Introduction
Due to the development of modern healthcare and
changes in dietary habits, gastric cancer (GC) is the fifth most
common lethal neoplasm (1), but it
remains the third leading cause of cancer-associated mortalities
worldwide, following lung and liver cancer (1,2).
Statistically, as an aggressive disease, ~950,000 new cases are
diagnosed each year and >720,000 cases of mortality have been
reported for GC in Europe (3). In
China, patients with GC account for ~42% of GC cases worldwide.
Despite improvements in surgical and oncological treatments, the
5-year survival rate for patients with GC remains low, reported as
<30% in developed countries (2). The mechanisms leading to the
occurrence and development of GC remain to be investigated.
MicroRNAs (miRNA/miRs) are short, ~22
nucleotides-long, non-coding RNA molecules, that negatively
regulate transcription via sequence-specific interactions with the
3′-untranslated regions (UTRs) of target genes (4,5).
Various studies have indicated that miRNAs serve a critical role in
tumorigenesis and development by targeting different genes
(5-9) and many are involved in controlling
GC-associated cellular processes, including proliferation,
migration, differentiation, apoptosis and cell cycle progression
(5,10-13).
As comprehensive care strategies for GC are improving, but still
remain unsatisfactory (14,15),
there is an urgency to determine and understand the molecular
mechanisms underlying GC progression. Thus, in the present study,
the role miRNA may serve in the development of GC was explored.
miR-3664-5P is a novel miRNA and its function in
tumors, particularly GC, remains to be investigated. According to
the TargetScan database (mirdb.org/cgi-bin/search.cgi), miR-3664-5P matches
with the mRNA sequence of metadherin (MTDH) without mismatch, thus
ranking first to target MTDH. MTDH, which is also known as
astrocyte elevated gene-1, 3D3 or LYRIC, is a transmembrane protein
that is induced in primary human fetal astrocytes infected with
human immunodeficiency virus (HIV)-1 or treated with recombinant
HIV-1 envelope glycoprotein (16).
MTDH has been recognized as an oncogene in various cancers,
including glioma, melanoma and neuroblastoma, and in carcinomas of
the breast, prostate, liver and esophagus, for its significant role
in promoting the proliferation, angiogenesis, invasion, metastasis
and chemoresistance of cancer cells via activation of the nuclear
factor (NF)-κB, phosphoinositide 3-kinase/protein kinase B,
mitogen-activated protein kinase and Wnt signaling pathways
(17–24). In GC, MTDH was demonstrated to
promote progression using a positive feedback Toll-like receptor
4/NF-κB signaling-associated mechanism, which is widely recognized
to serve a crucial role in inflammation and carcinogenesis
(23,25). Therefore, the aim of the present
study was to determine the role of miR-3664-5P in GC, and
investigated whether its function was associated with targeting
MTDH and regulating the NF-κB signaling pathway.
Materials and methods
Patients and tumor tissues
In the present study, 100 paired fresh GC and
adjacent normal tissues were collected from patients with GC (27-87
years old), who underwent resection of GC without local or systemic
treatment prior to operation at the Department of Gastrointestinal
Surgery in Changzhou No. 2 People's Hospital (Jiangsu, China)
between July 2010 and October 2012. All patients, consisting of 71
males and 29 females, were aged between 27 and 87 years old
(average age, 61.9 years). Patients provided written informed
consent and the experimental procedures were approved by the Ethics
Committee of the Nanjing Medical University Affiliated Changzhou
No. 2 People's Hospital (approval no. 2010-SR-077.A1). All
specimens were confirmed by clinical, radiographic and histological
examination for GC. Clinicopathological data such as age and
gender, as well as lymph node metastasis status, clinical stage,
differentiation, tumor size and T classification were obtained by
reviewing the patients' pathology records. Clinical stage was
determined according to the American Joint Committee on Cancer
staging criteria (26).
Cell lines
All GC cell lines (BGC823, MGC803, SGC7901, AGS and
MKN45) and normal epithelial gastric cells (GES-1) were obtained
from Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China). GC cell
lines were cultured to 70-80% confluence for subsequent experiments
in RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.) and 80 U/ml penicillin at 37°C in humidified air containing
5% CO2. To generate higher transfection efficiency and
better results in vitro and in vivo, MGC803 and MKN45
cells were chosen to construct miR-3664-5P overexpression- and
knockdown-cell lines, respectively, due to the highest miR-3664-5P
expression observed in MKN45 cells and the lowest miR-3664-5P
expression observed in MGC803 cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from tissues and cells was isolated using
TRIzol reagent (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
were reverse transcribed using the PrimeScript RT Master mix
(Takara Biotechnology Co., Ltd., Dalian, China) with the following
temperature protocol: 37°C for 15 min and 85°C for 5 sec, then held
at 4°C. Expression levels of miR-3664-5P and MTDH were determined
via qPCR with the SYBR Premix Ex Taq (Takara Biotechnology Co.,
Ltd.) on the ABI Prism 7900HT (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
thermocycling conditions were as follows: Hot-start DNA polymerase
activation at 95°C for 10 min, followed by 40 cycles of 95°C for 15
sec and 60°C for 1 min, and then one cycle of melt curve analysis
at 95°C for 15 sec, 60°C for 1 min and 95°C for 15 sec. Primer
sequences for qPCR were as follows: miR-3664-5P, forward,
5′-GCCGAGAACTCTGTC TTCAC-3′ and reverse, 5′-CTCAACTGGTGTCGTGGA-3′;
MTDH, forward, 5′-AAATGGGCGGACTGTTGAAGT-3′ and reverse,
5′-CTGTTTTGCACTGCTTTAGCAT-3′; GAPDH, forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′; interleukin (IL-8), forward,
5′-GTGCAGAGGGTTGTG GAGAAG TTT-3′ and reverse,
5′-TCACTGGCATCTTCACTGATT CTTG-3′; matrix metalloproteinase 9
(MMP9), forward, 5′-TG TACCGCTATGGTTACACTCG-3′ and reverse,
5′-GGCAGG GACAGTTGCTTCT-3′; vascular endothelial growth factor
(VEGF), forward, 5′-TGCATTCACATTTGTTGTGC-3′ and reverse,
5′-AGACCCTGGTGGACATCTTC-3′. The expression of miR-3664-5P, MTDH,
IL-8, MMP9 and VFGF were normalized to GAPDH and the expression
levels were calculated using 2−ΔΔCq methods (27).
Western blot assays
Radioimmunoprecipitation Assay buffer containing
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology, Haimen, China) was used to extract proteins from
cells and tissues according to the manufacturer's protocol. The
protein concentration was determined using a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology). Proteins
(40 µg) were then separated by 10% SDS-PAGE and transferred
onto polyvinylidene fluoride membranes (EMD Millipore, Billerica,
MA, USA). Then, the membranes were blocked in 5% non-fat milk for 2
h at room temperature and incubated with specific primary
antibodies at 4°C overnight. Following washing with 0.02 mmol/l
TBST (0.1% Tween; 3×10 min), the membranes were then incubated with
secondary antibodies (anti-rabbit or anti-mouse) at room
temperature for 2 h. The immunoreactive protein bands were
visualized using Enhance Chemiluminescence Plus (EMD Millipore)
with a bio-imaging system. The primary and secondary antibodies
employed in the present study were as follows: MTDH (1:1,000; cat.
no. ab45338; Abcam, Cambridge, UK), NF-κB inhibitor α (IκBα;
1:1,000; cat. no. 4814; Cell Signaling Technology, Inc., Danvers,
MA, USA), p65 (1:1,000; cat. no. 8242; Cell Signaling Technology,
Inc.), phosphorylated (p)-p65 (1:2,000; cat. no. 3033; Cell
Signaling Technology, Inc.), GAPDH (1:1,000; cat. no. 5174; Cell
Signaling Technology, Inc.), anti-rabbit horseradish peroxidase
(HRP)-conjugated secondary antibodies (1:5,000; cat. no. 7074; Cell
Signaling Technology, Inc.) and anti-mouse HRP-conjugated secondary
antibodies (1:5,000; cat. no. 7076; Cell Signaling Technology,
Inc.). ImageJ software 1.47 (National Institutes of Health,
Bethesda, MD, USA) was applied to quantify the integrated density
of the bands.
Knockdown and overexpression of
miRNA
miR-3664-5P mimics (5′-AACUCUGUCUUCACUCAUGAGU-3′)
and its negative control (NC) (5′-UUGUACUACACAAAAGUA CUG-3′), and
miR-3664-5P inhibitor (Sponge sequence:
5′-ACUCAUGAGUGAAGACAGAGUU-3′) and its NC (5′-UU
CUCCGAACGUGUCACGUAA-3′) were synthesized by Nanjing KeyGen Biotech
Co., Ltd. The miRNA mimics (50 nM) or inhibitor (100 nM), and their
NCs (50 and 100 nM, respectively), were transfected into GC cell
lines (4×105/well) using Lipofectamine® 3000
(5 µl/well; Invitrogen; Thermo Fisher Scientific, Inc.).
Cells were used for subsequent experiments at 48 h
post-transfection.
Knockdown of MTDH
Small interfering (si)-RNA targeting MTDH
(5′-AGCCGUAAUCAACCCUAUAUC-3′) and a NC sequence
(5′-UUCUCCGAACGUGUCACGUAA-3′) were obtained from Shanghai
GenePharma Co., Ltd. (Shanghai, China). siRNAs (50 nM) were
transfected into cells (4×105/well) in 6-well plates
using Lipofectamine® 3000 (5 µl/well; Invitrogen;
Thermo Fisher Scientific, Inc.). Cells were used for subsequent
experiments at 48 h post-transfection.
