Introduction
Diabetes mellitus (DM) induces various
cardiovascular complications in the diabetic population, which has
become the major cause of morbidity and mortality (1). Diabetic cardiomyopathy is a distinct
clinical entity inducing functional, biochemical and morphological
abnormalities in the heart, ultimately leading to heart failure
(2). However, the mechanisms
leading to cardiac changes are not fully comprehended.
Diabetic cardiomyopathy is accompanied by
mitochondrial injury (3,4). Mitochondria undergo frequent fusion
and fission, and the balance of these opposing processes regulates
mitochondrial morphology (5).
Mitochondrial fusion serves to keep up a tubular mitochondrial
network and to optimize mitochondrial function, which is regulated
by large GTPases, such as mitofusin-1 and -2 (Mfn1 and Mfn2).
Previous studies have reported that Mfn2 polymorphic genes were
present in type 1 and 2 diabetes patients. Type 2 diabetes
downregulates the expression of Mfn2 mRNA in skeletal muscle
(6). The overexpression of Mfn2 in
diabetic rats was reported to have the potential to protect the
kidney by inhibiting the activation of p38 and the accumulation of
ROS; to prevent mitochondrial dysfunction and reduce the synthesis
of collagen IV (7). Of all cell
types and tissues, Mfn2 is predominantly expressed in the heart
(8), at the same time its
functional role in the cardiac myocyte is poorly understood, and
there are no data regarding the changes of Mfn2 in the DM heart.
Thus, we wanted to examine whether Mfn2 changes in diabetic
cardiomyopathy and whether this change of cardiac Mfn2 is
correlated with the pathological process of DM, since it may
provide a novel strategy for the treatment of DM.
Excessive oxidative stress has been associated with
the pathology and complications of diabetes. Hyperglycemia causes
oxidative stress and cell death. Oxidative stress may induce
mitochondrial fragmentation, which is correlated with mitochondrial
fusion and fission. In the case that cardiac Mfn2 changes with the
development of DM, it is of importance to examine whether there is
any association with oxidative stress injury.
Apoptosis occurs in diabetic cardiomyopathy.
Caspases are a large protein family of cysteine proteases that have
been specifically linked with cell death (apoptosis). Among them,
caspase 3 is a frequently activated death protease, catalyzing the
specific cleavage of many key cellular proteins. Thus, we measured
the changes of caspase 3 during the development of DM, and aimed to
analyze the role of Mfn2 in the process.
Materials and methods
Animals
Thirty-six male Sprague-Dawley rats (200–250 g) were
purchased from the Animal Center of the Bengbu Medical College. The
rats were fed normal chow and had free access to water. They were
kept at a constant temperature of (21±1°C) with a fixed 12-h
light/dark cycle. All animal procedures were conducted in
accordance with the United States National Institutes of Health
Guide and were approved by the Animal Use and Care Committee of the
Bengbu Medical College.
Chemicals and reagents
Streptozotocin (STZ) was purchased from Sigma (St.
Louis, MO, USA). Malondialdehyde (MDA) and superoxide dismutase
(SOD) kits were purchased from the Nanjing Jiancheng Bioengineering
Institute (China). The caspase 3 kit was purchased from Shanghai
Genmed Scientifics Inc., (China). The primers for Mfn-2 were
5′-CTCAGGAGCAGCGGGTTTATTGTCT-3′ and
5′-TGTCGAGGGACCAGCATGTCTATCT-3′, the amplified fragment length was
412 bp, while the primers for β-actin were
5′-GATGGTGGGTATGGGTCAGAAGGAC-3′ and 5′-GCTCATTGCCGATAGTGATGACT-3′,
the amplified fragment length was 630 bp. Any other chemicals used
were of the highest purity available.
Induction of diabetes and experimental
protocol
Diabetes was induced in overnight-fasted rats by
administering a single intraperitoneal (i.p.) injection of 55 mg/kg
STZ freshly dissolved in 0.1 mol/l sodium citrate buffer (pH 4.5).
The control group was injected with a similar volume of sodium
citrate buffer alone. Rats with a fasting blood glucose (FBG)level
>16.7 mmol/l became diabetic 72 h subsequent to injection.
Animals were randomly divided into control groups of 4, 8 and 12W,
corresponding to diabetes at the fourth (DM4W), the eighth (DM8W)
and the twelfth week (DM12W) groups, respectively (n=6).
