Introduction
Acute pancreatitis (AP) is an inflammatory disorder
of the exocrine pancreas, which carries considerable morbidity and
mortality, and the pathophysiology of which remains unknown
(1,2). Parenchymal cell death is a major
complication of AP (3). Several
recent studies have indicated that pancreatic acinar cell death
occurs through both necrosis and apoptosis, and the severity of AP
directly correlates with the extent of necrosis and inversely with
that of apoptosis (3–5). Apoptosis is the process of programmed
cell death, during which no inflammatory reaction occurs. By
contrast, necrosis is a form of premature cell death, which is
capable of leading to the release of a variety of inflammatory
agents. The reported elevation of pancreatic acinar cell apoptosis
may indicate a beneficial response following the onset of
pancreatitis (6–8).
There are two main apoptotic pathways. The extrinsic
(death receptors) and the intrinsic (mitochondrial) pathways are
activated by caspase-8 and caspase-9, respectively. Activated
caspase-8 and -9 subsequently cleave and activate the executioner
caspases, such as caspase-3 and caspase-7, which then cleave
intracellular substrates, resulting in apoptosis (9–11).
By contrast, caspase activation may be tightly controlled by
endogenous inhibitors, such as the inhibitor of apoptosis proteins
(IAP), a family of structurally related proteins that negatively
regulate caspase activation. The X-linked IAP (XIAP) is the most
potent of the eight mammalian IAPs and blocks cell apoptosis
through inhibiting the activation of caspases-3, -7 and -9
(12,13).
The 78-kDa glucose-regulated protein (GRP78), also
known as BiP or HSPA5, is not only a major molecular chaperone but
also an anti-apoptotic molecule of the cell death signaling pathway
(14–16). Previous studies have indicated that
GRP78 plays an anti-apoptotic role, possibly through interaction
with XIAP (17,18). In addition, GRP78 exhibits
anti-apoptotic properties through interference with caspase
activation and release (14,19).
However, the pathological role of GRP78 and XIAP in AP remains
unclear.
It is unknown whether GRP78 and XIAP participate in
the pathogenesis of AP and whether GRP78 and XIAP correlate with
the regulation of cell death during AP. The aim of the present
study was to investigate the expression of GRP78 and XIAP and their
potential roles in the regulation of apoptosis during CAP in
rats.
Materials and methods
Animal experiments
Male Wistar rats (Experimental Animal Center of
Sichuan University, Chengdu, China), weighing 200 to 220 g, were
used in this study. All the animals were maintained at 23°C on a
12-h light/dark cycle and starved for 12 h prior to
experimentation, but were allowed free access to water. All the
animal experiments were conducted according to the guidelines of
the local animal use and care committees and executed according to
the National Animal Welfare Law of China. The rats were randomly
divided into 2 groups: the cerulein-treated AP group (CAP) and the
saline-treated control group. The CAP group was further divided
into 5 subgroups depending on the time of sampling: 1, 3, 6, 12 or
24 h (n=6). Acute pancreatitis was induced in the CAP group of rats
by injecting cerulein (Sigma, St. Louis, MO, USA) at a dose of 50
μg/kg intraperitoneally four times at 1-h intervals. Normal saline
was substituted for cerulein with the same volume for the controls.
After the last injection, rats were sacrificed at 1, 3, 6, 12 and
24 h under 1% pentobarbital anesthesia (50 mg/kg of body weight).
Fresh pancreatic tissue was removed, weighed immediately for the
pancreatic tissue edema assay, rinsed in TRIzol (Gibco, Carlsbad,
CA, USA) for subsequent RNA isolation, frozen at −80°C for western
blot analysis, and fixed with 10% formaldehyde prior to paraffin
sectioning for histological examination or
terminal-deoxynucleotidyl-transferase-mediated dUTP nick-end
labeling (TUNEL) assay. Inferior vena cava blood was collected,
centrifuged at 4°C, and the serum was stored at −80°C for enzyme
linked immunosorbent assay (ELISA).
Serum amylase and lipase
determination
Serum amylase and lipase activity were determined by
commercially available ELISA kits (R&D, Minneapolis, MN, USA),
according to the manufacturer’s instructions.
Pancreatic tissue edema assay
For the evaluation of pancreatic tissue edema, the
pancreas was removed and immediately weighed. It was then dried in
an oven at 80°C for 72 h, and reweighed. The extent of pancreatic
edema was determined by measuring tissue water content (wet weight
- dry weight/wet weight × 100 = percentage tissue water
content).
Histological examination
For routine histological examination, 4-μm sections
of 10% formalin-fixed, paraffin-embedded tissue were prepared and
stained with hematoxylin and eosin (HE). All microscopic sections
were evaluated in a blind fashion.
TUNEL assay
In the pancreatic tissue, apoptosis was detected by
use of the in situ cell death detection kit (Roche Applied
Science, Mannheim, Germany), according to the manufacturer’s
instructions. Briefly, tissue was fixed in 10% buffered
formaldehyde, embedded in paraffin, and 4-μm sections were adhered
to glass slides. After dewaxing and rehydration, the sections were
incubated with TUNEL reaction mixture at 37°C for 1 h. Finally, the
sections were analyzed under a fluorescence microscope (Olympus,
Tokyo, Japan) at 450–500 nm. Images of these tissues were obtained
using an image acquisition system (Olympus DD70 BX51).
