Introduction
The matrix metalloproteinases (MMPs) are a family of
peptidase enzymes responsible for the degradation of extracellular
matrix components, including collagen, gelatin, fibronectin,
laminin and proteoglycan (1).
Currently, more than 20 different members of the MMP family have
been identified. Based on sequence homology and substrate
specificity, MMPs are classified into collagenases, stromelysins,
gelatinases, membrane-type and other MMP sub-families (2). The activity of MMPs is known to be
associated with cardiovascular diseases, including atherosclerosis
(3), restenosis following
angioplasty (4), dilated
cardiomyopathy (5,6) and myocardial repair following
infarction (7).
MMP-9 is classified into the gelatinases sub-family.
MMP-9 cleaves gelatin, collagens IV, V and XIV, aggrecan, elastin,
entactin and vitronectin. Endothelial cells, smooth muscle cells,
fibroblasts and infiltrating inflammatory cells are able to produce
MMP-9. It is generally recognized that MMP-9 is involved in smooth
muscle cell migration in vivo and in vitro (8–10).
Accordingly, MMP-9 expression is markedly enhanced in vascular
stenosis following experimental balloon dilation and arterial
restenosis following angioplasty (11). The transcription of MMP-9 genes is
differentially activated by phorbol esters, lipopolysaccharides
(LPS) and staphylococcal enterotoxin B (SEB). Previous studies
demonstrated that the expression of MMP-9 is upregulated in the
liver, spleen and kidney following bacterial LPS administration
(12,13), and that selective deletion of the
MMP-9 gene protects against LPS-induced mortality (13). MMP-9 expression is also induced by
LPS in human aortic smooth muscle cells (HASMCs) (14). However, how LPS regulates MMP-9
expression remains to be determined. Since HASMCs express TLR4 and
MMP-9, and LPS upregulates MMP-9 expression in HASMCs, we
hypothesized that LPS upregulates MMP-9 expression in HASMCs
through the TLR4 pathway. In the present study, we demonstrated
that LPS upregulates MMP-9 expression in HASMCs through TLR4-NF-κB
signaling.
Materials and methods
Materials
HASMCs and medium 231 were obtained from Cascade
Biologics (Portland, OR, USA). LPS from Escherichia coli and
pyrrolidine dithiocarbamate (PDTC) were purchased from Sigma (St.
Louis, MO, USA). Neutralizing anti-TLR4 (HTA125) antibodies were
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
The anti-human MMP-9 polyclonal antibody used in the western blot
analysis was from Santa Cruz Biotechnology, Inc.
Cell culture
HASMCs were cultured in medium 231 supplemented with
5% SMGS [5% fetal bovine serum, recombinant human basic fibroblast
growth factor, recombinant human epidermal growth and insulin
(Cascade Biologics)]. After cells in the medium reached 80%
confluence, HASMCs were dislodged from the surface of the flask
using trypsin/EDTA solution, washed and resuspended in supplemented
medium 231. The cells were subcultured at ratios of 1:3. Subsequent
experiments were conducted using passages 5–7.
RT-PCR
Total RNA was extracted from resting or chemical
agent-stimulated HASMCs using TRIzol® reagent according
to the manufacturer's instructions (Invitrogen Life Technologies,
Carlsbad, CA, USA). Total RNA (1 μg) was used as a template to make
the cDNA using a reverse trancription kit (BioDev Scientific &
Technical Co., Ltd., China). PCR amplification was performed with
Taq polymerase for 32 cycles of 45 sec at 95°C, 30 sec at
62°C and 1 min at 72°C (for MMP-9 and GAPDH). The primer sets used
to amplify MMP-9 were 5′-gacatcgtcatccagtttgg-3′ (sense) and
5′-tggccttggaagatga atgg-3′ (antisense), yielding a 398-bp
fragment. The primer sets used to amplify GAPDH were
5′-cagggctgcttttaactctg-3′ (sense) and 5′-gaagatggtgatgggatttc-3′
(antisense), yielding a 175-bp fragment. The MMP-9 and GAPDH RT-PCR
products were confirmed by electrophoresis on a 1.5% agarose gel
and sequencing analysis.