Detection of fasting blood glucose (FBG),
body weight (BW) and heart weight (HW)
FBG level and BW were measured every 4 weeks
subsequent to injections of STZ. The ratio of heart weight to body
weight (HW/BW) was determined to indicate the degree of cardiac
hypertrophy.
Detection of MDA content and SOD
activity
At the end of the experimental period, 0.1 g heart
tissue was homogenized in ice-cold PBS. MDA content and SOD
activity were measured by commercially available kits, according to
the manufacturer’s instructions.
Detection of Mfn2 mRNA by RT-PCR
RT-PCR was used to detect Mfn2 mRNA expression in
the heart. Briefly, total RNA was extracted with TRIzol, according
to the manufacturer’s instructions. Total RNA (2 μg)were reverse
transcribed to cDNA, and PCR was performed following the routine
method. PCR products were analyzed on 1% agarose gel. Densitometry
result for Mfn2 gene was compared with the corresponding β-actin
levels to account for loading differences.
Detection of caspase-3 activity
Caspase-3 activity was measured by commercially
available kits, according to the manufacturer’s instructions.
Statistical analysis
Values were expressed as the mean ± SEM. Statistical
comparisons were carried out by one-way variance analysis and the
Newman-Keuls test. P<0.05 were considered to indicate a
statistically significant difference.
Results
Changes of FBG, BW, HW and the
HW/BW-ratio
The changes in FBG, BW, HW and the HW/BW-ratio at
different stages of diabetes are shown in Table I. In contrast to the control group,
the FBG in DM groups was increased significantly, and there were no
differences at different stages of DM. Compared with the control
animals, BW significantly decreased at 4, 8 and 12W after STZ
injection, and there were statistically significant differences in
the different stages of DM. HW/BW, however, was significantly
increased in different DM stages corresponding to the control
groups (Table I).
 | Table IChanges of fasting blood glucose,
heart weight, body weight and heart weight/body weight in the
different groups. |
Table I
Changes of fasting blood glucose,
heart weight, body weight and heart weight/body weight in the
different groups.
| Fasting blood glucose
(mmol/l) | 4W | 8W | 12W |
|---|
| Control | 6.05±1.02 | 5.88±1.14 | 5.75±0.63 |
| DM | 25.30±2.99a | 25.65±3.19a | 27.88±4.25a |
|
| Body weight (g) | 4W | 8W | 12W |
|
| Control | 382.67±5.95 | 440.53±3.84b | 525.00±5.36b,c |
| DM | 235.58±6.43a | 185.46±3.85a,d | 147.50±5.42a,d,e |
|
| Heart weight
(mg) | 4W | 8W | 12W |
|
| Control | 1392.66±35.76 | 1528.63±43.25b | 1861.7±181b,c |
| DM | 925.45±22.6a | 830.34±37.93a | 784.68±18.75a |
|
| HW/BW
(mgxg−1) | 4W | 8W | 12W |
|
| Control | 3.64±0.04 | 3.47±0.07 | 3.54±0.10 |
| DM | 3.93±0.21 | 4.48±0.11a,d | 5.32±0.06a,d,e |
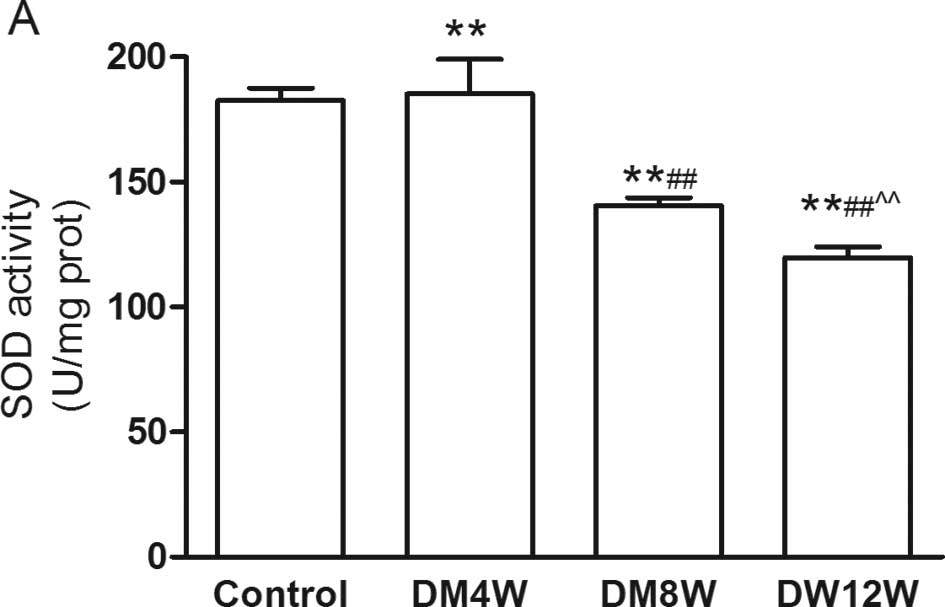
Changes of MDA content and SOD activity
in heart tissue
There were no statistically significant differences
of MDA content and SOD activity in the 4, 8 and 12W control groups
(data not shown). In contrast to the control group, MDA content was
increased, while SOD activity was decreased with the development of
DM (Fig. 1). This suggested that
with the development of DM, oxide stress injury was increased.
