691. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhao Y, Butler EB and Tan M: Targeting
cellular metabolism to improve cancer therapeutics. Cell Death Dis.
4:e5322013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hammoudi A, Song F, Reed KR, Jenkins RE,
Meniel VS, Watson AJ, Pritchard DM, Clarke AR and Jenkins JR:
Proteomic profiling of a mouse model of acute intestinal Apc
deletion leads to identification of potential novel biomarkers of
human colorectal cancer (CRC). Biochem Biophys Res Commun.
440:364–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Pichiorri F, Palmieri D, De Luca L,
Consiglio J, You J, Rocci A, Talabere T, Piovan C, Lagana A,
Cascione L, et al: In vivo NCL targeting affects breast cancer
aggressiveness through miRNA regulation. J Exp Med. 210:951–968.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Borer RA, Lehner CF, Eppenberger HM and
Nigg EA: Major nucleolar proteins shuttle between nucleus and
cytoplasm. Cell. 56:379–390. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Takagi M, Absalon MJ, McLure KG and Kastan
MB: Regulation of p53 translation and induction after DNA damage by
ribosomal protein L26 and nucleolin. Cell. 123:49–63. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen J, Guo K and Kastan MB: Interactions
of nucleolin and ribosomal protein L26 (RPL26) in translational
control of human p53 mRNA. J Biol Chem. 287:16467–16476. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Otake Y, Soundararajan S, Sengupta TK, Kio
EA, Smith JC, Pineda-Roman M, Stuart RK, Spicer EK and Fernandes
DJ: Overexpression of nucleolin in chronic lymphocytic leukemia
cells induces stabilization of bcl2 mRNA. Blood. 109:3069–3075.
2007. View Article : Google Scholar
|
|
67
|
Farin K, Di Segni A, Mor A and
Pinkas-Kramarski R: Structure-function analysis of nucleolin and
ErbB receptors interactions. PLoS One. 4:e61282009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Koutsioumpa M and Papadimitriou E: Cell
surface nucleolin as a target for anti-cancer therapies. Recent Pat
Anticancer Drug Discov. 9:137–152. 2014. View Article : Google Scholar
|
|
69
|
Watanabe T, Hirano K, Takahashi A,
Yamaguchi K, Beppu M, Fujiki H and Suganuma M: Nucleolin on the
cell surface as a new molecular target for gastric cancer
treatment. Biol Pharm Bull. 33:796–803. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wise JF, Berkova Z, Mathur R, Zhu H, Braun
FK, Tao RH, Sabichi AL, Ao X, Maeng H and Samaniego F: Nucleolin
inhibits Fas ligand binding and suppresses Fas-mediated apoptosis
in vivo via a surface nucleolin-Fas complex. Blood. 121:4729–4739.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Schokoroy S, Juster D, Kloog Y and
Pinkas-Kramarski R: Disrupting the oncogenic synergism between
nucleolin and Ras results in cell growth inhibition and cell death.
PLoS One. 8:e752692013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang B, Wang H, Jiang B, Liang P, Liu M,
Deng G and Xiao X: Nucleolin/C23 is a negative regulator of
hydrogen peroxide-induced apoptosis in HUVECs. Cell Stress
Chaperones. 15:249–257. 2010. View Article : Google Scholar :
|
|
73
|
Kirman DC, Renganathan B, Chui WK, Chen
MW, Kaya NA and Ge R: Cell surface nucleolin is a novel ADAMTS5
receptor mediating endothelial cell apoptosis. Cell Death Dis.
13:1722022. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Semenkovich CF, Ostlund RE Jr, Olson MO
and Yang JW: A protein partially expressed on the surface of HepG2
cells that binds lipoproteins specifically is nucleolin.
Biochemistry. 29:9708–9713. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Deng JS, Ballou B and Hofmeister JK:
Internalization of anti-nucleolin antibody into viable HEp-2 cells.
Mol Biol Rep. 23:191–195. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hovanessian AG, Puvion-Dutilleul F, Nisole
S, Svab J, Perret E, Deng JS and Krust B: The
cell-surface-expressed nucleolin is associated with the actin
cytoskeleton. Exp Cell Res. 261:312–328. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Fonseca NA, Rodrigues AS, Rodrigues-Santos
P, Alves V, Gregório AC, Valério-Fernandes Â, Gomes-da-Silva LC,
Rosa MS, Moura V, Ramalho-Santos J, et al: Nucleolin overexpression
in breast cancer cell sub-populations with different stem-like
phenotype enables targeted intracellular delivery of synergistic
drug combination. Biomaterials. 69:76–88. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen C, Chen L, Yao Y, Qin Z and Chen H:
Nucleolin overexpression is associated with an unfavorable outcome
for ependymoma: A multifactorial analysis of 176 patients. J
Neurooncol. 127:43–52. 2016. View Article : Google Scholar
|
|
79
|
Liu J, Wei T, Zhao J, Huang Y, Deng H,
Kumar A, Wang C, Liang Z, Ma X and Liang XJ: Multifunctional
aptamer-based nanoparticles for targeted drug delivery to
circumvent cancer resistance. Biomaterials. 91:44–56. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Mosafer J and Mokhtarzadeh A: Cell surface
nucleolin as a promising receptor for effective AS1411
aptamer-mediated targeted drug delivery into cancer cells. Curr
Drug Deliv. 15:1323–1329. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Ke J, Gu C, Zhang H, Liu Y, Zhang W, Rao
H, Li S and Wu F: Nucleolin promotes cisplatin resistance in
cervical cancer by the YB1-MDR1 pathway. J Oncol. 2021:99922182021.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Fu Z and Fenselau C: Proteomic evidence
for roles for nucleolin and poly[ADP-ribosyl] transferase in drug
resistance. J Proteome Res. 4:1583–1591. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Hu J, Chen Y, Wu Z, Wang L, Wen J, Jiang
P, Zhang Y and Lin M: Targeting nucleolin for reversal of
chemotherapy resistance in acute lymphoblastic leukemia. Blood.
