Introduction
Cervical cancer (CC) is one of the most common
malignant gynecological diseases and ranks fourth among all the
types of cancer affecting women worldwide, with an estimated
527,600 new cases and 265,700 deaths annually. In particular, this
type of cancer is more prevalent in developing countries and has
the second highest incidence and third highest death rate among all
malignancies affecting women (1).
As a developing country with one-fifth of the world population,
China accounts for approximately 98,900 new cases and 30,500 deaths
annually due to CC (2). The
traditional treatments of CC are surgery, chemotherapy and
radiotherapy. The earlier the diagnosis and treatment, the better
the outcome (3). At present, the
screening method for CC is a pap smear combined with an HPV test,
which is invasive and costly, thus resulting in lower screening
coverage in China (4). As a
well-studied serum biomarker for CC, the levels of squamous cell
carcinoma-related antigen (SCCA) have only been found to be
elevated in 28-85% of cervical squamous cell carcinomas (5). The sensitivity is lower for early
squamous cell carcinoma or adenocarcinoma (6,7).
Thus, there is an urgent need for the identification of novel,
convenient and effective detection methods for CC, such as novel
serum biomarkers.
Bradykinin (BK), one of the kinin peptides, consists
of 9 amino acids (Arg-Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg) and
preferentially binds to kinin B2 receptor (BDKRB2 or B2R). Its
degradation product des-[Arg9]-BK, has a high affinity to B1
receptor (B1R) (8). It is
generally considered that BK is released from H-kininogen by the
action of plasma kallikrein (KLK) (9). BK has been implicated in a number of
pathophysiological processes, including vasodilation, the increase
of vascular permeability, the sensing of pain, the contraction of
smooth muscle and the mediation of inflammation (9). It has recently been indicated that
the kallikrein-kinin system plays an important role in the
occurrence and progression of cancer. Kinins exhibit the ability to
stimulate cell proliferation, migration and angiogenesis, which may
contribute to the biological behavior of tumors (8). It has also been suggested that tumor
cells may bear a BK autocrine/paracrine mechanism to amplify its
tumor-promoting effect. However, there is little valid evidence to
support this hypothesis (8).
Villanueva et al found that BK and its degradation
production des[Arg9]-BK levels were higher in the sera of patients
with breast cancer and lower in patients with bladder cancer than
in control sera (10). Later, van
Winden et al further confirmed that des[Arg9]-BK levels were
increased in patients with breast cancer and were decreased to
normal levels following surgery, supporting its potential use as a
diagnostic biomarker (11).
Furthermore, several researchers have suggested that BK promotes
tumor progression through various pathways by activating B2R. For
example, BK can promote angiogenesis by increasing vascular
permeability in the early phase via B2 receptor in endothelial
cells and by promoting the upregulation of vascular endothelial
growth factor (VEGF) via B2 receptor in stromal fibroblasts in the
late phase in a mouse model bearing sarcoma 180 cell-derived tumors
(12). Ikeda et al also
reached a similar conclusion in that BK may facilitate
tumor-associated angiogenesis and tumor growth by stimulating
stromal B2 signaling to upregulate VEGF production mainly in
fibroblasts in Walker 256 carcinoma cell tumor-bearing rats and in
a kallikrein-kinin system-deficient rat model (13). Yu et al found that BK
operated through the B2 receptor, Akt, and mammalian target of
rapamycin (mTOR), which in turn activated nuclear factor-κB (NF-κB)
and activator protein 1 (AP-1), activating VEGF expression and
contributing to angiogenesis in human prostate cancer cells (PC3
and DU145 cells) (14). BK has
also been found to promote the migration and invasion of
hepatocellular carcinoma cells through transient receptor potential
cation channel subfamily M member 7 (TRPM7) and matrix
metalloproteinase-2 (MMP2) (15),
increasing the secretion and expression of endothelin-1 through
kinin B2 receptors in melanoma cells (16), and promoting cell proliferation,
migration, invasion and tumor growth of gastric cancer through the
extracellular signal-regulated kinase (ERK) signaling pathway
(17). However, to date, at least
to the best of our knowledge, related studies on cervical cancer
are limited. It would be interesting to determine whether BK may be
used as a diagnostic biomarker in patients with CC and whether BK
plays a role in the progression of CC.
Thus, in this study, we aimed to examine the role of
BK in CC and to determine whether it plays a role in the
progression of CC and whether it may be used as an effective
biomarker. We found that the serum BK levels were significantly
higher in patients with CC than in those with cervical
intraepithelial neoplasia (CIN) and the control patients, and its
levels were markedly decreased following surgery. The B2R levels
were also higher in CC tissues and CIN tissues than in normal
cervical tissues. Subsequently, we identified that BK promoted the
angiogenesis of HUVEC cells and upregulated the expression of VEGF
in CC cell lines. The antagonist of B2R, HOE140, exerted adverse
effects, downregulating the VEGF level. The knockdown or
overexpression of BDKRB2 in CC cell lines further facilitated the
identification of its carcinogenic role in CC and its promoting
effects on angiogenesis. In summary, BK may be a useful diagnostic
biomarker for CC and promotes tumor progression by upregulating
VEGF expression through BDKRB2.
Materials and methods
Patient specimens
Serum samples and tissues from patients with CC and
CIN, and control patients were obtained from the Biobank of the
Cancer Biology Research Center of Wuhan Tongji Hospital from
January, 2017 to December, 2017. Among the samples, the serum
samples were from 130 patients with CC (including 65 cases with
pre- and post-surgery paired samples, another 65 cases with only
pre-surgery samples; the surgery refers to radical surgery for
cervical cancer), 35 patients with CIN (pre- and post-surgery
paired; the surgery refers to cervical conization surgery) and 35
patients with uterine fibroids (as controls). Formalin-fixed and
paraffin-embedded tissue samples were also obtained from 44
patients with CC, 12 patients with CIN and 8 patients with uterine
fibroids. RNAlater frozen tissues were obtained from 15 patients
with CC and 9 patients with uterine fibroids. All the samples were
collected and used following the approval of the Ethics Committee
of Tongji Hospital.
