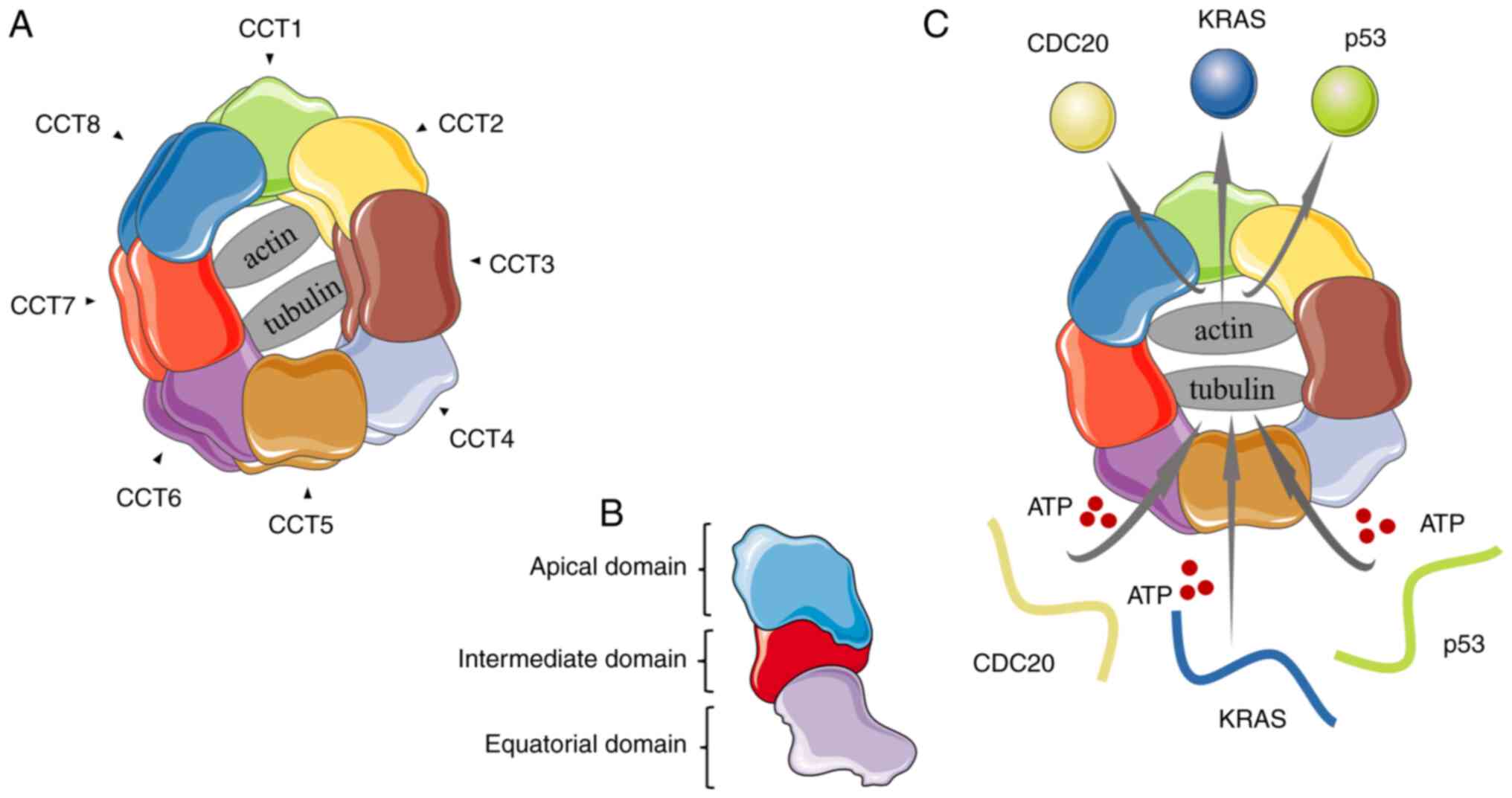
Chaperonins are functional proteins that help newly
synthesized polypeptide chains to fold and can be divided into two
categories. The first type is GroEL, which is present in fungi,
bacteria, mitochondria and chloroplasts. The second type is
Chaperonin Containing TCP-1 (CCT or TRiC), which exists in
eukaryotic cells (1,2). CCT has the most complex structure of
all chaperone proteins and is composed of two cyclic structures,
each of which consists of eight adjacent different subunits
(CCT1-8) with a molecular weight of ~60 kDa each (3,4).
CCT6 is also divided into two subunits, CCT6A and CCT6B (5). Each subunit can be divided into three
domains: Apical (substrate binding domain), intermediate
(connecting the other two domains) and the equatorial (contains the
ATP binding site). The sequences of the equatorial and intermediate
domains are conserved, while the sequences of the apical domains
are highly differentiated among the eight subunits (6,7).
There is a cavity in the middle of the CCT ring that can bind two
obligate folding substrates, actin and tubulin (7). The newly synthesized protein folds in
this cavity in an ATP-dependent manner (Fig. 1) (1,8). In
eukaryotic cells, CCT promotes the folding of ~10% of newly
synthesized proteins (1,4), including some cell cycle regulators
and tumor-associated proteins, such as cell division cycle protein
20, cadherin 1, STAT3, KRAS and p53 (9-11).
Malignant tumors are one of the diseases with the
highest mortality rates due to their concealment and high
invasiveness. Therefore, it is important to elucidate more tumor
markers. Previous studies have shown that upregulation of CCT can
promote the progression of malignant tumors (12-14).
Therefore, exploring genes that can inhibit CCT expression or
developing inhibitors against CCT may become the focus of future
research. For example, a study found that CCT6A is a target gene of
microRNA (miR)-148a/152 (15). The
upregulation of miR-148a/152 can inhibit the migration, invasion
and proliferation of CD44+/CD133+ colon
cancer stem cells and promote their apoptosis by inhibiting CCT6A
expression (15). A clinical study
has also demonstrated that treatment with the CCT inhibitor, CT20p,
can induce the death of neuroblastoma cells (16).
The purpose of the present review is to introduce
the structural characteristics of CCT and its role in the cell
cycle and to summarize its mechanism of action in the development
of certain cancer types. The present review also aims to provide a
new direction for the study of malignant tumors and to provide new
targets for early screening and treatment of tumors in the
future.
