Introduction
Laryngeal squamous cell carcinoma (LSCC) is a common
type of head and neck cancer with an increasing incidence and
mortality rate (1,2). In 2020, there were 184,615 new LSCC
cases and 99,840 LSCC-related deaths (3). Early-stage LSCC is often curable.
However, up to 60% of patients are already in the advanced stages
of LSCC (clinical stages III or IV) at the time of diagnosis
(2). Despite notable
advancements in diagnostic and treatment strategies, the 5-year
survival rate of patients with LSCC has declined from 66 to 61%
over the past 40 years (4). This
decline is mainly attributed to the proneness of LSCC to local
invasion, cervical lymph node metastasis and the unclear mechanisms
of LSCC tumorigenesis or development. Therefore, understanding the
molecular regulatory mechanisms underlying the progression of LSCC
is crucial for its diagnosis, treatment and prognosis.
Fascin actin-bundling protein 1 (FSCN1), a globular
filamentous actin-binding protein, is abnormally overexpressed in
multiple types of human cancer, such as laryngeal (5), bladder (6) and breast cancer (7), and elevated FSCN1 levels have been
shown to be a hallmark of an aggressive clinical progression and a
poor prognosis (8-10). In vitro functional studies
using cancer cell lines have revealed that FSCN1 promotes cell
growth, migration, invasion and metastasis in various types of
cancer, such as oral cancer (11), osteosarcoma (12) and non-small cell lung cancer
(8,13). Immunohistochemical studies have
also shown that FSCN1 protein expression is associated with
aggressive clinical phenotypes, a poor prognosis and shorter
survival outcomes (14,15). In addition, FSCN1 has been
established to play a role in cancer cell epithelial-mesenchymal
transition (5,16), glycolysis (17) and ferroptosis (18). In a previous study by the
authors, it was found that FSCN1 expression was substantially
upregulated in LSCC tissues compared with adjacent normal tissues
(19). Furthermore, FSCN1
knockdown using specific small interfering RNA (siRNA) has been
shown to inhibit LSCC cell migration, invasion and growth (5). Therefore, it is necessary to
further investigate the regulatory role of FSCN1 in LSCC and to
elucidate the molecular mechanisms through which FSCN1 promotes the
progression of LSCC.
In the present study, a comprehensive transcriptome
analysis using cells in which FSCN1 was knocked down compared with
control FSCN1-expressing cells was performed. It was found that
FSCN1 knockdown affected the expression of numerous genes. Through
a series of analyses, FSCN1 was linked to multiple biological
functions, such as transcriptional regulation, response to
radiation, focal adhesion, extracellular matrix (ECM)-receptor
interaction and steroid biosynthesis. The crosstalk analysis with
FSCN1-interacting proteins and co-immunoprecipitation (co-IP)
validation revealed that 24-dehydrocholesterol reductase (DHCR24)
is a new molecule associated with FSCN1 in LSCC. Further studies
focusing on the DHCR24-FSCN1 interaction may provide valuable
insight into the molecular mechanisms underlying LSCC.
Materials and methods
Cells and cell culture
The FD-LSC-1 human LSCC cell line [a gift from
Professor Liang Zhou (20)] was
cultured in BEGM™ Bronchial Epithelial Cell Growth Medium (Lonza
Group Ltd.) supplemented with 10% fetal bovine serum (Biological
Industries). 293T cells (China Center for Type Culture Collection)
and the TU-177 human LSCC cell line (Shanghai Bioleaf Biotech Co.,
Ltd.) were cultured in DMEM supplemented with 10% fetal bovine
serum. The cultures were maintained at 37°C in 5%
CO2.
Plasmid construction and cell
transfection
Human prostaglandin reductase 1 (PTGR1), DHCR24 and
solute carrier family 38 member 2 (SLC38A2) overexpression plasmids
were generated by inserting their respective CDS sequence into the
p3xFLAG-CMV-10 vector (Merck KGaA). The FSCN1 overexpression
plasmid was generated by inserting the FSCN1 coding sequence into
the pCMV-HA vector (Clontech; Takara Bio USA, Inc.). Empty
p3xFLAG-CMV-10 vector was used as a negative control. For DHCR24
overexpression, FD-LSC-1 and TU-177 cells were seeded in six-well
plates at a density of 5×105 cells/well. A total of 5
μl Lipofectamine 3000® (Invitrogen; Thermo Fisher
Scientific, Inc.) and 2.5 μg DHCR24 overexpression vector or
empty p3xFLAG-CMV-10 vector were mixed in 250 μl Opti-MEM™
(Gibco; Thermo Fisher Scientific, Inc.) and added to each well.
Following 6 h of incubation at 37°C with 5% CO2, the
medium was replaced with fresh medium. At 48 h following
transfection, the cells were collected for use in subsequent
experiments.
Small interfering RNA(RNA)
siRNA targeting FSCN1 (si-FSCN1) and scrambled siRNA
(si-NC) were synthesized by GenePharma Co., Ltd. The siRNA
sequences were as follows: si-FSCN1 sense
5′-GCAAGAAUGCCAGCUGCUACU-3′ and antisense
5′-AGUAGCAGCUGGCAUUCUUGC-3′; and si-NC sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. For transfection, TU-177 cells were
seeded in six-well plates at a density of 5×105
cells/well. A total of 5 μl Lipofectamine 3000®
(Invitrogen; Thermo Fisher Scientific, Inc.) and 100 nM siRNAs were
mixed in 250 μl Opti-MEM™ (Gibco; Thermo Fisher Scientific,
Inc.) and added to each well. Following 6 h of incubation at 37°C
with 5% CO2, the medium was replaced with fresh medium.
At 48 h following transfection, the cells were collected for use in
subsequent experiments.
Microarray assay and data analysis
TU-177 cells in which FSCN1 was knocked down and
control TU-177 cells were generated using siRNA transfection,
followed by total RNA extraction using TRIzol® reagent
(Thermo Fisher Scientific, Inc.). Total RNA was quantified using a
NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific,
Inc.), and the RNA integrity was assessed using Agilent Bioanalyzer
2100 (Agilent Technologies, Inc.). Sample labeling, microarray
hybridization and washing were performed according to the
manufacturer's instructions (Agilent Human lncRNA Microarray 2018
Version, 4×180k, Design ID: 085630). Briefly, total RNA was
transcribed into double-stranded cDNA (RNA Spike In Kit, one-color,
Agilent p/n 5188-5282; Agilent Technologies, Inc.), synthesized
into cRNA (RNA Spike In Kit, one-color, Agilent p/n 5188-5282;
Agilent Technologies, Inc.), and labeled with Cyanine-3-CTP (Low
Input Quick-Amp Labeling Kit, one-color. Agilent p/n 5190-2305;
Agilent Technologies, Inc.). The labeled cRNAs were hybridized onto
the microarray. Following washing, the arrays were scanned using
the Agilent Scanner G2505C (Agilent Technologies, Inc.).