Cell viability assay
To determine the effect of miR-3664-5P on the
proliferation of GC cells (MKN45 and MGC803), a Cell Counting Kit-8
(CCK-8) kit (Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
and a Cell-Light 5-Ethynyl-2′-deoxyuridine (EdU) Apollo567 In
vitro kit (Guangzhou RiboBio Co., Ltd., Guangzhou, China) were
utilized. For CCK-8 detection, 2×103 cells/well were
cultured in 96-well plates, each well containing 100 µl
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.) with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 80 U/ml
penicillin. Following 24, 48, 72 and 96 h, 10 µl CCK-8 assay
reagent was added to each well mixed with 90 µl serum-free
medium. The absorbance was then measured 2 h later using a
microplate reader at 450 nm, with 630 nm as reference wavelength.
For the EdU assay, cells (2×105) were seeded into Glass
Bottom Cell Culture Dishes (Wuxi Nest Biotechnology, Co., Ltd.,
Wuxi, China), then the cells were treated according to
manufacturer's instructions. Finally, images were captured under a
laser confocal scanning microscope (Carl Zeiss AG, Oberkochen,
Germany).
Plate colony formation assay
Cells (1,000/well) were cultured in 6-well plates to
investigate the effect of miR-3664-5P on the efficiency of colony
formation. Following 7 days, each well was washed with PBS three
times at room temperature. The cells were then fixed using ethyl
alcohol for 30 sec and stained for 20 min at room temperature using
crystal violet dye. Following washing with PBS, colonies (≥50
cells/colony) in each well were manually counted and images were
captured using a digital camera (Canon DS126211; Canon, Inc.,
Tokyo, Japan).
Transwell assay
Cell migration and invasion were analyzed in 24-well
plates with sterile polycarbonate chambers (8-µm filters; BD
Biosciences, Franklin Lakes, NJ, USA) with or without Matrigel
coating. Cells (2×104) were cultured in 100 µl
serum-free RPMI-1640 in the upper chamber and 600 µl
RPMI-1640 plus 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) in the lower chamber. Following 24 h, the lower
chamber was washed twice with PBS and stained with crystal violet
dye for 20 min. The chamber was then washed again with PBS three
times, and a cotton bud was used to remove cells and medium from
the upper chambers. The migrated and invaded cells in the lower
chambers were observed under a light microscope (NIKON ECLIPSE
TI-SR; Nikon Corporation, Tokyo, Japan; magnification, x200).
Flow cytometry analysis of cell cycle and
apoptosis
Cell cycle distribution was determined with a Cell
Cycle Assay kit (Vazyme, Piscataway, NJ, USA) according to the
manufacturer's instructions. Cells (6×105) were cultured
in 6-well plates and treated with 50 µM
H2O2 for 2 h at 37°C to induce apoptosis and
were then analyzed using an Annexin V-fluorescein
isothiocyanate/propidium iodide Apoptosis Detection kit (Vazyme).
Cell cycle distribution and the levels of apoptosis were analyzed
using a flow cytometer equipped with BD FACSDiva Software 6.0 (BD
Biosciences).
Animals and subcutaneous tumor growth
assays
A total of 20 male BALB/c nude mice (age, 4-8 weeks;
13-15 g) were purchased from the Laboratory Animal Center of
Yangzhou University (Yangzhou, China) and kept under specific
pathogen-free conditions. All experiments were approved by Animal
Ethics Committee of Nanjing Medical University (Nanjing, China).
Mice were maintained at room temperature (20-26°C) with a humidity
level of 50-60% and a 12-h light/dark cycle, and were bred under
specific pathogen-free conditions in accordance with the
institutional animal care and use committee regulations. Mice
received 5 g food and 100 ml water per 100 g body weight per day.
MGC803 cells (1×107) transfected with the miR-3664-5P
overexpression lentivirus (left groin) and NC (right groin) were
suspended in 100 µl PBS and injected subcutaneously into
nude mice. The tumor sizes were measured every five days using
micrometer calipers, and tumor volumes were calculated as follows:
Tumor volume = (width2 × length)/2. Following 30 days,
the mice were sacrificed by carbon dioxide inhalation (air
displacement rate: 20%/min; Yuyan Instruments Company, Shanghai,
China). All animal experiments were performed in compliance with
the relevant ethical regulations per an approved Institutional
Animal Care and Use Committee protocol, including a tumor size
limit of 2.0 cm in any dimension. The maximum tumor volume observed
in the mice was 1.0285 cm3 [(1.12 × 1.7)/2].
The maximum observed combined diameter in the cases where multiple
tumors occurred was 1.9 cm.
Lung metastasis model
A single-cell suspension was prepared with cells
stably overexpressing miR-3664-5P or NC lentivirus, suspended in
200 µl PBS and filtered through a sterile 70-µm nylon
mesh filter (BD Biosciences). Mice were separately injected with
MGC803 cells (5×106) with or without miR-3664-5P
overexpression via the tail vein to develop peripheral
intravascular-implanted models (10 mice/group). Mice were
sacrificed 5 weeks later and tumor metastases in the lung were
examined. Hematoxylin and eosin staining was used to evaluate the
number of tumors in lung tissues. Briefly, lung tissues were fixed
in 10% formalin at room temperature for 24 h, dehydrated with
different concentrations of alcohol (70, 85, 95 and 100%) and
xylene, embedded in paraffin and sliced into 4-µm sections.
Following deparaffinization in xylene and rehydration in different
concentrations of alcohol (100, 95, 85 and 70%) and distilled
water, sections were stained with hematoxylin for 5 min and eosin
for 2 min at room temperature, dehydrated with different
concentrations of alcohol (70, 85, 95 and 100%) and xylene, and
covered with a coverslip with mounting medium (Sangon Biotech Co.,
Ltd., Shanghai, China). The sections were viewed and images were
captured under a light microscope (NIKON ECLIPSE TI-SR; Nikon
Corporation; magnification, x200).
Immunohistochemistry (IHC)
For IHC, the UltraSensitive™ SP (Mouse/Rabbit) IHC
kit (Fuzhou Maixin Biotech Co., Ltd., Fuzhou, China) was used with
the streptavidin-biotin amplification method. Briefly, human
gastric cancer, paired adjacent normal and mice subcutaneous tumor
tissues were fixed in 10% formalin at room temperature for 24 h,
dehydrated with different concentrations of alcohol (70, 85, 95 and
100%) and xylene, embedded in paraffin and sliced into 4-µm
sections. Following deparaffinization in xylene and rehydration in
different concentrations of alcohol (100, 95, 85 and 70%) and
distilled water, sections were blocked with 0.3% hydrogen peroxide
and incubated with 0.03 M citrate buffer (pH 6.0) in a pressure
cooker at 121°C. Nonspecific tissue reactions were blocked using
10% bovine serum (Servicebio, Woburn, MA, USA) for 30 min at room
temperature and sections were incubated with the anti-MTDH (LYRIC)
primary antibody (1:300; cat. no. ab45338; Abcam) overnight at 4°C,
followed by incubation with horseradish peroxidase-conjugated
secondary antibody (1:1,000; polyclonal, cat. no. ab6721; Abcam)
for 15 min for at room temperature. Sections were then visualized
using 3,3′-diaminobenzidine solution (Fuzhou Maixin Biotech Co.,
Ltd.) for 1 min at room temperature. Following dehydration,
transparency and sealing, the sections were viewed and images were
captured under a light microscope (NIKON ECLIPSE TI-SR; Nikon
Corporation; magnification, x200).
Bioinformatics analysis and Luciferase
activity reporter assay
Target genes of miR-3664-5P were predicted using the
TargetScan database (www.targetscan.org; release no. 7.2) and miRDB
database (www.mirdb.org) by searching for the
presence of conserved orthologous 3′-UTRs of humans that matched
the seed region of miR-2664-5P (28,29).
The luciferase reporter assay was carried out using pDL-reporter
vectors (Guangzhou RiboBio Co.) containing the wild-type (WT) or
mutant (Mut) MTDH 3′-UTR sequences. Cells (1×105/well)
in a 24-well plate were cotransfected with miR-3664-5P
mimics/inhibitors or the corresponding controls along with the
WT/Mut MTDH 3′-UTR vectors using Lipofectamine® 3000 (1
µl/well; Invitrogen; Thermo Fisher Scientific, Inc.). The
luciferase activity was measured using a dual-luciferase assay kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions at 48 h post-transfection; luciferase
activities were normalized to Renilla luciferase activities as the
internal standard of transfection efficiency.
Statistical analysis
Patients were divided into low and high miR-3664-5P
expression groups according to the median RT-qPCR results.
Correlations between miR-3664-5P expression and various
clinicopathological parameters were analyzed by χ2 test.
Student's t-test was applied to evaluate the significance between
two samples. The expression levels of miR-3664-5P and MTDH in tumor
and normal tissues were analyzed via paired T tests, while the
tumor weight as well as the number of metastasis lesions in
vivo were analyzed using unpaired T test. One-way analysis of
variance and least-significant difference post hoc tests were used
to compare datasets containing multiple groups. The log rank test
was employed in the analysis of Kaplan-Meier curves. Clinical
characteristics that exhibited significant associations with
survival in univariate analyses (P<0.05) were entered into
multivariate analyses, performed using the Cox proportional hazard
model. Receiver operating characteristic (ROC) curve analysis was
used to detect the diagnostic efficiency of miR-3664-5P in GC. The
area under the curve was calculated, and the optimum sensitivity
and specificity were determined using the Youden index. All
statistical analyses were conducted with SPSS 17.0 (SPSS, Inc.,
Chicago, IL, USA) and GraphPad Prism 5 (GraphPad Software, Inc., La
Jolla, CA, USA). The results of the in vitro experiments
were presented as the mean ± standard error of the mean from three
independent experiments, while data obtained from the in
vitro experiments were presented as the mean ± standard
deviation. P<0.05 was considered to indicate a statistically
significant difference.
