Introduction
Epidermal growth factor receptor (EGFR) is a 175-kDa
trans-membrane glycoprotein in the tyrosine kinase family of growth
factor receptors (1). EGFR has
been reported to be overexpressed in various types of cancer,
including lung, colon and head and neck cancer. Additionally, the
activation of the EGFR cascades may result in various cell
responses including apoptosis, development, cell proliferation,
differentiation and survival (2–7). The
overexpression and activation of EGFR is a common mechanism by
which EGFR exerts its influence on tumorigenesis (1,8–11).
Thus, EGFR is a potential target for cancer therapy due to its
expression and activity in cancer tissues.
Natural products from medicinal plants have been
used for a long time due to their potential chemotherapeutic
activity (12,13). A recent review by Graham et
al reported the use of Plantago major L. (PML) as an
antitumor agent (13–15). Another plant that is used as a
medicinal agent for the treatment of dry cough, catarrhal
inflammation of the throat and oesophagus, as well as irregular or
absent menstrual cycles is Althaea rosea cavanil (ARC)
(16). However, the mechanisms of
PML and ARC involved in the suppression of various tumors are
unknown, although PML and ARC have been used as traditional
medicines to treat several diseases.
In this study, methanolic extracts from Plantago
major L. (MPML) and Althaea rosea Cavanil (MARC) were
studied for their anti-tumorigenic effects via the inhibition of
transformed normal cells. Consequently, the present study aimed to
identify the molecular mechanisms and direct targets of the
anti-tumorigenic effects of MARC and MPML in tumor promoter
(EGF)-induced neoplastic cell transformation.
Materials and methods
Reagents
Eagle’s minimal essential medium (MEM) and
Dulbecco’s modified essential medium (DMEM) were obtained from
Welgene, Inc. (Korea). EGF was purchased from Invitrogen (Carlsbad,
CA, USA). Antibodies to pEGFR and EGFR were obtained from Cell
Signaling, Inc. (Danvers, MA, USA). Anti-actin antibody was
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
MARC and MPML were kindly supplied by Professor Ki Han Kwon
(Gwangju University, Gwangju, Korea).
Cell culture and chemical treatments
EGFR+/+ and EGFR−/− mouse
embryonic fibroblasts (MEFs) were kindly provided by Professor Dong
(University of Minnesota, Hormel Institute, MN, USA) (11) and the JB6 P+ (JB6 CI 41–5a) mouse
epidermal cell line was obtained from the American Tissue Culture
Collection (Manassas, VA, USA). The JB6 P+ cells were cultured in
MEM containing 5% fetal bovine serum (FBS) and MEF cells were
maintained in DMEM containing 10% FBS, each with 100 U/ml of
penicillin (Welgene, Inc.), in a humidified atmosphere containing
5% CO2. An equal number of cells were seeded and allowed
to attach overnight. The cells were starved in 0.1% FBS/MEM or DMEM
for 24 h, incubated with MARC (30, 60 and 90 μg/ml) or MPML (20, 40
and 60 μg/ml) for 1 h and treated with 10 ng/ml of EGF.
MTS assay
The effects of MARC and MPML on cell viability were
estimated using the CellTiter 96® Aqueous One Solution
Cell Proliferation Assay (Promega, Madison, WI, USA) according to
the manufacturer’s instructions. In brief, cells were seeded in
96-well plates and then incubated with various doses of MARC (30,
60, 90 and 120 μg/ml) and MPML (20, 40, 60 and 80 μg/ml) for 72 h.
MTS
(3-(4,5-dimethylthiazol-20yl)-(3-carboxymethoxyphenyl)-2-(4-sulphophenyl)-2H-tetrazolium)
solution was added to each well and incubated for 2 h at 37°C. The
reaction buffer was analyzed using an ELISA microplate reader
(BioTeck Instruments Inc., Winooski, VT, USA) at 490 and 690 nm
(background).
Anchorage-independent cell transformation
assay
The effect of MARC and MPML on EGF-induced cell
transformation was estimated in JB6 P+ cells. The cells were
treated with EGF (10 ng/ml) with or without various doses of MARC
and MPML in 1 ml of 0.33% basal medium Eagle’s (BME) agar over 3 ml
of 0.5% BME agar containing 10% FBS. The cultures were maintained
at 37°C in a 5% CO2 incubator for 10 days and the cell
colonies were counted using a microscope.