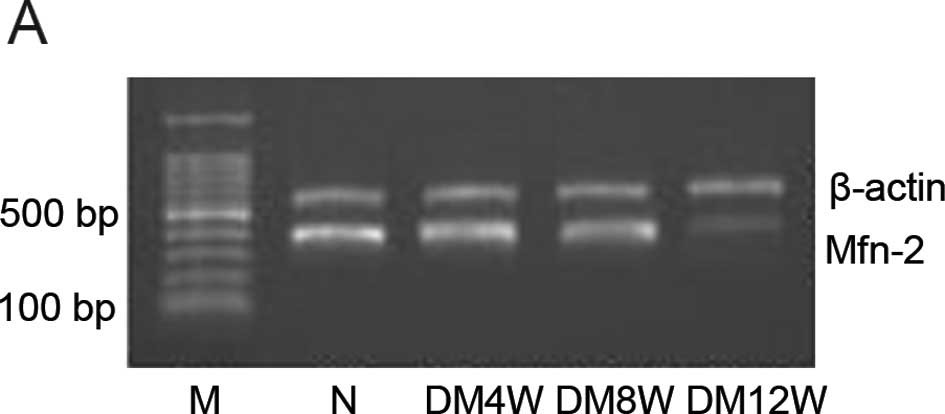
Change of Mfn2 mRNA level in heart
In contrast to control groups, the Mfn2 mRNA
expression level in the heart decreased in the different stages of
DM and was further decreased with the development of DM (Fig. 2).
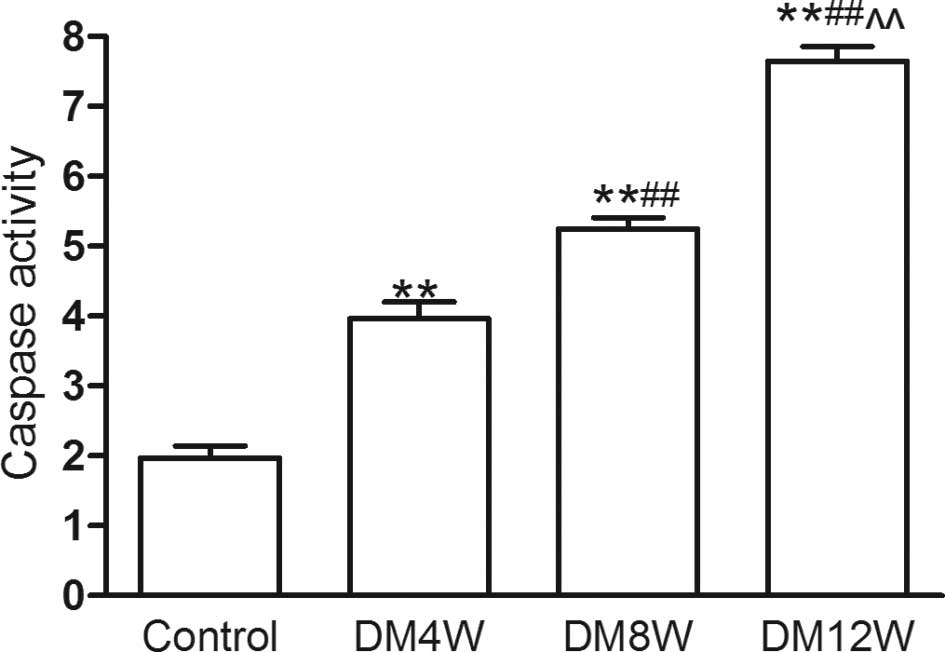
Change of caspase-3 activity in heart
tissue
In contrast to the control group, caspase-3 activity
increased with the development of DM (Fig. 3). It suggested that with the
development of DM apoptosis was aggravated.