Feature Extraction software (version 10.7.1.1,
Agilent Technologies) was used to analyze array images to obtain
raw data, and GeneSpring (version 13.1, Agilent Technologies) was
then used to complete the basic analysis of the raw data. First,
the raw data were normalized using the quantile algorithm. The
probes that at least 1 out of 2 conditions have flags in 'P' were
selected for further data analysis. Differentially expressed genes
(DEGs) or long non-coding RNAs (lncRNAs) were identified using fold
change (FC) and the P-value from the unpaired t-test. The
thresholds for up- and downregulated genes were a fold change of
≥1.5 and P≤0.05. Hierarchical clustering was performed to display
the expression patterns of distinguishable genes among samples. The
microarray assay data were deposited at the Gene Expression Omnibus
(GEO) database and are accessible via the accession no.
GSE255143.
Gene Ontology (GO) and Kyoto Encyclopedia
of Genes and Genomes Enrichment (KEGG) analysis
The Database for Annotation, Visualization and
Integrated Discovery (DAVID) Bioinformatics Resources (http://david.ncifcrf.gov) was used for functional
enrichment analysis of GO terms and KEGG pathways. P<0.05 was
selected as the cut-off criterion. The GO terms included biological
process, cellular component and molecular function. The top 20 GO
terms and KEGG pathways were plotted according to their
P-values.
Co-expression analysis
The RNA sequencing data of 57 pairs of LSCC and
matched adjacent normal mucosa tissues (GSE127165) were downloaded
from the GEO database. Of the 1,063 DEGs in the cells in which
FSCN1 was knocked down, 641 were expressed in the 57 LSCC tissues.
The co-expression between FSCN1 and the 641 DEGs was then
calculated using Pearson's correlation analysis, and Pearson's
correlation coefficients of <0.4 or >-0.4 were eliminated.
Both up- and downregulated DEGs that were positively and negatively
correlated with FSCN1, respectively, were eliminated.
Protein-protein interaction (PPI) network
analysis
The identified FSCN1-regulated genes were analyzed
using the Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING) online tool v11.5 (http://www.string-db.org) for interaction networks.
The PPI network was generated with a confidence score of ≥0.7 (high
confidence). The highly connected clusters from the PPI network
were identified using the 'Molecular Complex Detection (MCODE)'
plugin tool in Cytoscape v3.7.1 software. The top three high
ranking clusters were selected for further analysis.
Co-IP for western blot analysis
293T cells were cultured in 60-mm plates and each
plate was transfected with 5 μg FSCN1 overexpression vector
and 5 μg PTGR1 or DHCR24 or SLC38A2 overexpression vector
using Lipofectamine 3000® (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C with 5% CO2 for 6 h, according
to the manufacturer's instructions. At 48 h following transfection,
the cells were collected for use in subsequent co-IP experiments.
Whole-cell extracts were collected by lysing 1×107 cells
in Pierce IP Lysis buffer (Thermo Fisher Scientific, Inc.) with the
addition of a complete protease inhibitor mixture for 30 min on
ice, and clarified using centrifugation at 12,000 x g for 15 min at
4°C. For co-IP, protein extracts were incubated at 4°C overnight
with mouse monoclonal anti-flag antibody (1:500; cat. no. F1804;
Merck KGaA) or control mouse immunoglobulin G (IgG) (1:500; cat.no.
A7028; Beyotime Institute of Biotechnology), and precipitated
proteins were captured using Pierce Protein A/G magnetic beads
(Thermo Fisher Scientific, Inc.). Following three washes with
Pierce IP lysis buffer (Thermo Fisher Scientific, Inc.), bound
proteins were eluted in 2X SDS loading buffer and examined using
western blot analysis. Rabbit HA tag antibody (1:1,000; cat. no.
51064-2-AP; Proteintech Group, Inc.) was used for western blot
analysis.
Western blot analysis
The cells were lysed using Pierce RIPA lysis buffer
(Thermo Fisher Scientific, Inc.) containing protease inhibitor
cocktail for 30 min on ice. The protein concentration was
determined using a BCA Protein Assay kit (Thermo Fisher Scientific,
Inc.), and total protein was separated using 10% SDS-PAGE gels,
transferred to a PVDF membrane and blocked with 10% w/v non-fat
milk powder in TBST at room temperature for 2 h. The membranes were
incubated with primary antibodies against Flag (1:1,000; cat. no.
F1804; Merck KGaA), FSCN1 (1:1,000; cat. no. 54545S; Cell Signaling
Technology, Inc.), and GAPDH (1:1,000; cat.no. HC301-02; TransGen
Biotech Co., Ltd.)overnight at 4°C, followed by incubation with
appropriate horseradish peroxidase-conjugated secondary antibodies
(1:1,000; cat. no. A0216; Beyotime Institute of Biotechnology) at
room temperature for 2 h. Immunoreactive bands were visualized
using WesternBright® ECL HRP substrate (Advansta
Inc.).
Cell proliferation assay
Cell proliferation was determined using the Cell
Counting Kit-8 (TransGen Biotech Co., Ltd.), following the
manufacturer's instructions. Briefly, 3,000 cells were seeded into
each well of a 96-well plate. At 0, 12, 24, 48 and 72 h after
seeding, each well was replaced with 100 μl fresh complete
medium and 10 μl CCK-8 (TransGen Biotech Co., Ltd.) followed
by incubation at 37°C with 5% CO2 for 1 h. The
absorbance of the solution was measured at 450 nm using a Spectra
Max i3x Multifunctional microplate detection system (Molecular
Devices, LLC). A total of three independent experiments were
performed.