Western blot analysis
Whole cell lysates were collected in lysis buffer
for the western blot analysis. The protein supernatant fractions
were subjected to SDS-PAGE and then transferred on polyvinylidene
difluoride membranes and blocked with 5% skimmed milk followed by
hybridization with the indicated antibodies. The protein bands with
horseradish peroxidase-conjugated secondary antibody were observed
using a western blotting luminol reagent (Santa Cruz Biotechnology,
Inc.).
Statistical analysis
Data were assessed as the means ± SD. of triplicate
samples from at least three independent experiments. Statistical
significance was evaluated using a Student’s t-test or one-way
ANOVA and was considered significant when p<0.05.
Results
MARC and MPML do not affect cell
proliferation in JB6 P+ cells
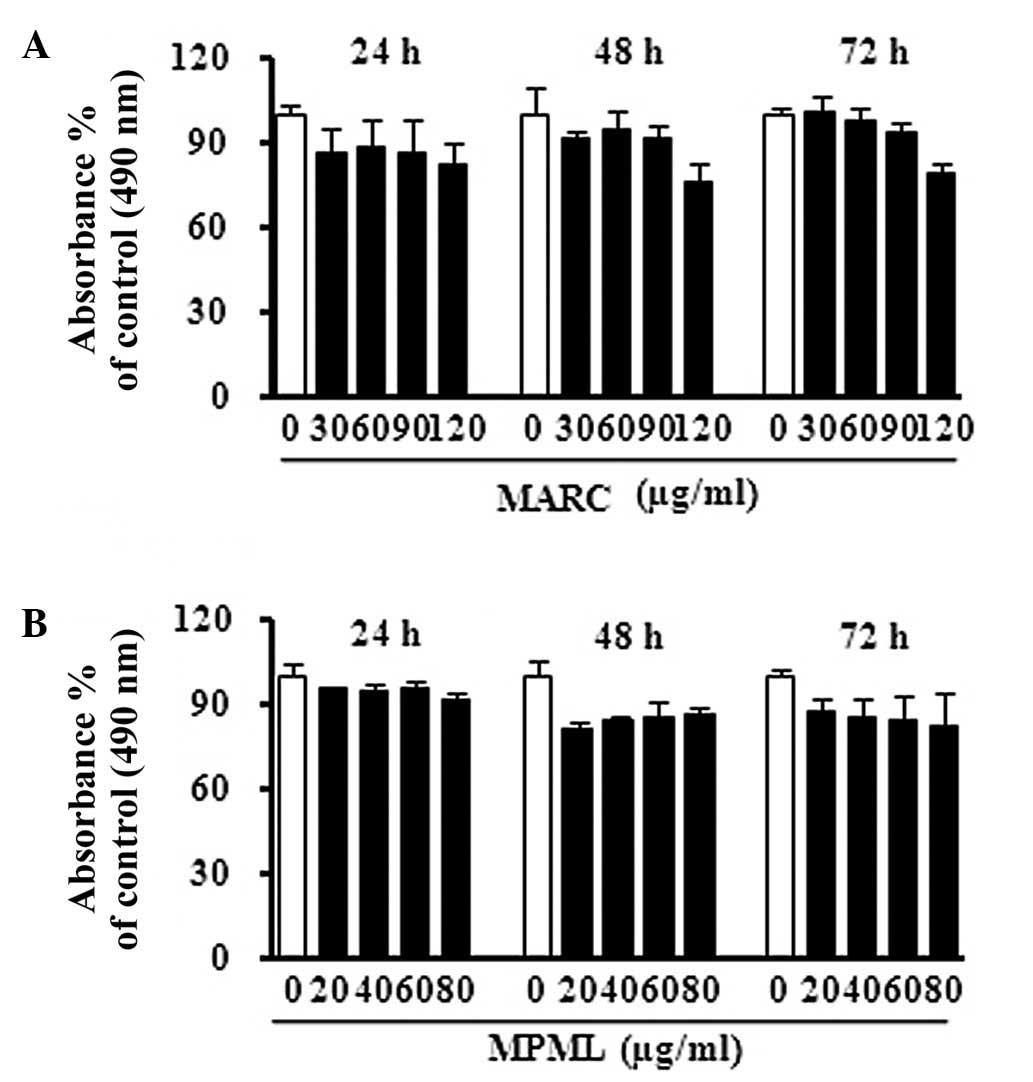
To determine whether MARC and MPML had cell growth
inhibitory effects, we treated JB6 P+ epidermal mouse skin cells
with MARC and MPML at a range of doses and assessed viability using
the MTS assay. The results showed that MARC and MPML did not
significantly affect cell proliferation at 24, 48 or 72 h after
treatment (Fig. 1).