134:50582019. View Article : Google Scholar
|
|
84
|
Cornelissen B, Waller A, Target C,
Kersemans V, Smart S and Vallis KA: 111In-BnDTPA-F3: An Auger
electron-emitting radiotherapeutic agent that targets nucleolin.
EJNMMI Res. 2:92012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Porkka K, Laakkonen P, Hoffman JA,
Bernasconi M and Ruoslahti E: A fragment of the HMGN2 protein homes
to the nuclei of tumor cells and tumor endothelial cells in vivo.
Proc Natl Acad Sci USA. 99:7444–7449. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Christian S, Pilch J, Akerman ME, Porkka
K, Laakkonen P and Ruoslahti E: Nucleolin expressed at the cell
surface is a marker of endothelial cells in angiogenic blood
vessels. J Cell Biol. 163:871–878. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Balça-Silva J, do Carmo A, Tão H, Rebelo
O, Barbosa M, Moura-Neto V, Sarmento-Ribeiro AB, Lopes MC and
Moreira JN: Nucleolin is expressed in patient-derived samples and
glioblastoma cells, enabling improved intracellular drug delivery
and cytotoxicity. Exp Cell Res. 370:68–77. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Moura V, Lacerda M, Figueiredo P, Corvo
ML, Cruz MEM, Soares R, de Lima MCP, Simoes S and Moreira JN:
Targeted and intracellular triggered delivery of therapeutics to
cancer cells and the tumor microenvironment: Impact on the
treatment of breast cancer. Breast Cancer Res Treat. 133:61–73.
2012. View Article : Google Scholar
|
|
89
|
Winer I, Wang S, Lee YEK, Fan W, Gong Y,
Burgos-Ojeda D, Spahlinger G, Kopelman R and Buckanovich RJ:
F3-targeted cisplatin-hydrogel nanoparticles as an effective
therapeutic that targets both murine and human ovarian tumor
endothelial cells in vivo. Cancer Res. 70:8674–8683. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Fonseca NA, Gomes-da-Silva LC, Moura V,
Simões S and Moreira JN: Simultaneous active intracellular delivery
of doxorubicin and C6-ceramide shifts the additive/antagonistic
drug interaction of non-encapsulated combination. J Control
Release. 196:122–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Reddy GR, Bhojani MS, McConville P, Moody
J, Moffat BA, Hall DE, Kim G, Koo YEL, Woolliscroft MJ, Sugai JV,
et al: Vascular targeted nanoparticles for imaging and treatment of
brain tumors. Clin Cancer Res. 12:6677–6686. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Drecoll E, Gaertner FC, Miederer M,
Blechert B, Vallon M, Müller JM, Alke A, Seidl C, Bruchertseifer F,
Morgenstern A, et al: Treatment of peritoneal carcinomatosis by
targeted delivery of the radio-labeled tumor homing peptide
213Bi-DTPA-[F3]2 into the nucleus of tumor
cells. PLoS One. 4:e57152009. View Article : Google Scholar
|
|
93
|
Brignole C, Bensa V, Fonseca NA, Del Zotto
G, Bruno S, Cruz AF, Malaguti F, Carlini B, Morandi F, Calarco E,
et al: Cell surface nucleolin represents a novel cellular target
for neuroblastoma therapy. J Exp Clin Cancer Res. 40:1802021.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cai Y, Xu Z, Shuai Q, Zhu F, Xu J, Gao X
and Sun X: Tumor-targeting peptide functionalized PEG-PLA micelles
for efficient drug delivery. Biomater Sci. 8:2274–2282. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chariou PL, Wang L, Desai C, Park J,
Robbins LK, von Recum HA, Ghiladi RA and Steinmetz NF: Let there be
light: Targeted photodynamic therapy using high aspect ratio plant
viral nanoparticles. Macromol Biosci. 19:18004072019. View Article : Google Scholar
|
|
96
|
Chen D, Yang D, Dougherty CA, Lu W, Wu H,
He X, Cai T, Van Dort ME, Ross BD and Hong H: In vivo targeting and
positron emission tomography imaging of tumor with intrinsically
radioactive metal-organic frameworks nanomaterials. ACS Nano.