Western blot analysis
HASMCs were washed with ice-cold phosphate-buffered
saline (PBS), and lysed for 20 min on ice with lysis buffer
containing 20 mM Tris-HCl (pH 8.0), 137 mM NaCl, 10% glycerol, 1%
NP-40, 2 mM EDTA, 5 mM DTT and 10 mM PMSF. Following lysis, the
lysates were centrifuged for 4 min at 12,000 rpm, and the
supernatants were collected in fresh tubes kept on ice. Protein
concentrations in each sample were determined using a bicinchoninic
acid (BCA) assay. Total protein (100 μg) was mixed with loading
buffer with the anionic denaturing detergent sodium dodecyl sulfate
(SDS), boiled at 100°C for 5 min, and then resolved by 10% SDS
polyacrylamide gel electrophoresis. The proteins were transferred
onto a polyvinylidene fluoride membrane. After blocking the
membrane in Tris-buffered saline Tween-20 (TBST) containing non-fat
milk for 1 h at 4°C under agitation, the membrane was washed three
times in TBST and incubated for 2 h with anti-human MMP-9 antibody
(1:200; Santa Cruz Biotechnology, Inc.,) or GAPDH monoclonal
antibody (1:200; Santa Cruz Biotechnology, Inc.,). After washing
three times in TBST, membranes were incubated with HRP-conjugated
mouse anti-human IgG (1:100; Santa Cruz Biotechnology, Inc.,) at
room temperature for 1 h and then washed three times with TBST. IgG
bands were detected using a streptavidin amplification reagent