Discussion
In the present study, we determined that with the
development of DM, cardiac oxidative stress and apoptosis were
aggravated, the cardiac Mfn2 mRNA level decreased, suggesting that
the downregulation of Mfn2 is likely to be correlated with
oxidative stress injury and apoptosis in DM. This is the first
study to report the correlation of DM and cardiac Mfn2
expressions.
STZ is a drug that causes sustained insulin
deficiency and elevated serum glucose levels by selectively
destroying pancreatic β-cells to induce DM. Investigators had
observed that STZ-induced DM imitated the structural and cellular
abnormalities of DM, including cardiac apoptosis, hypertrophy,
fibrosis and cardiac inflammation. Consequently, in our experiment
we selected STZ to induce DM. In the present study, FBG levels were
demonstrated to be elevated in different stages of DM, confirming
an occurring abnormality of the glucose metabolism in a STZ-induced
diabetes model.
Evidence has shown reactive oxygen species (ROS) to
be accumulated at the onset and throughout the development of
diabetic cardiomyopathy (9,10).
Free radical production was increased in DM patients, while
hyperglycemia appeared to be the key factor in the generating of
ROS, which lowers the concentrations of antioxidant enzymes.
Oxidative stress plays a crucial role in complications of diabetes
and induces the production of highly reactive oxygen radicals that
are toxic to cells, while having been linked to protein glycation
and/or glucose auto-oxidation. The present results have proven a
decreased SOD activity and an elevated MDA content in the diabetic
heart, indicating that oxidative stress injury is aggravated with
the development of diabetes. SOD scavenges super oxide radicals by
converting them to H2O2 and oxygen. The
decrease in SOD activity in diabetic rats may result from the
inefficient scavenging of ROS, indicating that oxidative enzymes
were inactivated, while deleterious effects occurred most likely
due to accumulated super oxide radicals, or because the enzymes
were glycosylated.
Mfn2 encodes a mitochondrial protein that is
involved in maintaining the mitochondrial network and regulates
mitochondrial metabolism and intracellular signaling. Elimination
of Mfn2 in fibroblasts, L6E9 myotubes or Opa1 in mouse embryonic
fibroblasts increased mitochondrial fission and decreased
mitochondrial membrane potential, oxygen consumption, glucose and
palmitate oxidation as well as respiratory complex activity
(11–13). In recent years, Mfn2 has been
reported to be correlated also with the pathological changes of
some diseases related to oxidative stress, energy metabolism and
mitochondrial apoptotic signaling. There have been different
opinions regarding the role of Mfn2. The overexpression of Mfn2
alleviated high-glucose-induced glomerular mesangial cell
proliferation and elevated apoptosis (14). Mfn2 gene can significantly promote
apoptosis via Bax and may inhibit proliferation in hepatocellular
carcinoma cells (15). Mfn2
silencing inhibited oxidative stress-induced apoptosis in H9c2
cells (16). Other authors,
however, reported that Mfn2 deficiency exacerbated renal epithelial
cell injury by promoting Bax-mediated mitochondrial outer membrane
injury and apoptosis (17). The
expression of Mfn2 was markedly downregulated in vascular smooth
muscle cells in spontaneously hypertensive rats (SHR) (18). The overexpression of Mfn2 inhibited
the proliferation of hyper-proliferative vascular smooth muscle
cells in vitro and in vivo (19). In our experiment, we observed that
with the development of diabetes, and the aggravation of oxidative
stress injury in DM, caspase-3 activity increased, while Mfn2 mRNA
expression was decreased, suggesting that the decrease of the Mfn2
expression may be crucial to diabetes pathophysiology, as well as
being closely associated with oxidative stress injury and apoptosis
in DM.
In conclusion, we found that with the development of
diabetes, along with an aggravation of oxidative stress and
apoptosis, Mfn2 expression was decreased, suggesting that Mfn2
might be an important regulator of diabetes. However, additional
studies are required to confirm this hypothesis.
Acknowledgements
This study was supported by research grants from the
Chinese National Natural Science Foundation (no. 81000074) and from
the Anhui Province Natural Science Foundation (no. 090413097).
References
|
1
|
Zimmet P, Alberti KG and Shaw J: Global
and societal implications of the diabetes epidemic. Nature.
