Introduction
Neuroblastoma (NB) is an extra-cranial pediatric
cancer. Effective neuroblastoma treatments are still limited
despite the advances in modern medicine, and we are in need of
novel chemotherapeutic strategies and more effective anticancer
agents (1–3). Many drugs used in cancer chemotherapy
is plant-derived or based on natural compounds (4–9). A
recent study screened a library of 500 plant extracts for
anti-proliferative and cytotoxic effects in neuroblastoma cell
lines in order to identify new active substances for chemotherapy.
The current study examines one extract from Scrophularia
orientalis L., as it was shown to have potent anticancer
effects in NB cells.
Members of the genus, Scrophularia, are
herbaceous flowering plants. There are 200 known species of
Scrophularia. Scrophularia species are used in
Traditional Chinese Medicine as a component in a formula used to
treat arthritis (10–13). It has anti-inflammatory effects and
is also a potent analgesic (14–17).
Several species of Scrophularia have been found to have
anticancer properties (18,19),
however, there are no studies documenting the medicinal effects of
Scrophularia orientalis L.
In the current study, a Scrophularia
orientalis dichloromethane extract was found to have anticancer
effects in NB, and was further examined to elucidate these effects.
Calcium is a second messenger that regulates many fundamental
physiological processes, including tumor progression and apoptosis
(20–24). Therefore, we used Fluo4-AM staining
and fluorescence microscopy while concurrently performing
Sulforhodamine B (SRB) staining on NB cells in order to determine
the effect of this extract on intracellular calcium signaling and
cell viability, respectively. The dichloromethane extract of
Scrophularia orientalis significantly increased
intracellular-free calcium levels and reduced NB cell viability.
The IC50 for the extract was ~5 μg/ml in NB cells with
and without MycN overexpression. The increase in intracellular-free
calcium appeared to be mediated by a calcium release mechanism
rather than calcium influx.
In addition, the mitochondrial transition pore assay
and western blot analysis revealed that cell death is associated
with opening of the mitochondrial permeability transition pore
(mPTP), and caspase-3 and PARP cleavage in NB cells. The results
from the current study suggest that the dichloromethane extract
derived from Scrophularia orientalis effectively kills NB
cells by inducing apoptosis via calcium release and consequently,
the opening of mPTP. The present study indicates that this extract
be used as an alternative strategy for treating NB, and may lead to
the development of more effective anticancer agents for NB.
Materials and methods
Cell culture
MYCN2 cells, a tetracycline inducible MycN
overexpression NB cell line was authenticated by the cell Line
Authentication testing Services at Genetica DNA Laboratories (USA)
using STR DNA typing to verify each cell line and verify pure cells
(no contamination). The cells were maintained in RPMI-1640
(Mediatech, Inc., Manassas, VA, USA) containing 10% (v/v)
heat-inactivated fetal bovine serum (FBS) (Atlanta Biologicals,
Lawrenceville, GA, USA), and grown at 37°C, 5% CO2, in a
95% humidity.
Chemicals
General chemicals were from VWR (West Chester, PA,
USA). Doxorubicin and ionomycin were from Calbiochem (Gibbstown,
NJ, USA). The Library of extracts, which included the
Scrophularia orientalis extracts, was obtained from Dr
Robert Borris.
Preparation of Scrophularia extracts
Samples of Scrophularia orientalis L. were
collected in the Sevan Pass between Dilijan and Tsovagyugh,
Gegharkunik Province, Armenia, in June 2006. Voucher specimens
(Tamanyan 42–2006) have been deposited in the herbaria of the New
York Botanical Garden and the Armenian National Academy of
Sciences. Fresh samples were air dried and then milled to a coarse
powder. A 1 kg (dry weight) portion of each sample was extracted
with methanol (3×4 liters) and the solvent removed in vacuo
to afford viscous oil. The resulting oil was dispersed in 1 liter
of methanol:water (9:1) and extracted with n-hexane (3×1 liters).
The depleted hydroalcoholic phase was freed of methanol, dispersed
in distilled water (1 liter) and extracted sequentially with
dichloromethane (3×1 liter) and water-saturated n-butanol (3×1
liters). The resulting solvent-soluble fractions were individually
evaporated to dryness in vacuo, while the residual aqueous
phase was freed of solvent and then lyophilized. Extracts and
fractions were maintained at −20°C until needed for use.
Calcium assay
MYCN2 cells were washed and incubated with 1 μM
Fluo-4 AM, the acetoxymethyl ester form of Fluo-4 (Molecular
Probes, Eugene, OR, USA), for 30 min at 37°C in a standard modified
Ringer's solution of the following composition (in mM): NaCl 145,
KCl 2.8, CsCl 10, CaCl2 2 (or 0), MgCl2 2,
glucose 10, Hepes·NaOH 10, pH 7.4, 330 mOsm. For nominally
calcium-free experiments 1 mM EGTA was added to the external
solution and calcium chloride was omitted. Cells were transferred
to 96-well plates at 10,000 cells/well and stimulated as indicated.
Epifluorescent measurements were performed using an Operetta High
Content Imaging System (PerkinElmer, Santa Clara, CA, USA).
Fluorescence intensity was quantified using Harmony
(PerkinElmer).
Fluorescence measurements
MYCN2 cells were incubated in a standard modified
Ringer's solution of the following composition (in mM): NaCl 145,
KCl 2.8, CsCl 10, CaCl2 2 (or 0), MgCl2 2,
glucose 10, Hepes·NaOH 10, pH 7.4, 330 mOsm. For nominally
calcium-free experiments, 1 mM EGTA was added to the external
solution and calcium chloride was omitted. Cells were loaded with
fura-2 AM, the acetoxymethyl ester form of fura-2 (Molecular
Probes). Cells were perfused with external solutions containing
Scrophularia orientalis extract, and cytosolic calcium was
measured in individual cells using a Zeiss microscope and
monochromatic light source tuned to excite fura-2 fluorescence at
360 and 390 nm for 20 msec each. Emission was detected at 450–550
nm using a photomultiplier.
Sulforhodamine B assay
The SRB colorimetric assay was used to determine
cell proliferation following the protocol previously described
(8). Briefly, cells were seeded at
a density of 10,000 cells/well on a transparent, flat-bottom,
96-well plate and allowed to settle overnight. At the initiation of
each experiment (t=0), and after drug treatments, 100 μl of 10%
(w/v) TCA were added to each well, incubated for 1 h at 4°C, washed
with deionized water, and dried at room temperature. One hundred
microliters of 0.057% (w/v) SRB solution were added to each well,
incubated for 30 min at room temperature, rinsed four times with 1%
(v/v) acetic acid, and allowed to dry at room temperature. Finally,
200 μl of 10 mM Tris base solution (pH 10.5) was added to each
well, and after shaking for 5 min at room temperature, the
absorbance was measured at 510 nm in a microplate reader. The
absorbance at t=0 was compared with the absorbance at the end of
the experiment to determine cell growth in treated cells compared
with control cells.