11:4315–4327. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Cruz AF, Caleiras MB, Fonseca NA,
Gonçalves N, Mendes VM, Sampaio SF, Moura V, Melo JB, Almeida RD,
Manadas B, et al: The enhanced efficacy of intracellular delivery
of doxorubicin/C6-ceramide combination mediated by the F3
peptide/nucleolin system is supported by the downregulation of the
PI3K/Akt pathway. Cancers (Basel). 13:30522021. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Essler M, Gärtner FC, Neff F, Blechert B,
Senekowitsch-Schmidtke R, Bruchertseifer F, Morgenstern A and Seidl
C: Therapeutic efficacy and toxicity of 225Ac-labelled vs.
213Bi-labelled tumour-homing peptides in a preclinical mouse model
of peritoneal carcinomatosis. Eur J Nucl Med Mol Imaging.
39:602–612. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ferrara B, Belbekhouche S, Habert D,
Houppe C, Vallée B, Bourgoin-Voillard S, Cohen JL, Cascone I and
Courty J: Cell surface nucleolin as active bait for nanomedicine in
cancer therapy: A promising option. Nanotechnology. 32:3220012021.
View Article : Google Scholar
|
|
100
|
Gomes-da-Silva LC, Santos AO, Bimbo LM,
Moura V, Ramalho JS, Pedroso de Lima MC, Simões S and Moreira JN:
Toward a siRNA-containing nanoparticle targeted to breast cancer
cells and the tumor microenvironment. Int J Pharm. 434:9–19. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Hu Q, Gu G, Liu Z, Jiang M, Kang T, Miao
D, Tu Y, Pang Z, Song Q, Yao L, et al: F3 peptide-functionalized
PEG-PLA nanoparticles co-administrated with tLyp-1 peptide for
anti-glioma drug delivery. Biomaterials. 34:1135–1145. 2013.
View Article : Google Scholar
|
|
102
|
Karamchand L, Kim G, Wang S, Hah HJ, Ray
A, Jiddou R, Koo Lee YE, Philbert MA and Kopelman R: Modulation of
hydrogel nanoparticle intracellular trafficking by multivalent
surface engineering with tumor targeting peptide. Nanoscale.
5:10327–10344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Lam PYH, Hillyar CRT, Able S and Vallis
KA: Synthesis and evaluation of an 18 F-labeled
derivative of F3 for targeting surface-expressed nucleolin in
cancer and tumor endothelial cells. J Labelled Comp Radiopharm.
59:492–499. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Lopes R, Shi K, Fonseca NA, Gama A,
Ramalho JS, Almeida L, Moura V, Simões S, Tidor B and Moreira JN:
Modelling the impact of nucleolin expression level on the activity
of F3 peptide-targeted pH-sensitive pegylated liposomes containing
doxorubicin. Drug Deliv Transl Res. 12:629–646. 2022. View Article : Google Scholar
|
|
105
|
Mäkelä AR, Närvänen A and Oker-Blom C:
Peptide-mediated interference with baculovirus transduction. J
Biotechnol. 134:20–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Orringer DA, Koo YEL, Chen T, Kim G, Hah
HJ, Xu H, Wang S, Keep R, Philbert MA, Kopelman R and Sagher O: In
vitro characterization of a targeted, dye-loaded nanodevice for
intraoperative tumor delineation. Neurosurgery. 64:965–972. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Pesarrodona M, Sánchez-García L,
Seras-Franzoso J, Sánchez-Chardi A, Baltá-Foix R, Cámara-Sánchez P,
Gener P, Jara JJ, Pulido D, Serna N, et al: Engineering a
nanostructured nucleolin-binding peptide for intracellular drug
delivery in triple-negative breast cancer stem cells. ACS Appl
Mater Interfaces. 12:5381–5388. 2020. View Article : Google Scholar
|
|
108
|
Pozdniakova NV, Ryabaya OV, Semkina AS,
Skribitsky VA and Shevelev AB: Using ELP repeats as a scaffold for
de novo construction of gadolinium-binding domains within
multifunctional recombinant proteins for targeted delivery of
gadolinium to tumour cells. Int J Mol Sci. 23:32972022. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Prickett WM, Van Rite BD, Resasco DE and
Harrison RG: Vascular targeted single-walled carbon nanotubes for
near-infrared light therapy of cancer. Nanotechnology.
22:4551012011. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Qin M, Zong H and Kopelman R: Click
conjugation of peptide to hydrogel nanoparticles for tumor-targeted
drug delivery. Biomacromolecules. 15:3728–3734. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Romano S, Moura V, Simões S, Moreira JN
and Gonçalves J: Anticancer activity and antibody-dependent
cell-mediated cytotoxicity of novel anti-nucleolin antibodies. Sci
Rep. 8:74502018. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Valério-Fernandes Â, Fonseca NA, Gonçalves
N, Cruz AF, Pereira MI, Gregório AC, Moura V, Ladeirinha AF,
Alarcão A, Gonçalves J, et al: Nucleolin overexpression predicts
patient prognosis while providing a framework for targeted
therapeutic intervention in lung cancer. Cancers (Basel).
14:22172022. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Xu G, Qin M, Mukundan A, Siddiqui J,
Takada M, Vilar-Saavedra P, Tomlins SA, Kopelman R and Wang X:
Prostate cancer characterization by optical contrast enhanced
photoacoustics. Proc SPIE Int Soc Opt Eng.