Cell lines and cell culture
The human cervical cancer cell lines, SiHa and HeLa,
were purchased from the American Type Culture Collection. Human
umbilical vein endothelial cells (HUVECs) was purchased from
Procell. The HPV16-positive immortalized human cervical
keratinocyte cell line, S12, was a gift from Professor Kenneth Raj
(Centre for Radiation, Chemical and Environmental Hazards, Health
Protection Agency, UK) and had been well applied in our laboratory
(18,19). All the cells were cultured at 37°C
in a humidified atmosphere containing 5% CO2. The SiHa
and HeLa cells grew in DMEM (Gibco) supplemented with 10% FBS
(Gibco). The S12 cells were cultured in a 1:3 mixture of DMEM and
Ham F-12 medium (Gibco) supplemented with 5% FBS (Gibco), 5
μg/ml insulin, 8.4 ng/ml cholera toxin, 24.3 μg/ml
adenine, 0.5 μg/ml hydrocortisone and 10 ng/ml epidermal
growth factor. These cell culture medium supplements were purchased
from Sigma-Aldrich. The HUVECs were maintained in endothelial cell
medium (ECM; Sciencell).
Establishment of cell lines with the
stable up- or downregulation of BDKRB2
The lentiviral-based shRNA targeting bdkrb2
gene was used to knockdown BDKRB2 in the SiHa and HeLa cells,
obtaining SiHa-sh-B2R and HeLa-sh-B2R. Lentiviral vector encoding
the bdkrb2 gene was transfected into the SiHa and HeLa cells
to construct BDKRB2-overexpressing cell lines, termed SiHa-oe-B2R
and HeLa-oe-B2R. An empty lentiviral vector was used as a control.
The transfection of the cells with lentivirus was conducted
according to the manufacturer's instructions. Briefly,
corresponding amount of lentivirus (the multiplicity of infection
for SiHa cells was 5 and for HeLa cells was 10) was added to a
6-well plate with a cell density of 30-40% and final concentration
of 5 μg/ml polybrene was added 10 min prior to the addition
of the lentivirus to promote transfection efficiency. Following
incubation at 37°C in a humidified atmosphere containing 5%
CO2 for 12 h, the medium in the well plate was changed
to fresh medium. Following a further incubation as with the
above-mentioned culture conditions for 72 h, the constructed cells
were selected by the supplementation of 2 μg/ml puromycin in
the medium. All the lentiviral vectors carried a luciferase tag for
the convenience in the following animal experiments. The
constructed cells were used for subsequent experiments within 3
months. All lentiviruses were obtained from Genechem.
Animal models
Twenty BALB/c nu mice (female, aged 4 weeks,
weighing 16-18 g) were purchased from HFK Bioscience Co. Ltd. and
housed in the Experimental Animal Center of Tongji Hospital. These
mice were maintained at a specific pathogen-free facility with a
constant humidity of 50% and a temperature of 26°C. Sterile feed
and water were changed twice a week. The animal experiments were
conducted according to the guidelines of Care and Use of Laboratory
Animals of Tongji Hospital and with the approval of the Ethics
Committee of Tongji Hospital, Tongji Medical College, Huazhong
University of Science and Technology. After a week of feeding
adaptation, these mice were randomly divided into 3 groups. The
SiHa-oe-B2R and SiHa-sh-B2R groups had 7 mice each and the SiHa-con
group had 6 mice. The established SiHa-oe-B2R, SiHa-sh-B2R and
SiHa-con cells were injected subcutaneously into the right flank of
each mouse. Tumor growth was monitored every 7 days and imaged
using the animal in vivo imaging system (PerkinElmer) every
2 weeks until death or tumor formation for 2 months. The humane
endpoint was that the tumor has a maximum diameter of 10 mm. Also,
the mice would be immediately sacrificed when the humane endpoint
was reached. Tumor volume was calculated as 1/2ab2,
where 'a' is the largest diameter and 'b' is the smallest diameter.
The in vivo imaging software 4.0 version from PerkinElmer
was used to quantitatively measure the fluorescence intensity of
tumors. The tumors were then removed and fixed in 4%
paraformaldehyde for sectioning and histological examination.
RNA isolation and RT-qPCR
Total RNA was extracted from the cervical tissues
and cells using a Total RNA kit (Omega Biotek) according to the
manufacturer's instructions and then reverse transcribed into cDNA
using M-MLV reverse transcriptase (Takara). Quantitative polymerase
chain reaction (qPCR) was performed using SYBR-Green Supermix
(Bio-Rad) in triplicate. The primer sequences were as follows:
Human BDKRB2 forward, 5'-CCGAAAGAAGTCTTGGGAGGT-3' and reverse,
5'-CTGGCGTTCCACGGAGATG-3'; human VEGF forward,
5'-AGGGCAGAATCATCACGAAGT-3' and reverse,
5'-AGGGTCTCGATTGGATGGCA-3'; and human GAPDH forward,
5'-GACAGTCAGCCGCATCTTCT-3' and reverse, 5'-TTAAAAGCAGCCCTGGTGAC-3'.
The cycling conditions were as follows: 3 min at 95°C for
polymerase activation, followed by 45 cycles of 95°C for 15 sec and
60°C for 30 sec.