Cell migration assay
Following transfection, the cells were suspended in
a serum-free medium. Serum-free DMEM (200 μl) containing
1×105 cells were added to the upper chamber. A total of
500 μl DMEM medium supplemented with 20% fetal bovine serum
(Biological Industries) was then added to the lower chamber. At 48
h following incubation in 37°C with 5% CO2, the cells in
the upper chamber were removed with cotton swabs and the lower side
of the chamber was gently washed twice with phosphate-buffered
saline (PBS), and fixed with 4% paraformaldehyde for 20 min at room
temperature. The cells stained with 0.1% crystal violet (Amresco,
LLC) for 10 min at room temperature, and images were then captured
using an inverted microscope (Leica Microsystems GmbH).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR) analysis
Total RNA was isolated from the cells in which FSCN1
was knocked down or control cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), following the
manufacturer's instructions. For RT-qPCR of messenger RNA (mRNA),
cDNA was synthesized using the HiScript II 1st Strand cDNA
Synthesis kit (Vazyme Biotech Co., Ltd. according to the following
conditions: 25°C for 5 min, 50°C for 15 min, and 85°C for 2 min.
ChamQ SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd.) was used for
qPCR. The following thermocycling conditions were used: 95°C for 30
sec, followed by 40 cycles at 95°C for 5 sec and 60°C for 10 sec.
The relative expression level of target genes was calculated using
the 2(−ΔΔCq) method (21). The
primer sequences used are listed in Table SI.
Statistical analysis
Statistical analysis was performed using SPSS
software v22.0 (IBM Corp.). The data were analyzed using the
two-tailed Student's t-test; P<0.05 was considered to indicate a
statistically significant difference.
Results
Screening of DEGs in LSCC TU-177 cells in
which FSCN1 was knocked down
To investigate genes regulated by FSCN1, the
microarray analysis of differentially up- or downregulated genes
was performed by knocking down FSCN1 using siRNAs in TU-177 cells.
The results of RT-qPCR indicated that FSCN1 was successfully
knocked down (Fig. 1A).
Following data preprocessing, 1,063 DEGs (462 up- and 601
downregulated) were identified in the TU-177 cells transfected with
FSCN1 siRNAs compared with those transfected with the control under
the thresholds of FC>1.5 and P<0.05. Based on the volcano
plot and hierarchical clustering analysis of these DEGs, the
control and FSCN1 knockdown groups were clearly distinguished
(Fig. 1B and C). In addition,
436 differentially expressed lncRNAs (118 up- and 318
downregulated) and 198 differentially expressed circular RNAs
(circRNAs) (44 up- and 154 downregulated) were obtained between the
FSCN1 knockdown and control groups (Fig. 1D). These genes are listed in
Table SII. In addition, the
expression of the top 10 up- and downregulated genes in TU-177
cells in which FSCN1 was knocked down compared with the control
cells based on microarray data are shown in Fig. 1E and F.
 | Figure 1Screening of differentially expressed
genes using microarray profiling. (A) Validation of FSCN1
knockdown. TU-177 cells were transfected with FSCN1 (si-FSCN1) or
negative (si-NC) siRNAs for 48 h, and the expression level of FSCN1
was determined using RT-qPCR and western blot analysis. In the
RT-qPCR experiment, data are presented as the mean ± SD of three
independent experiments. ***P<0.001. (B) Volcano
plots of differentially expressed mRNAs. The location of DHCR24 is
marked. (C) Hierarchical clustering heatmap of differentially
expressed mRNAs. (D) Number of differentially expressed genes,
including mRNAs, lncRNAs and circRNAs. (E and F) Expression of the
top 10 up- and 10 downregulated genes in TU-177 cells in which
FSCN1 was knocked down compared with control TU-177 cells based on
microarray data (n=3). *P<0.05,
**P<0.01 and ***P<0.001. siRNA, small
interfering RNA; lncRNA, long non-coding RNA; mRNA, messenger RNA;
FSCN1, fascin actin-bundling protein 1; RT-qPCR, reverse
transcription-quantitative PCR; MUC20, mucin 20, cell surface
associated; SLC26A1, solute carrier family 26 member 1; ZNF85, zinc
finger protein 85; HLA-DRA, major histocompatibility complex, class
II, DR alpha; ZNF618, zinc finger protein 618; SLC22A7, solute
carrier family 22 member 7; DOCK2, dedicator of cytokinesis 2;
ACOX3, acyl-CoA oxidase 3, pristanoyl; PLD5, phospholipase D family
member 5; RNF165, ring finger protein 165; GABRD,
gamma-aminobutyric acid type A receptor delta subunit; PTPN,
protein tyrosine phosphatase, non-receptor type 3; ADPRM,
ADP-ribose/CDP-alcohol diphosphatase, manganese dependent; OSCAR,
osteoclast associated, immunoglobulin-like receptor; USP9Y,
ubiquitin specific peptidase 9, Y-linked; WISP1, WNT1 inducible
signaling pathway protein 1; CAGE1, cancer antigen 1; ZNF518A, zinc
finger protein 518A; DCT, dopachrome tautomerase; ZBBX, zinc finger
B-box domain containing; DHCR24, 24-dehydrocholesterol
reductase. |
Function annotation of differentially
expressed mRNAs
To identify the putative functions and pathways
associated with the FSCN1-regulated genes, functional enrichment
analysis was performed through the DAVID Bioinformatics Resources
(22). The upregulated DEGs were
significantly enriched with biological functions of 'cilium
assembly', 'regulation of transcription from RNA polymerase II
promoter' and 'regulation of transcription, DNA-templated'
(Fig. 2A). By contrast, the
downregulated DEGs were associated with 'nervous system
development', 'sterol biosynthetic process' and 'response to
radiation' (Fig. 2B). In the
cellular components category, 'cilium' and 'cytoplasm' were
significantly enriched using the upregulated DEGs, whereas
'integral component of membrane' and 'plasma membrane' were
involved with the downregulated DEGs (Fig. 2A and B). For the GO molecular
functions category, the upregulated DEGs were enriched with
functions, such as 'metal ion binding', 'RNA polymerase II core
promoter proximal region sequence-specific DNA binding' and 'RNA
polymerase II transcription factor activity, sequence-specific DNA
binding' (Fig. 2A). The
downregulated DEGs were associated with 'calcium ion binding',
'endopeptidase inhibitor activity' and 'oxidoreductase activity'
(Fig. 2B). According to the
findings of KEGG pathway analysis, the upregulated DEGs
demonstrated significant enrichment in the pathways of 'herpes
simplex virus 1 infection', 'focal adhesion' and 'ECM-receptor
interaction' (Fig. 2C). By
contrast, the downregulated DEGs were involved in 'steroid
biosynthesis', 'metabolic pathways' and 'biosynthesis of cofactors'
(Fig. 2D). The complete results
are presented in Table
SIII.