9708:97080I2016.PubMed/NCBI
|
|
114
|
Yang J, Lu W, Xiao J, Zong Q, Xu H, Yin Y,
Hong H and Xu W: A positron emission tomography image-guidable
unimolecular micelle nanoplatform for cancer theranostic
applications. Acta Biomater. 79:306–316. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zeng Y, Xiao J, Cong Y, Liu J, He Y, Ross
BD, Xu H, Yin Y, Hong H and Xu W: PEGylated nanoscale metal-organic
frameworks for targeted cancer imaging and drug delivery. Bioconjug
Chem. 32:2195–2204. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhang H, Ingham ES, Gagnon MKJ, Mahakian
LM, Liu J, Foiret JL, Willmann JK and Ferrara KW: In vitro
characterization and in vivo ultrasound molecular imaging of
nucleolin-targeted microbubbles. Biomaterials. 118:63–73. 2017.
View Article : Google Scholar :
|
|
117
|
Zhang Y, Yang M, Park JH, Singelyn J, Ma
H, Sailor MJ, Ruoslahti E, Ozkan M and Ozkan C: A surface-charge
study on cellular-uptake behavior of F3-peptide-conjugated iron
oxide nanoparticles. Small. 5:1990–1996. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Henke E, Perk J, Vider J, De Candia P,
Chin Y, Solit DB, Ponomarev V, Cartegni L, Manova K, Rosen N and
Benezra R: Peptide-conjugated antisense oligonucleotides for
targeted inhibition of a transcriptional regulator in vivo. Nat
Biotechnol. 26:91–100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Ireson CR and Kelland LR: Discovery and
development of anticancer aptamers. Mol Cancer Ther. 5:2957–2962.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Morita Y, Leslie M, Kameyama H, Volk DE
and Tanaka T: Aptamer therapeutics in cancer: Current and future.
Cancers (Basel). 10:802018. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Gomes-da-Silva LC, Ramalho JS, Pedroso de
Lima MC, Simões S and Moreira JN: Impact of anti-PLK1
siRNA-containing F3-targeted liposomes on the viability of both
cancer and endothelial cells. Eur J Pharm Biopharm. 85:356–364.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Drecoll E, Gaertner FC, Miederer M,
Blechert B, Vallon M, Müller JM, Alke A, Seidl C, Bruchertseifer F,
Morgenstern A, et al: Treatment of peritoneal carcinomatosis by
targeted delivery of the radio-labeled tumor homing peptide
bi-DTPA-[F3]2 into the nucleus of tumor cells. PLoS One.
4:e57152009. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ayatollahi S, Salmasi Z, Hashemi M,
Askarian S, Oskuee RK, Abnous K and Ramezani M: Aptamer-targeted
delivery of Bcl-xL shRNA using alkyl modified PAMAM dendrimers into
lung cancer cells. Int J Biochem Cell Biol. 92:210–217. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Tong X, Ga L, Ai J and Wang Y: Progress in
cancer drug delivery based on AS1411 oriented nanomaterials. J
Nanobiotechnology. 20:572022. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Trinh TL, Zhu G, Xiao X, Puszyk W, Sefah
K, Wu Q, Tan W and Liu C: A Synthetic aptamer-drug adduct for
targeted liver cancer therapy. PLoS One. 10:e01366732015.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lohlamoh W, Soontornworajit B and Rotkrua
P: Anti-proliferative effect of doxorubicin-loaded AS1411 aptamer
on colorectal cancer cell. Asian Pac J Cancer Prev. 22:2209–2219.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Cheng Y, Zhao G, Zhang S, Nigim F, Zhou G,
Yu Z, Song Y, Chen Y and Li Y: AS1411-induced growth inhibition of
glioma cells by up-regulation of p53 and down-regulation of Bcl-2
and Akt1 via nucleolin. PLoS One. 11:e01670942016. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Ishimaru D, Zuraw L, Ramalingam S,
Sengupta TK, Bandyopadhyay S, Reuben A, Fernandes DJ and Spicer EK:
Mechanism of regulation of bcl-2 mRNA by nucleolin and A+ U-rich
element-binding factor 1 (AUF1). J Biol Chem. 285:27182–27191.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Wu J, Song C, Jiang C, Shen X, Qiao Q and
Hu Y: Nucleolin targeting AS1411 modified protein nanoparticle for
antitumor drugs delivery. Mol Pharm. 10:3555–3563. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Ai J, Xu Y, Lou B, Li D and Wang E:
Multifunctional AS1411-functionalized fluorescent gold
nanoparticles for targeted cancer cell imaging and efficient
photodynamic therapy. Talanta. 118:54–60. 2014. View Article : Google Scholar
|
|
131
|
Mongelard F and Bouvet P: AS-1411, a
guanosine-rich oligonucleotide aptamer targeting nucleolin for the
potential treatment of cancer, including acute myeloid leukemia.