Results
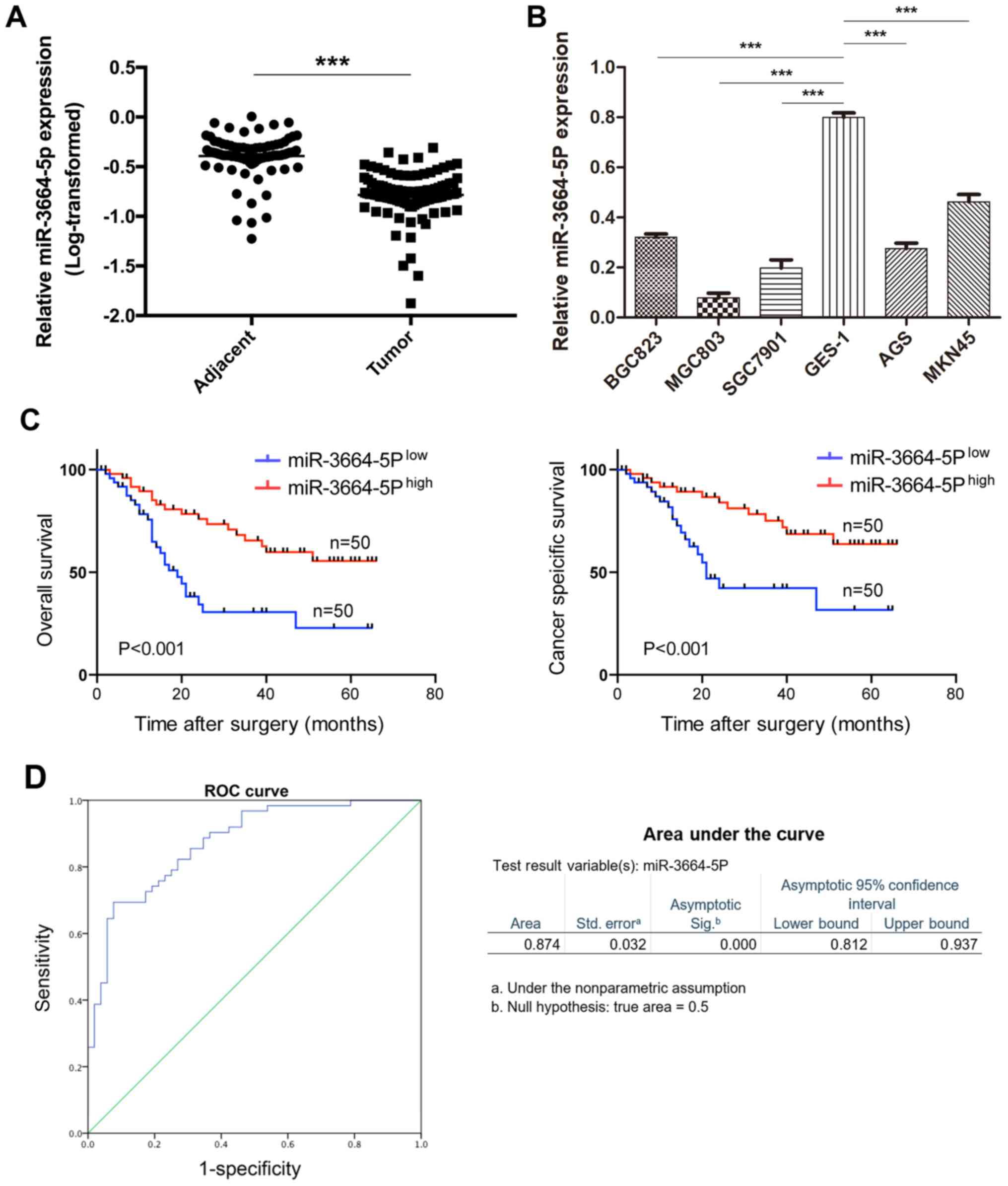
miR-3664-5P is significantly
downregulated in GC tissues and cell lines
To help understand the role of miR-3664-5P in GC,
RT-qPCR was performed in 100 GC and adjacent normal tissues, which
demonstrated that when compared with the normal tissues,
miR-3664-5P expression was significantly downregulated in GC
tissues (P<0.001; Fig. 1A).
Similar results were observed in the GC cell lines when compared
with the normal gastric epithelial cell line GES-1; miR-3664-5P
expression was suppressed in the BGC823, MGC803, SGC7901, AGS and
MKN45 GC cell lines (P<0.001; Fig.
1B).
High levels of miR-3664-5P are associated
with favorable prognosis in patients with GC
To further investigate the clinical relevance of
miR-3664-5P and its prognostic value in GC, patients were divided
into high and low miR-3664-5P expression groups according to the
median expression level as determined by RT-qPCR. Expression of
miR-3664-5P was correlated with differentiation (P=0.016) and tumor
size (P=0.016; Table I). However,
no significant association was identified when comparing sex, age,
clinical stage, lymph node metastasis and T classification. In
addition, patients with high miR-3664-5P expression had a higher
probability of a better overall (P<0.001) and cancer-specific
prognosis (P<0.001) compared with the low miR-3664-5P expression
group (Fig. 1C). Cox proportional
hazards regression analyses suggested that miR-3664-5P expression
was an independent prognostic predictor for overall survival
[hazard ratio (HR)=0.492; P=0.029] and cancer specific survival
(HR=0.038; P=0.01; Tables II and
III). ROC curve analysis was
performed to investigate the effectiveness of miR-3664-5P for GC
prediction. Serum samples of GC patients collected prior to
resections of GC (n=60) and control serum samples collected from
people undergoing physical examinations (n=40) were utilized. The
expression of miR-3664-5P in serum was detected by RT-qPCR. The
results indicated that miR-3664-5P may be an effective predictor
for GC diagnosis with a sensitivity of 0.923 and a specificity of
0.694 (Fig. 1D). These results
suggested that miR-3664-5P may serve a critical role in GC
development, and serve as a biomarker for GC diagnosis and
prognosis.
 | Table IAssociations between miR-3664-5P
expression and clinicopathological characteristics of patients with
GC (n=100). |
Table I
Associations between miR-3664-5P
expression and clinicopathological characteristics of patients with
GC (n=100).
| Clinicopathological
features | Total n
(n=100) | miR-3664-5P
expression
| P-value |
|---|
Low
| High
|
|---|
| No. of patients
(%) | No. of patients
(%) |
|---|
| Sex | | | | 0.826 |
| Male | 71 | 36 (72.0) | 35 (70.0) | |
| Female | 29 | 14 (28.0) | 15 (30.0) | |
| Age, years | | | | 0.406 |
| ≤50 | 64 | 30 (60.0) | 34 (68.0) | |
| >50 | 36 | 20 (40.0) | 16 (32.0) | |
|
Differentiation | | | | 0.016a |
| Well/
moderately | 56 | 22 (44.0) | 34 (68.0) | |
| Poor | 44 | 28 (56.0) | 16 (32.0) | |
| Clinical stage | | | | 0.069 |
| I-II | 43 | 26 (52.0) | 17 (34.0) | |
| III-IV | 57 | 24 (48.0) | 33 (66.0) | |
| Tumor size
(cm) | | | | 0.016a |
| ≤3.5 | 50 | 19 (38.0) | 31 (52.0) | |
| >3.5 | 50 | 31 (52.0) | 19 (38.0) | |
| Lymph node
metastasis | | | | 0.420 |
| No | 44 | 20 (40.0) | 24 (48.0) | |
| Yes | 56 | 30 (60.0) | 26 (52.0) | |
| T
classification | | | | 0.838 |
|
T1-T2 | 61 | 31 (62.0) | 30 (60.0) | |
|
T3-T4 | 39 | 19 (38.0) | 20 (40.0) | |
| Table IIUnivariate and multivariate survival
analyses evaluating miR-3664-5P expression influencing overall
survival in gastric cancer (n=100). |
Table II
Univariate and multivariate survival
analyses evaluating miR-3664-5P expression influencing overall
survival in gastric cancer (n=100).
| Variables | Univariate
| Multivariate
analysis
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Sex | | 0.427 | | NA |
| Male | 1 | | | |
| Female | 0.770
(0.404-1.468) | | | |
| Age, years | | 0.389 | | NA |
| ≤50 | 1 | | | |
| >50 | 0.753
(0.395-1.436) | | | |
|
Differentiation | | 0.010b | | 0.013a |
| Well/
moderately | 1 | | 1 | |
| Poor | 2.176
(1.207-3.923) | | 2.201
(1.178-4.113) | |
| Clinical stage | | 0.188 | | NA |
| I-II | 1 | | | |
| III-IV | 1.501
(0.820-2.750) | | | |
| Tumor size, cm | | 0.041a | | 0.170 |
| ≤3.5 | 1 | | 1 | |
| >3.5 | 1.867
(1.027-3.396) | | 1.546
(0.829-2.883) | |
| Lymph node
metastasis | | 0.009b | | 0.003b |
| No | 1 | | 1 | |
| Yes | 2.252
(1.225-4.139) | | 2.601
(1.379-4.907) | |
| T
classification | | 0.372 | | NA |
|
T1-T2 | 1 | | | |
|
T3-T4 | 1.306
(0.726-2.350) | | | |
| miR-3664-5P
expression | | 0.001c | | 0.029a |
| Low | 1 | | 1 | |
| High | 0.360
(0.195-0.666) | | 0.492
(0.260-0.931) | |
| Table IIIUnivariate and multivariate survival
analyses evaluating miR‐3664‐5P expression influencing cancer
specific survival in gastric cancer (n=100). |
Table III
Univariate and multivariate survival
analyses evaluating miR‐3664‐5P expression influencing cancer
specific survival in gastric cancer (n=100).