Mitochondrial permeability transition
pore assay
Treated and untreated NB cells were washed twice in
modified Hanks' Balanced Salt Solution (HBSS: sodium bicarbonate,
calcium, and magnesium that also included 10 mM HEPES, 2 mM
L-glutamine and 100 μM succinate), then labeled with 1.0 μM calcein
AM, 200 nM MitoTracker Red CMXRos, 1 μM Hoechst 33342 dye and 1.0
μM CoCl2. Cells were incubated for 15 min at 37°C, 5%
CO2, in 95% humidity, then washed in modified HBSS.
Ionomycin (1 μM) was used as a positive control for
calcium-mediated pore opening. Cells were labelled with Calcein Am
and MitoTracker Red. Epifluorescent measurements were performed
using an Operetta High Content Imaging System (PerkinElmer).
Fluorescence intensity was quantified using Harmony software
(PerkinElmer).
Western blot analysis
Cell lysates were prepared in
radioim-munoprecipitation assay buffer [20 mmol/l Tris-HCl (pH
7.5), 0.1% (w/v) sodium lauryl sulfate, 0.5% (w/v) sodium
deoxycholate, 135 mmol/l NaCl, 1% (v/v) Triton X-100, 10% (v/v)
glycerol, 2 mmol/l EDTA] supplemented with Complete protease
inhibitor cocktail (Roche Molecular Biochemicals) and phosphatase
inhibitors sodium fluoride (20 mmol/l) and sodium vanadate (0.27
mmol/l). Western blot analysis was performed as previously
described (20). The total protein
concentration was determined using the protein assay dye reagent
from Bio-Rad Laboratories. Cell lysates in SDS-sample buffer were
boiled for 5 min and equal amounts of total protein were analyzed
by 10% SDS-PAGE and western blotting. The antibodies used in this
study are mouse monoclonal p53 (1:250) from Santa Cruz
Biotechnology; rabbit polyclonal cleaved caspase-3 (1:1,000),
rabbit polyclonal cleaved PARP (1:1,000), mouse monoclonal GAPDH
(1:1,000), and mouse monoclonal PCNA (1:1000) from Cell Signaling
Technology. Proteins were detected using the Odyssey Infrared
Imaging System (LI-COR Biosciences, Lincoln, NB, USA) and analyzed
with Licor Image Studio 2.0 acquisition and analysis software.
Statistical analysis
Results are shown as the mean ± standard deviation.
Statistical significance was determined based on Student's t-test.
Adjacent to data points in the respective graphs, significant
differences were recorded as follows: single asterisk, P<0.05;
double asterisk, P<0.01; triple asterisk, P<0.001; no symbol,
P>0.05. SRB and FLuo-4 experiments are the number of at least 3,
in triplicates. All other experiments are reported as the mean of
at least 3.
Results
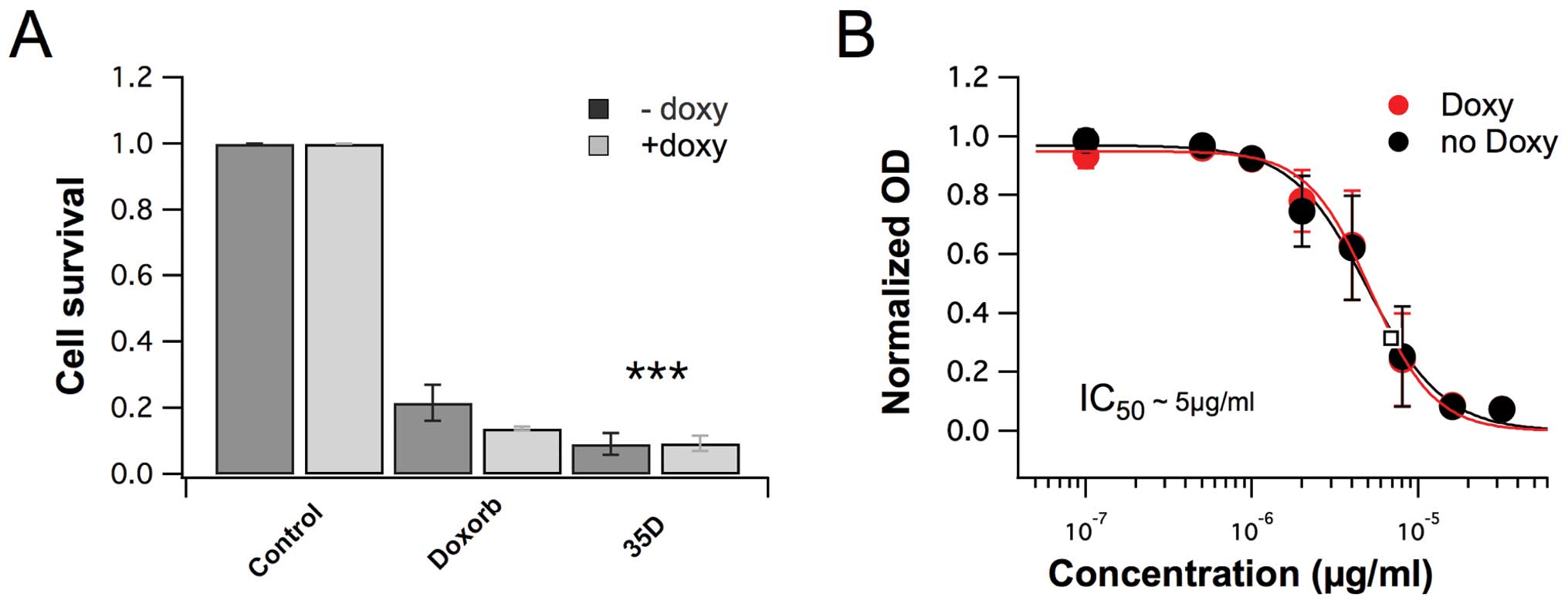
Scrophularia orientalis extracts reduce
NB cell viability
The library of 500 plant extracts was screened for
anticancer effects. The results clearly show that the
dichloromethane extracts of Scrophularia orientalis
significantly decreased viability in NB cells, compared to
untreated control cells. In NB cells with non-amplified MYCN
gene (NAM), extract at a concentration of 8 μg/ml, reduced cell
viability of NB cells by 91.6%, compared to the viability of
untreated control cells (Fig. 1A).
In NB cells with overexpression of MycN (MOE), the extract reduced
cell viability by 92.1% (Fig. 1A).
The cytotoxic effects were more potent than that of doxorubicin,
which reduced cell viability of NAM and MOE by 79.3% and 84.7%,
respectively (Fig. 1A). The
IC50 of the extract was determined, and the
IC50 of MNA and MOE were 8.33 μg/ml and 8.09 μg/ml,
respectively (Fig. 1B). The data
suggest that the dichloromethane extract of Scrophularia
orientalis has potent anticancer effects and effectively
decreases NB cell viability.