according to the manufacturer's instructions, and visualized using
a ProtoBlot®II AP system (Promega Corporation, Madison,
WI, USA).
Electrophoretic mobility shift assay
Nuclear extract was prepared according to the
Nuclear Extract kit manufacturer's instructions (Active Motif,
Carlsbad, CA, USA). The oligonucleotides containing the NF-κB
binding site (5′-agttgaggg gactttcccaggc-3′) were radiolabeled with
γ-32P-ATP using a T-4 polynucleotide kinase (Promega
Corporation). Binding reactions were performed with 10 mg of
nuclear protein in 10 mM Tris, 1 mM DTT, 1 mM EDTA, 5% glycerol,
0.1% Triton X-100, 1 mg poly (dIdC), 5 mg BSA and
γ-32P-labeled oligonucleotide. Subsequently, the
reaction mixture was separated on 5% non-denaturing polyacrylamide
gels for separation from the unbound oligonucleotides according to
the manufacturer's instructions (Promega Corporation).
Statistical analysis
Quantitative data were presented as the mean ±
standard deviation (SD). For comparison between multiple groups,
the data were analyzed by ANOVA and the Student-Newman-Keuls
post-hoc analysis. P<0.05 was considered statistically
significant. SPSS 11.5 (SPSS Inc., Chicago, IL, USA) was used for
statistical analysis.
Results
LPS induced MMP-9 expression in HASMCs in
a TLR4- dependent manner
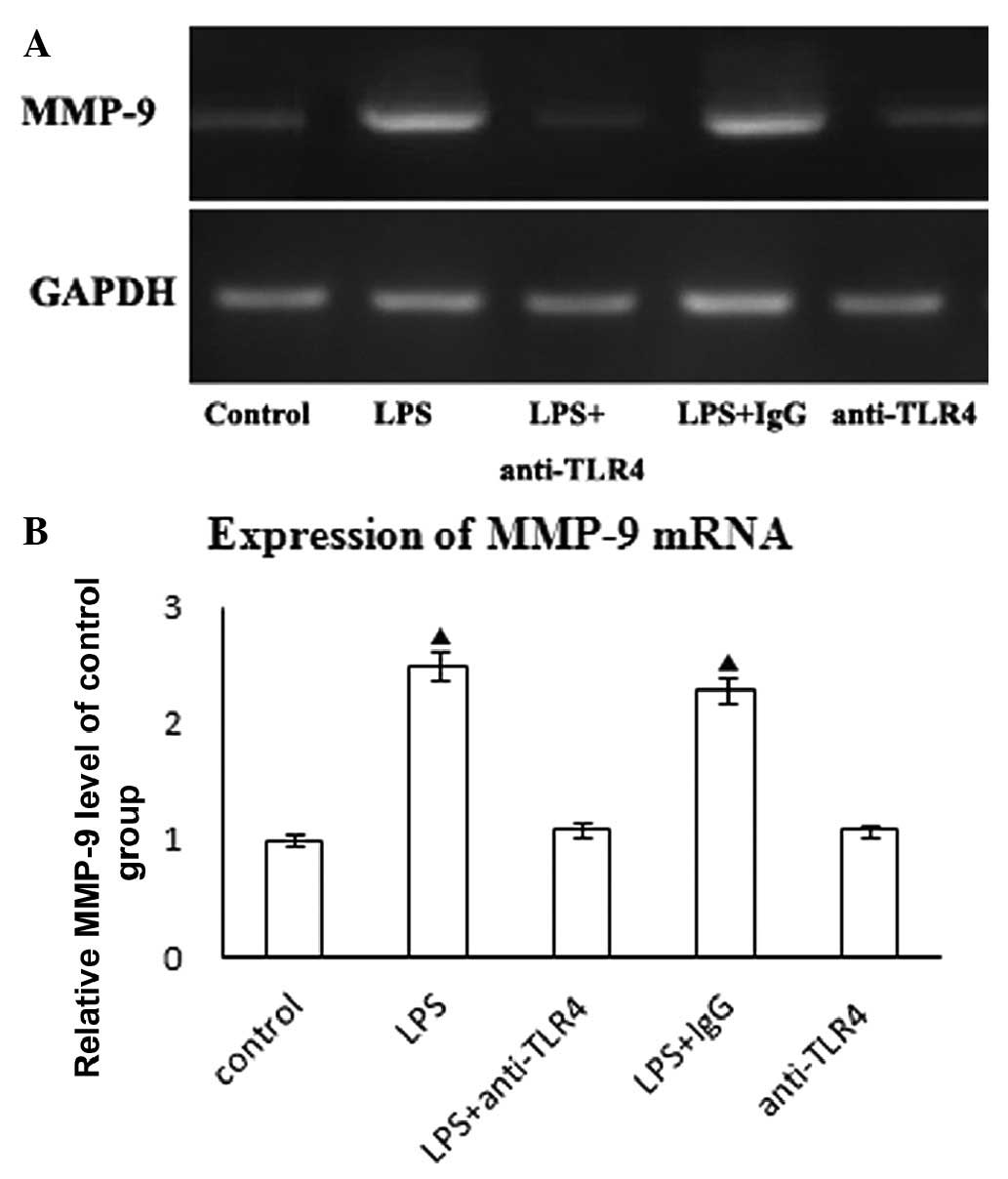