Western blot analysis
The cells were harvested and lysed by
radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime)
supplemented with a protease inhibitor cocktail (Roche). The
protein concentration was measured using the Coomassie brilliant
blue G-250 (Biofroxx) staining method. Protein samples (40
μg each) were separated by 10% SDS-PAGE, transferred onto a
polyvinylidene difluoride membrane and immunoblotted with primary
antibodies. The primary antibodies used were as follows: Rabbit
monoclonal anti-human BDKRB2 (1:500; ab134118, Abcam), rabbit
polyclonal anti-human VEGF (1:1,000; 102643, Genetex), rabbit
polyclonal anti-human α-tubulin (1:2,000; ANT014, Antgene). The
primary antibodies were incubated overnight at 4°C. The membranes
were then washed with phosphate-buffered saline Tween-20 (PBST, pH
7.5) and incubated with horseradish peroxidase-conjugated goat
anti-rabbit IgG (1:2,000; ANT020, Antgene) for 1 h at 37°C.
Finally, the proteins were detected using an enhanced
chemiluminescence system (Pierce/Thermo Fisher Scientific).
Immunohistochemistry (IHC)
The tissues were paraffin-embedded and cut to
4-μm-thick sections for immunohistochemical analysis using
the Avidin-Biotin Complex (ABC) Vectastain kit (Zsgb-Bio) according
to the manufacturer's instructions. Antigen retrieval was performed
with sodium citrate at 95°C for 8 min. Rabbit polyclonal anti-human
BDKRB2 (1:100; ab188797, Abcam), rabbit polyclonal anti-human VEGF
(1:200; GTX102643, Genetex) and rabbit polyclonal anti-human CD31
(1:100; GB11063-1, Servicebio) were used as the primary antibodies.
The staining intensity was graded as negative, weak, moderate or
strong, and the corresponding score was 0, 1, 2 and 3,
respectively. The staining area (100%) were confirmed by two
independent pathologists and the corresponding score was as
follows: 0-25%, 1; 26-50%, 2; 51-75%, 3 and 76-100%, 4. The
intensity score and proportion score of each sample were multiplied
to obtain a staining index ranging from 0 to 12 points. The slides
were independently blind-reviewed by 2 experienced observers. The
final score was the average of the 2 scores.
Enzyme-linked immunosorbent assay
(ELISA)
The serum BK level was measured using a bradykinin
ELISA kit (ADI-900-206, ENZO LifeScience) following the
manufacturer's instructions. VEGF levels in culture supernatants
were detected using a Human VEGF ELISA kit (EHC108; NeoBioscience).
These culture supernatants were collected after the cells were
cultured for 72 h in 6-well plates. All samples were performed in
duplicate. The optical density (OD) is measured at a wavelength of
450 nm on a microtiter plate reader (Bio-Rad).
Tube formation assay
A total of 60 μl Matrigel (10 mg/ml, Corning
Inc.) was coated in the wells of a 96-well plate, and
104 HUVECs in 100 μl medium were then placed onto
the layer of Matrigel in each well. The concentration variable
(0.1, 1 and 10 μM) Bradykinin (Tocris Bioscience) and the
bradykinin B2 receptor antagonist, HOE140, (Tocris Bioscience) were
supplemented to the corresponding wells. Medium alone served as the
control. Tube morphogenesis was assessed under an Olympus IX73
microscope (Olympus) within 12 h. Each group had triplicate wells
and the total tube length was quantified using ImageJ 1.8.0
software.
Oncomine data
We searched for data on the bdkrb2 gene on
the Oncomine database and used the Cervical Cancer vs. Normal
Analysis datasets to produce a P-value with the Oncomine online
database (https://www.oncomine.org). We set the
threshold by a P-value of 0.05, fold change to all, and gene rank
to top 10%.
Statistical analysis
Statistical analyses were performed using IBM SPSS
statistics 22 and GraphPad Prism 5. Data are expressed as the means
± SEM. An unpaired t-test or paired t-test was used to determine
the statistically significant differences among 2 groups. If
variances were unequal, Welch's correction was used. When comparing
3 sets of data, one-way ANOVA was used with the Bonferroni post hoc
test to compare any 2 sets. For ranked data (IHC scores), the
Kruskal-Wallis test and a post hoc test (Dunns) were applied. To
evaluate the diagnostic efficiency of the serum biomarkers, the
area under the curve (AUC) of the receiver operating characteristic
(ROC) curve was determined with a 95% confidence interval. Binary
logistic regression was used to calculate the combination
forecasting efficiency of 2 markers. MedCalc Version 18.6 was used
to analyze the ROC curves. The Kaplan-Meier method with the
log-rank test and post hoc analysis (Bonferroni) were used for
survival curve analysis. Student's t-test was used for Table I. Differences were considered to be
statistically significant at P<0.05.
 | Table IAssociation between the
clinicopathological characteristics of the patients with cervical
cancer and the serum BK and SCCA expression levels. |
Table I
Association between the
clinicopathological characteristics of the patients with cervical
cancer and the serum BK and SCCA expression levels.