MARC and MPML markedly suppress
EGF-induced neoplastic transformation in JB6 P+ cells
The JB6 P+ cell line is an excellent model to study
EGF-induced cell transformation (17). We evaluated whether MARC and MPML
is directly associated with EGF-induced neoplastic transformation.
The results showed that MARC and MPML significantly decreased the
EGF-promoted colony number and colony size in a dose-dependent
manner. Colony formation and colony size induced by EGF-induced JB6
P+ cells was also significantly less than that observed in the
non-treated JB6 P+ cells (Fig. 2A and
B).
MARC and MPML inhibit EGF-induced EGFR
phosphorylation in JB6 P+ cells
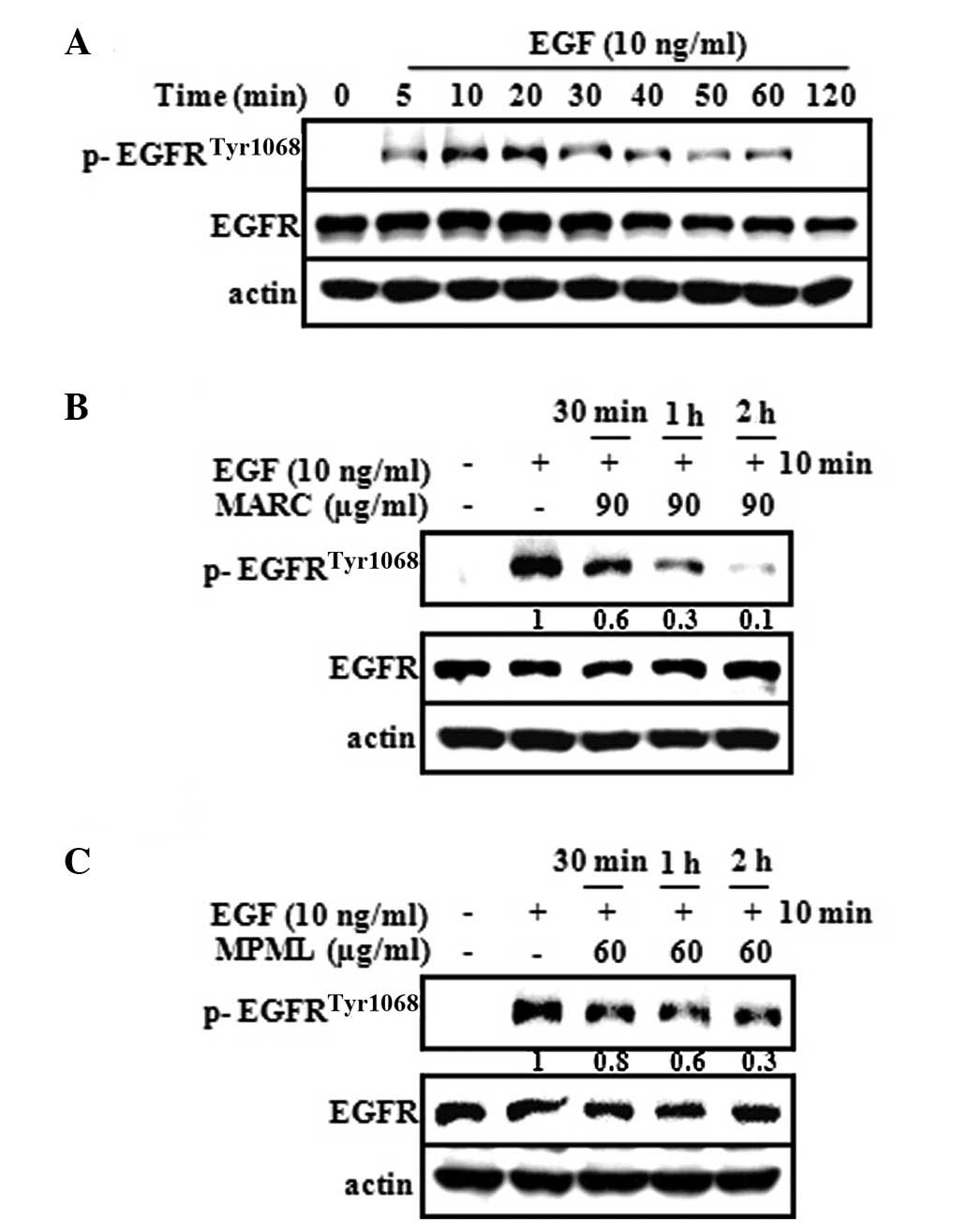
Numerous studies have shown that EGF promoted skin
carcinogenesis through EGFR activation (6,7). To
verify whether phosphorylation of EGFR was induced by EGF, we
tested the expression of EGFR phosphorylation with EGF for
increasing time periods. The results showed that the expression of
EGFR phosphorylation significantly increased within 5–20 min after
treatment with EGF (Fig. 3A). We
tested whether MARC and MPML downregulated the phosphorylation of
EGFR activated by EGF in JB6 P+ cells. Our results indicated that
MARC (90 μg/ml) and MPML (60 μg/ml) suppressed the EGF-induced
phosphorylation of EGFR (Fig. 3B and
C). These results suggested that the inactivation of EGFR by
MARC and MPML leads to the suppression of neoplastic
transformation.