| Variables | Univariate
| Multivariate
analysis
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Sex | | 0.880 | | NA |
| Male | 1 | | | |
| Female | 0.945
(0.458-1.952) | | | |
| Age, years | | 0.362 | | NA |
| ≤50 | 1 | | | |
| >50 | 0.700
(0.325-1.507) | | | |
|
Differentiation | | 0.006b | | 0.024a |
|
Well/moderately | 1 | | 1 | |
| Poor | 2.625
(1.310-5.260) | | 2.257
(1.114-4.573) | |
| Clinical stage | | 0.536 | | NA |
| I-II | 1 | | | |
| III-IV | 1.245
(0.622-2.493) | | | |
| Tumor size, cm | | 0.163 | | NA |
| ≤3.5 | 1 | | | |
| >3.5 | 1.641
(0.818-3.288) | | | |
| Lymph node
metastasis | | 0.150 | | NA |
| No | 1 | | | |
| Yes | 1.663
(0.832-3.326) | | | |
| T
classification | | 0.790 | | NA |
|
T1-T2 | 1 | | | |
|
T3-T4 | 1.009
(0.550-2.196) | | | |
| miR-3664-5P
expression | | 0.003b | | 0.010b |
| Low | 1 | | 1 | |
| High | 0.334
(0.163-0.686) | | 0.380
(0.183-0.791) | |
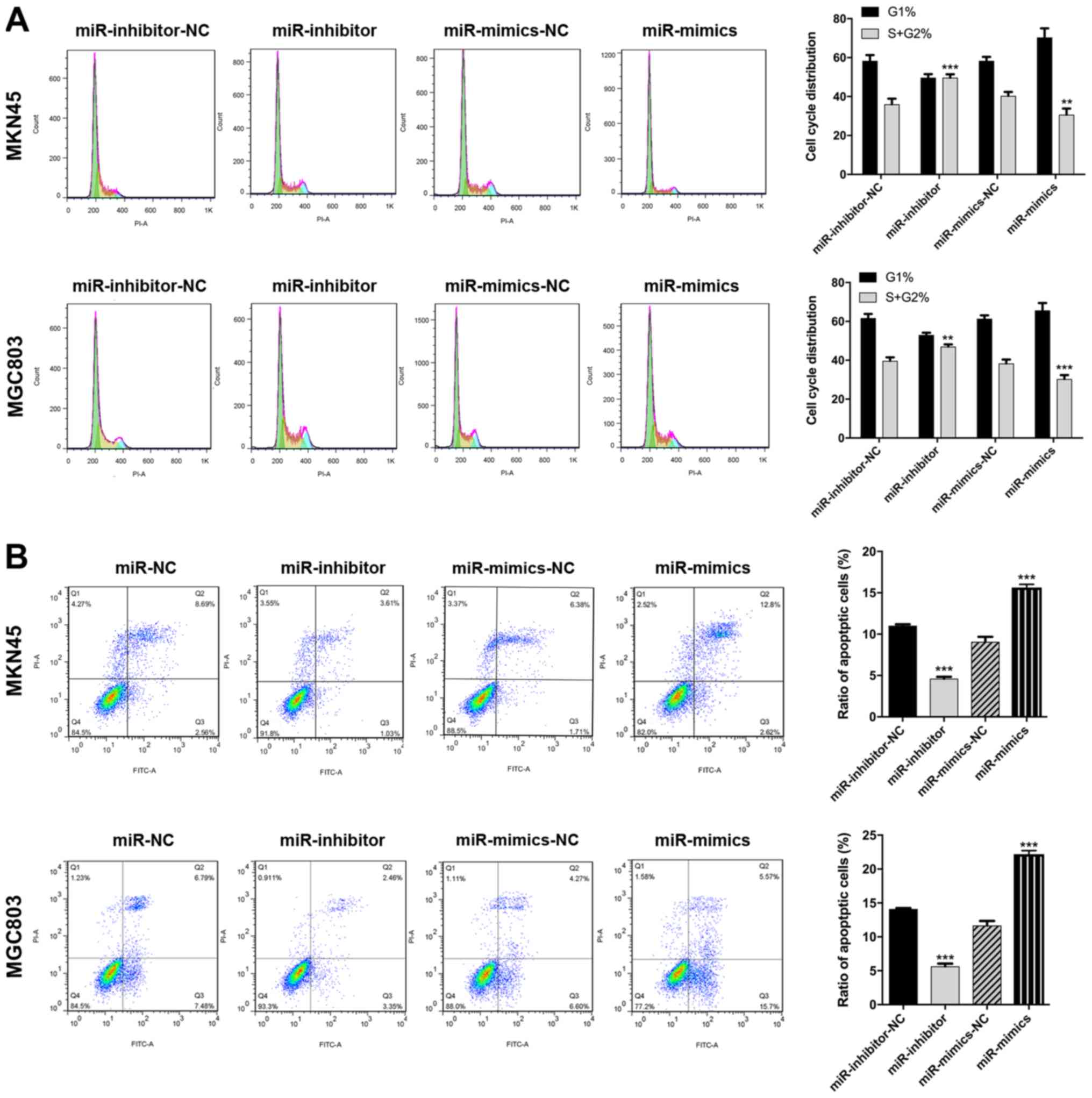
miR-3664-5P inhibits GC cell
proliferation, migration and invasion in vitro
MGC803 and MKN45 cells were chosen to construct
miR-3664-5P overexpression- and knockdown-cell lines, respectively,
due to their relatively low and high miR-3664-5P expression. The
miR-3664-5P inhibitors and mimics were transfected into MGC803 and
MKN45 cells in order to establish miR-3664-5P overexpression and
knockdown GC cell lines. The miR-3664-5P overexpression cell line
for in vivo experiments was constructed by transfecting a
lentivirus-plasmid into MGC803 cells to induce miR-3664-5P
overexpression. The transfection efficiency was confirmed by
RT-qPCR (P<0.001; Fig. 2A). It
was demonstrated that miR-3664-5P upregulation inhibited GC cell
proliferation and migration, and miR-3664-5P downregulation had the
reverse effect on CCK8, EdU and plate colony assays (P<0.01;
Fig. 2B–D). Flow cytometry assays
were performed to evaluate the effects of miR-3664-5P regulation on
apoptosis and the cell cycle. The results revealed that miR-3664-5P
overexpression in GC cells resulted in significant promotion of
apoptosis and cell cycle arrest, whereas miR-3664-5P knockdown
significantly suppressed apoptosis and promoted the accumulation of
cells in the S and G2 stage (P<0.05; Fig. 3A and B). The transwell assay
revealed that miR-3664-5P overexpression in decreased and
miR-3664-5P inhibition in increased migration and invasion in the
MGC803 and MKN45 cell lines (P<0.001; Fig. 4A and B). Taken together, the above
results demonstrated that miR-3664-5P inhibited GC cell
proliferation, migration and invasion in vitro.
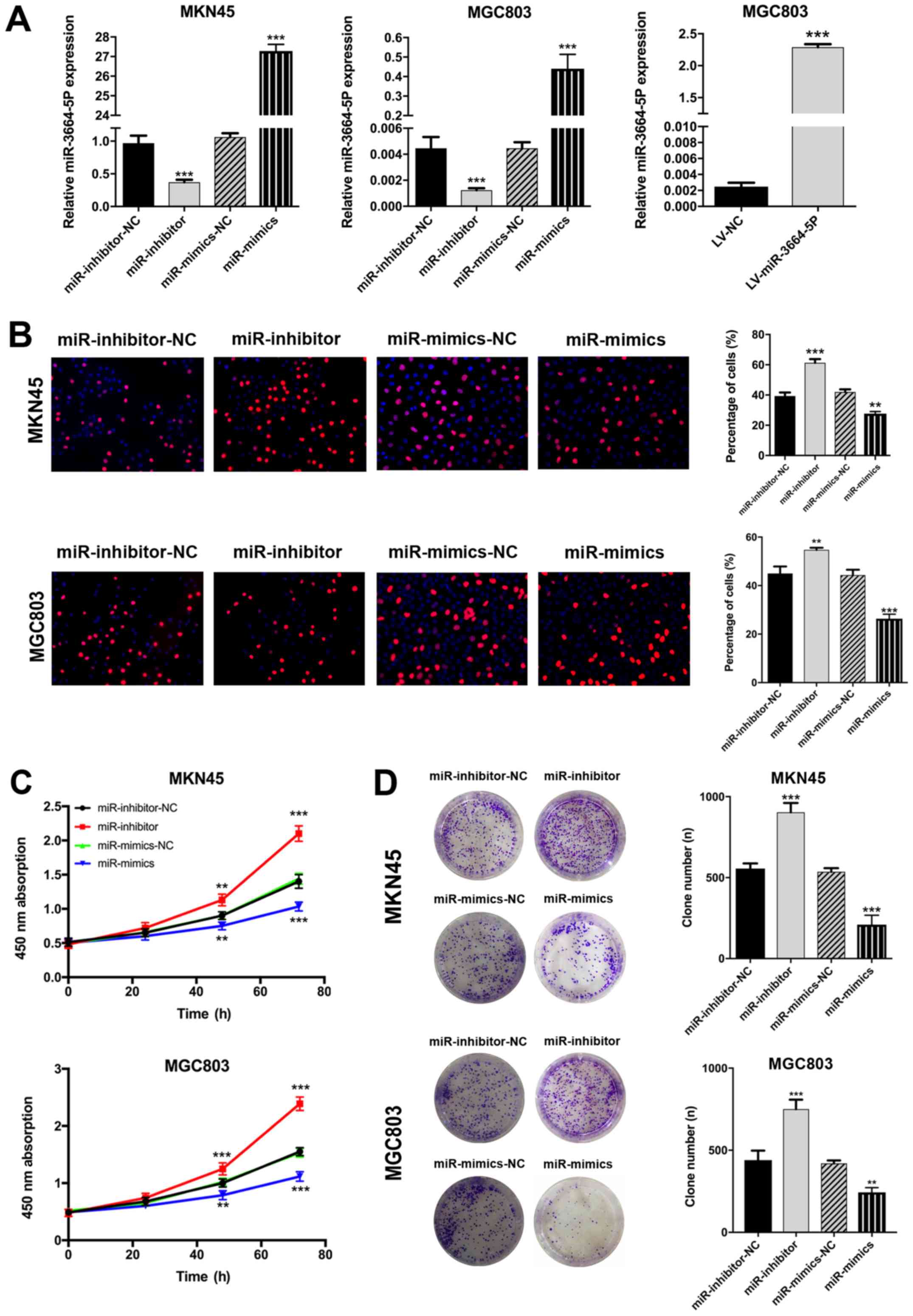 | Figure 2miR-3664-5P inhibits GC cell
proliferation in vitro. (A) Following transfection with
miR-3664-5P mimics or inhibitors, the expression of miR-3664-5P was
significantly upregulated or downregulated, respectively, in the
MKN45 and MGC803 cell lines. MGC803 was transfected with
miR-3664-5P overexpres-sion lentivirus for in vivo assays.