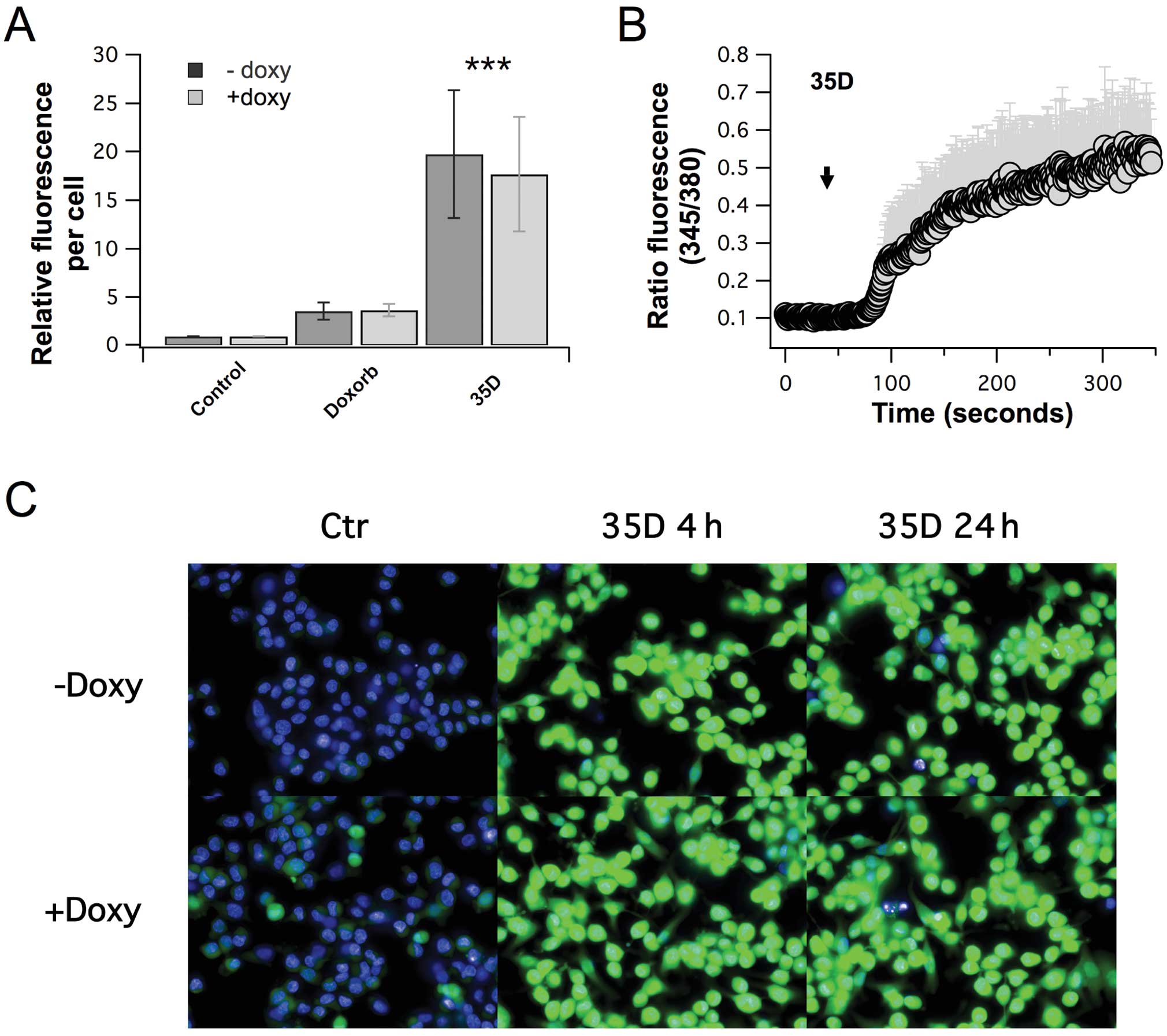
Scrophularia orientalis extract increases
intracellular calcium in NB cells
Calcium is a second messenger that plays a
fundamental role in a plethora of cellular processes, including
cell proliferation and cell death. Therefore, in order to determine
the effect of the Scrophularia orientalis extract on
calcium, intracellular-free calcium was measured in control,
untreated NB cells and NB cells treated with Scrophularia
orientalis extract. Treated and untreated cells were loaded
with Fluo-4, and relative fluorescence was measured using an
Operetta High Content Imaging System. Fig. 2A shows that there was a 20-fold
increase in intracellular-free calcium when NAM NB cells were
exposed to extract for 24 h, compared to untreated control.
Intracellular-free calcium increased by 18-fold when MOE NB cells
was exposed to extract, compared to untreated control (Fig. 2A). These changes in
intracellular-free calcium was significantly greater than that of
doxorubicin which increased intracellular-free calcium by 2.8- and
3.1-fold in NAM and MOE NB cells, respectively (Fig. 2A). Further examination of the
calcium inducing effects of the extract showed that external
application of extract to MOE NB cells increased intracellular-free
calcium steadily after a slight delay of approximately 30–45 sec,
whereas no signal was induced by vehicle control (Fig. 2B). This suggests that the extract
induces rapid changes in intracellular calcium. Of note, the
elevated calcium levels appeared to be sustained, as indicated by a
time course analysis of calcium levels after treatment of NB cells
with the extract for 4 and 24 h (Fig.
2C). These results clearly show that the Scrophularia
dichloromethane extract had significant and distinct effects on
calcium signaling in NB cells.
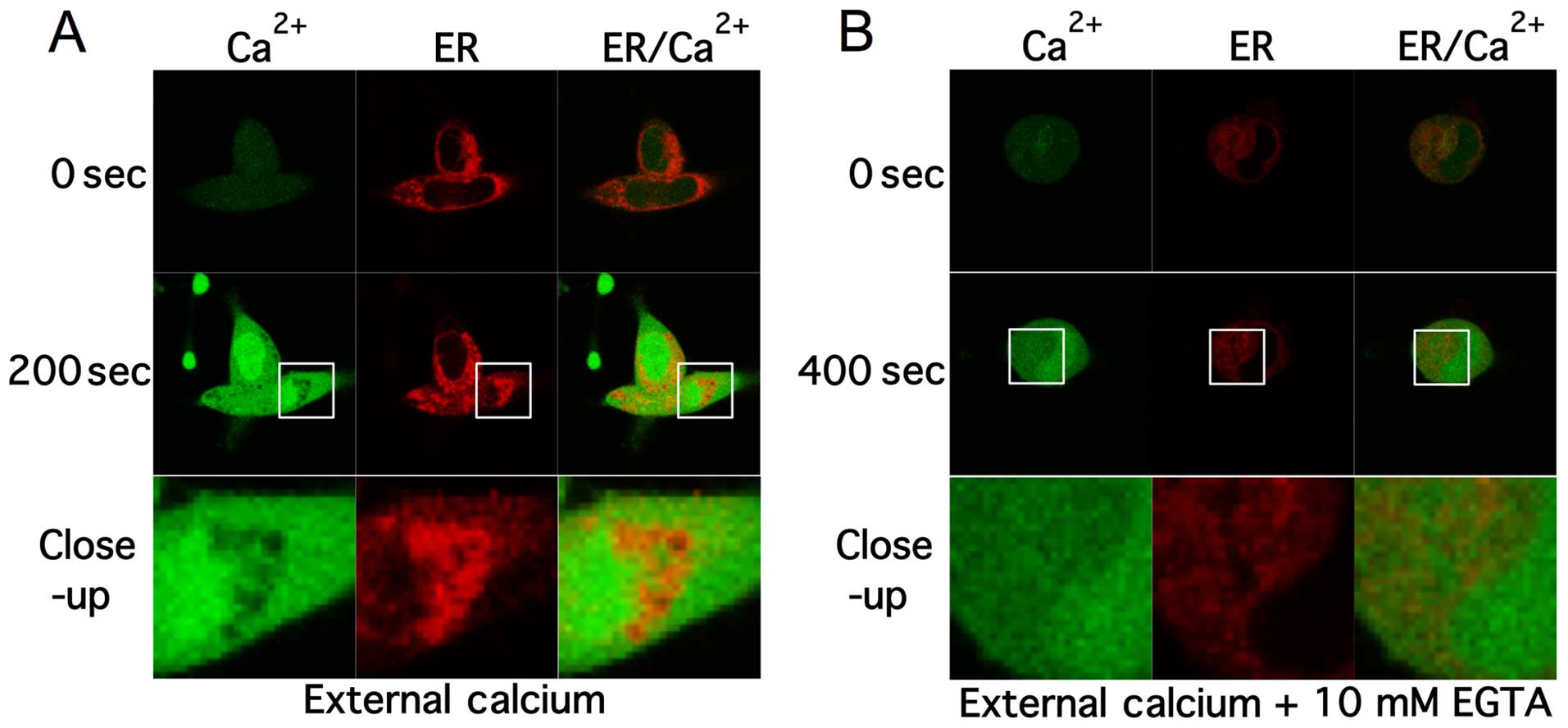
Scrophularia orientalis extracts induce
calcium release from intracellular stores and calcium influx
In order to decipher the source of
intracellular-free calcium time-lapse confocal experiments were
performed. Cells were labelled with Fluo-4 AM and ER-tracker.