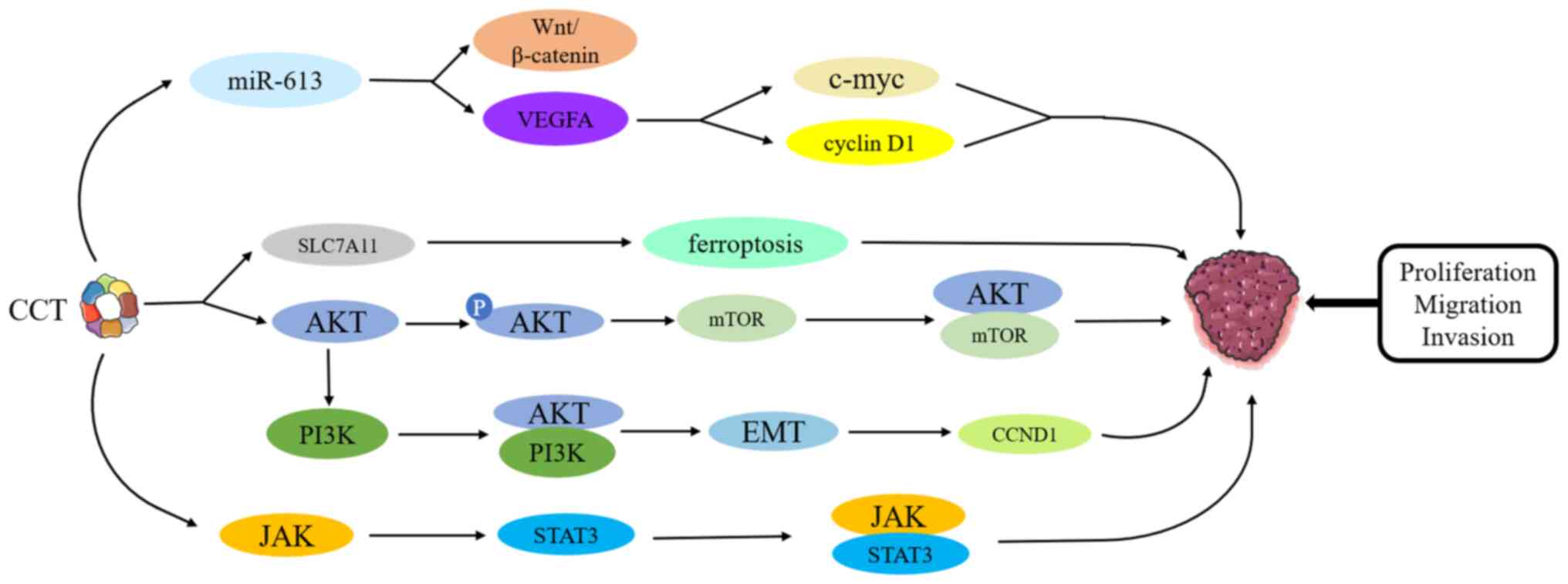
Studies have found that knockout of the CCT gene can
inhibit the phosphorylation and activation of AKT and that CCT can
interact with AKT and mTOR, activating AKT/mTOR signaling and
negatively regulating apoptosis and autophagy (12,24).
In particular, CCT3 has been shown to enhance AKT activity in lung
adenocarcinoma (LUAD) cells (25).
In addition, the AKT inhibitor, MK2206, significantly inhibits the
growth of CCT3-overexpressing lung cancer cells (25). In addition, a study has shown that
PI3K/AKT signaling pathway-related genes, CCNE1-2, GNB1-5 and TP53,
are significantly associated with changes in CCT expression,
suggesting that CCT may be used as a therapeutic target for
patients with malignant tumors (26). Meng et al (27) demonstrated that, knocking down CCT5
can inhibit LUAD cell migration and invasion in vitro by
inactivating PI3K/AKT and its downstream epithelial-mesenchymal
transition (EMT) signals, which could abrogate the accelerated
migration and invasion caused by Cyclin D1 overexpression. Guest
et al (28) used SUM-52
breast cancer cells to demonstrate that fibroblast growth factor
receptor 2 signaling regulates CCT expression through PI3K/AKT and
that this signaling does not require mTOR activity. In addition,
Wang et al (25) found that
CCT3 promotes LUAD cell growth by inhibiting SLC7A11-mediated iron
apoptosis and activating the AKT pathway. In summary, CCT can
regulate the proliferation, invasion and migration of cancer cells
through the PI3K/AKT/mTOR pathway.
STAT3 is an important oncogenic protein, which can
promote the proliferation and survival of cancer cells and control
cell cycle progression and apoptosis (29). It has been shown that STAT3 binds
to CCT in an ATP-dependent manner through its β-chain-rich DNA
binding domain and is a non-specific substrate of CCT (30). The addition of a second
TRiC-binding domain (TBD) to the N-terminus of STAT3 (the TBD of
pVHL, vTBD) can further increase the affinity of STAT3 for CCT
(30,31). STAT3 is present in primary small
cell lung cancer (SCLC) tissues and cell lines and is an important
target for inhibiting SCLC (32).
Carr et al (33) found that
the CCT inhibitor, CT20p (a polypeptide with cancer-specific
cytotoxicity), reduced STAT3 levels in SCLC cell lines and that
these cell lines were resistant to cell death induced by STAT3
inhibitors. It has been confirmed that the expression of CCT3 is
related to the JAK/STAT3 signaling pathway (34). Danni et al (35) found that CCT3 can promote the
cisplatin resistance of LUAD cells by activating the JAK2/STAT3
pathway and that CCT3 gene knockout can inhibit the JAK2/STAT3
pathway to re-sensitize A549/DDP cells to cisplatin. This suggests
that CCT3 may be a new molecular target for overcoming cisplatin
resistance in patients with LUAD. A study (36) showed that CCT3 may play a key role
in the transport of phosphorylated (p-)STAT3 and STAT3 from the
cytoplasm to the nucleus. CCT3 can enter the nucleus of hepatoma
cells with pSTAT3 and the expression of CCT3 is positively
correlated with the activation of the STAT3 signaling pathway
(36). Since CCT is essential for
the synthesis, refolding and activity of STAT3, it has become a
potential target for the treatment of cancerous diseases.
The Wnt/β-catenin signaling pathway serves an
important role in embryonic development and the homeostasis of cell
proliferation and differentiation in adult tissues. Excessive
activation of the Wnt signaling pathway has been found in a number
of cancer types, including colorectal cancer (CRC), breast cancer,
lung cancer and hematological malignancies (37). In addition, it has been shown that
CCT3 target genes are involved in the Wnt signaling pathway
(34). Li et al (38) found that circular CCT3 RNA directly
interacts with miR-613 and regulates the expression of VEGFA and
Wnt3 genes. CCT3 deficiency can inhibit the invasion and induce the
apoptosis of CRC cells through miR-613/Wnt3 or VEGFA (38). In addition, a study (39) confirmed that, when CCT3 expression
is knocked out in breast cancer cells, the expression of
Wnt/β-catenin and the downstream target genes, cyclin D1 and c-myc,
is inhibited. In addition, when CCT3 expression is upregulated,
these genes are also activated accordingly (39). These studies suggest that CCT3 may
promote the occurrence of breast cancer by activating the
Wnt/β-catenin signaling pathway. Tang et al (40) demonstrated that p53 can bind to the
second position of the Wnt7b promoter and that CCT interacts with
p53 to regulate the Wnt7b/β-catenin pathway. CCT gene knockout can
prevent the activation of the Wnt/β-catenin signaling pathway
through Wnt7b, thereby inhibiting the proliferation and migration
of hepatocellular carcinoma (HCC) cells (40). Therefore, further exploring the
relationship between CCT and the Wnt/β-catenin pathway may be a
viable direction for studying the mechanism of malignant tumor
development (Fig. 3).
p53 is a well-known tumor suppressor protein encoded
by the human TP53 (or TRp53) gene and is a major regulator of
genomic stability, the cell cycle, DNA repair, senescence and
apoptosis (41,42). The point mutation of p53 not only
leads to the loss of wild-type activity, but also leads to the
occurrence of cancer and the ability of p53 to promote invasion and
metastasis (43). A number of
studies (6,43,44)
have found that the N-terminus of p53 [including its subtype
Δ133p53 (45)] binds to seven
subunits of CCT to form a CCT-p53 complex to promote p53 folding.
When CCT is depleted or the binding is absent, the conformation of
p53 is altered, resulting in the accumulation of misfolded p53 and
thereby promoting the occurrence and development of tumors
(6,43,44).
It was found that there was a higher concentration of the more
acidic CCT3 variant protein in p53-deficient cells, indicating that
CCT3 is a protein rich in potential phosphorylation (46). Ooe et al (47) demonstrated that CCT5 was
upregulated in breast cancer carrying a p53 mutation, which may be
related to docetaxel resistance. Since p53 is one of the most
common tumor suppressor genes, it is necessary to further explore
the interaction between p53 and CCT.
Lung cancer is one of the most common malignant
tumors and the leading cause of cancer-related mortality (48,49).