MARC and MPML inhibit the EGF-induced
phosphorylation of EGFR and suppress the EGF-induced cell
proliferation in an MEF cell line
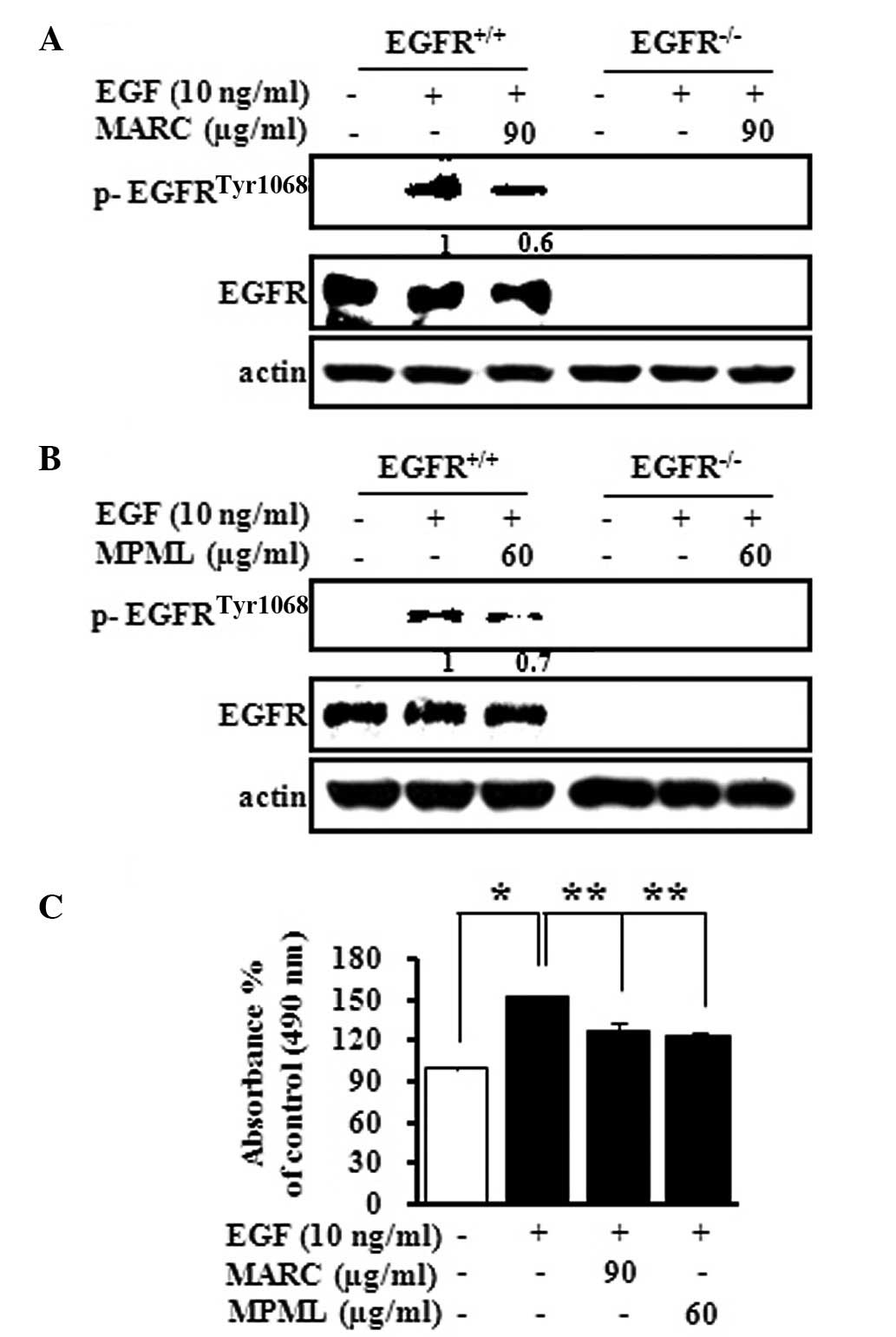
MARC and MPML strongly inhibited the EGF-induced
phosphorylation of EGFR in JB6 P+ cells (Fig. 3). Thus, we investigated the direct
effect of MARC and MPML on the phosphorylation of EGFR in EGF
knockout MEF cells (EGFR−/−) and EGFR-expressing MEF
cells (EGFR+/+). MARC and MPML decreased the EGF-induced
phosphorylation of EGFR in EGFR/WT MEF cells, but not in EGFR KO
MEF cells (Fig. 4A and B). To
further explore the inhibitory effect of MARC and MPML on
EGF-induced cell proliferation, EGFR+/+ cells were
cultured for 24 h in 96-well culture plates with MARC (90 μg/ml)
and MPML (60 μg/ml) and cell proliferation was measured using the
MTS assay. Treatment with EGF (10 ng/ml) resulted in an ~60%
increase compared to the DMSO control in the EGFR+/+
cells, but MARC (90 μg/ml) and MPML (60 μg/ml) inhibited
EGF-induced cell proliferation in EGFR+/+ cells. These
results indicated that EGFR+/+ MEF proliferation affects
the inhibition of EGFR activation by MARC and MPML.