Curr Opin Mol Ther. 12:107–114. 2010.PubMed/NCBI
|
|
132
|
Soundararajan S, Wang L, Sridharan V, Chen
W, Courtenay-Luck N, Jones D, Spicer EK and Fernandes DJ: Plasma
membrane nucleolin is a receptor for the anticancer aptamer AS1411
in MV4-11 leukemia cells. Mol Pharmacol. 76:984–991. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Reyes-Reyes EM, Teng Y and Bates PJ: A new
paradigm for aptamer therapeutic AS1411 action: Uptake by
macropinocytosis and its stimulation by a nucleolin-dependent
mechanism. Cancer Res. 70:8617–8629. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Laber DA, Sharma VR, Bhupalam L, Taft B,
Hendler FJ and Barnhart KM: Update on the first phase I study of
AGRO100 in advanced cancer. J Clin Oncol. 23(Suppl): S30642005.
View Article : Google Scholar
|
|
135
|
Storck S, Shukla M, Dimitrov S and Bouvet
P: Functions of the histone chaperone nucleolin in diseases.
Subcell Biochem. 41:125–144. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Stuart RK and Acton G: Relapsed and
refractory acute myeloid leukemia (AML) treated with AS1411 and
cytarabine: A randomized phase II trial. Blood. 112:19352008.
View Article : Google Scholar
|
|
137
|
Rizzieri D, Stockerl-Goldstein K, Wei A,
Herzig RH, Erlandsson F and Stuart RK: Long-term outcomes of
responders in a randomized, controlled phase II trial of aptamer
AS1411 in AML. J Clin Oncol. 28(Suppl): S65572010. View Article : Google Scholar
|
|
138
|
Stuart R, Stockerl-Goldstein K, Cooper M,
Devetten M, Herzig R, Medeiros B, Schiller G, Wei A, Acton G and
Rizzieri D: Randomized phase II trial of the nucleolin targeting
aptamer AS1411 combined with high-dose cytarabine in
relapsed/refractory acute myeloid leukemia (AML). J Clin Oncol.
27(Suppl): S70192009. View Article : Google Scholar
|
|
139
|
Rosenberg JE, Bambury RM, Van Allen EM,
Drabkin HA, Lara PN Jr, Harzstark AL, Wagle N, Figlin RA, Smith GW,
Garraway LA, et al: A phase II trial of AS1411 (a novel
nucleolin-targeted DNA aptamer) in metastatic renal cell carcinoma.
Invest New Drugs. 32:178–187. 2014. View Article : Google Scholar
|
|
140
|
Medicine NLo: Phase I open label study of
AS1411 in advanced solid tumours. https://clinicaltrials.gov/ct2/show/NCT00881244.
Accessed June 3, 2022
|
|
141
|
Medicine NLo: An open label randomized
controlled dose escalating phase II study of AS1411 combined with
cytarabine in the treatment of patients with primary refractory or
relapsed acute myeloid leukemia. https://clinicaltrials.gov/ct2/show/NCT00512083.
Accessed June 3, 2022
|
|
142
|
Medicine NLo: A phase II, open label,
single arm study of AS1411 in patients with metastatic renal cell
carcinoma. https://clinicaltrials.gov/ct2/show/NCT00740441.
Accessed June 3, 2022
|
|
143
|
Medicine NLo: An open-label randomized
controlled phase II study of AS1411 combined with cytarabine in the
treatment of patients with primary refractory or relapsed acute
myeloid leukemia. https://clinicaltrials.gov/ct2/show/NCT01034410.
Accessed June 3, 2022
|
|
144
|
Malik MT, O'Toole MG, Casson LK, Thomas
SD, Bardi GT, Reyes-Reyes EM, Ng CK, Kang KA and Bates PJ:
AS1411-conjugated gold nanospheres and their potential for breast
cancer therapy. Oncotarget. 6:22270–22281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Alibolandi M, Ramezani M, Abnous K and
Hadizadeh F: AS1411 aptamer-decorated biodegradable polyethylene
glycol-poly(lactic-co-glycolic acid) nanopolymersomes for the
targeted delivery of gemcitabine to non-small cell lung cancer in
vitro. J Pharm Sci. 105:1741–1750. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Lale SV, R G A, Aravind A, Kumar DS and
Koul V: AS1411 aptamer and folic acid functionalized pH-responsive
ATRP fabricated pPEGMA-PCL-pPEGMA polymeric nanoparticles for
targeted drug delivery in cancer therapy. Biomacromolecules.