There is evidence that risk factors for lung cancer often include
smoking, age, sex, family history, living environment and
occupational exposure (50). It
has been demonstrated that 85% of lung cancer cases are related to
smoking (51). Histologically,
lung cancer can be divided into SCLC and non-small cell lung cancer
(NSCLC), accounting for 15 and 85% of all lung cancers,
respectively (52). NSCLC can be
further divided into LUAD, squamous cell carcinoma, adenosquamous
cell carcinoma, large cell carcinoma and sarcomatoid carcinoma
(52-55). Current treatments for lung cancer
include surgical resection, targeted therapy, immunotherapy,
chemotherapy, radiotherapy and adjuvant therapy. Among them,
molecular targeting drugs are the main treatment for lung cancer.
However, the efficacy of these drugs is often limited and
treatment-related toxicity reduces their therapeutic effects.
Therefore, there is an urgent need to find new treatment strategies
for patients with lung cancer (56-58).
A large number of studies have shown that the
subunits of CCT interact with numerous proteins to affect the
development of lung cancer cells (59-62).
For example, Shi et al (59) found that CCT3 silencing inhibited
Yes-associated protein 1 (YAP1) and reduced the expression of YAP1
target genes in NSCLC cells. Activation of YAP1 by overexpression
of constitutively active YAP1 mutants can reverse the antitumor
effect of CCT3 inhibition on NSCLC cells (59). In addition, I-Trp can trigger
apoptosis in the highly metastatic CL1-5 NSCLC cell line by
interfering with the β-tubulin/CCT2 complex, then activating
endoplasmic reticulum stress and finally activating caspases
(60). In addition, at the
EC20 of I-Trp, the migration and invasion of cancer
cells were significantly inhibited (60). This strategy can be used to combat
highly metastatic LUAD with upregulated CCT2. Ying et al
(61) identified CCT6A as an
inhibitor and direct binding protein of SMAD2 and found that CCT6A
promoted metastasis in NSCLC cells. In addition, selective
inhibition of SMAD3 or CCT6A can effectively inhibit TGF-β-mediated
metastasis (61). This finding may
help to develop therapeutic strategies for targeting TGF-β in the
treatment of NSCLC. A study (62)
confirmed that CCT6A may promote cell proliferation by promoting
the transformation of NSCLC cells from G1 phase to S
phase, thereby accelerating tumor growth and determining TNM
staging in patients with advanced NSCLC (62). In addition, a study found that
CCT6A and CHCHD2 are co-amplified with EGFR in the chromosome 7p11
region and that both are potential driver genes for LUAD (63). The upregulation of CCT6A or CHCHD2
is related to the progression of LUAD (63). In addition, by inhibiting the
expression of CCT6A or CHCHD2, the acquired resistance to EGFR
tyrosine kinase inhibitors mediated by additional EGFR
amplification can be reduced (63).
Breast cancer is the most commonly diagnosed cancer
in women worldwide, with nearly 1.7 million confirmed new cases
each year, accounting for 25% of all new cancer cases in women (and
only 1% in men) (64,65). The number of women diagnosed with
breast cancer worldwide is increasing (66). Although the treatment of patients
with breast cancer has been greatly improved in recent years, the
prognosis of triple negative breast cancer (TNBC) is still
relatively poor (67). The lack of
drug targets is an obstacle to the treatment of patients with TNBC
(68). Since breast cancer is a
heterogeneous disease, treatment options and prognosis can vary
depending on the hormone receptor status and genetic
characteristics of the patient (69). Therefore, it is important to find
new tumor markers and targets.
The expression of CCT serves an important role in
the occurrence and development of breast cancer. Among the CCT
subunits, CCT1, CCT3, CCT4, CCT5 and CCT7 had the highest
expression levels in TNBC tissues (70). Furthermore, CCT6A and CCT8 were
abundant in HER2+ breast cancer tissues and CCT2 was
abundant in luminal breast cancer tissues (70). In different histological subtypes
of breast cancer, the high expression of CCT6A was negatively
correlated with estrogen receptor and progesterone receptor status
and positively correlated with nodular, basal and TNBC status
(71). CCT expression often
activates cell proliferation and abnormal cell proliferation is an
indicator of malignant tumor cells (72). Macario et al (72) showed that the proliferation index
and generation time of cells transfected with CCT2 lentivirus were
increased compared with the control cells. Evaluation of cell cycle
kinase (such as CDK2 and CDK4) levels found that they were
dependent on CCT. Therefore, increased cell proliferation
associated with CCT2 upregulation may be related to the kinases
that mediate the cell cycle (72).
Showalter et al (1)
observed the division of breast cancer cells and found that the
expression of a single CCT2 subunit may induce cell proliferation
through the activity of the chaperone protein complex, which
increases the levels of key cyclins, such as CDKs. Previous studies
have shown that CCT2 is a putative autocrine/paracrine factor and
has potential functions in regulating the growth of breast cancer
cells (73). Ghozlan et al
(74) demonstrated that CCT2 is
located on chromosome 12q15 and that genetic changes occur in
invasive breast cancer and other 12q15 oncogenes. Bassiouni et
al (75) demonstrated that
CT20p mediates its cytotoxicity by inhibiting CCT in breast cancer
cells. CT20p can decrease the levels of CCT target proteins, such
as actin, tubulin or STAT3, which are necessary for the
maintenance, adhesion and survival of cancer cells, leading to cell
death (75). For breast cancer
cases with high CCT expression, CCT inhibitors may be an effective
method for future treatment.
The liver is involved in the clearance of human
toxins, the uptake of nutrients and the regulation of blood volume
and can regulate metabolism and maintain homeostasis (76-78).
However, the development of liver tumors often leads to metabolic
disorders (76). Furthermore,
liver cancer (HCC accounts for 75-85%, cholangiocarcinoma accounts
for 10-15%) has become the fourth leading cause of cancer-related
mortality worldwide (76,79,80).
There are ~840,000 new cases of liver cancer and at least 78,000
individuals succumb to liver cancer each year (81). The global annual incidence rate is
~10.1/100,000 and the morbidity and mortality rates are steadily
rising (82-84). Due to the insidious nature of the
early stage of liver cancer and the limited accuracy of related
markers, treatment is difficult and the prognosis is extremely poor
(76,85,86).
The median overall survival time of untreated HCC at all stages is
estimated to be 9 months (84) and
the overall 5-year survival rate is ~20% (34% for local metastasis
and 3% for distant metastasis) (86). At present, early liver cancer is
mostly treated by surgical resection, but it is prone to
complications (87). For advanced
liver cancer, several tyrosine kinase inhibitors (such as sorafenib
and lenvatinib) and a monoclonal antibody against VEGFR2
(ranibizumab) are used for treatment, but long-term use can lead to
drug resistance (87,88). Therefore, it is important to find
new liver cancer markers. Tumor biomarkers are of great
significance in early screening, diagnosis, treatment evaluation,
recurrence and prognosis evaluation of cancer and CCT may provide a
new way for early detection and treatment of liver cancer.
The expression of CCT in liver cancer cells is
closely related to the diagnosis, treatment and prognosis of liver
cancer (26,36,89,90).