| Characteristic | No. of
patients | BK (ng/ml) | P-value | SCCA (ng/ml) | P-value |
|---|
| Age (years) | | | | | |
| ≥50 | 76 | 8.41±0.64 | 0.3469 | 2.24±0.31 | 0.0975 |
| <50 | 54 | 7.52±0.65 | | 1.56±0.27 | |
| FIGO stage | | | | | |
| IA - IB | 83 | 8.15±0.61 | 0.7567 | 1.68±0.23 | 0.1281 |
| IIA - IIIB | 47 | 7.85±0.69 | | 2.41±0.42 | |
| Lymph node
involvement | | | | | |
| Yes | 13 | 9.73±1.99 | 0.2845 | 3.28±0.69 | 0.0185 |
| No | 102 | 7.45±0.46 | | 1.31±0.13 | |
| Missed | 15 | | | | |
|
Differentiation | | | | | |
| Well-moderately
differentiated | 63 | 7.77±0.57 | 0.9889 | 1.94±0.28 | 0.8746 |
| Poorly
differentiated | 39 | 7.78±0.94 | | 2.00±0.33 | |
| Missed | 28 | | | | |
| Pathological
type | | | | | |
| Squamous
carcinoma | 102 | 7.76±0.48 | 0.2475 | 2.17±0.26 | 0.0025 |
|
Adenocarcinoma | 28 | 9.06±1.21 | | 1.08±0.24 | |
Results
Serum bradykinin is overexpressed in
patients with CC
Bradykinin expression was significantly higher in
patients with CC than in patients with CIN (P<0.05) and
non-cancerous control patients (P<0.01), as illustrated in
Fig. 1A. The medium concentration
in all cervical cancer cases prior to surgery was 8.04±0.46 ng/ml
(n=130). The corresponding expression level in CIN cases was
5.92±0.64 ng/ml (n=35) and in the control group was 4.93±0.59 ng/ml
(n=35) (data not shown). The clinicopathological characteristics of
the patients with CC and association analysis among age, FIGO
stage, lymph node metastasis, differentiation and pathological type
are demonstrated in Table I. The
missing data was due to not undergoing surgery in this hospital or
that the pathologists did not provide a clear judgment of the level
of tumor differentiation for some patients. However, none of these
factors was significantly associated with the higher serum BK
concentrations of patients with CC. We also obtained the serum SCCA
expression levels in the patients from the laboratory department. A
similar association analysis was conducted and we found that the
expression of SCCA in patients with lymph node metastasis was
significantly increased compared to that in patients who did not
have lymph node metastasis (P<0.05, Table I). Moreover, the expression of SCCA
in squamous carcinoma was markedly higher than that in patients
with adenocarcinoma (P<0.01, Table
I). These results are in accordance with the current research
(20). In addition, we compared
the serum BK level pre- and post-surgery, and a prominent decrease
was found in the patients with CC following surgery (P<0.0001,
Fig. 1B), whereas no statistically
significant difference was observed between the paired CIN cases
(P>0.05, Fig. 1B). We set up
the cut-off value at the maximum of Youden's index and the
corresponding sensitivity, specificity, positive predictive value
and negative predictive values are shown in Table II. SCCA held a high specificity
(96.97%), but a low sensitivity (32.52%). BK had a better
sensitivity (65.85%) but lower specificity (66.67%). To estimate
the diagnostic efficacy of BK and SCCA, as well as their
combination effect, ROC curves were depicted (Fig. 1C). The combination of the 2 led to
a better diagnostic effect. The AUC of BK was 0.671 (95% CI:
0.590-0.751) and that of SCCA 0.657 (95% CI: 0.579-0.735). When
both were combined, the AUC was 0.752 (95% CI: 0.682-0.821),
indicating a preferable diagnostic efficacy (P<0.01, compared
with any single indicator).
| Table IIDiagnostic values of serum BK and
SCCA at Youden's index maximum. |
Table II
Diagnostic values of serum BK and
SCCA at Youden's index maximum.
| Groups | Cut-off
(ng/ml) | Sensitivity
(%) | Specificity
(%) | Positive predictive
value (%) | Negative predictive
value (%) |
|---|
| BK | 5.15 | 65.85 | 66.67 | 78.64 | 51.16 |
| SCCA | 1.70 | 32.52 | 96.97 | 95.24 | 43.53 |
The receptor of bradykinin BDKRB2 is
overexpressed in cervical cancer tissues
We searched the Oncomine online database (https://www.oncomine.org) and found that BDKRB2
expression was significantly upregulated in the CC vs. the normal
samples in one dataset (Scotto Cervix Statistics) (Fig. 2A). We then further verified this
conclusion in CC tissues (n=15) and normal cervical tissues (n=9)
through RT-qPCR (P=0.0426, Fig.
2B). Apart from the mRNA level, we carefully detected the
BDKRB2 protein expression level by IHC. BDKRB2 was notably
overexpressed in the CC and CIN tissues, and typical images are
exhibited in Fig. 2C. The relevant
cartogram is illustrated in Fig.
2D.
Bradykinin promotes angiogenesis and
upregulates VEGF expression in CC cells
BK markedly promoted the angiogenesis of HUVECs and
the B2R antagonist, HOE140, inversely played an inhibitory role.
Representative pictures were collected 4h after cells implantation
and displayed in Fig. 3A. The
total tube length was quantified and is illustrated in Fig. 3B. We then detected BDKRB2
expression in the CC cell lines, SiHa and HeLa, as well as the CIN
cell line, S12. The expression level of BDKRB2 was higher in the
SiHa and HeLa cells than in the S12 cells (Fig. 3C). Following treatment with the B2R
agonist BK and the inhibitor, HOE140, by concentration gradients,
the VEGF expression level in the SiHa and HeLa cells was evaluated
by western blot analysis (Fig.
3D). Furthermore, following the addition of 10 μM BK or
HOE140 to the cell well plates, the VEGF in mRNA level and culture
supernatants (Fig. 3E and F) were
examined. We observed a common result in that BK promoted VEGF
expression and HOE140 downregulated VEGF expression. However, the
upregulation of VEGF expression in the cell culture supernatant
following treatment with BK was not statistically significant.