Unstimulated HASMCs expressed low levels of MMP-9
mRNA, however, the expression was upregulated upon treatment with
LPS at 10 ng/ml for 24 h (Fig.
1A). To confirm that the purified LPS signals through TLR4, we
treated HASMCs with anti-TLR4 neutralizing antibodies or control
IgG (10 μg/ml) for 2 h prior to the addition of LPS. Our results
demonstrate that anti-TLR4 neutralizing antibody pre-treatment
reduces LPS-mediated MMP-9 mRNA expression (Fig. 1B). However, the addition of control
IgG did not affect LPS-mediated MMP-9 mRNA expression. Furthermore,
neutralizing anti-TLR4 did not affect on MMP-9 mRNA expression in
HASMCs (Fig. 1B).
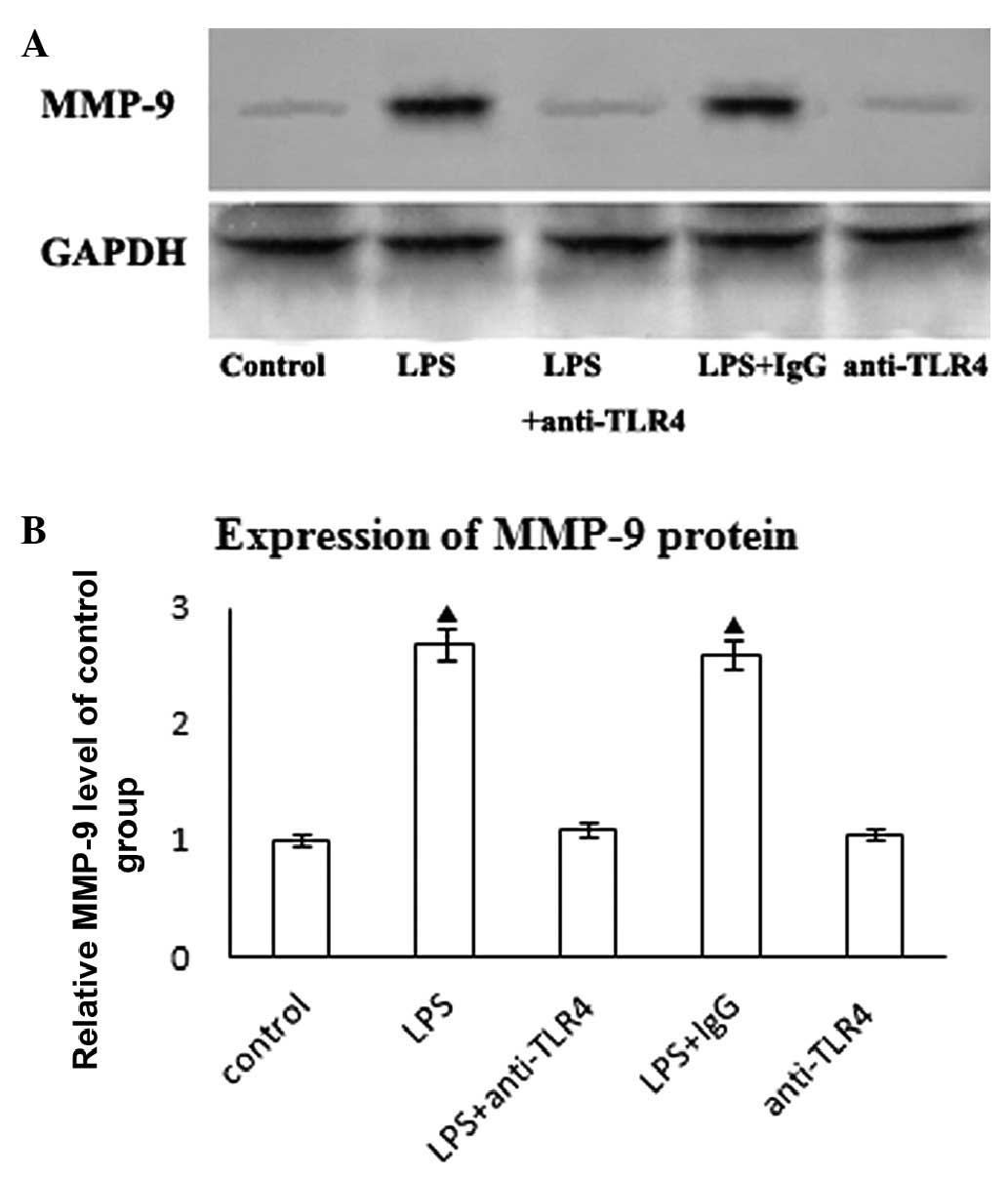
We examined the expression of MMP-9 protein in
response to LPS. Untreated HASMCs exhibited a low expression of
MMP-9 protein that was markedly upregulated in response to 10 ng/ml
LPS stimulation for 24 h (Fig.
2A). Anti-TLR4 neutralizing antibody pre-treatment
reducedLPS-mediated MMP-9 protein expression (Fig. 2B). However, the addition of control
IgG did not affect LPS-mediated MMP-9 protein expression.
Furthermore, neutralizing anti-TLR4 did not affect MMP-9 protein
expression in HASMCs (Fig.
2B).
Taken together, these results demonstrate that LPS
induces MMP-9 expression and production in HASMCs in a
TLR4-dependent manner.