Investigation of selected genes in patients with HCC and validation
using The Cancer Genome Atlas database demonstrated that CCT3 was
significantly correlated with the overall survival time of patients
with HCC (36,90). In addition, the overall survival
rate of patients with low expression of cancer cell nuclear CCT3
was higher compared with patients with high CCT3 expression. A
study has found that the incidence of CCT gene changes in HCC is as
high as 51.39%, which is related to the poor prognosis of patients
with HCC (26). When HCC is in the
early stage and α-fetoprotein (AFP) is negative, CCT3 can be
combined with IQ motif containing GTPase activating protein 3 as a
complementary marker of AFP and its expression is independent of
AFP (89). A study by Cui et
al (36) demonstrated that
CCT3 accumulates in the cytoplasm and nucleus in HCC cells, while
in non-HCC cells, CCT3 mainly accumulates in the cytoplasm. In
addition, CCT3 is highly expressed in the nuclei of moderately and
poorly differentiated HCC, but expression is lower in the nuclei of
highly differentiated and non-HCC (36). Li et al (5) found that abnormal expression of the
CCT gene was associated with macrophage infiltration and patients
with HCC with high CCT gene expression and a high macrophage score
had a poor prognosis. This may be due macrophages mainly existing
in the immune microenvironment of liver cancer tumors as M2 type
(one of the polarization types of macrophages), induced by
interleukin-4, is mainly involved in the anti-inflammatory response
and tissue repair (91). Huang
et al (3) demonstrated
that, compared with LO2 cells, the expression level of CCT8 in HCC
cells was significantly increased and the expression of CCT8 was
the most abundant in Huh7 cells. In addition, it was also found
that silencing CCT8 inhibited the proliferation of HCC cells by
inhibiting the S phase entry of HCC cells (3). Wong et al (92) found that the CCT3 gene expression
level in tumors was higher than that in adjacent non-malignant
liver tissues in 10 cases of HCC with amplicon 1q21-q22. In
addition, Liu et al (93)
demonstrated that nearly 50% of patients with HCC had CCT gene
mutations, of which 27% had CCT3 gene mutations and 18% had CCT5
gene mutations. Furthermore, patients with CCT gene mutations had a
worse clinical prognosis compared with patients without
mutations.
CRC is the third most common cancer worldwide and
the second leading cause of cancer-related mortality (101,102). CRC is a multifactorial disease
involving genetic, environmental and lifestyle risk factors
(103). The high heritability and
frequent recurrence of CRC leads to considerable mortality and the
association with poor prognosis (104). At 2020, there were >1.9
million new cases of CRC (including anal) and nearly 935,000
mortalities in worldwide (105).
CRC accounts for 9.2% of global deaths, with 5- and 10-year
survival rates of 65 and 58%, respectively (106). The incidence and mortality rates
are 25% higher in men than in women (106). Although the treatment strategy of
CRC has been greatly improved, including colon resection,
chemotherapy and immunotherapy, the high frequency of recurrence
and metastasis makes CRC a serious threat to human health (107,108). The 5-year survival rate of
patients diagnosed with advanced CRC, especially those with distant
metastasis, remains low, at ~13% (109,110). Therefore, it is necessary to find
new biomarkers for early diagnosis, accurate prediction of
metastasis and prognosis.
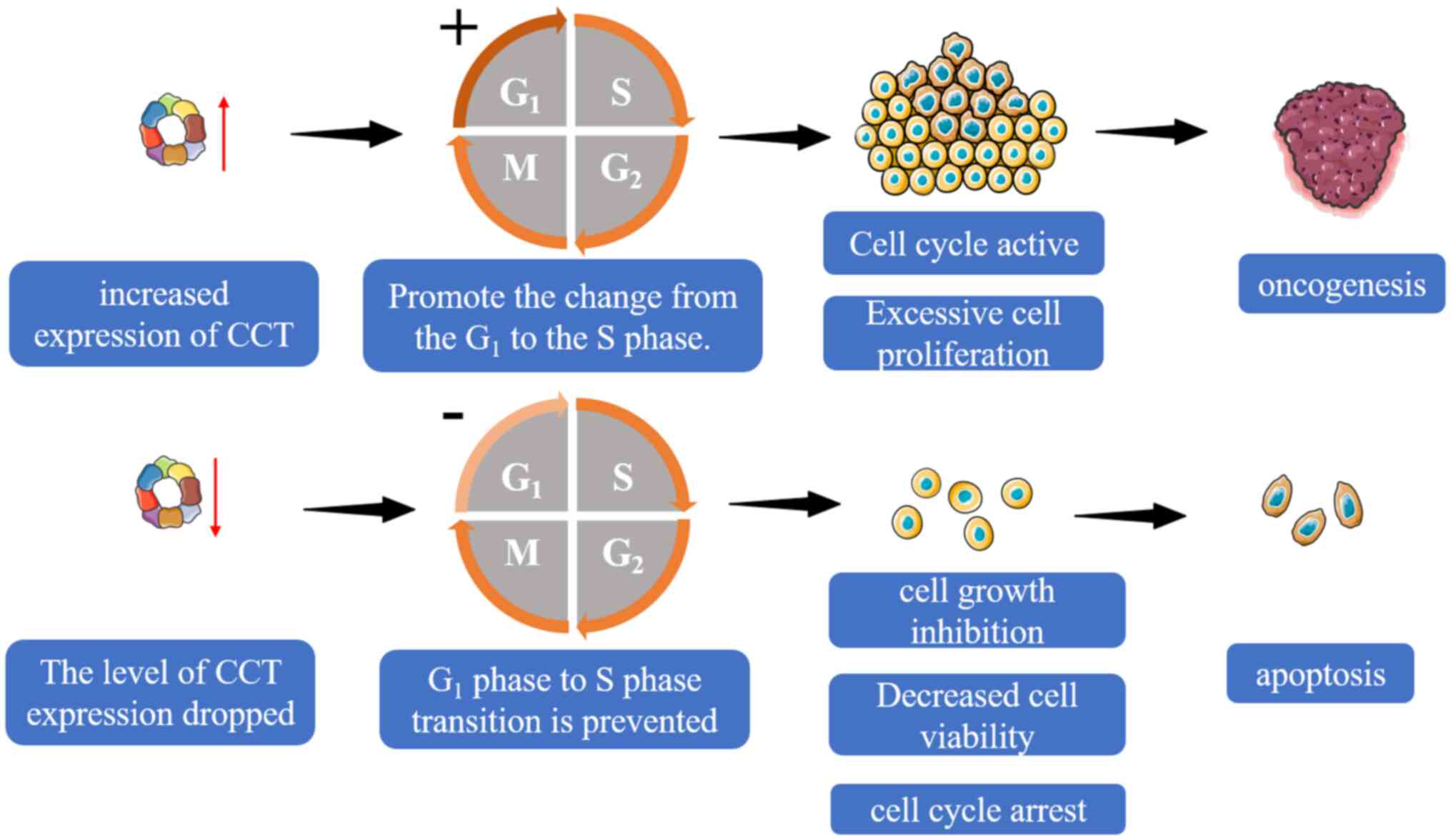
Cancer cells frequently have an unbalanced cell
cycle. Accumulating data indicates a crucial role for CCT in the
control of cell cycle progression. The development, proliferation
and death of eukaryotic cells are all influenced by CCT, which is
crucial for the folding of newly generated proteins. The expression
of CCT is significantly increased during cell division, which aids
in the folding of actin and other tumor-associated proteins
necessary for cell division. Finding new biomarkers is crucial to
advancing early detection and treatment of cancer, which is
especially critical given the rising incidence of the disease. CCT
is significant in a number of illnesses. Maintaining living cells
requires both the folding of newly generated proteins and the
preservation of proteins. Proteolytic turnover, intracellular
localization and protein folding are all closely regulated by the
molecular chaperone network. Therefore, CCT is a potential
diagnostic for malignant tumors since numerous important regulators
of cell growth and differentiation have been identified as target
proteins for molecular chaperones.
Data sharing is not applicable to this article, as
no data sets were generated or analyzed during the current
study.
LZheng and XC drafted the manuscript and
contributed equally. LZhang and NQ participated in the literature
search and analysis of the data to be included in the review. JA
and JZ were involved in the design of the study and assisted in the
preparation of the figures and tables. HJ and BT edited and revised
the manuscript. All authors have read and approved the final
version of the manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81960507, 82073087
and 82160112) and the Collaborative Innovation Center of Chinese
Ministry of Education (2020-39), the Science and Technology Bureau
fund of Zunyi City [grant no. ZUN SHI KE HE HZ ZI (2019)93-Hao] and
the Science and Technology Plan Project of Guizhou Province [grant
nos. QIAN KE HE JI CHU-ZK (2021) YI BAN451 and QIAN KE HE LH ZI
(2017)7095 HAO].
|
1
|
Showalter AE, Martini AC, Nierenberg D,
Hosang K, Fahmi NA, Gopalan P, Khaled AS, Zhang W and Khaled AR:
Investigating Chaperonin-Containing TCP-1 subunit 2 as an essential
component of the chaperonin complex for tumorigenesis. Sci Rep.