Confocal images of the 2 channels were acquired every 1.5 sec.
Standard extra cellular HBSS buffer was used that contained ~1.3
mM-free calcium and ~800 μM- free magnesium. Baseline fluorescence
was measured in cells for 30 sec then cells were perfused with
extracellular solution containing Scrophularia extract
through a glass application pipette. Upon exposure with extract,
intracellular calcium levels started to rise within 40–60 sec and
peaked at ~150–200 sec. Fig. 3A
shows confocal images of fluo-4 (green) and ER-tracker (red)
labelled NB cells pre- and post-application of extract. After 200
sec intracellular calcium rose to peak levels and remained elevated
up to 1000 sec, as shown by the increase in fluorescence.
Noteworthy, low levels of calcium within the cell co-localized with
endoplasmic reticulum (ER) staining, suggesting that the increase
in calcium may be due to calcium ER store depletion.
Intracellular-free calcium can be mobilized from either
transporters in the membrane of organelles, such as the ER, that
act as intracellular calcium stores, or the extracellular space
through ion channels and transporters. To further investigate the
source the Scrophularia extract mediated calcium
mobilisation, 10 mM EGTA was added to the HBSS buffer, chelating
extracellular-free calcium levels to ~10 nM (Fig. 3B).
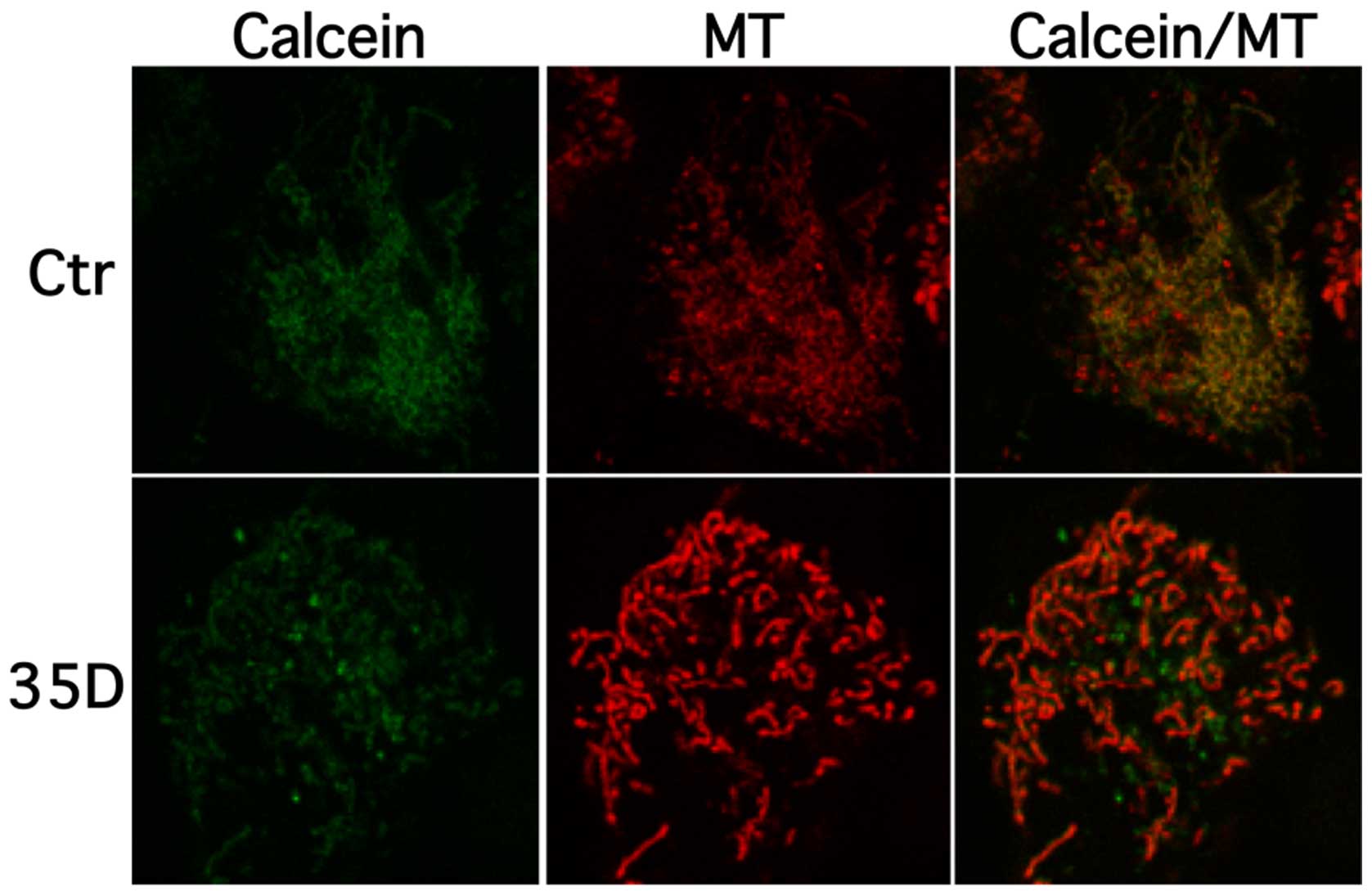
Scrophularia orientalis extracts induce
opening of mitochondrial permeability transition pore (MPTP) in NB
cells
To examine the effect of the Scrophularia
dichloromethane extract on loss of mitochondrial membrane
integrity, cells were stained with calcein AM cobalt
(CoCl2) and mitochondrial dye, and confocal measurements
were performed. In untreated healthy cells calcein-AM,
CoCl2 are taken up and non-specific esterase activity
cleaves and activates calcein. Cytoplasmic calcein is almost
entirely quenched by CoCl2 whereas mitochondrial calcein
is not accessed by CoCl2 freely leading to an overlap of
calcein and mitotracker signal in healthy cells. Upon induction of
mitochondrial pore opening CoCl2 enters the organelle
leading to quenching and subsequent drop of mitochondrial calcein
signal. Fig. 4 shows that
treatment of NAM cells with extract decreases mitochondrial calcein
co-localisations levels, whereas healthy cells display a clear
overlap of calcein (green) and mitochondrial (red) signal. These
results suggest that the Scrophularia extract induces loss
of mitochondrial membrane integrity, here indicated by calcein and
mitochondria distribution.