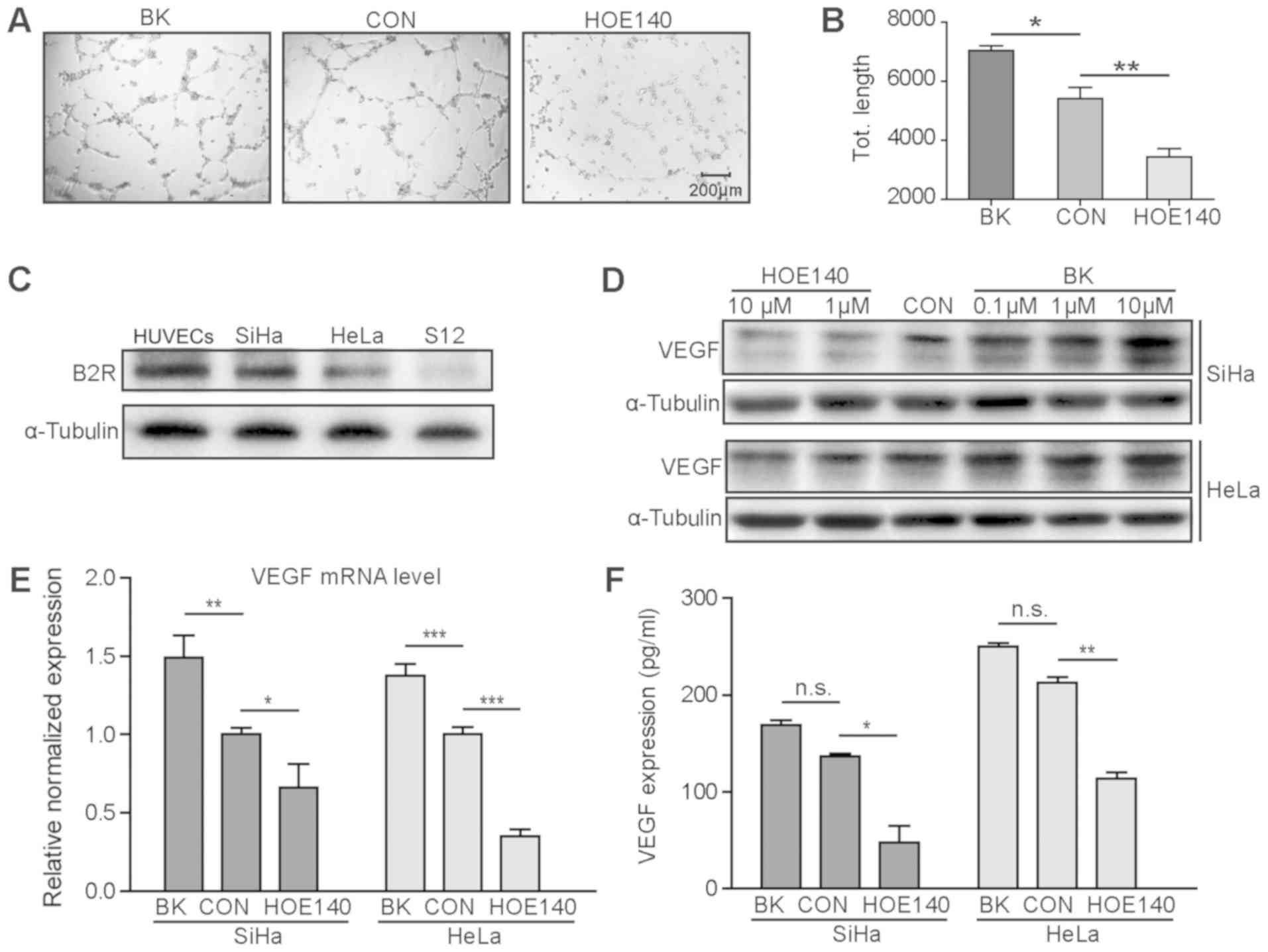 | Figure 3BK promotes angiogenesis and
upregulates VEGF expression in CC cells. (A) Representative images
of tube formation experiment of HUVECs cultured with BK or HOE140.
(B) The total tube length was quantified and statistics analysis
was conducted. BK promoted angiogenesis and HOE140 played an
inhibitory role. (C) The expression level of BDKRB2 was higher in
the CC cell lines, SiHa and HeLa, and lower in the CIN cell line,
S12. (D-F). BK upregulated the expression of VEGF in the SiHa and
HeLa cells at the protein level and mRNA level, and in culture
supernatants. HOE140 inversely suppressed VEGF expression.
*P<0.05, **P<0.01 and
***P<0.001, n.s. non-statistical significance. BK,
bradykinin; CC, cervical cancer; CON, control; VEGF, vascular
endothelial growth factor. |
Knockdown or overexpression of BDKRB2
affects the expression level of VEGF in CC cells
To further confirm whether B2R is involved in
BK-induced VEGF expression and angiogenesis, we constructed cells
(SiHa and HeLa cells) in which B2R was overexpressed or knocked
down. As shown in Fig. 4A and B,
the establishment of these cells was successful both at the protein
level and mRNA level. VEGF expression was then detected in these
cells at the protein level and mRNA level, and in culture
supernatants (Fig. 4A, C and D). A
total of 10 μM of BK was added to the supernatant of all
these cells during culture. The B2R-overexpressing CC cells
exhibited a markedly higher VEGF expression and the CC cells in
which B2R was knocked down exhibited a lower VEGF expression. Thus,
we came to the conclusion that BK upregulated VEGF expression and
promoted angiogenesis through B2R.
Effect of BDKRB2 on tumor growth in
vivo
We used the established SiHa cells in which BDKRB2
was overexpressed or knocked down to create a tumor-bearing mouse
model and observed tumor growth and the survival status. The
largest tumor diameter observed was 10 mm, which was in the
SiHa-oe-B2R group at 8 weeks following tumor implantation. The
total observation time was 9 weeks. In total, 17 mice died before 9
weeks. Specifically, 1 mouse died in the 5th week, 2 died in the
6th week, 3 died in the 7th week, 5 died in the 8th week, and 6
died in the 9th week. Among these, one of the deaths in the 8th
week was due to the fact that the mouse was sacrificed as the tumor
diameter was up to 10 mm; the other mice also had to be sacrificed
due to tumor growth. The SiHa-oe-B2R tumor-bearing mice exhibited a
significantly more rapid tumor growth and a shorter survival, while
the SiHa-sh-B2R tumor-bearing mice exhibited the opposite result
(Fig. 5A-C). The survival curve of
these tumor-bearing mice is depicted in Fig. 5A. The survival time of the
SiHa-sh-B2R tumor-bearing mice was significantly longer than that
of the SiHa-con group (P=0.024) and the SiHa-oe-B2R group
(P=0.003). The tumor size over time measured directly by tumor
diameter and indirectly by monitoring fluorescence intensity
produced by luciferase is depicted in Fig. 5B and C. Typical images were
displayed in Fig. 5D. The mice in
the SiHa-oe-B2R group exhibited a more rapid tumor growth and
tumors in the SiHa-oe-B2R group grew at a slower rate. We then
removed the tumor tissues and performed immunohistochemical
analysis (Fig. 5E). The BDKRB2
expression level was consistent with the established cells. Not
surprisingly, the SiHa-oe-B2R-derived tumors expressed higher
levels of VEGF, higher levels of the vascular marker, CD31, as well
as more neovascularization at the edge of the tumor (indicated by
black arrows). All the above suggested that BDKRB2 promoted tumor
growth in vivo and accelerated the formation of blood
vessels in tumors.