10:7982020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valpuesta JM, Martin-Benito J,
Gomez-Puertas P, Carrascosa JL and Willison KR: Structure and
function of a protein folding machine: The eukaryotic cytosolic
chaperonin CCT. FEBS Lett. 529:11–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang X, Wang X, Cheng C, Cai J, He S,
Wang H, Liu F, Zhu C, Ding Z, Huang X, et al: Chaperonin containing
TCP1, subunit 8 (CCT8) is upregulated in hepatocellular carcinoma
and promotes HCC proliferation. APMIS. 122:1070–1079.
2014.PubMed/NCBI
|
|
4
|
Tracy CM, Gray AJ, Cuellar J, Shaw TS,
Howlett AC, Taylor RM, Prince JT, Ahn NG, Valpuesta JM and
Willardson BM: Programmed cell death protein 5 interacts with the
cytosolic chaperonin containing tailless complex polypeptide 1
(CCT) to regulate beta-tubulin folding. J Biol Chem. 289:4490–4502.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li W, Liu J and Zhao H: Prognostic power
of a chaperonin containing TCP-1 subunit genes panel for
hepatocellular carcinoma. Front Genet. 12:6688712021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roh SH, Kasembeli M, Bakthavatsalam D,
Chiu W and Tweardy DJ: Contribution of the type II chaperonin,
TRiC/CCT, to oncogenesis. Int J Mol Sci. 16:26706–26720. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brackley KI and Grantham J: Activities of
the chaperonin containing TCP-1 (CCT): Implications for cell cycle
progression and cytoskeletal organisation. Cell Stress Chaperones.
14:23–31. 2009. View Article : Google Scholar :
|
|
8
|
Yam AY, Xia Y, Lin HT, Burlingame A,
Gerstein M and Frydman J: Defining the TRiC/CCT interactome links
chaperonin function to stabilization of newly made proteins with
complex topologies. Nat Struct Mol Biol. 15:1255–1262. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Liu P, Zhang Z, Wang J, Cheng Z
and Fan C: Identification of CCT3 as a prognostic factor and
correlates with cell survival and invasion of head and neck
squamous cell carcinoma. Biosci Rep. 41:BSR202111372021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu G, Bu S, Wang X, Zhang H and Ge H:
Suppression of CCT3 inhibits the proliferation and migration in
breast cancer cells. Cancer Cell Int. 20:2182020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lopez T, Dalton K and Frydman J: The
mechanism and function of group II chaperonins. J Mol Biol.
427:2919–2930. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Weng H, Feng X, Lan Y and Zheng Z: TCP1
regulates PI3K/AKT/mTOR signaling pathway to promote proliferation
of ovarian cancer cells. J Ovarian Res. 14:822021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dou L and Zhang X: Upregulation of CCT3
promotes cervical cancer progression through FN1. Mol Med Rep.
24:8562021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Liu C, Zhang X, Huang X, Liang S,
Xing F and Tian H: CCT5 induces epithelial-mesenchymal transition
to promote gastric cancer lymph node metastasis by activating the
Wnt/β-catenin signalling pathway. Br J Cancer. 126:1684–1694. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng X, Chen G, Lv B and Lv J:
MicroRNA-148a/152 cluster restrains tumor stem cell phenotype of
colon cancer via modulating CCT6A. Anticancer Drugs. 33:e610–e621.
2022. View Article : Google Scholar
|
|
16
|
Cox A, Nierenberg D, Camargo O, Lee E,
Khaled AS, Mazar J, Boohaker RJ, Westmoreland TJ and Khaled AR:
Chaperonin containing TCP-1 (CCT/TRiC) is a novel therapeutic and
diagnostic target for neuroblastoma. Front Oncol. 12:9750882022.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z: Regulation of cell cycle
progression by growth factor-induced cell signaling. Cells.
10:33272021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zatulovskiy E, Zhang S, Berenson DF,
Topacio BR and Skotheim JM: Cell growth dilutes the cell cycle
inhibitor Rb to trigger cell division. Science. 369:466–471. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hume S, Dianov GL and Ramadan K: A unified
model for the G1/S cell cycle transition. Nucleic Acids Res.
48:12483–12501. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Temiz E, Koyuncu İ and Sahin E: CCT3
suppression prompts apoptotic machinery through oxidative stress
and energy deprivation in breast and prostate cancers. Free Radic
Biol Med. 165:88–99. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen S, Tian Y, Ju A, Li B, Fu Y and Luo
Y: Suppression of CCT3 inhibits tumor progression by impairing ATP
production and cytoplasmic translation in lung adenocarcinoma. Int
J Mol Sci. 23:39832022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang W, Hu H, Zhang Q, Wu X, Wei F, Yang
F, Gan L, Wang N, Yang X and Guo AY: Regulatory networks in
mechanotransduction reveal key genes in promoting cancer cell
stemness and proliferation. Oncogene. 38:6818–6834. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Wang Y, Wei Y, Wu J, Zhang P,
Shen S, Saiyin H, Wumaier R, Yang X, Wang C and Yu L: Molecular
chaperone CCT3 supports proper mitotic progression and cell
proliferation in hepatocellular carcinoma cells. Cancer Lett.
372:101–109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen X, Chen X, Huang Y, Lin J, Wu Y and
Chen Y: TCP1 increases drug resistance in acute myeloid leukemia by
suppressing autophagy via activating AKT/mTOR signaling. Cell Death
Dis. 12:10582021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang K, He J, Tu C, Xu H, Zhang X, Lv Y
and Song C: Upregulation of CCT3 predicts poor prognosis and
promotes cell proliferation via inhibition of ferroptosis and
activation of AKT signaling in lung adenocarcinoma. BMC Mol Cell
Biol. 23:252022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fu R, Jiang S, Guan Z, Li J, Zhang X and
Chen H: Comprehensive analysis of the expression of chaperonin
containing TCP1 subunits (CCTs) and their influence on prognosis in
hepatocellular carcinoma. Transl Cancer Res. 9:1867–1883. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meng Y, Yang L, Wei X, Luo H, Hu Y, Tao X,
He J, Zheng X, Xu Q, Luo K, et al: CCT5 interacts with cyclin D1
promoting lung adenocarcinoma cell migration and invasion. Biochem
Biophys Res Commun. 567:222–229. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guest ST, Kratche ZR, Bollig-Fischer A,
Haddad R and Ethier SP: Two members of the TRiC chaperonin complex,
CCT2 and TCP1 are essential for survival of breast cancer cells and
are linked to driving oncogenes. Exp Cell Res. 332:223–235. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, Wang L, Cao F, Yu D, Yang J, Yu X,
Dong J, Qin JJ and Guan X: Design, synthesis, and biological
characterization of a potent STAT3 degrader for the treatment of
gastric cancer. Front Pharmacol. 13:9444552022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kasembeli M, Lau WC, Roh SH, Eckols TK,
Frydman J, Chiu W and Tweardy DJ: Modulation of STAT3 folding and
function by TRiC/CCT chaperonin. PLoS Biol. 12:e10018442014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vallin J, Córdoba-Beldad CM and Grantham
J: Sequestration of the transcription factor STAT3 by the molecular
chaperone CCT: A potential mechanism for modulation of STAT3
phosphorylation. J Mol Biol. 433:1669582021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bocchini CE, Nahmod K, Katsonis P, Kim S,
Kasembeli MM, Freeman A, Lichtarge O, Makedonas G and Tweardy DJ:
Protein stabilization improves STAT3 function in autosomal dominant
hyper-IgE syndrome. Blood. 128:3061–3072. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carr AC, Khaled AS, Bassiouni R, Flores O,
Nierenberg D, Bhatti H, Vishnubhotla P, Manuel JP, Santra S and
Khaled AR: Targeting chaperonin containing TCP1 (CCT) as a
molecular therapeutic for small cell lung cancer. Oncotarget.