15:1737–1752. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Teng Y, Girvan AC, Casson LK, Pierce WM
Jr, Qian M, Thomas SD and Bates PJ: AS1411 alters the localization
of a complex containing protein arginine methyltransferase 5 and
nucleolin. Cancer Res. 67:10491–10500. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Abdelmohsen K and Gorospe M: RNA-binding
protein nucleolin in disease. RNA Biol. 9:799–808. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Vigneron N: Human tumor antigens and
cancer immunotherapy. Biomed Res Int. 2015:9485012015. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Decker WK and Safdar A: Bioimmunoadjuvants
for the treatment of neoplastic and infectious disease: Coley's
legacy revisited. Cytokine Growth Factor Rev. 20:271–281. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Wojtukiewicz MZ, Rek MM, Karpowicz K,
Górska M, Polityńska B, Wojtukiewicz AM, Moniuszko M, Radziwon P,
Tucker SC and Honn KV: Inhibitors of immune checkpoints-PD-1,
PD-L1, CTLA-4-new opportunities for cancer patients and a new
challenge for internists and general practitioners. Cancer
Metastasis Rev. 40:949–982. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Wang QT, Nie Y, Sun SN, Lin T, Han RJ,
Jiang J, Li Z, Li JQ, Xiao YP, Fan YY, et al: Tumor-associated
antigen-based personalized dendritic cell vaccine in solid tumor
patients. Cancer Immunol Immunother. 69:1375–1387. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Chen CM, Chiang SY and Yeh NH: Increased
stability of nucleolin in proliferating cells by inhibition of its
self-cleaving activity. J Biol Chem. 266:7754–7758. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Fogal V, Sugahara KN, Ruoslahti E and
Christian S: Cell surface nucleolin antagonist causes endothelial
cell apoptosis and normalization of tumor vasculature.
Angiogenesis. 12:91–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Fernandes DJ and Tholanikunnel BG:
Abstract 1488: Development of anti-nucleolin antibodies with broad
spectrum anticancer activity and negligible toxicity to normal
cells. Cancer Res. 76(14 Suppl): S14882016. View Article : Google Scholar
|
|
156
|
De Greve H, Virdi V, Bakshi S and Depicker
A: Simplified monomeric VHH-Fc antibodies provide new opportunities
for passive immunization. Curr Opin Biotechnol. 61:96–101. 2020.
View Article : Google Scholar
|
|
157
|
Callebaut C, Blanco J, Benkirane N, Krust
B, Jacotot E, Guichard G, Seddiki N, Svab J, Dam E, Muller S, et
al: Identification of V3 loop-binding proteins as potential
receptors implicated in the binding of HIV particles to CD4(+)
cells. J Biol Chem. 273:21988–21997. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Nisole S, Krust B, Callebaut C, Guichard
G, Muller S, Briand JP and Hovanessian AG: The anti-HIV
pseudopeptide HB-19 forms a complex with the cell-surface-expressed
nucleolin independent of heparan sulfate proteoglycans. J Biol
Chem. 274:27875–27884. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Nisole S, Krust B, Dam E, Bianco A,
Seddiki N, Loaec S, Callebaut C, Guichard G, Muller S, Briand JP
and Hovanessian AG: The HB-19 pseudopeptide 5[Kpsi(CH2N) PR]-TASP
inhibits attachment of T lymophocyte- and macrophage-tropic HIV to
permissive cells. AIDS Res Hum Retroviruses. 16:237–249. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Nisole S, Said EA, Mische C, Prevost MC,
Krust B, Bouvet P, Bianco A, Briand JP and Hovanessian AG: The
anti-HIV pentameric pseudopeptide HB-19 binds the C-terminal end of
nucleolin and prevents anchorage of virus particles in the plasma
membrane of target cells. J Biol Chem. 277:20877–20886. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Said EA, Krust B, Nisole S, Svab J, Briand
JP and Hovanessian AG: The anti-HIV cytokine midkine binds the cell
surface-expressed nucleolin as a low affinity receptor. J Biol
Chem. 277:37492–37502. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Legrand D, Vigié K, Said EA, Elass E,
Masson M, Slomianny MC, Carpentier M, Briand JP, Mazurier J and
Hovanessian AG: Surface nucleolin participates in both the binding
and endocytosis of lactoferrin in target cells. Eur J Biochem.
271:303–317. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Said EA, Courty J, Svab J, Delbé J, Krust
B and Hovanessian AG: Pleiotrophin inhibits HIV infection by
binding the cell surface-expressed nucleolin. FEBS J.
272:4646–4659. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Hovanessian AG: Midkine, a cytokine that
inhibits HIV infection by binding to the cell surface expressed
nucleolin. Cell Res. 16:174–181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Barel M, Hovanessian AG, Meibom K, Briand
JP, Dupuis M and Charbit A: A novel receptor-ligand pathway for
entry of Francisella tularensis in monocyte-like THP-1 cells:
Interaction between surface nucleolin and bacteria elongation
factor Tu. BMC Microbiol. 8:1452008. View Article : Google Scholar
|
|
166
|
Losfeld ME, Khoury DE, Mariot P,
Carpentier M, Krust B, Briand JP, Mazurier J, Hovanessian AG and
Legrand D: The cell surface expressed nucleolin is a glycoprotein
that triggers calcium entry into mammalian cells. Exp Cell Res.
315:357–369. 2009. View Article : Google Scholar
|
|
167
|
El Khoury D, Destouches D, Lengagne R,
Krust B, Hamma-Kourbali Y, Garcette M, Niro S, Kato M, Briand JP,
Courty J, et al: Targeting surface nucleolin with a multivalent
pseudopeptide delays development of spontaneous melanoma in RET
transgenic mice. BMC Cancer. 10:3252010. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Destouches D, Page N, Hamma-Kourbali Y,
Machi V, Chaloin O, Frechault S, Birmpas C, Katsoris P, Beyrath J,
Albanese P, et al: A simple approach to cancer therapy afforded by
multivalent pseudopeptides that target cell-surface nucleoproteins.