TUNEL-positive cells displayed brilliant green fluorescence. For
each test, negative controls were included. Apoptotic index (AI)
was determined as the percentage of TUNEL positive cells in 10
randomly selected high-power fields by averaging 10 counts per
tissue section.
Quantitative real-time reverse
transcriptase PCR
Fresh pancreatic tissue (50–80 mg) was collected per
rat, and total RNA was isolated using TRIzol (Gibco). Total RNA (5
μg) was reverse transcribed and 1 μg of the RT product was
subjected to PCR in the presence of specific primers. The sequences
of the primers were as follows: (GRP78: 5′-GAA
ACTGCCGAGGCGTAT-3′/5′-ATGTTCTTCTCTCCCTCT CTCTTA-3′;
Caspase-3: 5′-CGGACCTGTGGACCTGAA
A-3′/5′-GGGTGCGGTAGAGTAAGC-3′; Caspase-7: 5′-TCT
ATGTGCCCCGTCAGTA-3′/5′-ACATCCATACCTGTCGC TTT-3′; Caspase-9:
5′-ACGACCTGACTGCTAAGAAA-3′/5′-AGCCATGAGAGAGGATGAC-3′; XIAP:
5′-TGTGAG TGCTCAGAAAGATAAT-3′/5′-TGCTTCTGCACACTG TTTACA-3′;
β-actin:
5′-CGTGAAAAGATGACCCAGAT-3′/5′-ACCCTCATAGATGGGCACA-3′). Conditions
for all PCRs were optimized on an iCycler IQ (Bio-Rad, Hercules,
CA, USA) system for a 30-μl reaction using the following 40 cycle
program: 94°C for 20 sec, 53°C for 30 sec, and 72°C for 30 sec. All
samples were amplified simultaneously in triplicate in one
assay-run. β-actin was included in each reaction as an internal
standard and relative quantitative gene expression was calculated
using the 2−ΔΔCt method as described previously
(20).
Western blot assay
Total proteins were prepared from pancreatic tissue
using a total protein extraction kit (KeyGen Biotech., Co., Ltd.,
Nanjing, China) and the concentrations were determined using a
bicinchoninic acid protein assay kit (Pierce, Rockford, IL, USA).
Each 20-μg aliquot of total protein was loaded onto a 12% sodium
dodecyl sulfate-polyacrylamide gel for electrophoresis, and then
transferred onto polyvinylidene difluoride membranes (Millipore,
Billerica, MA, USA). Following complete protein transfer, the
membranes were blocked with 5% milk powder solution for 2 h and
incubated with primary antibodies overnight. The primary antibodies
used in this study included polyclonal GRP78 (Abcam, Cambridge, UK)
and monoclonal XIAP (Cell Signaling, Beverly, MA, USA) in a 1:1,000
dilution individually. For internal reference, a monoclonal rabbit
anti-rat β-actin antibody (1:1,000 dilution; Cell Signaling) was
used. After washing the membranes, the goat polyclonal anti-rabbit
immunoglobulin G secondary antibody (Cell Signaling) conjugated to
horseradish peroxidase was applied in a 1:5,000 dilution and
incubated for 2 h at room temperature. Finally, antibody binding
was visualized using the enhanced chemiluminescence system
(Pierce), and the semi-quantitative grayscale intensity was
generated with Multi Gauge V3.0 software (Fujifilm, Tokyo,
Japan).
Statistical analysis
The experimental data are expressed as the means ±
SE. The differences between 2 groups were compared by the
non-paired Student t-test. Pearson correlation coefficient was
calculated where indicated. P<0.05 was considered to indicate a
statistically significant difference. All tests were performed
using the statistical package SPSS software 13.0 (SPSS, Chicago,
IL, USA).