8:110273–110288. 2017. View Article : Google Scholar
|
|
34
|
Qian T, Cui L, Liu Y, Cheng Z, Quan L,
Zeng T, Huang W, Dai Y, Chen J, Liu L, et al: High expression of
chaperonin-containing TCP1 subunit 3 may induce dismal prognosis in
multiple myeloma. Pharmacogenomics J. 20:563–573. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Danni X, Jiangzheng Z, Huamao S, Yinglian
P, Changcheng Y and Yanda L: Chaperonin containing TCP1 subunit 3
(CCT3) promotes cisplatin resistance of lung adenocarcinoma cells
through targeting the Janus kinase 2/signal transducers and
activators of transcription 3 (JAK2/STAT3) pathway. Bioengineered.
12:7335–7347. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cui X, Hu ZP, Li Z, Gao PJ and Zhu JY:
Overexpression of chaperonin containing TCP1, subunit 3 predicts
poor prognosis in hepatocellular carcinoma. World J Gastroenterol.
21:8588–8604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morimoto E, Inagaki K, Komatsubara M,
Terasaka T, Itoh Y, Fujisawa S, Sasaki E, Nishiyama Y, Hara T and
Wada J: Effects of Wnt-β-Catenin signaling and sclerostin on the
phenotypes of rat pheochromocytoma PC12 cells. J Endocr Soc.
6:bvac1212022. View Article : Google Scholar
|
|
38
|
Li W, Xu Y, Wang X, Cao G, Bu W, Wang X,
Fang Z, Xu Y, Dong M and Tao Q: circCCT3 modulates vascular
endothelial growth factor A and Wnt signaling to enhance colorectal
cancer metastasis through sponging miR-613. DNA Cell Biol.
39:118–125. 2020. View Article : Google Scholar
|
|
39
|
Qu H, Zhu F, Dong H, Hu X and Han M:
Upregulation of CCT-3 induces breast cancer cell proliferation
through miR-223 competition and Wnt/β-Catenin signaling pathway
activation. Front Oncol. 10:5331762020. View Article : Google Scholar
|
|
40
|
Tang N, Cai X, Peng L, Liu H and Chen Y:
TCP1 regulates Wnt7b/β-catenin pathway through P53 to influence the
proliferation and migration of hepatocellular carcinoma cells.
Signal Transduct Target Ther. 5:1692020. View Article : Google Scholar
|
|
41
|
Wang Q, Huang WR, Chih WY, Chuang KP,
Chang CD, Wu Y, Huang Y and Liu HJ: Cdc20 and molecular chaperone
CCT2 and CCT5 are required for the Muscovy duck reovirus
p10.8-induced cell cycle arrest and apoptosis. Vet Microbiol.
235:151–163. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rivlin N, Katz S, Doody M, Sheffer M,
Horesh S, Molchadsky A, Koifman G, Shetzer Y, Goldfinger N, Rotter
V and Geiger T: Rescue of embryonic stem cells from cellular
transformation by proteomic stabilization of mutant p53 and
conversion into WT conformation. Proc Natl Acad Sci USA.
111:7006–7011. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Trinidad AG, Muller PA, Cuellar J, Klejnot
M, Nobis M, Valpuesta JM and Vousden KH: Interaction of p53 with
the CCT complex promotes protein folding and wild-type p53
activity. Mol Cell. 50:805–817. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Monteith J and McMahon SB: p53: The TRiC
is knowing when to fold 'em. Mol Cell. 50:781–782. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Arsic N, Slatter T, Gadea G, Villain E,
Fournet A, Kazantseva M, Allemand F, Sibille N, Seveno M, de Rossi
S, et al: Δ133p53β isoform pro-invasive activity is regulated
through an aggregation-dependent mechanism in cancer cells. Nat
Commun. 12:54632021. View Article : Google Scholar
|
|
46
|
Lee SC and Chan J: Proteomic
identification of chaperonin-containing tail-less complex
polypeptide-1 gamma subunit as a p53-responsive protein in colon
cancer cells. Cancer Genomics Proteomics. 9:101–108.
2012.PubMed/NCBI
|
|
47
|
Ooe A, Kato K and Noguchi S: Possible
involvement of CCT5, RGS3, and YKT6 genes up-regulated in
p53-mutated tumors in resistance to docetaxel in human breast
cancers. Breast Cancer Res Treat. 101:305–315. 2007. View Article : Google Scholar
|
|
48
|
Liu Q, Liu J, He N, Zhang M, Wu L, Chen X,
Zhu J, Ran F, Chen Q and Zhang H: CRISPR/Cas12a coupling with
magnetic nanoparticles and cascaded strand displacement reaction
for ultrasensitive fluorescence determination of exosomal miR-21.
Molecules. 27:53382022. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ke L, Ma H, Zhang Q, Wang Y, Xia P, Yu L,
Lv W and Hu J: The pattern of lymph node metastasis in peripheral
pulmonary nodules patients and risk prediction models. Front Surg.
9:9813132022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shao F, Chen Y, Xu H, Chen X, Zhou J, Wu
Y, Tang Y, Wang Z, Zhang R, Lange T, et al: Metabolic obesity
phenotypes and risk of lung cancer: A prospective cohort study of
450,482 UK biobank participants. Nutrients. 14:33702022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Villalobos-Manzo R, Ríos-Castro E,
Hernández-Hernández JM, Oza G, Medina MA and Tapia-Ramírez J:
Identification of transferrin receptor 1 (TfR1) overexpressed in
lung cancer cells, and internalization of magnetic Au-CoFe(2)O(4)
core-shell nanoparticles functionalized with its ligand in a
cellular model of small cell lung cancer (SCLC). Pharmaceutics.
14:17152022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu M and Gong J: Prognostic signature,
immune features, and therapeutic responses of a novel
ubiquitination-related gene signature in lung adenocarcinoma. J
Oncol. 2022:25246492022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Platini H, Ferdinand E, Kohar K, Prayogo
SA, Amirah S, Komariah M and Maulana S: Neutrophil-to-lymphocyte
ratio and platelet-to-lymphocyte ratio as prognostic markers for
advanced non-small-cell lung cancer treated with immunotherapy: A
systematic review and meta-analysis. Medicina (Kaunas).
58:10692022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou F, Qian C, Chen T and Zang X:
MiR-96-5p facilitates lung adenocarcinoma cell phenotypes by
inhibiting FHL1. Comput Math Methods Med. 2022:78912222022.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu S, Cao P, Kong K, Han P, Yue J, Deng Y,
Li F and Zhao B: circCNN2 accelerates cell proliferation and
invasion in lung squamous cell carcinoma via regulating
miR-184/E2F1 and Activating MAPK signaling pathway. Dis Markers.
2022:63290972022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Barak D, Engelberg S, Assaraf YG and
Livney YD: Selective targeting and eradication of various human
non-small cell lung cancer cell lines using self-assembled
aptamer-decorated nanoparticles. Pharmaceutics. 14:16502022.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Skonieczna M, Kasprzycka A, Jelen M and
Morak-Mlodawska B: Tri- and pentacyclic azaphenothiazine as
pro-apoptotic agents in lung carcinoma with a protective potential
to healthy cell lines. Molecules. 27:52552022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cui XY, Park SH and Park WH: Anti-cancer
effects of auranofin in human lung cancer cells by increasing
intracellular ROS levels and depleting GSH levels. Molecules.