miR-3664-5P levels were determined by reverse
transcription-quantitative polymerase chain reaction. (B) EdU, (C)
Cell Counting Kit-8 and (D) plate colony assays were performed to
detect the viability of GC cell lines (magnification, x200).
miR-3664-5P upregulation inhibited and miR-3664-5P downregulation
promoted the proliferation of GC cells. Data are presented as the
mean ± standard error of the mean, from three independent
experiments. **P<0.01 and ***P<0.001
vs. the corresponding NC. NC, negative control; miR-inhibitor-NC,
cells transfected with the negative control of the miR-3664-5P
inhibitor; miR-inhibitors, cells transfected with miR-3664-5P
inhibitors; miR-mimics-NC, cells transfected with the negative
control of the miR-3664-5P mimics; miR-mimics, cells transfected
with miR-3664-5P mimics; miR, microRNA; GC, gastric cancer; LV,
lentivirus. |
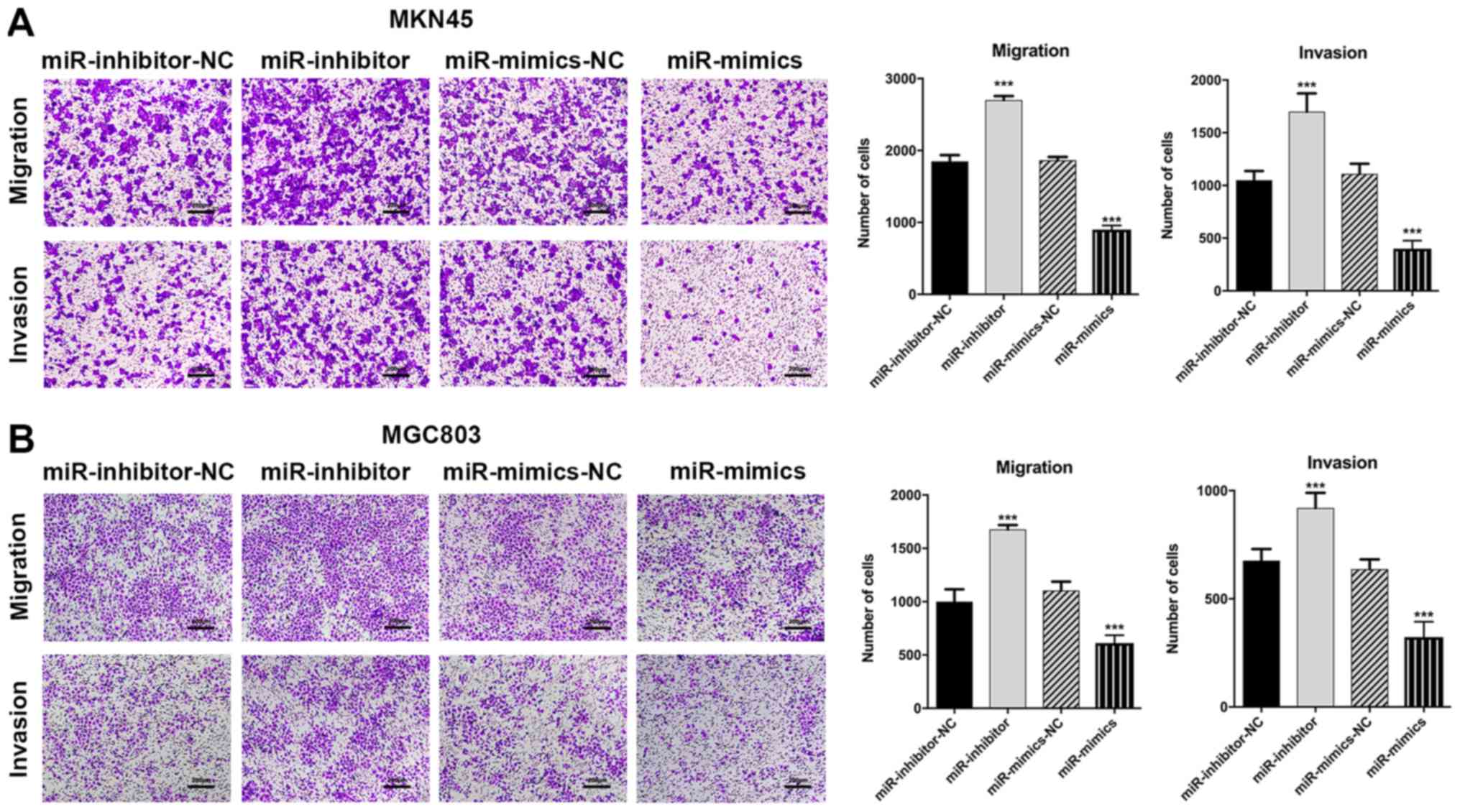 | Figure 4miR-3664-5P inhibits GC cell
migration and invasion in vitro. Transwell assays were
performed to evaluate the migration and invasion abilities of GC
cells. (A and B) The inhibition of miR-3664-5P enhanced, while the
overexpression of miR-3664-5P suppressed, the migration and
invasion of (A) MKN45 and (B) MGC803 cells. Magnification, x200;
scale bars, 200 µm. Data are presented as the mean ±
standard error of the mean, from three independent experiments.
***P<0.001 vs. the corresponding NC. NC, negative
control; miR-inhibitor-NC, cells transfected with the negative
control of the miR-3664-5P inhibitor; miR-inhibitors, cells
transfected with miR-3664-5P inhibitors; miR-mimics-NC, cells
transfected with the negative control of the miR-3664-5P mimics;
miR-mimics, cells transfected with miR-3664-5P mimics; miR,
microRNA; GC, gastric cancer. |
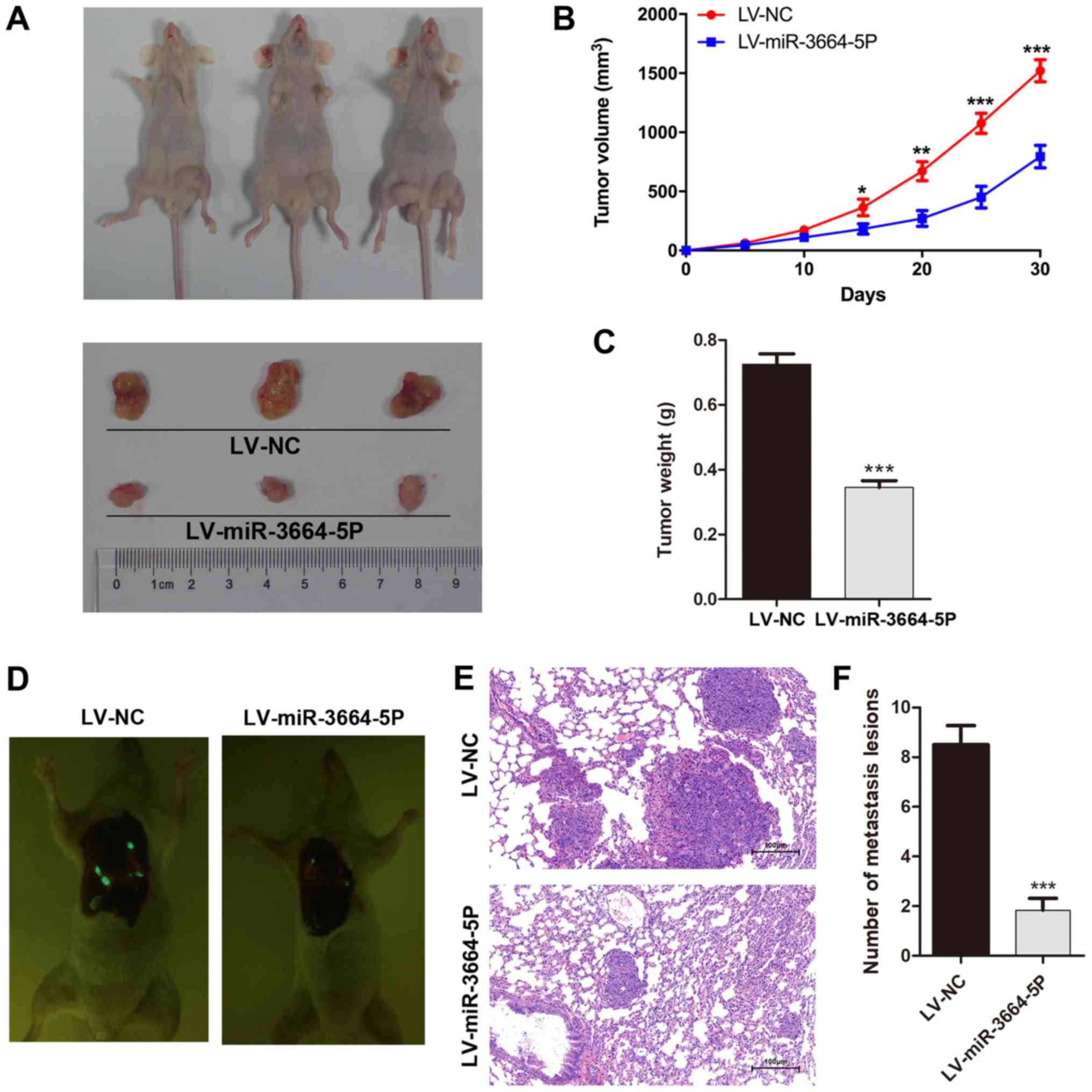
miR-3664-5P acts as an inhibitor of tumor
growth in vivo
A xenotransplantation model was established in order
to gain better understanding of the role of miR-3664-5P in
vivo. MGC803 cells transfected with miR-3664-5P overexpressing
lentivirus were injected into nude mice subcutaneously. The tumor
size was monitored every 5 days until sacrifice. Compared with the
control group, miR-3664-5P overexpression markedly inhibited tumor
growth in vivo (Fig. 5A–C),
verifying the inhibitory role of miR-3664-5P in the proliferation
of GC cells. Consistently, detection of lung metastasis in the nude
mice injected through the tail vein with miR-3664-5P overexpressing
MGC308 or control cells revealed that an increased expression of
miR-3664-5P was significantly associated with reduced lung
metastases (Fig. 5D–F).