Discussion
SCCA is a well-studied biomarker of cervical
squamous cell cancer and it is used for the diagnosis, prognosis,
monitoring therapy and detecting the recurrence of CC. However,
only 10% of pure adenocarcinomas have elevated serum SCCA levels at
diagnosis (20). It was previously
shown that the positive predictive value for lymph node metastasis
at >8.6 ng/ml was 100% with a sensitivity of only 22.6%
(21). In this study, the
diagnostic efficacy (AUC) of SCCA was 0.657 and its sensitivity was
only 32.52%; thus, it is not an ideal diagnostic indicator when
used alone. The AUC of BK was 0.671, and when used in combination
with SCCA, the diagnostic efficacy increased to 0.751. In
particular, patients with adenocarcinoma had slightly higher levels
of BK than patients with squamous cell carcinoma, indicating a
favorable supplement in the diagnosis of adenocarcinoma. Thus, BK
used in combination with SCCA wound be a better choice for the
rough screening of cervical cancer, in the case that the cervical
smear is not available.
BK plays a role in the progression of various tumors
through multiple mechanisms (14-17).
We hypothesized that BK may also facilitate CC progression. We
detected the levels of its receptor B2R in CC and normal cervical
tissues and found that it was overexpressed both at the protein
level and mRNA level. The growth of human tumors and development of
metastases depend on the de novo formation of blood vessels
(22). The VEGF signaling pathway
has been implicated as the key endothelial cell-specific factor
signaling pathway required for tumor neovascularization (23,24).
In this study, we also conducted HUVEC tube formation assay and
found that BK promoted angiogenesis and the B2R antagonist, HOE140,
exerted an inhibitory effect. Consistent with this finding, the
VEGF expression levels in the BK-treated CC cell lines (SiHa and
HeLa) were significantly increased, while they were decreased in
the HOE140-treated cells. To further explore the mechanisms through
which BK promotes cervical cancer progression, we constructed CC
cell lines in which B2R was overexpressed or knocked down.
Following treatment with BK, the B2R-overexpressing cells
correspondingly exhibited a higher VEGF expression and the cells in
which B2R was knocked down exhibited a decreased VEGF expression. A
tumor-bearing mouse model with the established SiHa cells further
suggested that B2R-overexpressing tumors grew at a more rapid rate
and exhibited more neovascularization; in addition, mice with
B2R-overexpressing tumors exhibited a shorter survival time. These
data suggested that BK promoted VEGF expression by activating B2R
to accelerate the progression of CC. In a recent study, BK was
found to promote the progression of neuroblastoma through a variety
of mechanisms, including the upregulation of VEGF expression,
increasing metalloproteinase activity and inducing the adhesion of
neuroblastoma cells (25). Tumor
progression is a complex process with multiple factors and there
may be other mechanisms through which BK promotes the development
of CC. Thus, further investigations are required into this
matter.
Kinin receptors have been regarded as targets for
cancer therapy (26), and may also
provide an alternative for the treatment of CC. B2R is considered
to be widely expressed in human cancer and in experimental murine
tumors (27). Targeting B2R using
cell-penetrating antagonists has been shown to arrest the growth
and induce the apoptosis of human triple-negative breast cancer
(28). The BK antagonist dimer,
CU201, has been shown to inhibit the growth of small cell lung
cancer (SCLC) and non-small cell lung cancers (NSCLC) cell lines
but to have no effect on the growth of normal lung cells (29). When combined with antitumor agents
doxorubicin, etoposide, cisplatin, vinorelbine, paclitaxel or an
epidermal growth factor receptor tyrosine kinase inhibitor
(ZD1839), CU201 has been shown to exert synergistic growth
inhibitory effects (30). In this
study, the B2R antagonist, HOE140, inhibited HUVEC tube formation
and downregulated VEGF expression in CC cells, suggesting an
applicability in the treatment of CC. In addition, B1R is also
known as a more attractive target for the development of tumor
imaging probes or therapeutic opportunities due to its low
expression in normal tissues (26). Radionuclide-labeled B1R ligand
analogs successfully target B1R-positive tumors in vivo
(31-33). Cell-penetrant B1R antagonists have
been shown to exert an anticancer effect on MDA-MB-231 cells and
can cooperate with chemotherapeutic agents to promote
triple-negative breast cancer cell death (34). The role and application of B1R in
CC is also worth exploring in future research.
In the present study, an issue worthy of discussion
is to determine the reasons why the serum BK level was elevated in
patients with CC. The reasons for this remain undetermined. Tumor
cells may bear a BK autocrine/paracrine mechanism which can promote
the action of kinins, leading to signal amplification for tumor
growth (8). Kinins are generated
from kininogen by KLK (9). On the
one hand, the elevated BK level may be caused by an increase in its
precursor, kininogen. A recent study reported that BK precursors
(kininogens-1 and 2) were detected at much higher concentrations in
bone marrow cells of γ-irradiated mice when compared to
non-irradiated animals, which increased the BK release and
contributed to a favorable microenvironment for metastasis
formation in neuroblastoma (25).