Construction of a network of genes
co-expressed with FSCN1 in LSCC
RNA sequencing (RNA-seq) was previously performed in
57 pairs of LSCC and matched adjacent normal mucosa tissues to
construct mRNA expression profiles (23). Based on the aforementioned
RNA-seq data and 1,063 DEGs in the cells in which FSCN1 was knocked
down, a network of genes co-expressed with FSCN1 in LSCC was
constructed using the flowchart illustrated in Fig. 3A. Finally, 48 DEGs (10 up- and 38
downregulated DEGs) were found to be co-expressed with FSCN1 in
LSCC with Pearson's correlation coefficients of 0.4. A network was
constructed and visualized using Cytoscape software (Fig. 3B). In addition, functional
enrichment analysis revealed that the 48 genes co-expressed with
FSCN1 were significantly associated with the functions of 'defense
response to virus', 'response to virus', 'cellular response to
interferon-α', 'innate immune response' and 'neutral amino acid
transport' (Fig. 3C). KEGG
pathway analysis demonstrated that 'biosynthesis pathways of
cofactors' and 'metabolic pathways' were significantly enriched by
genes co-expressed with FSCN1 (Fig.
3D). The complete results are presented in Table SIV.
PPI network of DEGs
To explore the interactions among these
FSCN1-regulated genes and identify specific functional complexes, a
PPI network was constructed using the STRING database. In the PPI
network, the minimum required interaction score was set as
>0.700 (high confidence), and disconnected nodes in the network
were hidden. A PPI network containing 455 nodes and 570 edges was
constructed (Fig. 4A). In
addition, three highly connected clusters were identified from the
PPI network using the MCODE plugin tool (24) in Cytoscape (Fig. 4B-D). The GO and KEGG enrichment
analyses revealed that Cluster 1 was mainly involved with functions
of defense response to virus (GO:0051607) and response to virus
(GO:0009615). By contrast, Cluster 2 was associated with the
function of cholesterol biosynthetic process (GO:0006695) and the
KEGG pathway of steroid biosynthesis (hsa00100). The genes in
Cluster 3 were mainly associated with cilium assembly
(GO:0060271).
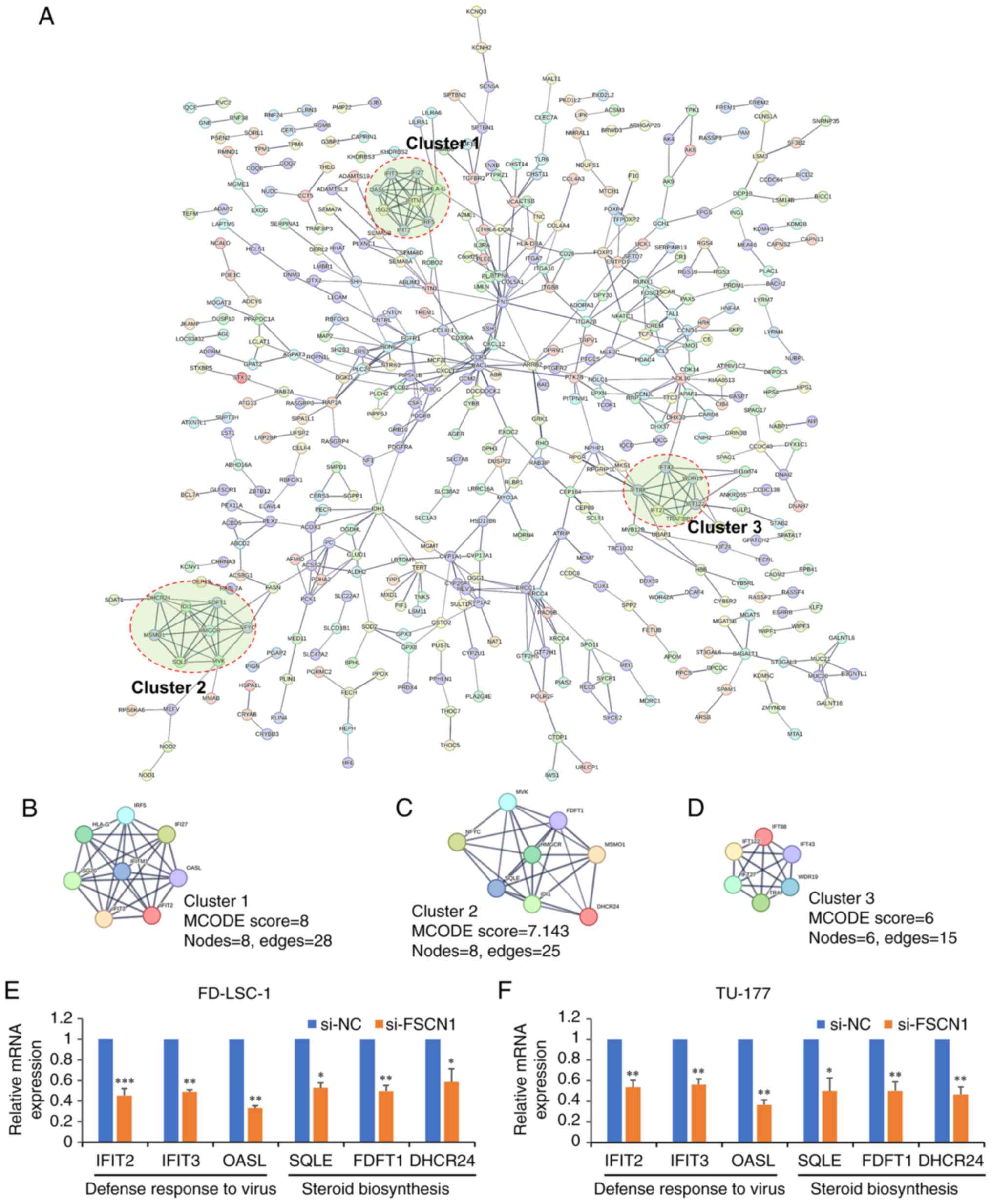 | Figure 4PPI network of DEGs in cells in which
FSCN1 was knockdown. (A) The PPI network obtained from the STRING
database with a confidence score of >0.7. The network contained
455 nodes and 570 edges. (B-D) The three tightly connected network
clusters obtained with MCODE were rendered as separate modules. (E
and F) Validation of FSCN1 affected functional or signaling pathway
molecules. (E) FD-LSC-1 and (F) TU-177 LSCC cells were transfected
with FSCN1 siRNAs or NC siRNAs for 48 h, and then the expression
level of genes involved in defense response to virus and steroid
biosynthesis the pathway was determined using RT-qPCR. In the
RT-qPCR experiment, data are presented as the mean ± SD of three
independent experiments. *P<0.05,
**P<0.01 and ***P<0.001. PPI,
protein-protein interaction; DEGs, differentially expressed genes;
FSCN1, fascin actin-bundling protein 1; MCODE, Molecular Complex
Detection; LSCC, laryngeal squamous cell carcinoma; siRNA, small
interfering RNA; RT-qPCR, reverse transcription-quantitative PCR;
IFIT2, tetratricopeptide repeats 2; IFIT3, tetratricopeptide
repeats 3; OASL, 2′-5′-oligoadenylate synthetase like; SQLE,
squalene epoxidase; FDFT1, farnesyl-diphosphate farnesyltransferase
1; DHCR24, 24-dehydrocholesterol reductase. |
Subsequently, RT-qPCR was used to validate the
expression of genes involved in the function of defense response to
virus and steroid biosynthesis pathway. Compared with the control
group, interferon induced protein with tetratricopeptide repeats
(IFIT)2, IFIT3, 2′-5′-oligoadenylate synthetase like (defense
response to virus; QASL), squalene epoxidase (SQLE),
farnesyl-diphosphate farnesyltransferase 1 (FDFT1) and DHCR24
(steroid biosynthesis) were significantly downregulated in the LSCC
cells in which FSCN1 was knocked down (Fig. 4E and F), indicating that FSCN1
affected these functions and pathways.