Cancer Res. 71:3296–3305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Krust B, El Khoury D, Soundaramourty C,
Nondier I and Hovanessian AG: Suppression of tumorigenicity of
rhabdoid tumor derived G401 cells by the multivalent HB-19
pseudopeptide that targets surface nucleolin. Biochimie.
93:426–433. 2011. View Article : Google Scholar
|
|
170
|
Birmpas C, Briand JP, Courty J and
Katsoris P: Nucleolin mediates the antiangiogenesis effect of the
pseudopeptide N6L. BMC Cell Biol. 13:322012. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Birmpas C, Briand JP, Courty J and
Katsoris P: The pseudopeptide HB-19 binds to cell surface nucleolin
and inhibits angiogenesis. Vasc Cell. 4:212012. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Destouches D, Huet E, Sader M, Frechault
S, Carpentier G, Ayoul F, Briand JP, Menashi S and Courty J:
Multivalent pseudopeptides targeting cell surface nucleoproteins
inhibit cancer cell invasion through tissue inhibitor of
metalloproteinases 3 (TIMP-3) release. J Biol Chem.
287:43685–43693. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Koutsioumpa M, Polytarchou C, Courty J,
Zhang Y, Kieffer N, Mikelis C, Skandalis SS, Hellman U, Iliopoulos
D and Papadimitriou E: Interplay between αvβ3 integrin and
nucleolin regulates human endothelial and glioma cell migration. J
Biol Chem. 288:343–354. 2013. View Article : Google Scholar
|
|
174
|
Benedetti E, Antonosante A, d'Angelo M,
Cristiano L, Galzio R, Destouches D, Florio TM, Dhez AC, Astarita
C, Cinque B, et al: Nucleolin antagonist triggers autophagic cell
death in human glioblastoma primary cells and decreased in vivo
tumor growth in orthotopic brain tumor model. Oncotarget.
6:420912015. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Diamantopoulou Z, Gilles ME, Sader M,
Cossutta M, Vallée B, Houppe C, Habert D, Brissault B, Leroy E,
Maione F, et al: Multivalent cationic pseudopeptide polyplexes as a
tool for cancer therapy. Oncotarget. 8:90108–90122. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
De Cola A, Franceschini M, Di Matteo A,
Colotti G, Celani R, Clemente E, Ippoliti R, Cimini A, Dhez A,
Vallée B, et al: N6L pseudopeptide interferes with nucleophosmin
protein-protein interactions and sensitizes leukemic cells to
chemotherapy. Cancer Lett. 412:272–282. 2018. View Article : Google Scholar
|
|
177
|
Dhez AC, Benedetti E, Antonosante A,
Panella G, Ranieri B, Florio TM, Cristiano L, Angelucci F,
Giansanti F, Di Leandro L, et al: Targeted therapy of human
glioblastoma via delivery of a toxin through a peptide directed to
cell surface nucleolin. J Cell Physiol. 233:4091–4105. 2018.
View Article : Google Scholar
|
|
178
|
Darche M, Cossutta M, Caruana L, Houppe C,
Gilles ME, Habert D, Guilloneau X, Vignaud L, Paques M, Courty J
and Cascone I: Antagonist of nucleolin, N6L, inhibits
neovascularization in mouse models of retinopathies. FASEB J.
34:5851–5862. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Chalabi-Dchar M, Cruz E, Mertani HC, Diaz
JJ, Courty J, Cascone I and Bouvet P: Nucleolin aptamer N6L
reprograms the translational machinery and acts synergistically
with mTORi to inhibit pancreatic cancer proliferation. Cancers
(Basel). 13:49572021. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Lamprou M, Koutsioumpa M, Kaspiris A,
Zompra K, Tselios T and Papadimitriou E: Binding of pleiotrophin to
cell surface nucleolin mediates prostate cancer cell adhesion to
osteoblasts. Tissue Cell. 76:1018012022. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Thongchot S, Jirapongwattana N,
Luangwattananun P, Chiraphapphaiboon W, Chuangchot N, Sa-Nguanraksa
D, O-Charoenrat P, Thuwajit P, Yenchitsomanus PT and Thuwajit C:
Adoptive transfer of anti-nucleolin T cells combined with PD-L1
inhibition against triple-negative breast cancer. Mol Cancer Ther.
21:727–739. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Pincha M, Sundarasetty BS, Salguero G,
Gutzmer R, Garritsen H, Macke L, Schneider A, Lenz D, Figueiredo C,
Blasczyk R, et al: Identity, potency, in vivo viability, and
scaling up production of lentiviral vector-induced dendritic cells
for melanoma immunotherapy. Hum Gene Ther Methods. 23:38–55. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Nakayama M: Antigen presentation by
MHC-dressed cells. Front Immunol. 5:6722015. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Mangare C, Tischer-Zimmermann S, Riese SB,
Dragon AC, Prinz I, Blasczyk R, Maecker-Kolhoff B and Eiz-Vesper B:
Robust identification of suitable T-cell subsets for personalized
CMV-specific T-cell immunotherapy using CD45RA and CD62L
microbeads. Int J Mol Sci. 20:14152019. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Li ZL, Zhang HL, Huang Y, Huang JH, Sun P,
Zhou NN, Chen YH, Mai J, Wang Y, Yu Y, et al: Autophagy deficiency
promotes triple-negative breast cancer resistance to T
cell-mediated cytotoxicity by blocking tenascin-C degradation. Nat
Commun. 11:38062020. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Gurung S, Khan F, Gunassekaran GR, Yoo JD,
Poongkavithai Vadevoo SM, Permpoon U, Kim SH, Kim HJ, Kim IS, Han
H, et al: Phage display-identified PD-L1-binding peptides
reinvigorate T-cell activity and inhibit tumor progression.