Results
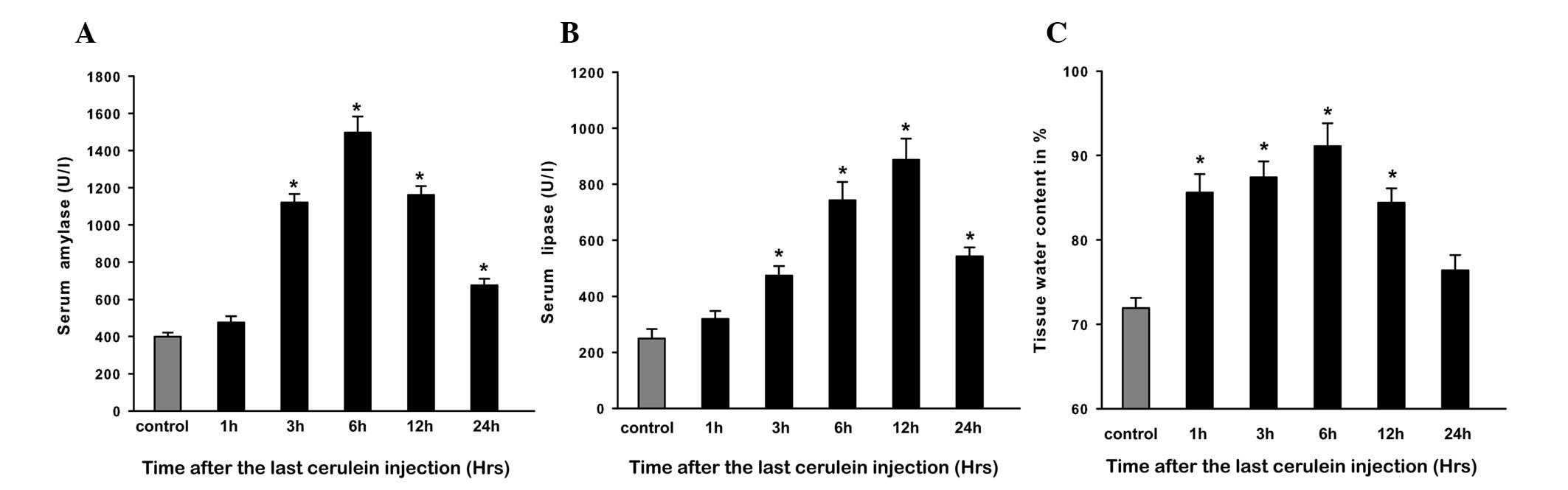
Evaluation of CAP
In this study, intraperitoneal injections of 50
μg/kg cerulein led to the induction of AP, and the severity of CAP
was assessed by serum amylase and lipase levels and pancreatic
edema. Compared with control rats, amylase and lipase levels in CAP
rats were significantly increased within 3 h and peaked at 6 and 12
h, respectively (P<0.05), then a trend of return to baseline
levels was observed at 24 h (Fig. 1A
and B). Injections of cerulein induced prominent pancreatic
edema as assessed by tissue water content, which showed that tissue
water content was significantly increased at 1 h and peaked at 6 h;
thereafter it gradually decreased and returned to baseline at 24 h
(Fig. 1C).
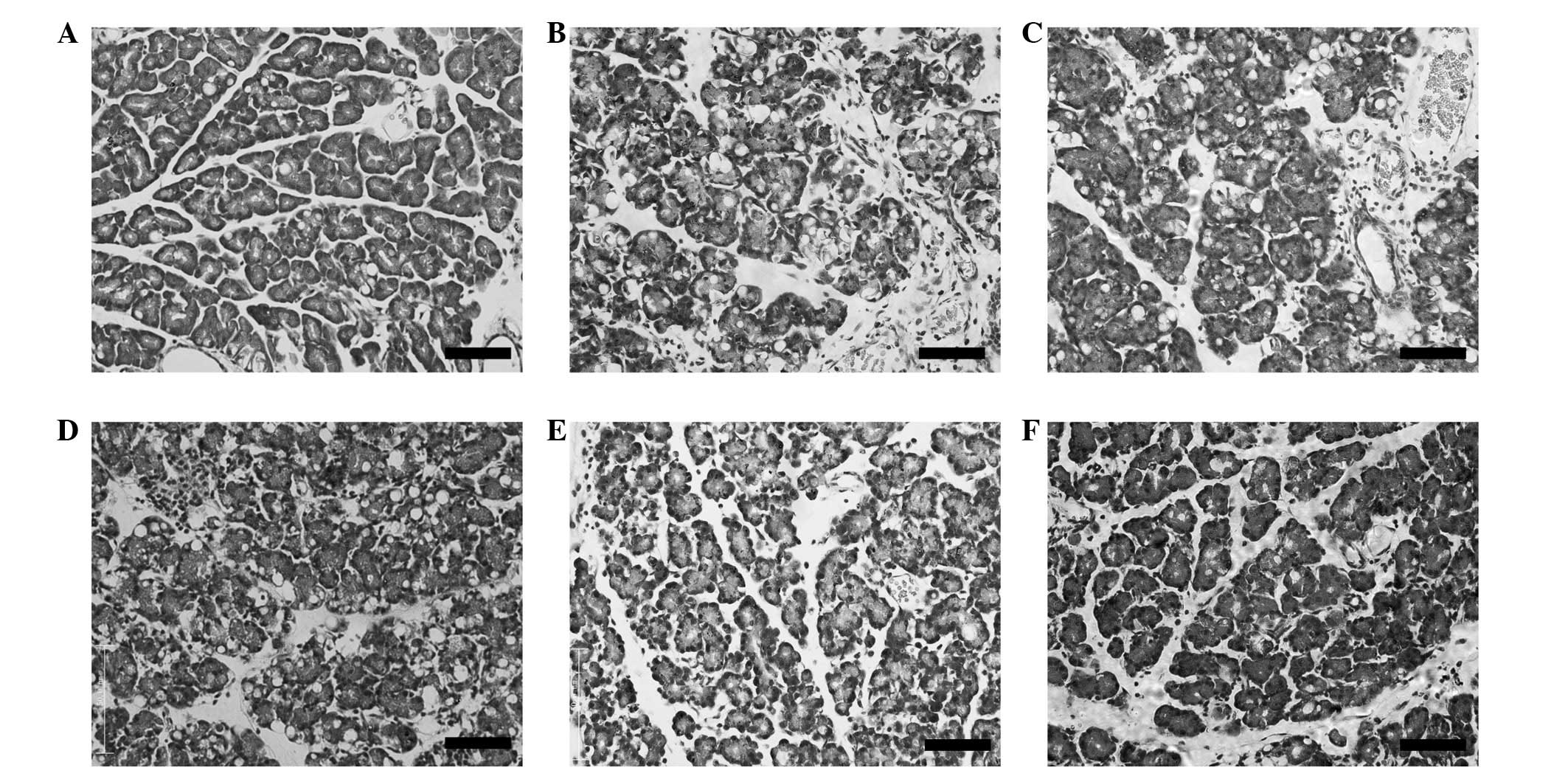
Histological findings
Rat pancreatic tissues were stained with HE for
morphological examination. None of the control tissues showed
characteristics of AP at any time point (Fig. 2A). CAP tissues showed evidence of
interstitial edema, cellular vacuolation and inflammatory cell
infiltration. The most severe interstitial edema and cellular
vacuolation were observed at 6 h (Fig.