FSCN1-interacting proteins in LSCC and
crosstalk analysis
FSCN1-interacting proteins were previously
characterized in LSCC cell lines using a mass spectrometry-based
proteomics approach, and 238 proteins were identified as
interacting partners of FSCN1 in TU-177 cells (25). Following Venn analysis, seven
overlapped target genes [DHCR24, SLC38A2, PTGR1, peroxiredoxin 4
(PRDX4), tropomyosin 4 (TMP4), abhydrolase domain containing 16A
(ABHD16A), phospholipase and capping protein regulator and myosin 1
linker 1 (CARMIL1)] were identified (Fig. 5A). Of note, it was found that a
proven FSCN1-regulated gene (DHCR24; Fig. 4E and F), interacted with FSCN1 in
TU-177 cells. SLC38A2, a gene co-expressed with FSCN1 in LSCC
(Fig. 3B) interacted with FSCN1
in TU-177 cells. Furthermore, another DEG (PTGR1) in cells in which
FSCN1 was knocked down was identified as an FSCN1-interacting
protein in Hep-2 cells (25).
RT-qPCR was performed to verify the relative expression levels of
PTGR1 and SLC38A2 in LSCC cells in which FSCN1 was knocked down. As
shown in Fig. 5B, SLC38A2 was
significantly downregulated in LSCC cells in which FSCN1 was
knocked down, whereas the expression of PTGR1 was not significantly
altered.
 | Figure 5Crosstalk analysis and validation.
(A) Venn diagram confirmed 7 target genes that overlapped between
the DEGs in FSCN1-knockdown TU-177 cells and FSCN1 interacting
proteins in TU-177 cells. (B) Validation of the expression of FSCN1
regulated genes. FD-LSC-1 and TU-177 LSCC cells were transfected
with FSCN1 siRNAs or NC siRNAs for 48 h, and the expression level
of PTGR1 and SLC38A2 was then determined using RT-qPCR. In the
RT-qPCR experiment, data are presented as the mean ± SD of three
independent experiments (*P<0.05). (C) Validation of
the interactions between PTGR1, DHCR24 and SLC38A2 with FSCN1.
HA-tagged FSCN1 and Flag-tagged PTGR1, DHCR24 and SLC38A2 were
transiently co-expressed in 293T cells, respectively. For western
blot analysis, antibodies against Flag, HA and GAPDH were used to
detect protein expression. Total cell lysates were used to validate
the interaction by co-IP. Antibodies against Flag tag were used to
capture proteins-FSCN1 complexes, and normal mouse IgG served as a
negative control. On western blot analysis, rabbit anti-HA antibody
was used to detect FSCN1. DEGs, differentially expressed genes;
FSCN1, fascin actin-bundling protein 1; siRNAs, small interfering
RNA; LSCC, laryngeal squamous cell carcinoma; RT-qPCR, reverse
transcription-quantitative PCR; PTGR1, prostaglandin reductase 1;
DHCR24, 24-dehydrocholesterol reductase; SLC38A2, solute carrier
family 38 member 2; PRDX4, peroxiredoxin 4; TPM4, tropomyosin 4;
ABHD16A, abhydrolase domain containing 16A; CARMIL1, capping
protein regulator and myosin 1 linker 1. |
The present study then attempted to validate the
interaction between FSCN1, and PTGR1, DHCR24 and SLC38A2 through
co-IP. The three genes were cloned into vectors with a 3xFlag tag,
and the FSCN1 gene was cloned into vectors with an HA tag. FSCN1
and Flag-tagged proteins were transiently co-expressed in 293T
cells. The expression of all three Flag-tagged proteins and
HA-tagged FSCN1 was successfully confirmed using western blot
analysis (Fig. 5C). Co-IP was
performed on whole cell extracts with Flag-tag antibodies, with
matched normal IgG used as a negative control. Out of these three
proteins, two were confirmed to bind to FSCN1 (Fig. 5C), and one protein (SLC38A2) did
not interact with FSCN1.
Overexpression of DHCR24 promotes the
proliferation and migration of LSCC cells
As DHCR24 binds to the FSCN1 protein and its
expression is regulated by FSCN1, the expression levels of DHCR24
in the FD-LSC-1 and TU-177 LSCC cell lines were first investigated.
Compared with the normal cell line (293T), DHCR24 was significantly
downregulated in the FD-LSC-1 and TU-177 LSCC cell lines (Fig. 6A). To further investigate the
functional role of DHCR24 in LSCC cells, the DHCR24 overexpression
vector (OE-DHCR24) and its empty vector (OE-NC) were transfected
into FD-LSC-1 and TU-177 cells. The efficiency of DHCR24 protein
overexpression was further verified using western blot analysis
(Fig. 6B). Compared with the
negative control group, DHCR24 overexpression promoted LSCC cell
proliferation (Fig. 6C and D).