MTDH is the functional target of
miR-3664-5P
Sufficient evidence has demonstrated that miRNAs
function by regulating the expression of a target gene (4). To obtain an in-depth understanding of
the association between miR-3664-5P and GC, two bioinformatics
databases (TargetScan, www.targetscan.org; and miRDB, www.mirdb.org) were utilized to predict the target
gene of miR-3664-5P (29,30). The results suggested that MTDH was
the candidate with highest potential for binding sites, including 3
conserved putative target sites for miR-3664-5P (70-91, 3,358-3,379
and 5,000-5,021 nt) (data not shown). MTDH mRNA and protein
expressions were upregulated in tumor tissues when compared with
paired normal tissues (Fig. 6A and
B). A Pearson correlation analysis of 100 GC tissues revealed
that MTDH expression was negatively correlated with miR-3664-5P
expression (Fig. 6C).
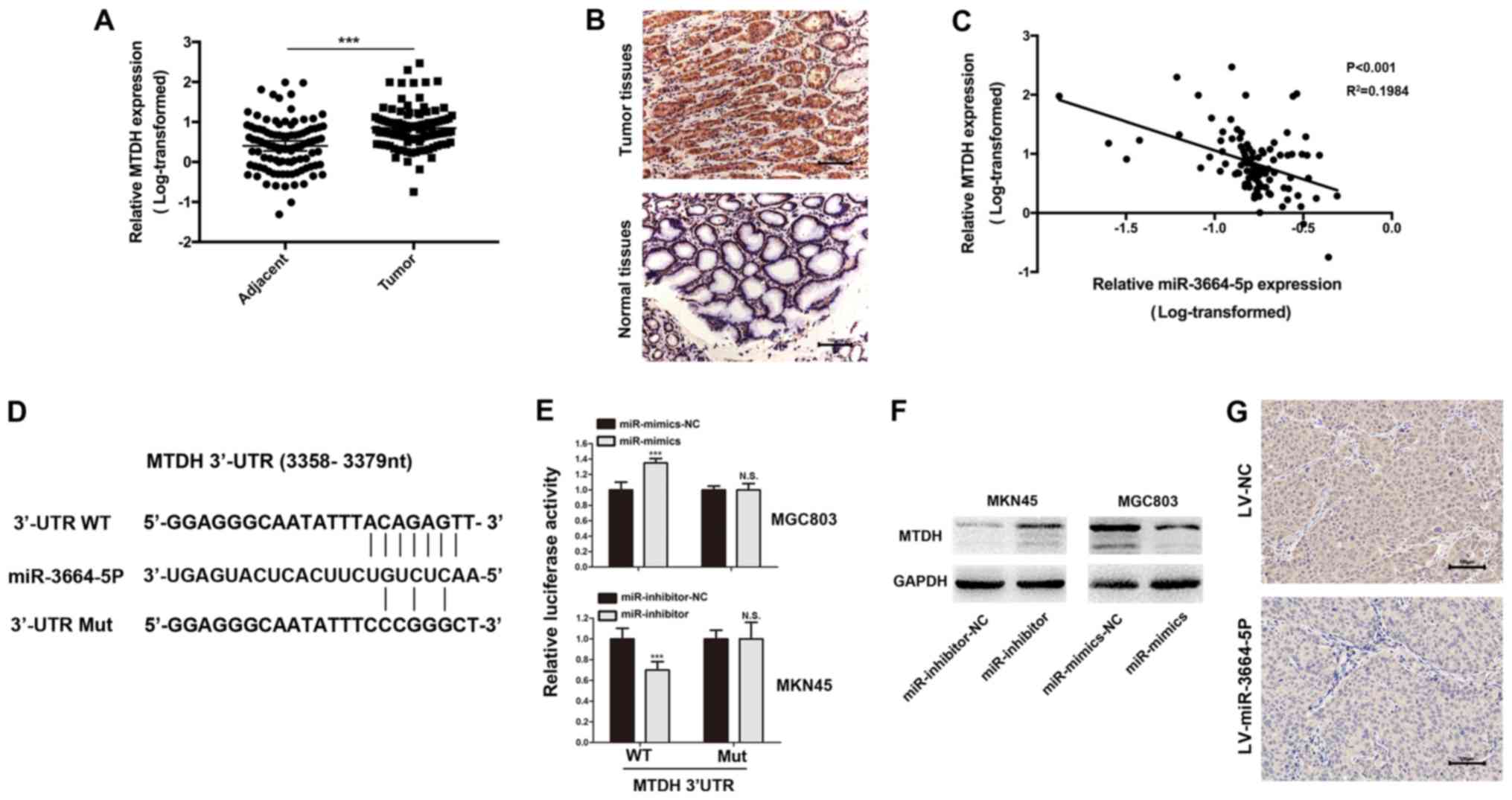 | Figure 6MTDH is the functional target of
miR-3664-5P. (A) MTDH levels were significantly increased in GC
tissues when compared with the control. ***P<0.001,
as indicated. (B) IHC was performed to investigate MTDH expression
in GC and adjacent normal tissues. Magnification, x200; scale bars,
100 µm. (C) Pearson correlation analysis revealed that
miR-3664-5P expression was negatively correlated with MTDH
expression in GC tissues. (D) miR-3664-5P targeted the 3′-UTR of
MTDH and the corresponding mutations of miR-3664-5P were assessed.
(E) Luciferase reporter assays were performed to evaluate
interactions between miR-3664-5P and candidate genes in GC cell
lines. ***P<0.001 vs. the corresponding NC. (F) MTDH
protein levels in GC cells with miR-3664-5P knockdown or
overexpression were detected. (G) IHC was performed in the tumor
tissues collected from the mouse xenotransplantation model.
Magnification, x200; scale bars, 100 µm. Data are presented
as the mean ± standard error of the mean. NC, negative control;
miR-inhibitor-NC, cells transfected with the negative control of
the miR-3664-5P inhibitor; miR-inhibitors, cells transfected with
miR-3664-5P inhibitors; miR-mimics-NC, cells transfected with the
negative control of the miR-3664-5P mimics; miR-mimics, cells
transfected with miR-3664-5P mimics; LV, lentivirus; LV-NC, cells
transfected with NC LV; LV-miR-3664-5P, cells transfected with
miR-3664-5P overexpression LV; IHC, immunohistochemistry; N.S., not
significant; miR, microRNA; GC, gastric cancer; UTR, untranslated
region; Mut, mutant; WT, wild-type; MTDH, metadherin. |
For further verification of the interactions between
miR-3664-5P and the 3′-UTR of MTDH mRNA, three pairs of WT
(pDL-MTDH-30UTR-wt) or Mut (pDL-MTDH-30UTR-mut) reporter vectors
were constructed based on the 3 potential binding sites and
co-transfected with miR-3664-5P mimics or inhibitors into MGC803 or
MKN45 cells, respectively. Among these 3 potential binding sites,
the most pronounced decrease in luciferase activity when compared
with the WT reporter in MGC803 cells was observed for the second
site (3,358-3,379 nt; Fig. 6D and
E). MTDH protein expression in MGC803 cells, where miR-3664-5P
was knocked down, was further investigated. The results suggested
that MTDH expression was decreased in MGC803 cells treated with
miR-3664-5P mimics and increased in MKN45 cells treated with
miR-3664-5P inhibitors (Fig. 6F).
Furthermore, MTDH protein expression was decreased in tumor tissues
transfected with miR-3664-5P overexpressing lentivirus in the
xenotransplantation model (Fig.
6G). Taken together, these results demonstrated that
miR-3664-5P functioned by targeting MTDH.
MTDH promotes the antitumor function of
miR-3664-5P through the NF-κB signaling pathway
Considering a previous study (23), it was hypothesized that miR-3664-5P
may suppress GC by regulating the NF-κB signaling pathway, with
MTDH as the key protein connecting miR-3664-5P and the NF-κB
signaling pathway. Rescue assays in MKN45 cells were performed
utilizing siRNAs targeting MTDH. CCK8, EdU, plate colony and
Transwell assays were performed using MKN45 miR-3664-5P modified
cells. The effect of miR-3664-5P inhibitors on miR-3664-5P
antitumor function was counteracted by the ectopic expression of
MTDH (Fig. 7A–D). Then the
expression levels of certain key proteins of the NF-κB signaling
pathway and further downstream targets, including the intercellular
adhesion molecule IL-8, MMP9 and VEGF, were assessed. The results
revealed a decreased degradation of IκBα and an increased
phosphorylation of p65, which induced the activation of the NF-κB
signaling pathway in miR-3664-5P inhibited MKN45 cells. These
observations were reversed to the pre-treatment expression levels
following co-transfection with MTDH siRNAs (Fig. 7E). The downstream targets of the
NF-κB signaling pathway were significantly upregulated in
miR-3664-5P inhibited MKN45 cells, and MTDH siRNAs reversed these
observations (Fig. 7F). The
results indicated that miR-3664-5P suppressed the proliferation of
GC cells by regulating MTDH expression, and the underlying
mechanism may be associated with targeting the NF-κB signaling
pathway.