TLR4 mediates LPS-induced activation of
NF-κB in HASMCs
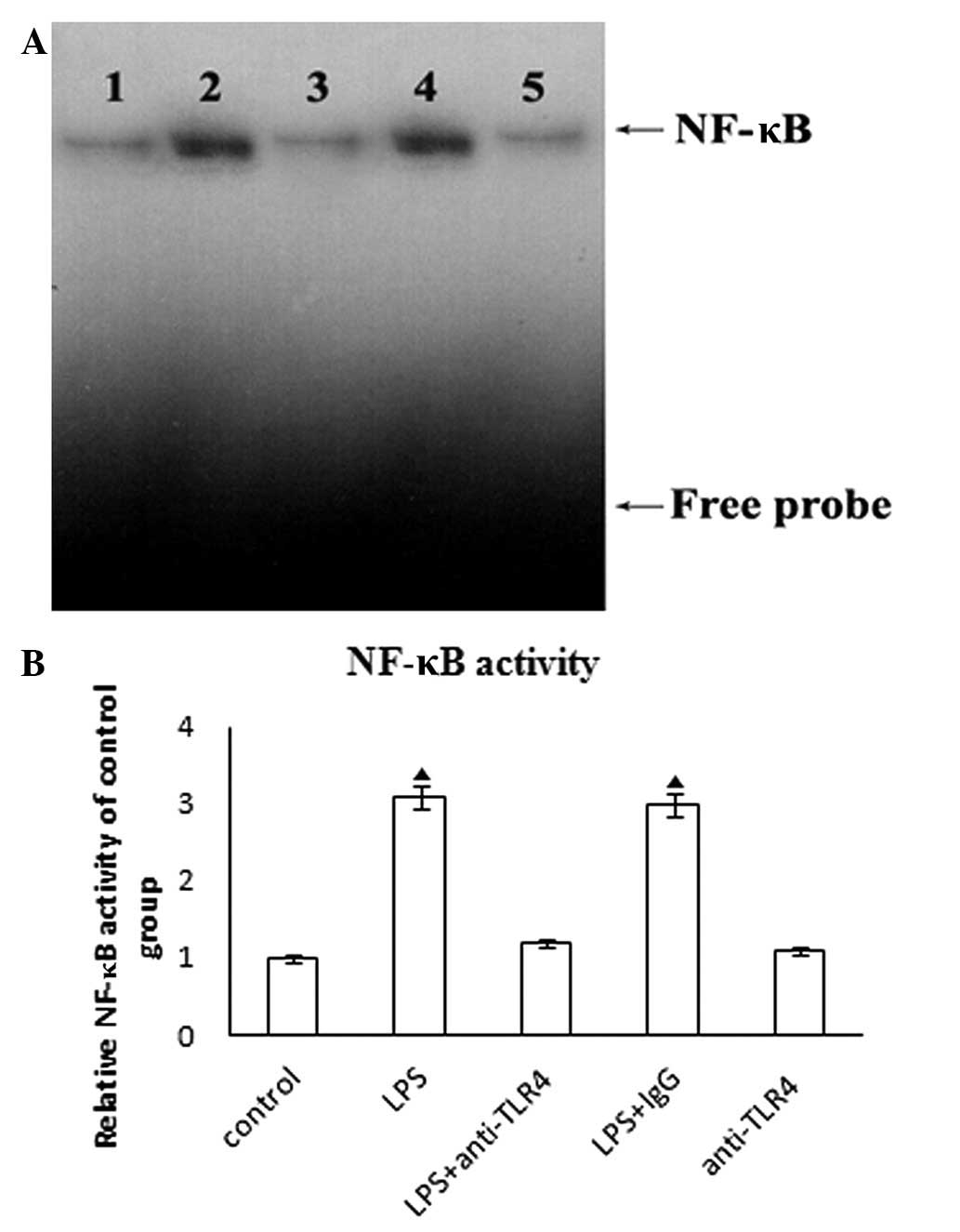
Nuclear extracts of HASMCs treated with LPS
demonstrated a significant increase in NF-κB activation. The
anti-TLR4 neutralizing antibody pre-treatment blocked this effect.
The addition of control IgG did not affect LPS-induced NF-κB
activation (Fig. 3A). Furthermore,
neutralizing anti-TLR4 had no effect on NF-κB activation in HASMCs
(Fig. 3B). These results
demonstrate that LPS induces NF-κB activation in a TLR4-dependent
manner in HASMCs.
LPS-induced MMP-9 expression is dependent
on NF-κB activation
LPS potently induced NF-κB activation in HASMCs, and
this effect was blocked by neutralizing anti-TLR4. We investigated
whether the changes in NF-κB activation correlated with the
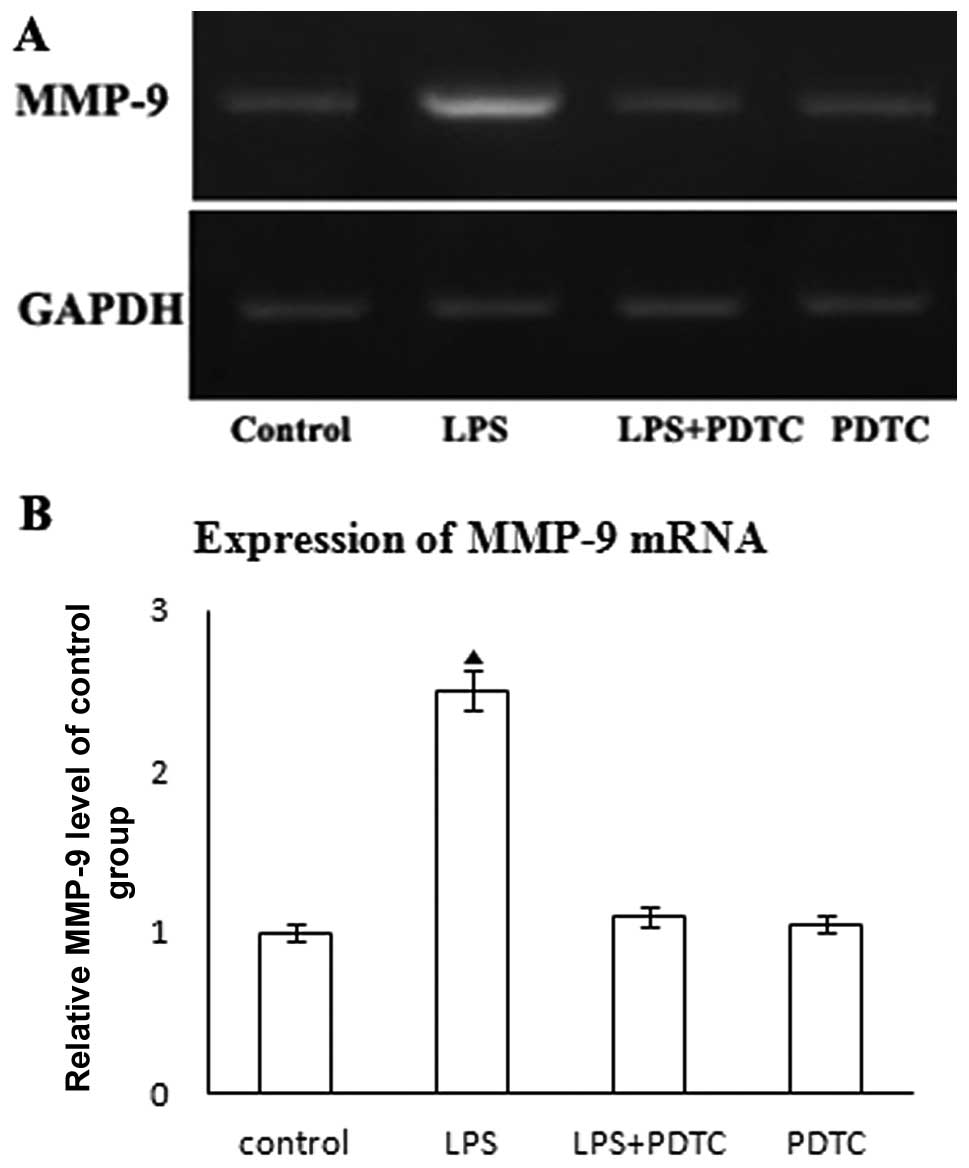
expression of MMP-9. PDTC has been demonstrated to be a potent
antioxidant and inhibitor of NF-κB in in vivo and in
vitro studies (15). In our
study, NF-κB activation was targeted by PDTC (100 μM in PBS for 1
h) (data not shown). To determine the role of NF-κB in LPS-mediated
MMP-9 expression, HASMCs were treated with PDTC (100 μM in PBS for
1 h) prior to the addition of LPS. Treatment with PDTC attenuated
LPS-induced MMP-9 mRNA expression (Fig. 4A and B). Our results indicate that
LPS induces MMP-9 upregulation in an NF-κB-dependent manner.