27:52072022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shi H, Zhang Y, Wang Y, Fang P, Liu Y and
Li W: Restraint of chaperonin containing T-complex protein-1
subunit 3 has antitumor roles in non-small cell lung cancer via
affection of YAP1. Toxicol Appl Pharmacol. 439:1159262022.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liu YJ, Chang YJ, Kuo YT and Liang PH:
Targeting β-tubulin/CCT-β complex induces apoptosis and suppresses
migration and invasion of highly metastatic lung adenocarcinoma.
Carcinogenesis. 41:699–710. 2020. View Article : Google Scholar
|
|
61
|
Ying Z, Tian H, Li Y, Lian R, Li W, Wu S,
Zhang HZ, Wu J, Liu L, Song J, et al: CCT6A suppresses SMAD2 and
promotes prometastatic TGF-β signaling. J Clin Invest.
127:1725–1740. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang H, Wang X, Xu L, Lin Y and Zhang J:
CCT6A and CHCHD2 are coamplified with EGFR and associated with the
unfavorable clinical outcomes of lung adenocarcinoma. Dis Markers.
2022:15601992022.PubMed/NCBI
|
|
63
|
Zhang T, Shi W, Tian K and Kong Y:
Chaperonin containing t-complex polypeptide 1 subunit 6A correlates
with lymph node metastasis, abnormal carcinoembryonic antigen and
poor survival profiles in non-small cell lung carcinoma. World J
Surg Oncol. 18:1562020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tewelde B, Tamire M and Kaba M: Breast
self-examination practice and predictors among female secondary
school teachers in Addis Ababa, Ethiopia: Using the health belief
model. BMC Womens Health. 22:3172022. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liu N, Yang DW, Wu YX, Xue WQ, Li DH,
Zhang JB, He YQ and Jia WH: Burden, trends, and risk factors for
breast cancer in China from 1990 to 2019 and its predictions until
2034: An up-to-date overview and comparison with those in Japan and
South Korea. BMC Cancer. 22:8262022. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
de Freitas GB, Penteado L, Miranda MM,
Filassi JR, Baracat EC and Linhares IM: The circulating 70 kDa heat
shock protein (HSPA1A) level is a potential biomarker for breast
carcinoma and its progression. Sci Rep. 12:130122022. View Article : Google Scholar
|
|
67
|
Tang M, O'Grady S, Crown J and Duffy MJ:
MYC as a therapeutic target for the treatment of triple-negative
breast cancer: Preclinical investigations with the novel MYC
inhibitor, MYCi975. Breast Cancer Res Treat. 195:105–115. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Tam S, Al-Zubaidi Y, Rahman MK, Bourget K,
Zhou F and Murray M: The ixabepilone and vandetanib combination
shows synergistic activity in docetaxel-resistant MDA-MB-231 breast
cancer cells. Pharmacol Rep. 74:998–1010. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Kotani H, Masuda N, Yamashita T, Naito Y,
Taira T, Inoue K, Takahashi M, Yonemori K, Toyoizumi S, Mori Y, et
al: Efficacy and safety of talazoparib in Japanese patients with
germline BRCA-mutated locally advanced or metastatic breast cancer:
Results of the phase 1 dose-expansion study. Breast Cancer.
29:1088–1098. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xu WX, Song W, Jiang MP, Yang SJ, Zhang J,
Wang DD and Tang JH: Systematic characterization of expression
profiles and prognostic values of the eight subunits of the
chaperonin TRiC in breast cancer. Front Genet. 12:6378872021.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Huang K, Zeng Y, Xie Y, Huang L and Wu Y:
Bioinformatics analysis of the prognostic value of CCT6A and
associated signalling pathways in breast cancer. Mol Med Rep.
19:4344–4352. 2019.PubMed/NCBI
|
|
72
|
Macario AJL and Conway de Macario E:
Chaperonins in cancer: Expression, function, and migration in
extracellular vesicles. Semin Cancer Biol. 86(Pt 1): 26–35. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu Q, Qi Y, Kong X, Wang X, Zhang W, Zhai
J, Yang Y, Fang Y and Wang J: Molecular and clinical
characterization of CCT2 expression and prognosis via large-scale
transcriptome profile of breast cancer. Front Oncol. 11:6144972021.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Ghozlan H, Showalter A, Lee E, Zhu X and
Khaled AR: Chaperonin-Containing TCP1 Complex (CCT) promotes breast
cancer growth through correlations with key cell cycle regulators.
Front Oncol. 11:6638772021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Bassiouni R, Nemec KN, Iketani A, Flores
O, Showalter A, Khaled AS, Vishnubhotla P, Sprung RW Jr, Kaittanis
C, Perez JM and Khaled AR: Chaperonin Containing TCP-1 protein
level in breast cancer cells predicts therapeutic application of a
cytotoxic peptide. Clin Cancer Res. 22:4366–4379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Satriano L, Lewinska M, Rodrigues PM,
Banales JM and Andersen JB: Metabolic rearrangements in primary
liver cancers: Cause and consequences. Nat Rev Gastroenterol
Hepatol. 16:748–766. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Trefts E, Gannon M and Wasserman DH: The
liver. Curr Biol. 27:R1147–R1151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Cheng ML, Nakib D, Perciani CT and
MacParland SA: The immune niche of the liver. Clin Sci (Lond).
135:2445–2466. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Villanueva A: Hepatocellular carcinoma. N
Engl J Med. 380:1450–1462. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gao YX, Yang TW, Yin JM, Yang PX, Kou BX,
Chai MY, Liu XN and Chen DX: Progress and prospects of biomarkers
in primary liver cancer (Review). Int J Oncol. 57:54–66.
2020.PubMed/NCBI
|
|
81
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Hepatocellular carcinoma. Nat Rev Dis
Primers. 7:72021. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Moon H, Park H, Chae MJ, Choi HJ, Kim DY
and Ro SW: Activated TAZ induces liver cancer in collaboration with
EGFR/HER2 signaling pathways. BMC Cancer. 22:4232022. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Keenan BP, Fong L and Kelley RK:
Immunotherapy in hepatocellular carcinoma: The complex interface
between inflammation, fibrosis, and the immune response. J
Immunother Cancer. 7:2672019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang H, Lu Z and Zhao X: Tumorigenesis,
diagnosis, and therapeutic potential of exosomes in liver cancer. J
Hematol Oncol. 12:1332019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Mintz KJ and Leblanc RM: The use of
nanotechnology to combat liver cancer: Progress and perspectives.
Biochim Biophys Acta Rev Cancer. 1876:1886212021. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Anwanwan D, Singh SK, Singh S, Saikam V
and Singh R: Challenges in liver cancer and possible treatment
approaches. Biochim Biophys Acta Rev Cancer. 1873:1883142010.