Scrophularia orientalis extract induces
apoptosis in NB cells
To determine whether the Scrophularia extract
indeed induced mitochondrial calcium overload and subsequently
promoted apoptosis, whole cell lysates were prepared from NB cells
treated with the extracts or left untreated, and the lysates were
analysed by western blot for detection of apoptotic markers,
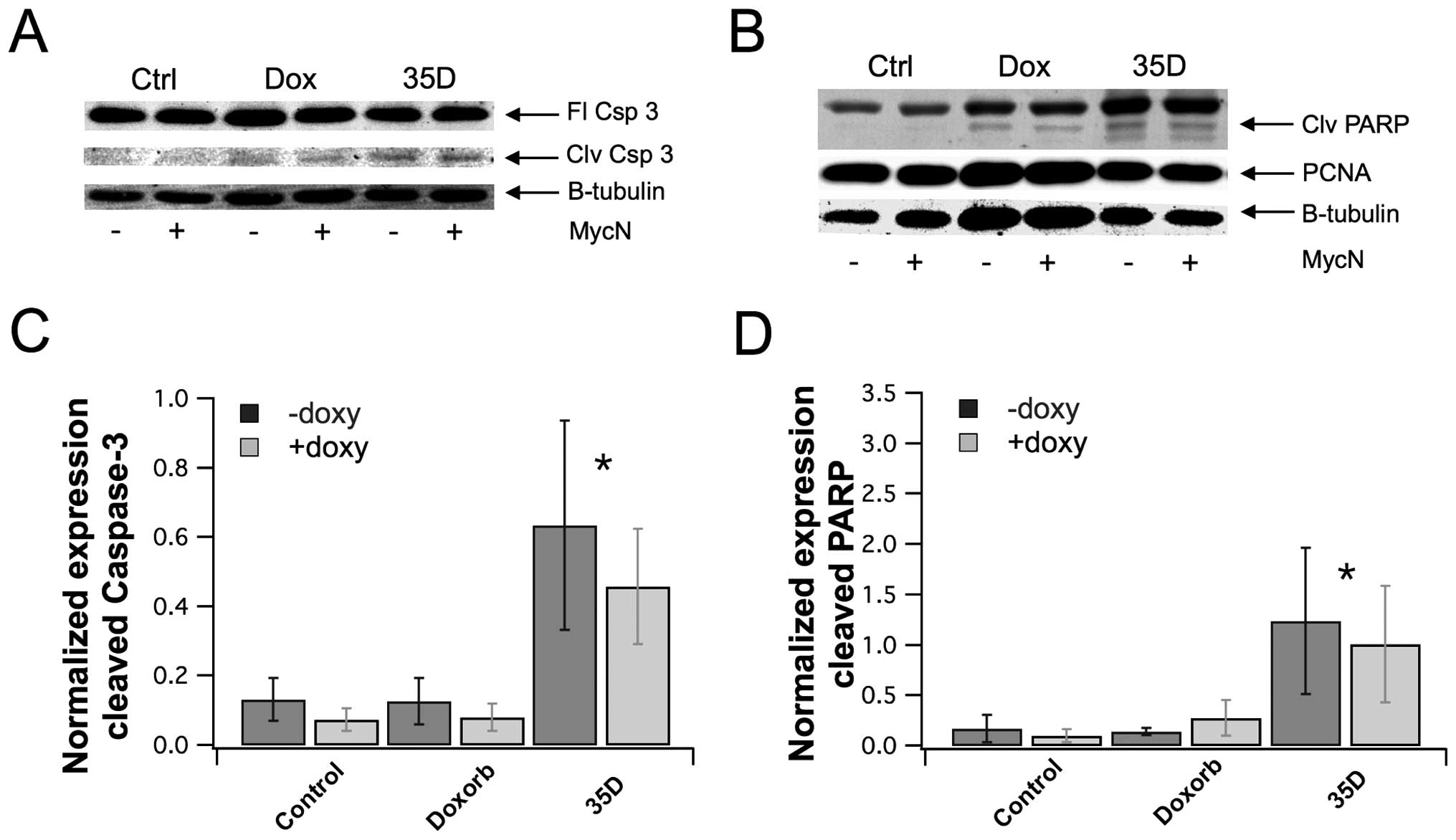
cleaved caspase-3 and cleaved PARP. Fig. 5A and B shows that extract increased
the cleavage of caspase-3 and PARP. However, there was no effect on
total caspase expression. In addition, there was no effect on
proliferation marker, proliferating cell nuclear antigen (PCNA).
The band density in each lane of the western blot was quantified,
and the level of cleaved caspase-3 and cleaved PARP were normalized
to GAPDH expression. The quantification of cleaved caspase-3 showed
that the extract significantly increased caspase-3 cleavage by
4.8-fold for NAM and 6.1-fold for MOE, compared to control
(Fig. 5C). The quantification of
cleaved PARP showed that the extract significantly increased PARP
cleavage by 7.2 and 9.9 for NAM and MOE, respectively, compared to
control (Fig. 5D). These results
suggest that the dichloromethane Scrophularia extract induce
mitochondrial calcium overload, opening of the mitochondrial
permeability transition pore and promoting apoptosis.
Discussion
Scrophularia are herbaceous flowering plants.
There are 200 known species of Scrophularia, of which
several species have been found to have medicinal effects. For
example, Scrophularia is used in Traditional Chinese
medicine as a component in a formula used to treat arthritis
(25). Previous studies have shown
that Scrophularia has anti-inflammatory effects and may also
be used as an analgesic (14–17).
Several species have been found to have anticancer properties
(18,19). However, there are no studies
investigating the medicinal effects of Scrophularia
orientalis. In the current study, an extract from
Scrophularia orientalis which was part of a library of 500
plant extracts that was screened in a previous study was found to
have potent anticancer effects in NB cells. The anticancer effects
of the Scrophularia orientalis dichloromethane extract were
further examined, and the results showed that the extract
significantly decreased cell viability of NB cells with and without
MycN overexpression. MycN is the protein that is the product of
MYCN gene expression. MycN is a transcription factor that contains
a C-terminal basic helix-loop-helix zipper motif and an N-terminal
transactivation domain. MycN binds to genes that contain the E-box
consensus sequence in the promoter (26). MYCN gene amplification is a
negative prognostic indicator of NB, and is associated with
high-risk and advanced stage NB (27,28).