Biomaterials. 247:1199842020. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Liao F, Liu L, Luo E and Hu J: Curcumin
enhances anti-tumor immune response in tongue squamous cell
carcinoma. Arch Oral Biol. 92:32–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Zhang J, Dang F, Ren J and Wei W:
Biochemical aspects of PD-L1 regulation in cancer immunotherapy.
Trends Biochem Sci. 43:1014–1032. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Wang X, Wu WKK, Gao J, Li Z, Dong B, Lin
X, Li Y, Li Y, Gong J, Qi C, et al: Autophagy inhibition enhances
PD-L1 expression in gastric cancer. J Exp Clin Cancer Res.
38:1402019. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Abreu TR, Godinho-Santos A, Moreira J and
Gonçalves J: P06. 02 Anti-nucleolin CAR (chimeric antigen
receptor)-T cell targeting triple-negative breast cancer-critical
parameters in CAR-T cells generation. J Immunother Cancer. 10(Suppl
1): A232022.
|
|
191
|
Vigneron N, Stroobant V, Van den Eynde BJ
and van der Bruggen P: Database of T cell-defined human tumor
antigens: The 2013 update. Cancer Immun. 13:152013.PubMed/NCBI
|
|
192
|
Wu D, Gao Y, Qi Y, Chen L, Ma Y and Li Y:
Peptide-based cancer therapy: opportunity and challenge. Cancer
Lett. 351:13–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Kim JH, Bae C, Kim MJ, Song IH, Ryu JH,
Choi JH, Lee CJ, Nam JS and Kim JI: A novel nucleolin-binding
peptide for cancer theranostics. Theranostics. 10:9153–9171. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Shadidi M and Sioud M: Selective targeting
of cancer cells using synthetic peptides. Drug Resist Updat.
6:363–371. 2003. View Article : Google Scholar
|
|
196
|
Knutson KL, Schiffman K and Disis ML:
Immunization with a HER-2/neu helper peptide vaccine generates
HER-2/neu CD8 T-cell immunity in cancer patients. J Clin Invest.
107:477–484. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Tobias J, Garner-Spitzer E, Drinić M and
Wiedermann U: Vaccination against Her-2/neu, with focus on
peptide-based vaccines. ESMO Open. 7:1003612022. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Carmichael MG, Benavides LC, Holmes JP,
Gates JD, Mittendorf EA, Ponniah S and Peoples GE: Results of the
first phase 1 clinical trial of the HER-2/neu peptide (GP2) vaccine
in disease-free breast cancer patients: United States military
cancer institute clinical trials group study I-04. Cancer.
116:292–301. 2010. View Article : Google Scholar
|
|
199
|
Abbaspour M and Akbari V: Cancer vaccines
as a targeted immunotherapy approach for breast cancer: An update
of clinical evidence. Expert Rev Vaccines. 21:337–353. 2022.
View Article : Google Scholar
|
|
200
|
Mitchell MS, Lund TA, Sewell AK, Marincola
FM, Paul E, Schroder K, Wilson DB and Kan-Mitchell J: The cytotoxic
T cell response to peptide analogs of the HLA-A*0201-restricted
MUC1 signal sequence epitope, M1.2. Cancer Immunol Immunother.
56:287–301. 2007. View Article : Google Scholar
|
|
201
|
Thueng-in K, Thanongsaksrikul J,
Jittavisutthikul S, Seesuay W, Chulanetra M, Sakolvaree Y,
Srimanote P and Chaicumpa W: Interference of HCV replication by
cell penetrable human monoclonal scFv specific to NS5B polymerase.
MAbs. 6:1327–1339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Medicine NLo: A non-randomized,
open-label, multi-centric dose-finding adaptive phase I/IIa study
to assess safety, tolerability, pharmacokinetics and preliminary
efficacy of repeated intravenous IPP-204106N administrations in
adult patients with advanced solid tumors. Available online:
https://clinicaltrials.gov/ct2/show/NCT01711398.
Accessed June 3, 2022
|
|
203
|
Ramos KS, Moore S, Runge I, Tavera-Garcia
MA, Cascone I, Courty J and Rayes-Reyes EM: The nucleolin
antagonist N6L inhibits LINE1 retrotransposon activity in non-small
cell lung carcinoma cells. J Cancer. 11:733–740. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Hovanessian AG, Soundaramourty C, El
Khoury D, Nondier I, Svab J and Krust B: Surface expressed
nucleolin is constantly induced in tumor cells to mediate
calcium-dependent ligand internalization. PLoS One. 5:e157872010.
View Article : Google Scholar
|