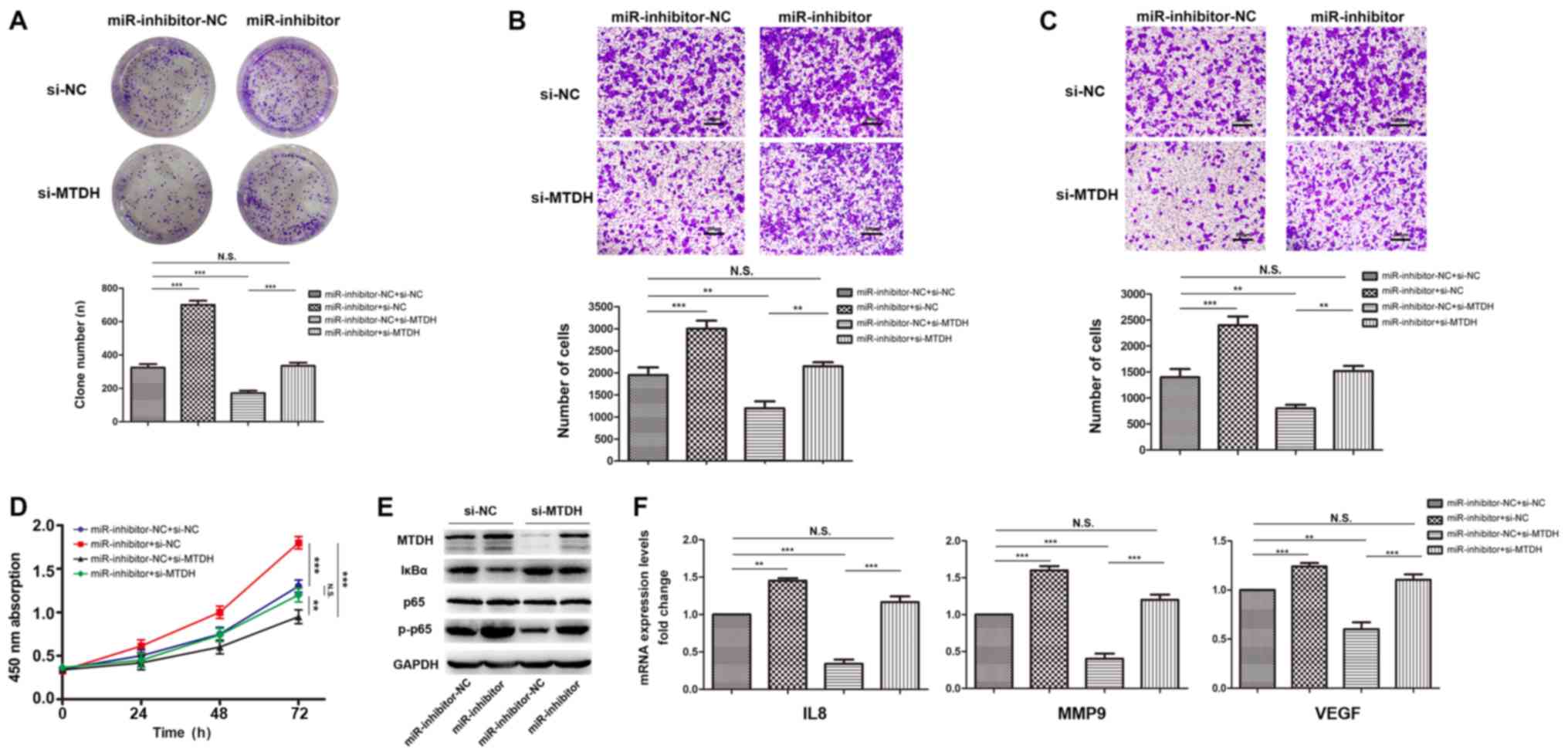 | Figure 7MTDH mediates the antitumor function
of miR-3664-5P through the NF-κB signaling pathway. (A-D)
miR-3664-5P inhibition enhanced the proliferation, migration and
invasion of GC cells as determined by (A) plate colony assays, (B)
transwell migration assays, (C) transwell invasion assays and (D)
Cell Counting Kit-8 assays, which was reversed following
transfection with MTDH siRNA. Magnification, x200; scale bars, 200
µm. (E) MKN45 transfected with miR-3664-5P inhibitors
exhibited significant upregulation of MTDH and key molecules of the
NF-κB signaling. Following transfection with MTDH siRNA, expression
levels were restored to the original (untransfected) levels. (F)
mRNA expression levels of the downstream targets of the NF-κB
signaling pathway were upregulated in miR-3664-5P inhibited MKN45
cells and this effect was reversed following transfection with MTDH
siRNA. Data are presented as the mean ± standard error of the mean,
from three independent experiments. **P<0.01 and
***P<0.001, as indicated. NC, negative control;
miR-inhibitor-NC, cells transfected with the negative control of
the miR-3664-5P inhibitor; miR-inhibitors, cells transfected with
miR-3664-5P inhibitors; si-/ siRNA, small interfering RNA; si-NC,
siRNA of NC; si-MTDH, siRNA targeting MTDH; N.S., not significant;
miR, microRNA; GC, gastric cancer; MTDH, metadherin; NF-κB, nuclear
factor-κB; IκBα, NF-κB inhibitor α; p-, phosphorylated; IL8,
interleukin 8; MMP9, matrix metalloproteinase; VEGF, vascular
endothelial growth factor. |
Discussion
The tumorigenesis of GC is a complicated process
that has not been completely elucidated. Among the associated risk
factors, familial inheritance and Helicobacter pylori (H.
pylori) infection account for the largest proportion, followed
by chronic atrophic gastritis, including pernicious anemia, toxic
and dietary agents, previous gastric surgery with bile reflux,
hypertrophic gastropathy including Metenier's disease, gastric
polyps, low socioeconomic status and obesity (12,31-33).
In developing countries particularly, including China, H.
pylori infection may be the greatest risk factor of GC
(25,32). It is widely accepted that H.
pylori infection promotes the genesis of GC by enhancing the
production of free radicals and long-term inflammation of the
gastric mucosa (23,25,32).
Thus, investigation into the conversion process from the gastric
mucosa inflammation to GC will further the understanding of the
underlying mechanisms of GC genesis.
NF-κB is a protein complex of transcription factors
observed in the κ-light chain of immunoglobins in B-cells and is
composed of five members: Rel (c-Rel), RelA (p65), RelB, NF-κB1
(p50 and its precursor p105) and NF-κB2 (p52 and its precursor
p100) (34,35). The classical or canonical NF-κB
signaling pathway is activated by tumor necrosis factor (TNF), IL-1
and Toll-like receptor ligands, including lipopolysaccharides. Upon
stimulation, IκBα is phosphory-lated and ubiquitinated, triggering
the release, translocation and transcription of RelA (P65)
(36). Previous studies have
demonstrated that the activation of the NF-κB signaling pathway is
involved in the initiation and progression of a spectrum of
different types of gastrointestinal cancers, including liver,
pancreatic, prostate and lung cancers (37-42).
In GC, chronic inflammation, including that induced by H.
pylori infection, greatly increases the risk of cancer via a
process that involves the production of protumorigenic cytokines,
including TNF, IL-1, IL-6, IL-17A and IL-23, promoting the
activation of the NF-κB and signal transducer and activator of
transcription 3 signaling pathways (43). NF-κB acts in epithelial and myeloid
cells, and mainly suppresses cell death and sustains cell survival
in epithelial cells, consequently aggravating disease progression
(34). MTDH, a classical
oncoprotein, was reported to be involved in the activation of the
NF-κB signaling pathway (23).
Upon the engagement of the upstream cytokines to receptors, MTDH
will be phos-phorylated by IκB kinase β, then interact with P65,
which finally promotes the translocation and transcriptional
activity of P65 (44,45). In the present study, the
significant role of miR-3664-5P in the suppression of the NF-κB
signaling pathway was highlighted by targeting MTDH and inhibiting
the proliferation and metastasis of GC, which was validated in
vitro and in vivo. Analysis of the association between
miR-3664-5P and the clinical data from patients with GC revealed
that miR-3664-5P was significantly associated with the clinical
features of GC tumor differentiation and tumor size, and positively
correlated with the outcome of patients with GC. It should be noted
that the present results do not agree with previously established
data, in which GC prognosis was determined by the invasion of
layers in the gastric mucosa (T classification) and lymph node
metastasis (46,47), these were also not significantly
associated with the expression of miR-3664-5P in the present
cohort. Considering that correlation and prognostic analyses can
provide supporting evidence to the conclusions in vitro and
in vivo, and the results are cohort- and cohort
capacity-dependent, it should be further investigated in a large
cohort to confirm the results of the present study. Notably, the
potential of miR-3664-5P to serve as a diagnosis marker in patient
serum was also uncovered and verified in present study.
In conclusion, the present study suggested that
during the tumorigenesis and progression of GC, miR-3664-5P
suppressed the proliferation and metastasis of GC by attenuating
the NF-κB signaling pathway through targeting MTDH. This highlights
the potential of miR-3664-5P of being a prognostic and diagnostic
indicator in GC.
Funding
The present study was supported in part by the
National Natural Science Foundation of China (grant no.
81700537).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LT conceived and designed the study. YJ, HY and JQ
acquired the data, and performed the experiments and statistical
analysis. YG and HL carried out the patient follow-up procedures.
SW and LC analyzed and interpreted the data. YJ and LT drafted and
edited the manuscript. All authors have given final approval of the
version to be published.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Nanjing Medical University Affiliated Changzhou
No. 2 People's Hospital (approval no. 2010-SR-077.A1; Jiangsu,
China) and all patients provided written informed consent. All
experiments were approved by Animal Ethics Committee of Nanjing
Medical University (Jiangsu, China).
Patient consent for publication
The present study obtained consent for publication
from all patients.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr Hao Han (The
First Affiliated Hospital of Nanjing Medical University, Jiangsu,
China) for providing language and technological support.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe:
estimates for 40 countries in 2012. Eur J Cancer. 49:1374–1403.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Peng Y, Zhang X, Ma Q, Yan R, Qin Y, Zhao
Y, Cheng Y, Yang M, Wang Q, Feng X, et al: MiRNA-194 activates the
Wnt/β-catenin signaling pathway in gastric cancer by targeting the
negative Wnt regulator, SUFU. Cancer Lett. 385:117–127. 2017.