Furthermore, the results of Transwell assay demonstrated that
DHCR24 overexpression markedly promoted LSCC cell migration
(Fig. 6E). These findings
suggested that DHCR24 promotes the proliferation and migration of
LSCC cells.
Discussion
FSCN1 has been widely reported to be overexpressed
in several types of human cancer, such as laryngeal (5), bladder (6) and breast cancer (7), and its expression has been
associated with an aggressive clinical course, a poor prognosis and
shorter survival rates (8,10,26). Previous studies have indicated
that FSCN1 expression levels in LSCC tissues are significantly
higher than those in adjacent normal tissues, and that FSCN1
knockdown inhibits cell growth, migration and invasion in LSCC
(5,19). In addition, a high expression of
FSCN1 has been shown to be significantly associated with clinical
features and poor outcomes in LSCC (5,19,27,28). However, the regulatory mechanisms
of FSCN1 in LSCC remain unclear. In the present study, to gain more
insight into the function of FSCN1 in LSCC progression, microarray
analysis was performed to screen the DEGs in TU-177 LSCC cells in
which FSCN1 was knocked down. Following data preprocessing, 462
genes were upregulated and 601 genes were downregulated in the
FSCN1-knockdown cells relative to the control cells with a 1.5-FC
cut-off value. According to functional enrichment analysis, these
FSCN1-regulated genes were associated with gene transcription
(29-31), radioresistance (32) and focal adhesion (33,34), which was consistent with the
results of previous studies. Of note, to the best of our knowledge,
there is no evidence to date that FSCN1 plays a role in the sterol
biosynthetic process, ECM-receptor interaction and the steroid
biosynthesis pathway, suggesting that FSCN1 may be linked to
previously unknown functions.
To further understand the functional association
between these DEGs in TU-177 cells in which FSCN1 was knocked down
and FSCN1 in LSCC, a network of genes co-expressed with FSCN1 in
LSCC was constructed. The results revealed that 10 up- and 38
downregulated DEGs were found to be co-expressed with FSCN1 in LSCC
tissues. Furthermore, functional enrichment analysis and RT-qPCR
validation revealed that numerous genes co-expressed with FSCN1
were associated with the functions of the defense response to virus
and response to virus, indicating that FSCN1 may play a role in the
defense response to a virus. This result was also consistent with
the results of the PPI analysis.
Studies have reported that dysregulated cholesterol
metabolism is exhibited in cancer cells, with cholesterol and its
metabolites acting as signaling molecules that drive tumor
development (35,36). Although FSCN1 plays a role in
cancer progression, there is no evidence to date to indicate that
FSCN1 is associated with cholesterol metabolism. Herein, the
results of PPI analysis revealed another highly connected cluster
(Cluster 2, Fig. 4C), in which
several FSCN1-regulated genes were associated with the cholesterol
biosynthetic process and steroid biosynthesis pathway. For example,
SQLE is a key rate-limiting enzyme that catalyzes the conversion of
squalene to 2,3-epoxysqualene in cholesterol biosynthesis (37,38). FDFT1 is an upstream enzyme of
SQLE in cholesterol biosynthesis and plays a key regulatory role in
catalyzing the dimerization of two molecules of farnesyl
diphosphate to produce squalene (39). DHCR24 is the final enzyme in
cholesterol biosynthesis that catalyzes the reduction of the
delta-24 double bond in sterol intermediates to form cholesterol
(40). To address whether FSCN1
affects cholesterol metabolism, the expression of the
aforementioned three relevant genes was validated using RT-qPCR.
The expression levels of SQLE, FDFT1 and DHCR24 were significantly
downregulated in the LSCC cells in which FSCN1 was knocked down
(Fig. 4E and F), suggesting that
FSCN1 plays a role in cholesterol synthetic metabolism. Studies
have reported that these three genes are involved in cancer
development and progression (41-44). Therefore, FSCN1 may regulate LSCC
progression by maintaining intracellular cholesterol metabolic
homeostasis.
Furthermore, through a crosstalk analysis with
FSCN1-interacting proteins in LSCC cells, three genes/proteins
(PTGR1, DHCR24 and SLC38A2) were shown to regulate LSCC progression
by FSCN1. Following co-IP validation, it was confirmed that DHCR24
and PTGR1 could bind to FSCN1. The interaction between FSCN1 and
DHCR24 further strengthened the role of FSCN1 in cholesterol
metabolism.
Previous studies have reported that DHCR24 is
dysregulated in various types of cancer (41,42,45-47). Its high expression has been shown
to be associated with aggressiveness and disease recurrence in
endometrial, urothelial and hepatocellular carcinoma (41,42,47). Consistently, the data of the
present study revealed that the overexpression of DHCR24 promoted
LSCC cell proliferation and migration.
Finally, several studies have been conducted on the
potential mechanisms through which FSCN1 affects the expression of
a large number of genes (29,31). For example, Saad et al
(31) revealed that
phosphorylated FSCN1 (p-FSCN1) localized in the nucleus and
regulated histone methylation and gene transcription.
Mechanistically, p-FSCN1 was shown to specifically interact with
the H3K4 methyltransferase core subunit RbBP5 form H3K4me3. Nuclear
pFSCN1 interactions with the RNA polymerase II complex further
elucidate the role of FSCN1 in gene transcription (31). In addition, FSCN1 knockdown has
been reported to affect the expression of a number of genes in
various types of cancer cells, such as esophageal squamous cell
carcinoma, triple-negative breast cancer and cervical cancer cells
(29,30,48). These results indicate that FSCN1
may be a transcriptional regulator.
In conclusion, the present study demonstrated that
FSCN1 knockdown affected the expression of up to 1,063 genes, with
462 upregulated and 601 downregulated genes in TU-177 cells.
Through crosstalk analysis and validation, FSCN1 was shown to be
linked to novel functions, including tje defense response to virus
and steroid biosynthesis. In particular, it was found that DHCR24,
a key enzyme in cholesterol biosynthesis, interacted with FSCN1,
suggesting that FSCN1 may promote LSCC progression by mediating
cholesterol metabolism-related signaling pathways. Overall, the
present study provided a comprehensive understanding of the diverse
functions of FSCN1 and found that the FSCN1-DHCR24 interaction may
play a key role in LSCC progression.
Supplementary Data
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HL and WH designed the study and wrote the first
draft of the manuscript. CZ and HL revised the manuscript. WH, XW
and YZ conducted the bioinformatics analysis. HL, XX and LH
developed the methods and performed the validation. LH, XW and XX
participated in data analysis and tabulation. YZ, JY and CZ
performed the statistical analysis. HL and CZ confirmed the
authenticity of all the raw data. All authors have read and agreed
to the published version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 81802793), the Basic Research
Program of Shanxi Province (Free Exploration) (grant nos.