View Article : Google Scholar
|
|
88
|
Rebouissou S and Nault JC: Advances in
molecular classification and precision oncology in hepatocellular
carcinoma. J Hepatol. 72:215–229. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Qian EN, Han SY, Ding SZ and Lv X:
Expression and diagnostic value of CCT3 and IQGAP3 in
hepatocellular carcinoma. Cancer Cell Int. 16:552016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang F, Wang R, Li Q, Qu X, Hao Y, Yang J,
Zhao H, Wang Q, Li G, Zhang F, et al: A transcriptome profile in
hepatocellular carcinomas based on integrated analysis of
microarray studies. Diagn Pathol. 12:42017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yunna C, Mengru H, Lei W and Weidong C:
Macrophage M1/M2 polarization. Eur J Pharmacol. 877:1730902020.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wong N, Chan A, Lee SW, Lam E, To KF, Lai
PB, Li XN, Liew CT and Johnson PJ: Positional mapping for amplified
DNA sequences on 1q21-q22 in hepatocellular carcinoma indicates
candidate genes over-expression. J Hepatol. 38:298–306. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Liu J, Huang L, Zhu Y, He Y, Zhang W, Lei
T, Xuan J, Xiao B, Li L, Zhou Q and Sun Z: Exploring the expression
and prognostic value of the TCP1 ring complex in hepatocellular
carcinoma and overexpressing its subunit 5 Promotes HCC
tumorigenesis. Front Oncol. 11:7396602021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zeng G, Wang J and Huang Y, Lian Y, Chen
D, Wei H, Lin C and Huang Y: Overexpressing CCT6A contributes to
cancer cell growth by affecting the G1-To-S phase transition and
predicts a negative prognosis in hepatocellular carcinoma. Onco
Targets Ther. 12:10427–10439. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Shaath H, Vishnubalaji R, Elango R,
Kardousha A, Islam Z, Qureshi R, Alam T, Kolatkar PR and Alajez NM:
Long non-coding RNA and RNA-binding protein interactions in cancer:
Experimental and machine learning approaches. Semin Cancer Biol.
86(Pt 3): 325–345. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Pereira B, Billaud M and Almeida R:
RNA-Binding proteins in cancer: Old players and new actors. Trends
Cancer. 3:506–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Iino K, Mitobe Y, Ikeda K, Takayama KI,
Suzuki T, Kawabata H, Suzuki Y, Horie-Inoue K and Inoue S:
RNA-binding protein NONO promotes breast cancer proliferation by
post-transcriptional regulation of SKP2 and E2F8. Cancer Sci.
111:148–159. 2020. View Article : Google Scholar
|
|
98
|
Sondergaard JN, Sommerauer C, Atanasoai I,
Hinte LC, Geng K, Guiducci G, Brautigam L, Aouadi M, Stojic L,
Barragan I and Kutter C: CCT3-LINC00326 axis regulates
hepatocarcinogenic lipid metabolism. Gut. 71:2081–2092. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Zhang Z, Xu L and Sun C: Comprehensive
characterization of cancer genes in hepatocellular carcinoma
genomes. Oncol Lett. 15:1503–1510. 2018.PubMed/NCBI
|
|
100
|
Xu J, Zhang Y, Liu C, Yan P and Yang Z:
Roles of the miR-139-5p/CCT5 axis in hepatocellular carcinoma: A
bioinformatic analysis. Int J Med Sci. 18:3556–3564. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zayegh O, Hmidi ZS, Nawlo AA, Al-Mouakeh
A, Amin B, Banjah B, Chammout A and Alsayid M: Awareness and
knowledge of colorectal cancer screening among medical students at
the university of aleppo: A cross-sectional study. Avicenna J Med.
12:54–60. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Cummings A, Foster R, Calman L, Permyakova
NV, Bridges J, Wiseman T, Corbett T, Smith PWF and Foster C:
Quality of life and health status in older adults (>/=65 years)
up to five years following colorectal cancer treatment: Findings
from the ColoREctal Wellbeing (CREW) cohort study. PLoS One.
17:e02700332022. View Article : Google Scholar
|
|
103
|
La Vecchia S and Sebastián C: Metabolic
pathways regulating colorectal cancer initiation and progression.
Semin Cell Dev Biol. 98:63–70. 2020. View Article : Google Scholar
|
|
104
|
Liu X, Yang K, Li Z and Liu J: MMP2
polymorphisms and colorectal cancer susceptibility in a Chinese Han
Population. Int J Gen Med. 15:6009–6019. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Pan S, Mei W, Huang L, Tao Y, Xu J and
Ruan Y: Prediction of postoperative survival in young colorectal
cancer patients: A cohort study based on the SEER database. J
Immunol Res. 2022:27366762022. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Li J, Ma X, Chakravarti D, Shalapour S and
DePinho RA: Genetic and biological hallmarks of colorectal cancer.
Genes Dev. 35:787–820. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Dienstmann R, Vermeulen L, Guinney J,
Kopetz S, Tejpar S and Tabernero J: Consensus molecular subtypes
and the evolution of precision medicine in colorectal cancer. Nat
Rev Cancer. 17:79–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Watson AJ and Collins PD: Colon cancer: A
civilization disorder. Dig Dis. 29:222–228. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Goldstein DA, Zeichner SB, Bartnik CM,
Neustadter E and Flowers CR: Metastatic colorectal cancer: A
systematic review of the value of current therapies. Clin
Colorectal Cancer. 15:1–6. 2016. View Article : Google Scholar :
|
|
110
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Qian-Lin Z, Ting-Feng W, Qi-Feng C,
Min-Hua Z and Ai-Guo L: Inhibition of cytosolic chaperonin CCTζ-1
expression depletes proliferation of colorectal carcinoma in vitro.
J Surg Oncol. 102:419–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Coghlin C, Carpenter B, Dundas SR, Lawrie
LC, Telfer C and Murray GI: Characterization and over-expression of
chaperonin t-complex proteins in colorectal cancer. J Pathol.
210:351–357. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Yokota S, Yamamoto Y, Shimizu K, Momoi H,
Kamikawa T, Yamaoka Y, Yanagi H, Yura T and Kubota H: Increased
expression of cytosolic chaperonin CCT in human hepatocellular and
colonic carcinoma. Cell Stress Chaperones. 6:345–350. 2001.
View Article : Google Scholar
|
|
114
|
Yang X, Tong Y, Ye W and Chen L: HOXB2
increases the proliferation and invasiveness of colon cancer cells
through the upregulation of CCT6A. Mol Med Rep. 25:1742022.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Liu YJ, Kumar V, Lin YF and Liang PH:
Disrupting CCT-β : β-tubulin selectively kills CCT-β overexpressed
cancer cells through MAPKs activation. Cell Death Dis. 8:e30522017.
View Article : Google Scholar
|
|
116
|
Park SH, Jeong S, Kim BR, Jeong YA, Kim
JL, Na YJ, Jo MJ, Yun HK, Kim DY, Kim BG, et al: Activating CCT2
triggers Gli-1 activation during hypoxic condition in colorectal
cancer. Oncogene. 39:136–150. 2020. View Article : Google Scholar
|
|
117
|
Sun H, Wang Y, Jing HY, Yang XY, Shi XX,
Zhang JH, Yu YX, Gao L, Wang XY, Li WH and Yu L:
Chaperonin-Containing TCP1 Subunit 6A Is a prognostic potential
biomarker that correlates with the presence of immune infiltrates
in colorectal cancer. Front Genet. 12:6298562021. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Hu J, Han C, Zhong J, Liu H, Liu R, Luo W,
Chen P and Ling F: Dynamic network biomarker of pre-exhausted
CD8(+) T cells contributed to T cell exhaustion in colorectal
cancer. Front Immunol. 12:6911422021. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Lu L, Li ZJ, Li LF, Wu WK, Shen J, Zhang
L, Chan RL, Yu L, Liu YW, Ren SX, et al: Vascular-targeted TNFα
improves tumor blood vessel function and enhances antitumor
immunity and chemotherapy in colorectal cancer. J Control Release.
210:134–146. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Lu L, Li ZJ, Li LF, Shen J, Zhang L, Li
MX, Xiao ZG, Wang JH and Cho CH: A novel vascular-targeting peptide
for gastric cancer delivers low-dose TNFα to normalize the blood
vessels and improve the anti-cancer efficiency of 5-fluorouracil.
Peptides. 97:54–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Liao Q, Ren Y, Yang Y, Zhu X, Zhi Y, Zhang
Y, Chen Y, Ding Y and Zhao L: CCT8 recovers WTp53-suppressed cell
cycle evolution and EMT to promote colorectal cancer progression.
Oncogenesis. 10:842021. View Article : Google Scholar : PubMed/NCBI
|