In addition, MYCN amplification may alter the
anticancer effects of chemotherapeutic drugs and/or confer
resistance to some treatments (29). Identification of alternative
treatments with selectivity for MYCN amplified NB or NB with
MycN overexpression would be beneficial for the treatment of
advanced stage, high-risk NB. Therefore, this study examined the
effect of the dichloromethane Scrophularia orientalis
extract in NB cells with and without MycN overexpression.
Interestingly, the IC50 of the extracts were similar in
NB cells with and without MycN overexpression, suggesting that
components of the extracts are effective in NB cells regardless of
MycN status.
The Scrophularia extract also altered
intracellular calcium regulation. Of note, application of the
extracts induced a rapid increase in intracellular calcium.
Confocal analysis showed that Scrophularia extract increased
intracellular-free calcium through a process that involved calcium
release from the ER, when calcium was present in the external
media. In the absence of external calcium, exposure of NB cells to
Scrophularia extract mobilized intracellular calcium to a
lower extent and with a ~200 second delay, compared to calcium
mobilization in the presence of external calcium. This suggests
that calcium influx, which requires external calcium, is involved
in the elevated calcium induced by Scrophularia extract.
Also, lower levels of intracellular calcium co-localized with ER
marker suggesting that calcium was mobilized, in part, from the ER.
Taken together the data suggest that exposure of
Scrophularia extract triggers at least two components of
calcium mobilisation one being sourced by ER calcium and the other
through plasma membrane mediated calcium flux. One of the
possibilities in this context would be the induction of store
operated calcium entry as one of the main influx pathways (20,21).
To confirm the ER-calcium depletion, experiments were also conduced
with Rhod-4 that has been reported to have greater access to ER
stores confirming the results (data not shown). Further the effect
of magnesium was ruled out by using buffer deficient of calcium and
magnesium (data not shown).
The increase in cytosolic calcium levels was
maintained for over 24 h. This effect may be due to increased or
dysregulated activity of calcium release transporters (e.g. IP3 or
Ryanodine receptors) or plasma membrane Store-operated calcium
channels (Orai) (30–35). Alternatively, activation of
transporter proteins that remove cytosolic calcium such as ATPases
that transport calcium into intracellular stores (SERCA) or out of
the cell (Na/Ca-exchanger, Ca-ATPase), may be hampered (36,37).
Inhibition of these processes could lead to the sustained increase
in intracellular calcium observed in this study.
Noteworthy, sustained high calcium levels in the
cytosol could lead to transport of calcium into the mitochondria.
The mitochondria is an interesting organelle. It requires calcium
in order to generate energy in the form of adenosine triphosphate
(ATP) (38,39). It also serves as a calcium sink and
can transiently buffer cytosolic calcium, as a cytoprotective
mechanism (40,41). In addition, there are complex
signaling between the ER and mitochondria that allows for
spatio-temporal patterns of calcium signaling between the two
organelles to tightly control cellular function and cell fate
(42–47). Transport of calcium from the ER to
the mitochondria at mitochondria associated membranes will help to
buffer cytosolic calcium levels and induce ATP production. The
increased ATP may feed back to the calcium-ATPase pump in the ER
membrane (SERCA) to pump calcium back to the ER (48,49).
However, alterations in the function of proteins that regulate
calcium signaling between these organelles could lead to a
sustained increase in mitochondria calcium that could lead to
opening of the mitochondria permeability transition pore (mPTP)
which eventually leads to activation of the apoptotic pathway
calcium (50–52).
In the current study, the data suggest that the
Scrophularia extract may alter the function of the calcium
transporters in the ER and/or mitochondria that may prevent the
transport of calcium from mitochondria and cytosol back into the ER
or out of the cell, which results in sustained cytosolic and
mitochondria calcium and eventually opening of the mPTP. The
opening of the mPTP resulted in activation of apoptosis in the NB
cells. Western blot analyses showed that the opening of the mPTP
consequently induced apoptosis as shown by the increase in both
cleaved caspase-3 and cleaved PARP, apoptosis markers. However, the
extract did not affect proliferation, as levels of (12) pCNA (proliferating cell nuclear
antigen, a proliferation marker) were unchanged when cells were
treated with the extract.
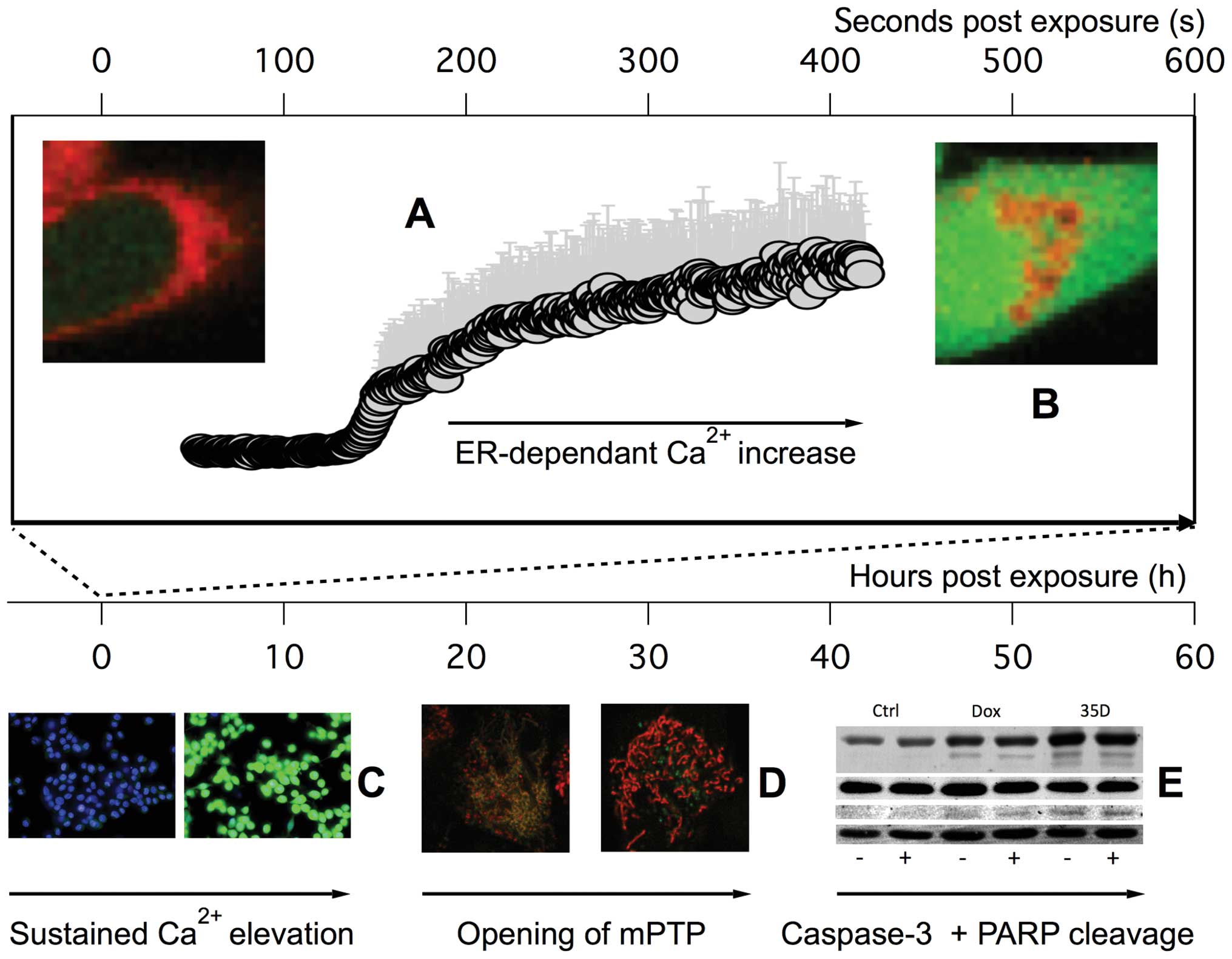
In conclusion, the current study showed that
Scrophularia orientalis dichloromethane extract has potent
anticancer effects. This extract rapidly increased in
intracellular-free calcium and altered calcium handling that led to
sustained elevation in intracellular calcium and ultimately,
opening of the mitochondria permeability transition pore and
apoptosis in NB cells (Fig. 6).