202203021211015, 20210302124704, 20210302124594 and
202103021223430), the Science Research Start-up Fund for Doctor of
Shanxi Medical University (XD1801), Scientific and Technological
Innovation Programs of Higher Education Institutions in Shanxi
(STIP; grants 2021-185), and the Medical Science and Technology
Innovation Team of Shanxi Province (grant no. 2020TD26).
References
|
1
|
Johnson DE, Burtness B, Leemans CR, Lui
VWY, Bauman JE and Grandis JR: Head and neck squamous cell
carcinoma. Nat Rev Dis Primers. 6:922020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steuer CE, El-Deiry M, Parks JR, Higgins
KA and Saba NF: An update on larynx cancer. CA Cancer J Clin.
67:31–50. 2017. View Article : Google Scholar
|
|
3
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gao W, Zhang C, Li W, Li H, Sang J, Zhao
Q, Bo Y, Luo H, Zheng X, Lu Y, et al: Promoter
methylation-regulated miR-145-5p inhibits laryngeal squamous cell
carcinoma progression by targeting FSCN1. Mol Ther. 27:365–379.
2019. View Article : Google Scholar :
|
|
6
|
Chiyomaru T, Enokida H, Tatarano S,
Kawahara K, Uchida Y, Nishiyama K, Fujimura L, Kikkawa N, Seki N
and Nakagawa M: miR-145 and miR-133a function as tumour suppressors
and directly regulate FSCN1 expression in bladder cancer. Br J
Cancer. 102:883–891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ghebeh H, Al-Khaldi S, Olabi S, Al-Dhfyan
A, Al-Mohanna F, Barnawi R, Tulbah A, Al-Tweigeri T, Ajarim D and
Al-Alwan M: Fascin is involved in the chemotherapeutic resistance
of breast cancer cells predominantly via the PI3K/Akt pathway. Br J
Cancer. 111:1552–1561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu H, Zhang Y, Li L, Cao J, Guo Y, Wu Y
and Gao W: Fascin actin-bundling protein 1 in human cancer:
Promising biomarker or therapeutic target? Mol Ther Oncolytics.
20:240–264. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li CH, Chan MH, Liang SM, Chang YC and
Hsiao M: Fascin-1: Updated biological functions and therapeutic
implications in cancer biology. BBA Adv. 2:1000522022. View Article : Google Scholar
|
|
10
|
Li Z, Shi J, Zhang N, Zheng X, Jin Y, Wen
S, Hu W, Wu Y and Gao W: FSCN1 acts as a promising therapeutic
target in the blockade of tumor cell motility: A review of its
function, mechanism, and clinical significance. J Cancer.
13:2528–2539. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alam H, Bhate AV, Gangadaran P, Sawant SS,
Salot S, Sehgal L, Dange PP, Chaukar DA, D'cruz AK, Kannanl S, et
al: Fascin overexpression promotes neoplastic progression in oral
squamous cell carcinoma. BMC Cancer. 12:322012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Arlt MJ, Kuzmanov A, Snedeker JG, Fuchs B,
Silvan U and Sabile AA: Fascin-1 enhances experimental osteosarcoma
tumor formation and metastasis and is related to poor patient
outcome. BMC Cancer. 19:832019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao J, Zhou Y, Zhang Z, Tian F, Ma N, Liu
T, Gu Z and Wang Y: Upregulated fascin1 in non-small cell lung
cancer promotes the migration and invasiveness, but not
proliferation. Cancer Lett. 290:238–247. 2010. View Article : Google Scholar
|
|
14
|
Adams JC: Fascin-1 as a biomarker and
prospective therapeutic target in colorectal cancer. Expert Rev Mol
Diagn. 15:41–48. 2015. View Article : Google Scholar
|
|
15
|
Tan VY, Lewis SJ, Adams JC and Martin RM:
Association of fascin-1 with mortality, disease progression and
metastasis in carcinomas: A systematic review and meta-analysis.
BMC Med. 11:522013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang L, Jia Y, Jiang Z, Gao W and Wang B:
FSCN1 is upregulated by SNAI2 and promotes epithelial to
mesenchymal transition in head and neck squamous cell carcinoma.
Cell Biol Int. 41:833–841. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin S, Li Y, Wang D, Huang C, Marino D,
Bollt O, Wu C, Taylor MD, Li W, DeNicola GM, et al: Fascin promotes
lung cancer growth and metastasis by enhancing glycolysis and
PFKFB3 expression. Cancer Lett. 518:230–242. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen C, Xie B, Li Z, Chen L, Chen Y, Zhou
J, Ju S, Zhou Y, Zhang X, Zhuo W, et al: Fascin enhances the
vulnerability of breast cancer to erastin-induced ferroptosis. Cell
Death Dis. 13:1502022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao W, Zhang C, Feng Y, Chen G, Wen S,
Huangfu H and Wang B: Fascin-1, ezrin and paxillin contribute to
the malignant progression and are predictors of clinical prognosis
in laryngeal squamous cell carcinoma. PLoS One. 7:e507102012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu CP, Zhou L, Gong HL, Du HD, Tian J, Sun
S and Li JY: Establishment and characterization of a novel
HPV-negative laryngeal squamous cell carcinoma cell line, FD-LSC-1,
with missense and nonsense mutations of TP53 in the DNA-binding
domain. Cancer Lett. 342:92–103. 2014. View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Zhang Y, Zheng X, Dai F, Lu Y, Dai
L, Niu M, Guo H, Li W, Xue X, et al: Circular RNA circCORO1C
promotes laryngeal squamous cell carcinoma progression by
modulating the let-7c-5p/PBX3 axis. Mol Cancer. 19:992020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu H, Cui J, Zhang Y, Niu M, Xue X, Yin
H, Tang Y, Dai L, Dai F, Guo Y, et al: Mass spectrometry-based
proteomic analysis of FSCN1-interacting proteins in laryngeal
squamous cell carcinoma cells. IUBMB Life. 71:1771–1784. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ristic B, Kopel J, Sherazi S, Gupta S,
Sachdeva S, Bansal P, Ali A, Perisetti A and Goyal H: Emerging role
of fascin-1 in the pathogenesis, diagnosis, and treatment of the
gastrointestinal cancers. Cancers (Basel). 13:25362021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zou J, Yang H, Chen F, Zhao H, Lin P,
Zhang J, Ye H, Wang L and Liu S: Prognostic significance of
fascin-1 and E-cadherin expression in laryngeal squamous cell
carcinoma. Eur J Cancer Prev. 19:11–17. 2010. View Article : Google Scholar
|
|
28
|
Durmaz A, Kurt B, Ongoru O, Karahatay S,
Gerek M and Yalcin S: Significance of fascin expression in
laryngeal squamous cell carcinoma. J Laryngol Otol. 124:194–198.