2D), whereas inflammatory cell infiltration in the interstitial
space was most predominant at 12 h (Fig. 2E). Some recovery was apparent by 24
h after cerulein treatment, as pancreatic tissue appeared more
similar to the control (Fig.
2F).
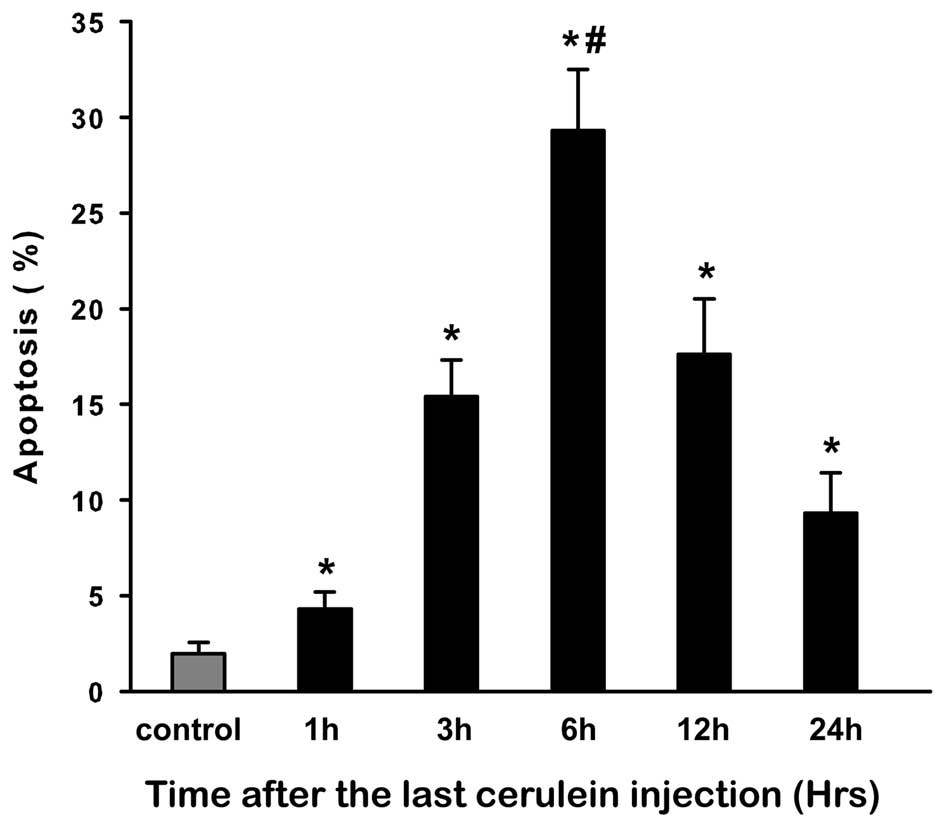
Apoptosis assay during CAP
Apoptosis in pancreatic acinar cells was determined
by TUNEL assay (Fig. 3). The
results of TUNEL assay showed that cerulein induced a
time-dependent increase in apoptosis in comparison with control
rats. We observed that AI was markedly increased at 3 h and peaked
at 6 h (P<0.05). Although this increase began to decrease after
6 h, it was significantly increased at 24 h in comparison with the
control (P<0.05; Fig. 4).