Components in the extracts may provide an alternative anticancer
treatment for NB, and should be investigated in future studies.
This study shows that targeting calcium signaling at the ER and/or
mitochondria may be a unique strategy for the development of more
effective, and novel drugs for the treatment of NB.
Acknowledgements
This study was supported by the Alex's Lemonade
Stand Foundation Young Investigator Award 439744 (D.-L.K), the NIH
grants from the National Cancer Institute Mentored Research
Scientist Development Award (K01) CA154758, (D.-L.K), and the
National Institutes of General Medical Sciences P20GM 103466
(D.-L.K).
References
|
1
|
Maris JM and Matthay KK: Molecular biology
of neuroblastoma. J Clin Oncol. 17:2264–2279. 1999.PubMed/NCBI
|
|
2
|
Combaret V, Delattre O, Bénard J and
Favrot MC: Biological markers for the prognosis of neuroblastoma:
Proposal of a method of analysis. Bull Cancer. 85:262–266. 1998.(In
French). PubMed/NCBI
|
|
3
|
Park JR, Eggert A and Caron H:
Neuroblastoma: Biology, prognosis, and treatment. Hematol Oncol
Clin North Am. 24:65–86. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sawadogo WR, Schumacher M, Teiten MH,
Dicato M and Diederich M: Traditional West African pharmacopeia,
plants and derived compounds for cancer therapy. Biochem Pharmacol.
84:1225–1240. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tan W, Lu J, Huang M, Li Y, Chen M, Wu G,
Gong J, Zhong Z, Xu Z, Dang Y, et al: Anti-cancer natural products
isolated from Chinese medicinal herbs. Chin Med. 6:272011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jo H, Loison F, Hattori H, Silberstein LE,
Yu H and Luo HR: Natural product Celastrol destabilizes tubulin
heterodimer and facilitates mitotic cell death triggered by
microtubule-targeting anti-cancer drugs. PLoS One. 5:e103182010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Desai AG, Qazi GN, Ganju RK, El-Tamer M,
Singh J, Saxena AK, Bedi YS, Taneja SC and Bhat HK: Medicinal
plants and cancer chemoprevention. Curr Drug Metab. 9:581–591.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Konkimalla VB and Efferth T: Anti-cancer
natural product library from traditional Chinese medicine. Comb
Chem High Throughput Screen. 11:7–15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mukherjee AK, Basu S, Sarkar N and Ghosh
AC: Advances in cancer therapy with plant based natural products.
Curr Med Chem. 8:1467–1486. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang LQ, Guo FJ, Wang SC and Li YM: A new
triterpenoid tetrasaccharide from the root of Scrophularia
ningpoensis. Yao Xue Xue Bao. 47:1358–1362. 2012.
|
|
11
|
Chen C, Duan LN, Zhou XL, Chen BL and Fu
CX: Molecular authentication of geo-authentic Scrophularia
ningpoensis. J Zhejiang Univ Sci B. 12:393–398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang CG, Shang YJ, Zhang J, Zhang JR, Li
WJ and Jiao BH: Hypouricemic effects of phenylpropanoid glycosides
acteoside of Scrophularia ningpoensis on serum uric acid levels in
potassium oxonate-pretreated mice. Am J Chin Med. 36:149–157. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang S, Zheng Z, Weng Y, Yu Y, Zhang D,
Fan W, Dai R and Hu Z: Angiogenesis and anti-angiogenesis activity
of Chinese medicinal herbal extracts. Life Sci. 74:2467–2478. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ahmad M, Muhammad N, Mehjabeen, Jahan N,
Alam SM, Ahmad M and Obaidullah: Biological screening of
Scrophularia nodosa extract and its fractions. Pak J Pharm Sci.
25:307–313. 2012.PubMed/NCBI
|
|
15
|
García D, Fernández A, Sáenz T and Ahumada
C: Anti-inflammatory effects of different extracts and harpagoside
isolated from Scrophularia frutescens L. Farmaco. 51:443–446.
1996.
|
|
16
|
Giner RM, Villalba ML, Recio MC, Máñez S,
Cerdá-Nicolás M and Ríos J: Anti-inflammatory glycoterpenoids from
Scrophularia auriculata. Eur J Pharmacol. 389:243–252. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fernández MA, Sáenz MT and García MD:
Anti-inflammatory activity in rats and mice of phenolic acids
isolated from Scrophularia frutescens. J Pharm Pharmacol.
50:1183–1186. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Giessrigl B, Yazici G, Teichmann M, Kopf
S, Ghassemi S, Atanasov AG, Dirsch VM, Grusch M, Jäger W, Ozmen A,
et al: Effects of Scrophularia extracts on tumor cell
proliferation, death and intravasation through lymphoendothelial
cell barriers. Int J Oncol. 40:2063–2074. 2012.PubMed/NCBI
|
|
19
|
Ardeshiry Lajimi A, Rezaie-Tavirani M,
Mortazavi SA, Barzegar M, Moghadamnia SH and Rezaee MB: Study of
anti-cancer property of Scrophularia striata extract on the human
astrocytoma cell line (1321). Iran J Pharm Res. 9:403–410.
2010.PubMed/NCBI
|
|
20
|
Qu B, Al-Ansary D, Kummerow C, Hoth M and
Schwarz EC: ORAI-mediated calcium influx in T cell proliferation,
apoptosis and tolerance. Cell Calcium. 50:261–269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu H, Hughes JD, Rollins S, Chen B and
Perkins E: Calcium entry via ORAI1 regulates glioblastoma cell
proliferation and apoptosis. Exp Mol Pathol. 91:753–760. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang W, Lu C, Wu Y, Ouyang S and Chen Y:
T-type calcium channel antagonists, mibefradil and NNC-55-0396
inhibit cell proliferation and induce cell apoptosis in leukemia
cell lines. J Exp Clin Cancer Res. 34:542015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi DL, Jang SJ, Cho S, Choi HE, Rim HK,
Lee KT and Lee JY: Inhibition of cellular proliferation and
induction of apoptosis in human lung adenocarcinoma A549 cells by
T-type calcium channel antagonist. Bioorg Med Chem Lett.