On the other hand, it may be that the over-expression of KLKs leads
to the increase in BK levels. KLKs are overexpressed in a number of
types of cancer and may be potential biomarkers for diagnosis and
therapy (35). The broad use of
prostate-specific antigen (PSA/KLK3) in the routine clinical
management of patients with prostate cancer was a good illustration
(36). There was another notable
phenomenon that serum BK levels did not increase, while B2R
expression was significantly upregulated among patients with CIN.
It may be that the microenvironment has not promoted BK elevation,
but a high expression of B2R in HPV-integrated CIN cells has
emerged. No relevant research has yet been conducted on this
matter, at least to the best of our knowledge. The underlying
mechanisms warrant further investigation. BDKRB2 as an oncoprotein
may also function through many other pathways to promote the
progression of CC.
In conclusion, and to the best of our knowledge,
this study was the first to explore the association between BK/B2R
and CC. We found that the serum BK levels and the levels of its
receptor, B2R, were overexpressed in patients with CC. B2R
exhibited a high expression even in CIN tissues. The serum BK
levels in combination with SCCA may provide a more effective and
convenient diagnostic method. One potential mechanism through which
BK promotes the development of CC may be the upregulation of VEGF
expression, thus accelerating angiogenesis through B2R.
Funding
This study was financially supported by the Natural
Science Foundation Committee of China (grant nos. 81472444 and
81630060) and the National Science-technology Supporting Plan
Projects (grant no. 2015BAI13B05).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
YZ, LX and DM conceived and designed the study. YZ,
WW, RW and GJ performed the experiments. YZ, FL, XC, XW and SL
analyzed the data. YZ wrote the manuscript. LX planned and
supervised the study as well as revised the manuscript. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
This study was conducted under the approval of
Ethics Committee of Tongji Hospital (Wuhan, China). Since the
samples were from the Biobank of Cancer Biology Research Center of
Tongji hospital, the patient's informed consent was waived. Animal
experiments were conducted according to the guidelines of Care and
Use of Laboratory Animals of Tongji Hospital and with the approval
of the Ethics Committee of Tongji Hospital, Tongji Medical College,
Huazhong University of Science and Technology.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Koh WJ, Greer BE, Abu-Rustum NR, Apte SM,
Campos SM, Cho KR, Chu C, Cohn D, Crispens MA, Dorigo O, et al:
Cervical Cancer, Version 2.2015. J Natl Compr Canc Netw.
13:395–404; quiz 404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Di J, Rutherford S and Chu C: Review of
the cervical cancer burden and population-based cervical cancer
screening in China. Asian Pac J Cancer Prev. 16:7401–7407. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ueda Y, Enomoto T and Kimura T, Miyatake
T, Yoshino K, Fujita M and Kimura T: Serum biomarkers for early
detection of gynecologic cancers. Cancers (Basel). 2:1312–1327.
2010. View Article : Google Scholar
|
|
6
|
Pectasides D, Economides N, Bourazanis J,
Pozadzizou P, Gogou L, Koutsiouba P and Athanassiou A: Squamous
cell carcinoma antigen, tumor-associated trypsin inhibitor, and
carcinoembryonic antigen for monitoring cervical cancer. Am J Clin
Oncol. 17:307–312. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nam JH, Chang KC, Chambers JT, Schwartz PE
and Cole LA: Urinary gonadotropin fragment, a new tumor marker.
III. Use in cervical and vulvar cancers. Gynecol Oncol. 38:66–70.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
da Costa PL, Sirois P, Tannock IF and
Chammas R: The role of kinin receptors in cancer and therapeutic
opportunities. Cancer Lett. 345:27–38. 2014. View Article : Google Scholar
|
|
9
|
Bhoola KD, Figueroa CD and Worthy K:
Bioregulation of kinins: Kallikreins, kininogens, and kininases.
Pharmacol Rev. 44:1–80. 1992.PubMed/NCBI
|
|
10
|
Villanueva J, Shaffer DR, Philip J,
Chaparro CA, Erdjument-Bromage H, Olshen AB, Fleisher M, Lilja H,
BrogiE Boyd J, et al: Differential exoprotease activities confer
tumor-specific serum peptidome patterns. J Clin Invest.
116:271–284. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
van Winden AW, van den Broek I, Gast MC,
Engwegen JY, Sparidans RW, van Dulken EJ, Depla AC, Cats A,
Schellens JH, Peeters PH, et al: Serum degradome markers for the
detection of breast cancer. J Proteome Res. 9:3781–3788. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ishihara K, Kamata M, Hayashi I, Yamashina
S and Majima M: Roles of bradykinin in vascular permeability and
angiogenesis in solid tumor. Int Immunopharmacol. 2:499–509. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ikeda Y, Hayashi I, Kamoshita E, Yamazaki
A, Endo H, Ishihara K, Yamashina S, Tsutsumi Y, Matsubara H and
Majima M: Host stromal bradykinin B2 receptor signaling facilitates
tumor-associated angiogenesis and tumor growth. Cancer Res.
64:5178–5185. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu HS, Wang SW, Chang AC, Tai HC, Yeh HI,
Lin YM and Tang CH: Bradykinin promotes vascular endothelial growth
factor expression and increases angiogenesis in human prostate
cancer cells. Biochem Pharmacol. 87:243–253. 2014. View Article : Google Scholar
|
|
15
|
Chen Y, Yu Y, Sun S, Wang Z, Liu P, Liu S
and Jiang J: Bradykinin promotes migration and invasion of
hepatocellular carcinoma cells through TRPM7 and MMP2. Exp Cell
Res. 349:68–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Andoh T, Akira A, Saiki I and Kuraishi Y:
Bradykinin increases the secretion and expression of endothelin-1
through kinin B2 receptors in melanoma cells. Peptides. 31:238–241.