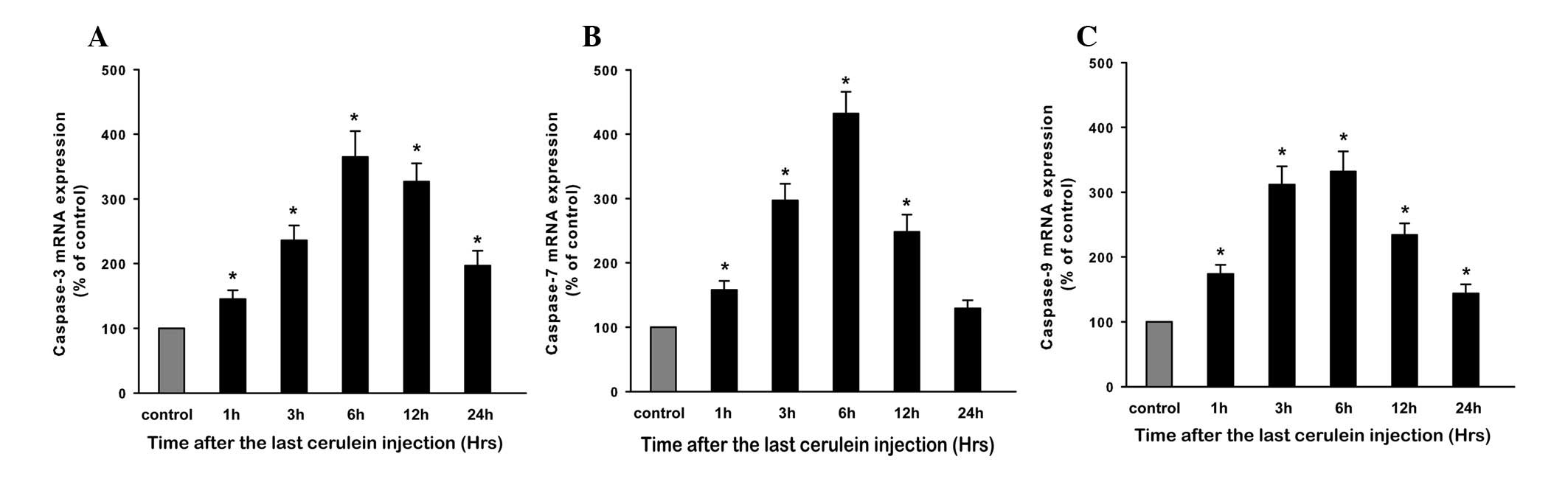
Expression of caspase genes during
CAP
In order to further understand the apoptotic
signaling pathway, we determined the initiator caspase-9 and the
effector caspase-3 and -7 mRNA expression by real time RT-PCR
(Fig. 5). Compared with control
rats, the mRNA levels of caspase-3, -7 and -9 in CAP rats showed a
basically consistent change, in that all levels were markedly
increased at 1 h and peaked at 6 h (P<0.05, respectively), then
began to decrease and gradually returned to baseline at 24 h.
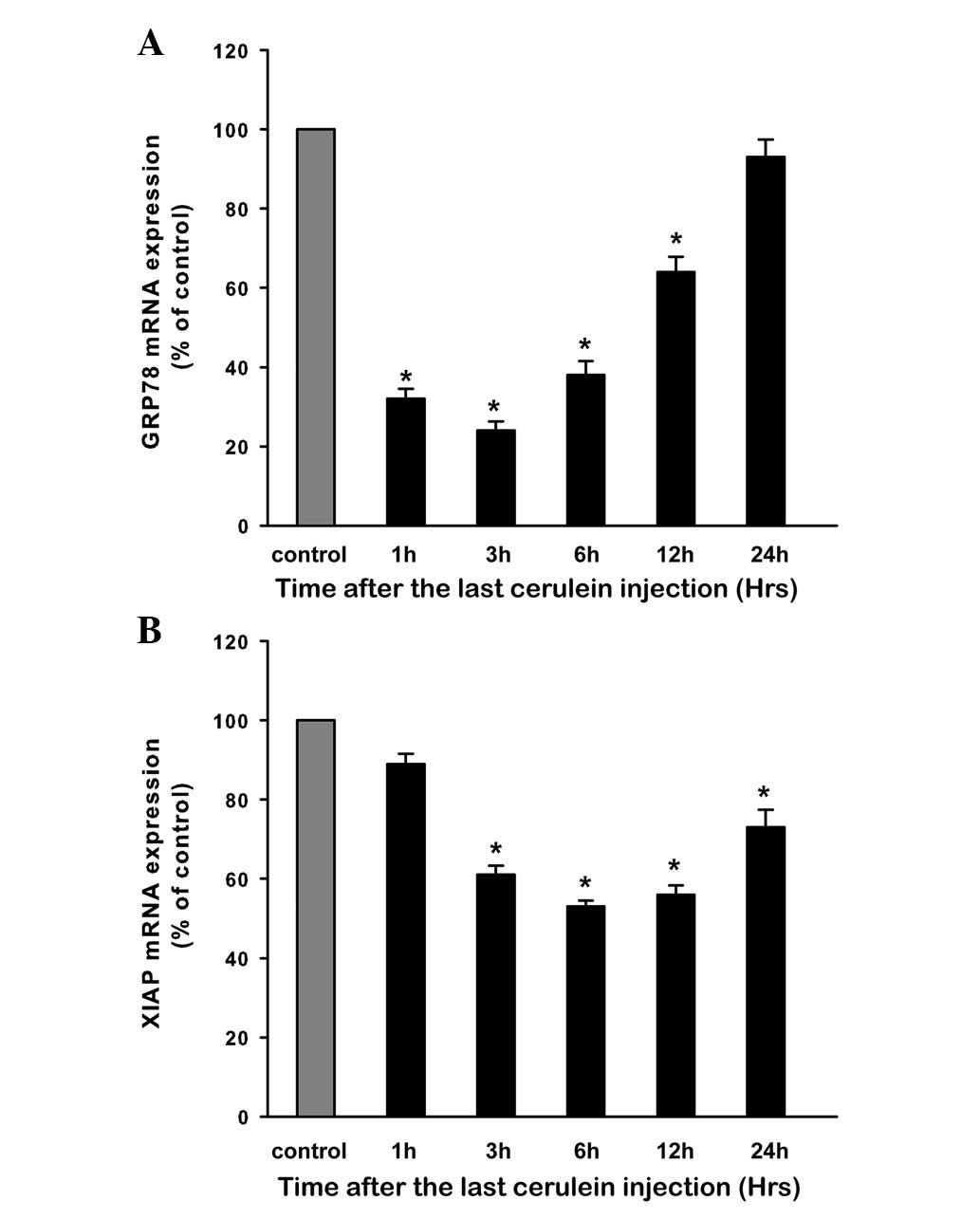
Expression of GRP78 during CAP
Expression of GRP78 mRNA in the pancreatic tissue