24:1565–1570. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ohkubo T and Yamazaki J: T-type
voltage-activated calcium channel Cav3.1, but not Cav3.2, is
involved in the inhibition of proliferation and apoptosis in MCF-7
human breast cancer cells. Int J Oncol. 41:267–275. 2012.PubMed/NCBI
|
|
25
|
Zhu T, Zhang L, Ling S, Duan J, Qian F, Li
Y and Xu JW: Scropolioside B inhibits IL-1β and cytokines
expression through NF-κB and inflammasome NLRP3 pathways. Mediators
Inflamm. 2014:8190532014. View Article : Google Scholar
|
|
26
|
Murphy DM, Buckley PG, Bryan K, Das S,
Alcock L, Foley NH, Prenter S, Bray I, Watters KM, Higgins D, et
al: Global MYCN transcription factor binding analysis in
neuroblastoma reveals association with distinct E-box motifs and
regions of DNA hypermethylation. PLoS One. 4:e81542009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chan HS, Gallie BL, DeBoer G, Haddad G,
Ikegaki N, Dimitroulakos J, Yeger H and Ling V: MYCN protein
expression as a predictor of neuroblastoma prognosis. Clin Cancer
Res. 3:1699–1706. 1997.
|
|
28
|
Tonini GP, Boni L, Pession A, Rogers D,
Iolascon A, Basso G, Cordero di Montezemolo L, Casale F, Pession A,
Perri P, et al: MYCN oncogene amplification in neuroblastoma is
associated with worse prognosis, except in stage 4s: The Italian
experience with 295 children. J Clin Oncol. 15:85–93.
1997.PubMed/NCBI
|
|
29
|
Gogolin S, Dreidax D, Becker G, Ehemann V,
Schwab M and Westermann F: MYCN/MYC-mediated drug resistance
mechanisms in neuroblastoma. Int J Clin Pharmacol Ther. 48:489–491.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo RW and Huang L: New insights into the
activation mechanism of store-operated calcium channels: Roles of
STIM and Orai. J Zhejiang Univ Sci B. 9:591–601. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peel SE, Liu B and Hall IP: ORAI and
store-operated calcium influx in human airway smooth muscle cells.
Am J Respir Cell Mol Biol. 38:744–749. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hewavitharana T, Deng X, Soboloff J and
Gill DL: Role of STIM and Orai proteins in the store-operated
calcium signaling pathway. Cell Calcium. 42:173–182. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vig M, Beck A, Billingsley JM, Lis A,
Parvez S, Peinelt C, Koomoa DL, Soboloff J, Gill DL, Fleig A, et
al: CRACM1 multimers form the ion-selective pore of the CRAC
channel. Curr Biol. 16:2073–2079. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Peinelt C, Vig M, Koomoa DL, Beck A,
Nadler MJ, Koblan-Huberson M, Lis A, Fleig A, Penner R and Kinet
JP: Amplification of CRAC current by STIM1 and CRACM1 (Orai1). Nat
Cell Biol. 8:771–773. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vig M, Peinelt C, Beck A, Koomoa DL, Rabah
D, Koblan-Huberson M, Kraft S, Turner H, Fleig A, Penner R, et al:
CRACM1 is a plasma membrane protein essential for store-operated
Ca2+ entry. Science. 312:1220–1223. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carafoli E and Guerini D: Molecular and
cellular biology of plasma membrane calcium ATPase. Trends
Cardiovasc Med. 3:177–184. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Newton T, Black JP, Butler J, Lee AG, Chad
J and East JM: Sarco/endoplasmic-reticulum calcium ATPase SERCA1 is
maintained in the endoplasmic reticulum by a retrieval signal
located between residues 1 and 211. Biochem J. 371:775–782. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martínez F, Uribe A, Espinosa-García MT,
Flores-Herrera O, García-Pérez C and Milán R: Calcium modulates the
ATP and ADP hydrolysis in human placental mitochondria. Int J
Biochem Cell Biol. 34:992–1003. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tanaka J, Kono Y, Shimahara Y, Sato T,
Jones RT, Cowley RA and Trump BF: A study of oxidative
phosphorylative activity and calcium-induced respiration of rat
liver mitochondria following living Escherichia coli injection. Adv
Shock Res. 7:77–90. 1982.PubMed/NCBI
|
|
40
|
García-Sancho J: The coupling of plasma
membrane calcium entry to calcium uptake by endoplasmic reticulum
and mitochondria. J Physiol. 592:261–268. 2014. View Article : Google Scholar :
|
|
41
|
Santo-Domingo J and Demaurex N: Calcium
uptake mechanisms of mitochondria. Biochim Biophys Acta.
1797:907–912. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Patergnani S, Suski JM, Agnoletto C,
Bononi A, Bonora M, De Marchi E, Giorgi C, Marchi S, Missiroli S,
Poletti F, et al: Calcium signaling around Mitochondria Associated
Membranes (MAMs). Cell Commun Signal. 9:192011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Smaili SS, Hsu YT, Carvalho AC, Rosenstock
TR, Sharpe JC and Youle RJ: Mitochondria, calcium and pro-apoptotic
proteins as mediators in cell death signaling. Braz J Med Biol Res.
36:183–190. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ermak G and Davies KJ: Calcium and
oxidative stress: From cell signaling to cell death. Mol Immunol.
38:713–721. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lewis A, Hayashi T, Su TP and Betenbaugh
MJ: Bcl-2 family in inter-organelle modulation of calcium
signaling; roles in bioen-ergetics and cell survival. J Bioenerg
Biomembr. 46:1–15. 2014. View Article : Google Scholar
|
|
46
|
Annunziata I and d'Azzo A: Interorganellar
membrane microdomains: Dynamic platforms in the control of calcium
signaling and apoptosis. Cells. 2:574–590. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Krebs J, Agellon LB and Michalak M: Ca(2+)
homeostasis and endoplasmic reticulum (ER) stress: An integrated
view of calcium signaling. Biochem Biophys Res Commun. 460:114–121.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Balaban RS: The role of Ca(2+) signaling
in the coordination of mitochondrial ATP production with cardiac
work. Biochim Biophys Acta. 1787:1334–1341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Malli R, Frieden M, Trenker M and Graier
WF: The role of mitochondria for Ca2+ refilling of the
endoplasmic reticulum. J Biol Chem. 280:12114–12122. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dahlem YA, Wolf G, Siemen D and Horn TF:
Combined modulation of the mitochondrial ATP-dependent potassium
channel and the permeability transition pore causes prolongation of
the biphasic calcium dynamics. Cell Calcium. 39:387–400. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Deniaud A, Sharaf el Dein O, Maillier E,
Poncet D, Kroemer G, Lemaire C and Brenner C: Endoplasmic reticulum
stress induces calcium-dependent permeability transition,
mitochondrial outer membrane permeabilization and apoptosis.
Oncogene. 27:285–299. 2008. View Article : Google Scholar
|
|
52
|
Ma X, Tian X, Huang X, Yan F and Qiao D:
Resveratrol-induced mitochondrial dysfunction and apoptosis are
associated with Ca2+ and mCICR-mediated MPT activation
in HepG2 cells. Mol Cell Biochem. 302:99–109. 2007. View Article : Google Scholar : PubMed/NCBI
|