2010. View Article : Google Scholar
|
|
17
|
Wang G, Sun J, Liu G, Fu Y and Zhang X:
Bradykinin promotes cell proliferation, migration, invasion, and
tumor growth of gastric cancer through ERK signaling pathway. J
Cell Biochem. 118:4444–4453. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bechtold V, Beard P and Raj K: Human
papillomavirus type 16 E2 protein has no effect on transcription
from episomal viral DNA. J Virol. 77:2021–2028. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu Z, Ding W, Zhu D, Yu L, Jiang X, Wang
X, Zhang C, Wang L, Ji T, Liu D, et al: TALEN-mediated targeting of
HPV oncogenes ameliorates HPV-related cervical malignancy. J Clin
Invest. 125:425–436. 2015. View
Article : Google Scholar :
|
|
20
|
de Bruijn HW, Duk JM, van der Zee AG, Pras
E, Willemse PH, Boonstra H, Hollema H, Mourits MJ, de Vries EG and
Aalders JG: The clinical value of squamous cell carcinoma antigen
in cancer of the uterine cervix. Tumour Biol. 19:505–516. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bolger BS, Dabbas M, Lopes A and Monaghan
JM: Prognostic value of preoperative squamous cell carcinoma
antigen level in patients surgically treated for cervical
carcinoma. Gynecol Oncol. 65:309–313. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Folkman J: The role of angiogenesis in
tumor growth. Semin Cancer Biol. 3:65–71. 1992.PubMed/NCBI
|
|
23
|
McMahon G: VEGF receptor signaling in
tumor angiogenesis. Oncologist. 5(Suppl 1): 3–10. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Amini A, Masoumi Moghaddam S, Morris DL
and Pourgholami MH: The critical role of vascular endothelial
growth factor in tumor angiogenesis. Curr Cancer Drug Targets.
12:23–43. 2012. View Article : Google Scholar
|
|
25
|
Ulrich H, Ratajczak MZ, Schneider G,
Adinolfi E, Orioli E, Ferrazoli EG, Glaser T, Corrêa-Velloso J,
Martins PCM, Coutinho F, et al: Kinin and purine signaling
contributes to neuroblastoma metastasis. Front Pharmacol.
9:5002018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Figueroa CD, Ehrenfeld P and Bhoola KD:
Kinin receptors as targets for cancer therapy. Expert Opin Ther
Targets. 16:299–312. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu J, Akaike T, Hayashida K, Miyamoto Y,
Nakagawa T, Miyakawa K, Müller-Esterl W and Maeda H: Identification
of bradykinin receptors in clinical cancer specimens and murine
tumor tissues. Int J Cancer. 98:29–35. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dubuc C, Savard M, Bovenzi V, Lessard A,
Fortier A, Côté J, Neugebauer W, Rizzolio F, Geha S, Giordano A, et
al: Targeting intracellular B2 receptors using novel
cell-penetrating antagonists to arrest growth and induce apoptosis
in human triple-negative breast cancer. Oncotarget. 9:9885–9906.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chan D, Gera L, Stewart J, Helfrich B,
Verella-Garcia M, Johnson G, Baron A, Yang J, Puck T and Bunn P Jr:
Bradykinin antagonist dimer, CU201, inhibits the growth of human
lung cancer cell lines by a 'biased agonist' mechanism. Proc Natl
Acad Sci USA. 99:4608–4613. 2002. View Article : Google Scholar
|
|
30
|
Chan DC, Gera L, Stewart JM, Helfrich B,
Zhao TL, Feng WY, Chan KK, Covey JM and Bunn PA Jr: Bradykinin
antagonist dimer, CU201, inhibits the growth of human lung cancer
cell lines in vitro and in vivo and produces synergistic growth
inhibition in combination with other antitumor agents. Clin Cancer
Res. 8:1280–1287. 2002.PubMed/NCBI
|
|
31
|
Lin KS, Pan J, Amouroux G, Turashvili G,
Mesak F, Hundal-Jabal N, Pourghiasian M, Lau J, Jenni S, Aparicio
S, et al: In vivo radioimaging of bradykinin receptor B1, a widely
overex-pressed molecule in human cancer. Cancer Res. 75:387–393.
2015. View Article : Google Scholar
|
|
32
|
Kuo HT, Pan J, Lau J, Zhang C, Zeisler J,
Colpo N, Bénard F and Lin KS: Radiolabeled R954 derivatives for
imaging bradykinin B1 receptor expression with positron emission
tomography. Mol Pharm. 14:821–829. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Amouroux G, Zhang Z, Pan J, Jenni S, Zhang
C, Hundal-Jabal N, Colpo N, Zeisler J, Lin KS and Bénard F:
Synthesis and evaluation of a 68Ga-labeled bradykinin B1 receptor
agonist for imaging with positron emission tomography. Bioorg Med
Chem. 25:690–696. 2017. View Article : Google Scholar
|
|
34
|
Dubuc C, Savard M, Bovenzi V, Lessard A,
Côté J, Neugebauer W, Geha S, Chemtob S and Gobeil F Jr: Antitumor
activity of cell-penetrant kinin B1 receptor antagonists in human
triple-negative breast cancer cells. J Cell Physiol. 234:2851–2865.
2019. View Article : Google Scholar
|
|
35
|
Kontos CK, Mavridis K, Talieri M and
Scorilas A: Kallikrein-related peptidases (KLKs) in
gastrointestinal cancer: Mechanistic and clinical aspects. Thromb
Haemost. 110:450–457. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hong SK: Kallikreins as biomarkers for
prostate cancer. BioMed Res Int. 2014:5263412014. View Article : Google Scholar : PubMed/NCBI
|


