was examined by real-time RT-PCR. Compared with control rats, the
GRP78 mRNA level in CAP rats was decreased rapidly at 1 h and
reached a low peak at 3 h (P<0.05). The level then began to
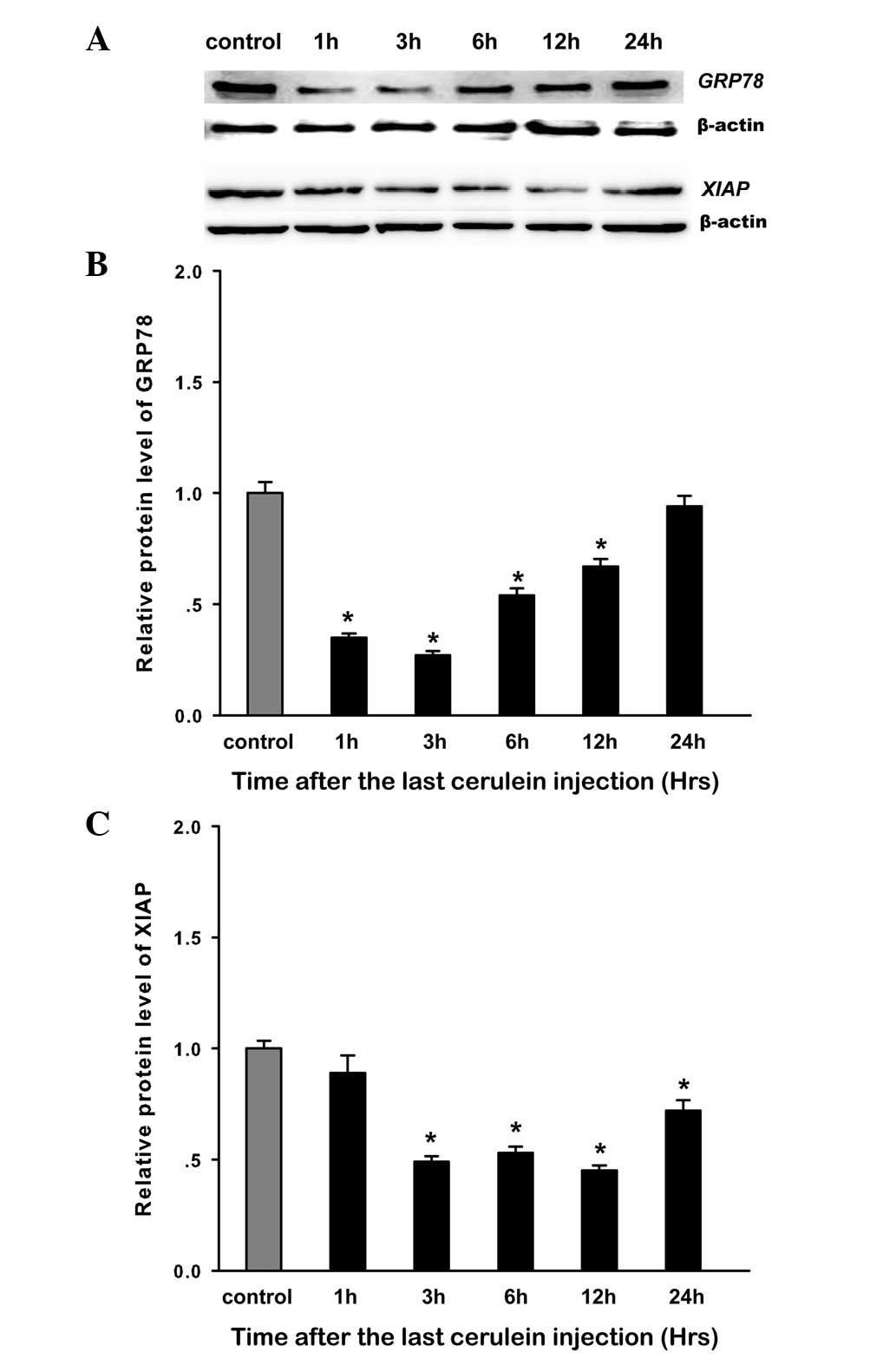
increase, and returned to baseline at 24 h (Fig. 6A). The expression of GRP78 protein
was evaluated by western blot analysis. The results showed a
similar trend to mRNA change in the expression of GRP78 protein
(Fig. 7A and B). A negative and
significant correlation was demonstrated between GRP78 expression
and the AI, and the expression of caspase-3, -7, and -9 (Pearson’s
correlation of −0.757, −0.669, −0.871, −0.704, respectively,
P<0.05 for all).