Discussion
Endotoxin is important in the development and
progression of atherosclerosis via the induction of TLR4-dependent
proinflammatory cytokines, chemokines and adhesion molecule
expression (16). MMP-9 expression
is markedly enhanced in vascular stenosis following experimental
balloon dilation and arterial restenosis following angioplasty
(11). LPS has been demonstrated
to stimulate MMP-9 expression in smooth muscle cells (14). However, how LPS regulates MMP-9
expression in HASMCs remains to be determined. In the present
study, we demonstrated that LPS upregulates MMP-9 expression in
HASMCs through TLR4/NF-κB signaling.
MMP-9 is classified into the gelatinase sub-group of
MMPs. Previous studies have demonstrated that the expression of
MMP-9 is upregulated in the liver, spleen and kidney following
bacterial LPS administration (12,13).
In the present study, we demonstrated that LPS upregulates MMP-9
expression in HASMCs. These results are in accordance with results
of other studies (14). HASMCs
expressed MMP-9 at low levels under basal conditions. LPS
upregulated MMP-9 expression, and anti-TLR4 neutralizing antibody
pre-treatment abolished LPS-mediated MMP-9 mRNA and protein
expression. These results demonstrate that LPS induces MMP-9
expression and production in a TLR4-dependent manner in HASMCs.