2010. View Article : Google Scholar
|
|
29
|
Guo F, Liu Y, Cheng Y, Zhang Q, Quan W,
Wei Y and Hong L: Transcriptome analysis reveals the potential
biological function of FSCN1 in HeLa cervical cancer cells. PeerJ.
10:e129092022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Barnawi R, Al-Khaldi S, Majid S, Qattan A,
Bakheet T, Fallatah M, Ghebeh H, Alajez NM and Al-Alwan M:
Comprehensive transcriptome and pathway analyses revealed central
role for fascin in promoting triple-negative breast cancer
progression. Pharmaceuticals (Basel). 14:12282021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saad A, Bijian K, Qiu D, da Silva SD,
Marques M, Chang CH, Nassour H, Ramotar D, Damaraju S, Mackey J, et
al: Insights into a novel nuclear function for Fascin in the
regulation of the amino-acid transporter SLC3A2. Sci Rep.
6:366992016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li S, Huang XT, Wang MY, Chen DP, Li MY,
Zhu YY, Yu Y, Zheng L, Qi B and Liu JQ: FSCN1 promotes radiation
resistance in patients with PIK3CA gene alteration. Front Oncol.
11:6530052021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elkhatib N, Neu MB, Zensen C, Schmoller
KM, Louvard D, Bausch AR, Betz T and Vignjevic DM: Fascin plays a
role in stress fiber organization and focal adhesion disassembly.
Curr Biol. 24:1492–1499. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Villari G, Jayo A, Zanet J, Fitch B,
Serrels B, Frame M, Stramer BM, Goult BT and Parsons M: A direct
interaction between fascin and microtubules contributes to adhesion
dynamics and cell migration. J Cell Sci. 128:4601–4614.
2015.PubMed/NCBI
|
|
35
|
Ediriweera MK: Use of cholesterol
metabolism for anti-cancer strategies. Drug Discov Today.
27:1033472022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu H, Zhou S, Tang Q, Xia H and Bi F:
Cholesterol metabolism: New functions and therapeutic approaches in
cancer. Biochim Biophys Acta Rev Cancer. 1874:1883942020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gill S, Stevenson J, Kristiana I and Brown
AJ: Cholesterol-dependent degradation of squalene monooxygenase, a
control point in cholesterol synthesis beyond HMG-CoA reductase.
Cell Metab. 13:260–273. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
You W, Ke J, Chen Y, Cai Z, Huang ZP, Hu P
and Wu X: SQLE, a key enzyme in cholesterol metabolism, correlates
with tumor immune infiltration and immunotherapy outcome of
pancreatic adenocarcinoma. Front Immunol. 13:8642442022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Coman D, Vissers LELM, Riley LG, Kwint MP,
Hauck R, Koster J, Geuer S, Hopkins S, Hallinan B, Sweetman L, et
al: Squalene synthase deficiency: Clinical, biochemical, and
molecular characterization of a defect in cholesterol biosynthesis.
Am J Hum Genet. 103:125–130. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Luu W, Zerenturk EJ, Kristiana I, Bucknall
MP, Sharpe LJ and Brown AJ: Signaling regulates activity of DHCR24,
the final enzyme in cholesterol synthesis. J Lipid Res. 55:410–420.
2014. View Article : Google Scholar :
|
|
41
|
Dai M, Zhu XL, Liu F, Xu QY, Ge QL, Jiang
SH, Yang XM, Li J, Wang YH, Wu QK, et al: Cholesterol synthetase
DHCR24 induced by insulin aggravates cancer invasion and
progesterone resistance in endometrial carcinoma. Sci Rep.
7:414042017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee GT, Ha YS, Jung YS, Moon SK, Kang HW,
Lee OJ, Joung JY, Choi YH, Yun SJ, Kim WJ and Kim IY: DHCR24 is an
independent predictor of progression in patients with
non-muscle-invasive urothelial carcinoma, and its functional role
is involved in the aggressive properties of urothelial carcinoma
cells. Ann Surg Oncol. 21(Suppl 4): S538–S545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang H, Tang E, Chen Y, Liu H, Zhao Y,
Lin M and He L: Squalene synthase predicts poor prognosis in stage
I-III colon adenocarcinoma and synergizes squalene epoxidase to
promote tumor progression. Cancer Sci. 113:971–785. 2022.
View Article : Google Scholar :
|
|
44
|
Wang S, Dong L, Ma L, Yang S, Zheng Y,
Zhang J, Wu C, Zhao Y, Hou Y, Li H and Wang T: SQLE facilitates the
pancreatic cancer progression via the lncRNA-TTN-AS1/miR-133b/SQLE
axis. J Cell Mol Med. 26:3636–3647. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Battista MC, Guimond MO, Roberge C, Doueik
AA, Fazli L, Gleave M, Sabbagh R and Gallo-Payet N: Inhibition of
DHCR24/seladin-1 impairs cellular homeostasis in prostate cancer.
Prostate. 70:921–933. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fuller PJ, Alexiadis M, Jobling T and
McNeilage J: Seladin-1/DHCR24 expression in normal ovary, ovarian
epithelial and granulosa tumours. Clin Endocrinol (Oxf).
63:111–115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu J, Guo L, Qiu X, Ren Y, Li F, Cui W and
Song S: Genkwadaphnin inhibits growth and invasion in
hepatocellular carcinoma by blocking DHCR24-mediated cholesterol
biosynthesis and lipid rafts formation. Br J Cancer. 123:1673–1685.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Du ZP, Wu BL, Xie JJ, Lin XH, Qiu XY, Zhan
XF, Wang SH, Shen JH, Li EM and Xu LY: Network analyses of gene
expression following fascin knockdown in esophageal squamous cell
carcinoma cells. Asian Pac J Cancer Prev. 16:5445–5451. 2015.
View Article : Google Scholar : PubMed/NCBI
|

