View Article : Google Scholar
|
|
6
|
Luo H and Liang C: MicroRNA-148b inhibits
proliferation and the epithelial-mesenchymal transition and
increases radiosensi-tivity in non-small cell lung carcinomas by
regulating ROCK1. Exp Ther Med. 15:3609–3616. 2018.PubMed/NCBI
|
|
7
|
Gruszka R and Zakrzewska M: The oncogenic
relevance of miR-17-92 cluster and its paralogous miR-106b-25 and
miR-106a-363 clusters in brain tumors. Int J Mol Sci.
19:192018.
|
|
8
|
Inoue A, Mizushima T, Wu X, Okuzaki D,
Kambara N, Ishikawa S, Wang J, Qian Y, Hirose H, Yokoyama Y, et al:
A miR-29b Byproduct Sequence Exhibits Potent Tumor-Suppressive
Activities via Inhibition of NF-κB Signaling in KRAS-Mutant Colon
Cancer Cells. Mol Cancer Ther. 17:977–987. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murray-Stewart T, Sierra JC, Piazuelo MB,
Mera RM, Chaturvedi R, Bravo LE, Correa P, Schneider BG, Wilson KT
and Casero RA: Epigenetic silencing of miR-124 prevents spermine
oxidase regulation: Implications for Helicobacter pylori-induced
gastric cancer. Oncogene. 35:5480–5488. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao Y, Tan S, Tu Y, Zhang G, Liu Y, Li D,
Xu S, Le Z, Xiong J, Zou W, et al: MicroRNA-125a-5p inhibits
invasion and metastasis of gastric cancer cells by targeting BRMS1
expression. Oncol Lett. 15:5119–5130. 2018.PubMed/NCBI
|
|
12
|
Yu X, Ma C, Fu L, Dong J and Ying J:
MicroRNA-139 inhibits the proliferation, migration and invasion of
gastric cancer cells by directly targeting ρ-associated protein
kinase 1. Oncol Lett. 15:5977–5982. 2018.PubMed/NCBI
|
|
13
|
Hu L, Wu H, Wan X, Liu L, He Y, Zhu L, Liu
S, Yao H and Zhu Z: MicroRNA-585 suppresses tumor proliferation and
migration in gastric cancer by directly targeting MAPK1. Biochem
Biophys Res Commun. 499:52–58. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan C, Yu J, Liu Y, Kang W, Ma Z and Zhou
L: MiR-32 promotes gastric carcinoma tumorigenesis by targeting
Kruppel-like factor 4. Biochem Biophys Res Commun. 467:913–920.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Delaunoit T: Latest developments and
emerging treatment options in the management of stomach cancer.
Cancer Manag Res. 3:257–266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su ZZ, Kang DC, Chen Y, Pekarskaya O, Chao
W, Volsky DJ and Fisher PB: Identification and cloning of human
astrocyte genes displaying elevated expression after infection with
HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid
subtraction hybridization, RaSH. Oncogene. 21:3592–3602. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee SG, Kim K, Kegelman TP, Dash R, Das
SK, Choi JK, Emdad L, Howlett EL, Jeon HY, Su ZZ, et al: Oncogene
AEG-1 promotes glioma-induced neurodegeneration by increasing
glutamate excitotoxicity. Cancer Res. 71:6514–6523. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park SY, Choi M, Park D, Jeong M, Ahn KS,
Lee J, Fisher PB, Yun M and Lee SG: AEG-1 promotes mesenchymal
transition through the activation of Rho GTPases in human
glioblastoma cells. Oncol Rep. 36:2641–2646. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liang Y, Hu J, Li J, Liu Y, Yu J, Zhuang
X, Mu L, Kong X, Hong D, Yang Q, et al: Epigenetic Activation of
TWIST1 by MTDH Promotes Cancer Stem-like Cell Traits in Breast
Cancer. Cancer Res. 75:3672–3680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu G, Chong RA, Yang Q, Wei Y, Blanco MA,
Li F, Reiss M, Au JL, Haffty BG and Kang Y: MTDH activation by 8q22
genomic gain promotes chemoresistance and metastasis of
poor-prognosis breast cancer. Cancer Cell. 15:9–20. 2009.
View Article : Google Scholar :
|
|
21
|
Robertson CL, Srivastava J, Siddiq A,
Gredler R, Emdad L, Rajasekaran D, Akiel M, Shen XN, Guo C,
Giashuddin S, et al: Genetic deletion of AEG-1 prevents
hepatocarcinogenesis. Cancer Res. 74:6184–6193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Srivastava J, Siddiq A, Emdad L,
Santhekadur PK, Chen D, Gredler R, Shen XN, Robertson CL, Dumur CI,
Hylemon PB, et al: Astrocyte elevated gene-1 promotes
hepatocarcinogenesis: Novel insights from a mouse model.
Hepatology. 56:1782–1791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li G, Wang Z, Ye J, Zhang X, Wu H, Peng J,
Song W, Chen C, Cai S, He Y, et al: Uncontrolled inflammation
induced by AEG-1 promotes gastric cancer and poor prognosis. Cancer
Res. 74:5541–5552. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu H, Song X, Liu C, Xie L, Wei L and Sun
R: Knockdown of astrocyte elevated gene-1 inhibits proliferation
and enhancing chemo-sensitivity to cisplatin or doxorubicin in
neuroblastoma cells. J Exp Clin Cancer Res. 28:192009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dicken BJ, Bigam DL, Cass C, Mackey JR,
Joy AA and Hamilton SM: Gastric adenocarcinoma: Review and
considerations for future directions. Ann Surg. 241:27–39.
2005.
|
|
26
|
Ikoma N, Blum M, Estrella JS, Das P,
Hofstetter WL, Fournier KF, Mansfield P, Ajani JA and Badgwell BD:
Evaluation of the American Joint Committee on Cancer 8th edition
staging system for gastric cancer patients after preoperative
therapy. Gastric Cancer. 21:74–83. 2018. View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43(D1): D146–D152. 2015. View Article : Google Scholar :
|
|
30
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
eLife. 4:42015. View Article : Google Scholar
|
|
31
|
Hu Y, Yu K, Wang G, Zhang D, Shi C, Ding
Y, Hong D, Zhang D, He H, Sun L, et al: Lanatoside C inhibits cell
proliferation and induces apoptosis through attenuating
Wnt/β-catenin/c-Myc signaling pathway in human gastric cancer cell.
Biochem Pharmacol. 150:280–292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang F, Meng W, Wang B and Qiao L:
Helicobacter pylori-induced gastric inflammation and gastric
cancer. Cancer Lett. 345:196–202. 2014. View Article : Google Scholar
|
|
33
|
Van Cutsem E, Dicato M, Geva R, Arber N,
Bang Y, Benson A, Cervantes A, Diaz-Rubio E, Ducreux M,
Glynne-Jones R, et al: The diagnosis and management of gastric
cancer: expert discussion and recommendations from the 12th
ESMO/World Congress on Gastrointestinal Cancer, Barcelona, 2010.
Ann Oncol. 22(Suppl 5): v1–v9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He G and Karin M: NF-κB and STAT3 - key
players in liver inflammation and cancer. Cell Res. 21:159–168.
2011. View Article : Google Scholar
|
|
35
|
Ghosh S and Karin M: Missing pieces in the
NF-kappaB puzzle. Cell. 109(Suppl): S81–S96. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zandi E, Rothwarf DM, Delhase M, Hayakawa
M and Karin M: The IkappaB kinase complex (IKK) contains two kinase
subunits, IKKalpha and IKKbeta, necessary for IkappaB
phosphorylation and NF-kappaB activation. Cell. 91:243–252. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L, et al: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E and
Ben-Neriah Y: NF-kappaB functions as a tumour promoter in
inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
He G, Yu GY, Temkin V, Ogata H, Kuntzen C,
Sakurai T, Sieghart W, Peck-Radosavljevic M, Leffert HL and Karin
M: Hepatocyte IKKbeta/NF-kappaB inhibits tumor promotion and
progression by preventing oxidative stress-driven STAT3 activation.
Cancer Cell. 17:286–297. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jin R, Yi Y, Yull FE, Blackwell TS, Clark
PE, Koyama T, Smith JA Jr and Matusik RJ: NF-κB gene signature
predicts prostate cancer progression. Cancer Res. 74:2763–2772.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Todoric J, Antonucci L, Di Caro G, Li N,
Wu X, Lytle NK, Dhar D, Banerjee S, Fagman JB, Browne CD, et al:
Stress-Activated NRF2-MDM2 Cascade Controls Neoplastic Progression
in Pancreas. Cancer Cell. 32:824–839.e8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen W, Li Z, Bai L and Lin Y: NF-kappaB
in lung cancer, a carcinogenesis mediator and a prevention and
therapy target. Front Biosci. 16:1172–1185. 2011. View Article : Google Scholar :
|
|
43
|
Karin M: NF-kappaB as a critical link
between inflammation and cancer. Cold Spring Harb Perspect Biol.
1:a0001412009. View Article : Google Scholar
|
|
44
|
Krishnan RK, Nolte H, Sun T, Kaur H,
Sreenivasan K, Looso M, Offermanns S, Krüger M and Swiercz JM:
Quantitative analysis of the TNF-α-induced phosphoproteome reveals
AEG-1/MTDH/LYRIC as an IKKβ substrate. Nat Commun. 6:66582015.
View Article : Google Scholar
|
|
45
|
Emdad L, Sarkar D, Su ZZ, Randolph A,
Boukerche H, Valerie K and Fisher PB: Activation of the nuclear
factor kappaB pathway by astrocyte elevated gene-1: Implications
for tumor progression and metastasis. Cancer Res. 66:1509–1516.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jiao X, Lu HJ, Zhai MM, Tan ZJ, Zhi HN,
Liu XM, Liu CH and Zhang DP: Overexpression of kallikrein gene 10
is a biomarker for predicting poor prognosis in gastric cancer.
World J Gastroenterol. 19:9425–9431. 2013. View Article : Google Scholar
|
|
47
|
Deng JY and Liang H: Clinical significance
of lymph node metastasis in gastric cancer. World J Gastroenterol.
20:3967–3975. 2014. View Article : Google Scholar : PubMed/NCBI
|