Findings of another study suggested that LPS induces the expression
of genes encoding the elastin-degrading enzyme via TLR4 in human
cervical smooth muscle cells (17). These proteinases allow SMCs to
migrate to the site of inflammation or injury in response to
chemokines (8). A clinical
observation suggested that MMP-9 promoter is positively correlated
with more rapid intima-media thickening (IMT) and constrictive
remodeling (CR) progression (18).
These studies lead to a pathogenic role for MMP-9 in vascular
occlusive diseases.
NF-κB was first identified in the B-lymphocyte
nucleus as a transcription factor that binds to an enhancer of the
immunoglobulin κ light chain gene. NF-κB is able to mediate gene
transcription by binding to specific sequences in the promoter
regions of its target genes. LPS has been shown to induce
activation of NF-κB in vivo (19). Our results also suggest that LPS
induces NF-κB activation in a TLR4-dependent manner in HASMCs. In
our study, the nuclear extracts of HASMCs treated with LPS revealed
significant increases in NF-κB activation, and anti-TLR4
neutralizing antibody pre-treatment blocked this effect. Addition
of the control IgG did not affect LPS-induced NF-κB activation. To
determine the role of NF-κB in LPS-mediated MMP-9 expression,
HASMCs were treated with PDTC prior to the addition of LPS. The
results demonstrated that LPS stimulated MMP-9 expression in
HASMCs, and that inhibition of NF-κB blocks LPS-mediated MMP-9
upregulation, indicating that MMP-9 is an NF-κB-responsive gene.
Bea et al found that inhibition of the activation of
proinflammatory transcription factors might reduce the synthesis of
MMP-9 in vivo (20). These
results suggest that LPS modulates MMP-9 expression through NF-κB
activation. However, other studies have suggested that there are
other pathways involved in LPS-induced MMP-9 expression (21,22).
One of the explanations for this may be differences in species.
In conclusion, findings of the present study have
shown that LPS upregulates MMP-9 expression in HASMCs through
TLR4/NF-κB signaling.
Acknowledgements
This study was supported by the Shanghai Rising-Star
Program (08QA1404100) and the National Natural Science Foundation
of China (30971265).
References
|
1
|
Nagase H: Activation mechanisms of matrix
metalloproteinases. Biol Chem. 378:151–160. 1997.PubMed/NCBI
|
|
2
|
Creemers EE, Cleutjens JP, Smits JF and
Daemen MJ: Matrix metalloproteinase inhibition after myocardial
infarction: a new approach to prevent heart failure? Circ Res.
89:201–210. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lancelot E, Amirbekian V, Brigger I,
Raynaud JS, Ballet S, David C, Rousseaux O, Le Greneur S, Port M,
Lijnen HR, Bruneval P, Michel JB, Ouimet T, Roques B, Amirbekian S,
Hyafil F, Vucic E, Aguinaldo JG, Corot C and Fayad ZA: Evaluation
of matrix metalloproteinases in atherosclerosis using a novel
noninvasive imaging approach. Arterioscler Thromb Vasc Biol.
28:425–432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Furman C, Luo Z, Walsh K, Duverger N,
Copin C, Fruchart JC and Rouis M: Systemic tissue inhibitor of
metalloproteinase-1 gene delivery reduces neointimal hyperplasia in
balloon-injured rat carotid artery. FEBS Lett. 531:122–126. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fedak PW, Moravec CS, McCarthy PM,
Altamentova SM, Wong AP, Skrtic M, Verma S, Weisel RD and Li RK:
Altered expression of disintegrin metalloproteinases and their
inhibitor in human dilated cardiomyopathy. Circulation.
113:238–245. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tziakas DN, Chalikias GK, Papaioakeim M,
Hatzinikolaou EI, Stakos DA, Tentes IK, Papanas N, Kortsaris A,
Maltezos E and Hatseras DI: Comparison of levels of matrix
metalloproteinase-2 and -3 in patients with ischemic cardiomyopathy
versus nonischemic cardiomyopathy. Am J Cardiol. 96:1449–1451.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun Y, Zhang JQ, Zhang J and Lamparter S:
Cardiac remodeling by fibrous tissue after infarction in rats. J
Lab Clin Med. 135:316–323. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ogawa M, Suzuki J, Hishikari K, Takayama
K, Tanaka H and Isobe M: Clarithromycin attenuates acute and
chronic rejection via matrix metalloproteinase suppression in
murine cardiac transplantation. J Am Coll Cardiol. 51:1977–1985.