Expression of XIAP during CAP
Expression of XIAP mRNA in the pancreatic tissue was
examined by quantitative RT-PCR. In CAP rats, XIAP mRNA was
decreased at 3 h and the low level was maintained up to 24 h in
comparison with control rats (P<0.05; Fig. 6B). Western blot analysis showed a
similar trend for mRNA change in the expression of XIAP protein
(Fig. 7). XIAP expression was
negatively correlated with the AI and the expression of caspase-3,
-7 and -9 (Pearson’s correlation of −0.911, −0.950, −0.827, −0.855
respectively, P<0.05 for all).
Discussion
AP is characterized not only by inflammation but
also by parenchymal cell death (2,3,21).
Apoptosis and necrosis are observed in clinical as well as
experimental pancreatitis (5). Of
note, the severity of experimental pancreatitis directly correlates
with the extent of necrosis and inversely with that of apoptosis
(4,22). Thus, apoptosis appears to be a
beneficial response in the setting of AP. However, the mechanisms
regulating apoptosis in AP are unclear. To explore the mechanisms
regulating apoptosis response in AP, in the present study, we
analyzed the apoptosis response in CAP, the most commonly used and
well-characterized rat model of AP. Consistent with previous
studies, the induction of CAP was demonstrated by the elevation of
serum amylase activity, pancreatic edema, vacuolization,
inflammation and apoptosis. However, little necrosis was present in
CAP rats.
To explore the apoptotic signaling pathway during
CAP, we analyzed the pro-apoptotic caspase genes by real-time PCR.
We found the mRNA levels of the initiator caspase-9 and the
effector caspase-3 and -7 were rapidly and significantly increased
within 1 h and peaked at 6 h. Correspondingly, these increased
caspase levels were consistent with AI in CAP rats, as calculated
by TUNEL assay. These results indicated that the pro-apoptotic
signaling pathway was activated in CAP.
To further study the potential role of GRP78 in CAP,
we analyzed the expression of GRP78 by real time RT-PCR and western
blotting. The results showed that the mRNA and protein levels of
GRP78 were markedly downregulated in early CAP (within 1 h),
although the expression levels returned to baseline at 24 h. It is
notable that GRP78 was upregulated in the early stage of
arginine-induced severe rat AP, which exhibited low apoptosis in
pancreatic acinar cells (23). By
contrast, downregulation of GRP78 expression was found in a
cerulein-induced in vitro mild model of AP, which showed
high levels of apoptosis in AR42J cells (24). Taken together, these findings
suggest that GRP78 participates in the pathogenesis of AP, and it
may regulate apoptosis during AP. Moreover, previous studies have
indicated that GRP78 serves an anti-apoptotic role by forming
complexes with caspase-7 and caspase-12 and preventing their
activation and release (14,20).
In this study, the downregulated expression of GRP78 was negatively
correlated with the expression of caspase-3, -7 and -9 during
CAP.
There are other data indicating that GRP78 plays an
anti-apoptotic role, perhaps through interacting with XIAP
(18), which is the most potent
member of IAPs that negatively regulate caspase activation
(13). Indeed, it has been
indicated that XIAP was degraded rapidly and fully in rat CAP, and
that its degradation caused caspase activation (22). In the present study, the mRNA and
protein levels of XIAP were markedly downregulated within 3 h,
which was negatively correlated with the caspase gene expression
during CAP. Thus, these findings suggest that XIAP is an important
caspase regulator in AP. In particular, high XIAP levels block
caspase activation, whereas XIAP degradation facilitates caspase
activation.
In conclusion, our study revealed that GRP78 and
XIAP are downregulated in the process of CAP in rats. The
expression changes of GRP78 and XIAP were negatively correlated
with caspase gene expression and AI in CAP. These findings indicate
that downregulation of GRP78 and XIAP was correlated with apoptosis
through negatively regulating caspase activation during CAP in
rats.
Acknowledgements
This work was supported by grants from the National
Natural Science Fund of China (NSFC key project No. 30830100;
project No. 30972924; project No. 81170439), the Project sponsored
by SRF for ROCS, SEM (No. 20101174-4-2) and the Research Fund for
the Doctoral Program of Higher Education, SEM (No.