414:782–787. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Voulgari C, Papadogiannis D and
Tentolouris N: Diabetic cardiomyopathy: from the pathophysiology of
the cardiac myocytes to current diagnosis and management
strategies. Vasc Health Risk Manag. 6:883–903. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bugger H and Abel ED: Mitochondria in the
diabetic heart. Cardiovasc Res. 88:229–240. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Duncan JG: Mitochondrial dysfunction in
diabetic cardiomyopathy. Biochim Biophys Acta. 1813:1351–1359.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang P, Galloway CA and Yoon Y: Control
of mitochondrial morphology through differential interactions of
mitochondrial fusion and fission proteins. PLoS One. 6:e206552011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bach D, Naon D, Pich S, Soriano FX, Vega
N, Rieusset J, Laville M, Guillet C, Boirie Y, Wallberg-Henriksson
H, Manco M, Calvani M, Castagneto M, Palacín M, Mingrone G, Zierath
JR, Vidal H and Zorzano A: Expression of Mfn2, the
Charcot-Marie-Tooth neuropathy type 2A gene, in human skeletal
muscle: effects of type 2 diabetes, obesity, weight loss, and the
regulatory role of tumor necrosis factor alpha and interleukin-6.
Diabetes. 54:2685–2693. 2005. View Article : Google Scholar
|
|
7
|
Tang WX, Wu WH, Zeng XX, Bo H and Huang
SM: Early protective effect of mitofusion 2 overexpression in
STZ-induced diabetic rat kidney. Endocrine. 41:236–247. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Papanicolaou KN, Khairallah RJ, Ngoh GA,
Chikando A, Luptak I, O’Shea KM, Riley DD, Lugus JJ, Colucci WS,
Lederer WJ, Stanley WC and Walsh K: Mitofusin-2 maintains
mitochondrial structure and contributes to stress-induced
permeability transition in cardiac myocytes. Mol Cell Biol.
31:1309–1328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cai L and Kang YJ: Oxidative stress and
diabetic cardiomyopathy: a brief review. Cardiovasc Toxicol.
1:181–193. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wold LE, Ceylan-Isik AF and Ren J:
Oxidative stress and stress signaling: menace of diabetic
cardiomyopathy. Acta Pharmacol Sin. 26:908–917. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pich S, Bach D, Briones P, Liesa M, Camps
M, et al: The Charcot-Marie-Tooth type 2A gene product, Mfn2,
up-regulates fuel oxidation through expression of OXPHOS system.
Hum Mol Genet. 14:1405–1415. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bach D, Pich S, Soriano FX, Vega N,
Baumgartner B, et al: Mitofusin-2 determines mitochondrial network
architecture and mitochondrial metabolism. A novel regulatory
mechanism altered in obesity. J Biol Chem. 278:17190–17197. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen H, Chomyn A and Chan DC: Disruption
of fusion results in mitochondrial heterogeneity and dysfunction. J
Biol Chem. 280:26185–26192. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wan-Xin T, Tian-Lei C, Ben W, Wei-Hua W
and Ping F: Effect of mitofusin 2 overexpression on the
proliferation and apoptosis of high-glucose-induced rat glomerular
mesangial cells. J Nephrol. Feb 7–2012.(Epub ahead of print).
|
|
15
|
Wang W, Lu J, Zhu F, Wei J, Jia C, Zhang
Y, Zhou L, Xie H and Zheng S: Pro-apoptotic and anti-proliferative
effects of mitofusin-2 via Bax signaling in hepatocellular
carcinoma cells. Med Oncol. 29:70–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen T, Zheng M, Cao C, Chen C, Tang J,
Zhang W, Cheng H, Chen KH and Xiao RP: Mitofusin-2 is a major
determinant of oxidative stress-mediated heart muscle cell
apoptosis. J Biol Chem. 282:23354–23361. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gall JM, Wang Z, Liesa M, Molina A, Havasi
A, Schwartz JH, Shirihai O, Borkan SC and Bonegio RG: Role of
mitofusin 2 in the renal stress response. PLoS One. 7:e310742012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fang L, Moore XL, Gao XM, Dart AM, Lim YL
and Du XJ: Down-regulation of mitofusin-2 expression in cardiac
hypertrophy in vitro and in vivo. Life Sci. 80:2154–2160. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen KH, Guo X, Ma D, Guo Y, Li Q, Yang D,
Li P, Qiu X, Wen S, Xiao RP and Tang J: Dysregulation of HSG
triggers vascular proliferative disorders. Nat Cell Biol.
6:872–883. 2004. View
Article : Google Scholar : PubMed/NCBI
|