2008. View Article : Google Scholar
|
|
9
|
Lee SO, Chang YC, Whang K, Kim CH and Lee
IS: Role of NAD(P)H:quinone oxidoreductase 1 on tumor necrosis
factor-alpha-induced migration of human vascular smooth muscle
cells. Cardiovasc Res. 76:331–339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Otero-Viñas M, Llorente-Cortés V, Peña E,
Padró T and Badimon L: Aggregated low density lipoproteins decrease
metalloproteinase-9 expression and activity in human coronary
smooth muscle cells. Atherosclerosis. 194:326–333. 2007.PubMed/NCBI
|
|
11
|
Cho A and Reidy MA: Matrix
metalloproteinase-9 is necessary for the regulation of smooth
muscle cell replication and migration after arterial injury. Circ
Res. 91:845–851. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abe K, Ikeda T, Wake K, Sato T, Sato T and
Inoue H: Glycyrrhizin prevents
lipopolysaccharide/D-galactosamine-induced liver injury through
down-regulation of matrix metalloproteinase-9 in mice. J Pharm
Pharmacol. 60:91–97. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pagenstecher A, Stalder AK and Kincaid CL:
Regulation of matrix metalloproteinases and their inhibitor genes
in lipopolysaccharide-induced endotoxemia in mice. Am J Pathol.
157:197–210. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin SJ, Lee IT, Chen YH, Lin FY, Sheu LM,
Ku HH, Shiao MS, Chen JW and Chen YL: Salvianolic acid B attenuates
MMP-2 and MMP-9 expression in vivo in apolipoprotein-E-deficient
mouse aorta and in vitro in LPS-treated human aortic smooth muscle
cells. J Cell Biochem. 100:372–384. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Beswick RA, Zhang H, Marable D, Catravas
JD, Hill WD and Webb RC: Long-term antioxidant administration
attenuates mineralocorticoid hypertension and renal inflammatory
response. Hypertension. 37:781–786. 2001. View Article : Google Scholar
|
|
16
|
Li Hongli and Sun Baogui: Toll-like
receptor 4 in atherosclerosis. J Cell Mol Med. 11:88–95. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Watari M, Watari H, Nachamkin I and
Strauss JF: Lipopolysaccharide induces expression of genes encoding
pro-inflammatory cytokines and the elastin-degrading enzyme,
cathepsin S, in human cervical smooth-muscle cells. J Soc Gynecol
Invest. 7:190–198. 2000. View Article : Google Scholar
|
|
18
|
Fiotti N, Altamura N, Fisicaro M, Carraro
N, Adovasio R, Sarra VM, Uxa L, Guarnieri G, Baxter BT and
Giansante C: MMP-9 microsatellite polymorphism: association with
the progression of intima-media thickening and constrictive
remodeling of carotid atherosclerotic plaques. Atherosclerosis.
182:287–292. 2005. View Article : Google Scholar
|
|
19
|
Tsai CS, Lin FY, Chen YH, Yang TL, Wang
HJ, Huang GS, Lin CY, Tsai YT, Lin SJ and Li CY: Cilostazol
attenuates MCP-1 and MMP-9 expression in vivo in LPS-administrated
balloon-injured rabbit aorta and in vitro in LPS-treated monocytic
THP-1 cells. J Cell Biochem. 103:54–66. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bea F, Kreuzer J, Preusch M, Schaab S,
Isermann B, Rosenfeld ME, Katus H and Blessing E: Melagatran
reduces advanced atherosclerotic lesion size and may promote plaque
stability in apolipoprotein E-deficient mice. Arterioscler Thromb
Vasc Biol. 26:2787–2792. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee SY, Kim HJ, Lee WJ, Joo SH, Jeon SJ,
Kim JW, Kim HS, Han SH, Lee J, Park SH, Cheong JH, Kim WK, Ko KH
and Shin CY: Differential regulation of matrix metalloproteinase-9
and tissue plasminogen activator activity by the cyclic-AMP system
in lipopolysaccharide-stimulated rat primary astrocytes. Neurochem
Res. 33:2324–2334. 2008. View Article : Google Scholar
|
|
22
|
Marcet-Palacios M, Ulanova M, Duta F,
Puttagunta L, Munoz S, Gibbings D, Radomski M, Cameron L, Mayers I
and Befus AD: The transcription factor Wilms tumor 1 regulates
matrix metalloproteinase-9 through a nitric oxide-mediated pathway.
J Immunol. 179:256–265. 2007. View Article : Google Scholar : PubMed/NCBI
|