200806100058).
Abbreviations:
|
CAP
|
cerulein-induced acute
pancreatitis
|
|
GRP78
|
78-kDa glucose-regulated protein
|
|
XIAP
|
X-linked inhibitor of apoptosis
protein
|
References
|
1
|
Frossard JL, Steer ML and Pastor CM: Acute
pancreatitis. Lancet. 371:143–152. 2008. View Article : Google Scholar
|
|
2
|
Pandol SJ, Saluja AK, Imrie CW and Banks
PA: Acute pancreatitis: bench to the bedside. Gastroenterology.
132:1127–1151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gukovskaya AS, Mareninova OA, Odinokova
IV, et al: Cell death in pancreatitis: Effects of alcohol. J
Gastroenterol Hepatol. 213(Suppl 3): S10–S13. 2006. View Article : Google Scholar
|
|
4
|
Bhatia M: Apoptosis of pancreatic acinar
cells in acute pancreatitis: is it good or bad? J Cell Mol Med.
8:402–409. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bhatia M: Apoptosis versus necrosis in
acute pancreatitis. Am J Physiol Gastrointest Liver Physiol.
286:G189–G196. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fiers W, Beyaert R, Declercq W and
Vandenabeele P: More than one way to die: apoptosis, necrosis and
reactive oxygen damage. Oncogene. 18:7719–7730. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chao KC, Chao KF, Chuang CC and Liu SH:
Blockade of interleukin 6 accelerates acinar cell apoptosis and
attenuates experimental acute pancreatitis in vivo. Br J Surg.
93:332–338. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao M, Xue DB, Zheng B, Zhang WH, Pan SH
and Sun B: Induction of apoptosis by artemisinin relieving the
severity of inflammation in caerulein-induced acute pancreatitis.
World J Gastroenterol. 13:5612–5617. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wolf BB and Green DR: Suicidal tendencies:
apoptotic cell death by caspase family proteinases. J Biol Chem.
274:20049–20052. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee HC and Wei YH: Mitochondrial role in
life and death of the cell. J Biomed Sci. 7:2–15. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lomberk G and Urrutia R: Primers on
molecular pathways - caspase pathway. Pancreatology. 9:6–8. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Deveraux QL, Takahashi R, Salvesen GS and
Reed JC: X-linked IAP is a direct inhibitor of cell-death
proteases. Nature. 388:300–304. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salvesen GS and Duckett CS: IAP proteins:
blocking the road to death’s door. Nat Rev Mol Cell Biol.
3:401–410. 2002.
|
|
14
|
Rao RV, Peel A, Logvinova A, et al:
Coupling endoplasmic reticulum stress to the cell death program:
role of the ER chaperone GRP78. FEBS Lett. 514:122–128. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BiP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pfaffenbach KT and Lee AS: The critical
role of GRP78 in physiologic and pathologic stress. Curr Opin Cell
Biol. 23:150–156. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Misra UK, Deedwania R and Pizzo SV:
Activation and cross-talk between Akt, NF-kappaB, and unfolded
protein response signaling in 1-LN prostate cancer cells consequent
to ligation of cell surface-associated GRP78. J Biol Chem.
281:13694–13707. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moon K, Zhou SY, Lee SH, Owyang C and
DiMagno MJ: Hsp70 and XIAP are potential key molecular mechanisms
causing impaired apoptosis and severe acute pancreatitis (AP) in CF
mice. Pancreas. 35:4172007. View Article : Google Scholar
|
|
19
|
Reddy RK, Mao C, Baumeister P, Austin RC,
Kaufman RJ and Lee AS: Endoplasmic reticulum chaperone protein
GRP78 protects cells from apoptosis induced by topoisomerase
inhibitors: role of ATP binding site in suppression of caspase-7
activation. J Biol Chem. 278:20915–20924. 2003. View Article : Google Scholar
|
|
20
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative CT method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gukovskaya AS, Gukovsky I, Jung Y, Mouria
M and Pandol SJ: Cholecystokinin induces caspase activation and
mitochondrial dysfunction in pancreatic acinar cells. Roles in cell
injury processes of pancreatitis. J Biol Chem. 277:22595–22604.
2002. View Article : Google Scholar
|
|
22
|
Mareninova OA, Sung KF, Hong P, et al:
Cell death in pancreatitis: caspases protect from necrotizing
pancreatitis. J Biol Chem. 281:3370–3381. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kubisch CH, Sans MD, Arumugam T, Ernst SA,
Williams JA and Logsdon CD: Early activation of endoplasmic
reticulum stress is associated with arginine-induced acute
pancreatitis. Am J Physiol Gastrointest Liver Physiol.
291:G238–G245. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu JH, Lim JW, Kim KH, Morio T and Kim H:
NADPH oxidase and apoptosis in cerulein-stimulated pancreatic
acinar AR42J cells. Free Radical Biol Med. 39:590–602. 2005.
View Article : Google Scholar : PubMed/NCBI
|