Discussion
Cancer is a major public health problem. Numerous
studies have indicated that chemotherapeutic drugs cannot
successfully treat cancer as they have severe side effects
including various types of rash, hair loss, painful paronychia and
xerosis cutis (18). Plants as
medicinal agents have been used in the treatment of various types
of diseases in humans (19–23)
and a number of investigators have also reported that natural
products may be safe and free from side effects (24–27).
Particularly, PML as a natural product, has been reported to exert
its anticancer effect via DNA damage in cancer cells (15). Wang et al also reported that
ARC inhibited inflammation in rat paw edema (28). However, no study currently exists
on the anti-cancer activity and related molecular targets of ARC
and PML.
In this study, we focused on three primary
objectives in relation to the chemopreventive effect of ARC and PML
in EGF-induced neoplastic cell transformation. The first was to
examine the effect of ARC and PML on the cell toxicity of mouse
epidermal JB6 P+ cells. The second was to determine the
anti-tumorigenic effect of ARC and PML on EGF-induced neoplastic
cell transformation. The third was to identify critical key
molecules in mouse skin anti-cancer activities by ARC and PML. For
this study, ARC and PML were extracted with methanol. We
investigated whether MARC and MPML were cytotoxic in the mouse
epidermal JB6 P+ cells. The results showed that MARC and MPML did
not have any effect on the viability of JB6 P+ cells when treated
at various doses.
The irregular biological process of tumorigenesis is
associated with the abnormal regulation of growth signaling. EGF is
one of the tumor promoters that is capable of causing the induction
of abnormal cell growth (29) and
the EGF-EGFR transduction system has been confirmed to be
significantly increased in human cancer cells of the skin, breast,
colon, lung and prostate to stimulate cell proliferation,
invasiveness and angiogenesis (11,30,31).
The JB6 P+ cell line is a well-established system extensively used
as an in vitro model for the study of tumor promotion
(32,33) and provides an ideal model for the
investigation of the molecular mechanisms involved in neoplastic
transformation, promotion and progression (11,34–36).
Notably, it has been reported that EGF induced the formation of
anchorage-independent colonies in JB6 P+ cells using a soft agar
assay (11,33,37).
Therefore, we investigated whether MARC and MPML are capable of
suppressing the EGF-induced neoplastic cell transformation. We
showed that MARC and MPML clearly inhibited EGF-induced neoplastic
cell transformation as well as colony number and size in
EGF-treated JB6 P+ cells, indicating that MARC and MPML could be
acting as anti-cancer agents. EGFR has been recognized as a
convergence point for diverse signal transduction pathways
(37–39). After EGF binds with the ectodomain
of the EGFR, which exists as homodimers or heterodimers, this leads
to the autophosphorylation of tyrosine residues in the cytoplasmic
domain and activation of the receptor’s intrinsic kinase activity
(11,40). Notably, EGFR is frequently
overexpressed and abnormally activated in many types of cancer.
Thus, we examined whether MARC and MPML inhibited EGF-induced EGFR
phosphorylation to suppress neoplastic cell transformation in
EGFR-expressing cells (JB6 P+, EGFR+/+). These results
suggested that MARC and MPML significantly inhibited the
phosphorylation of EGFR by EGF on JB6 P+ and EGFR+/+
cells, but not EGFR−/− cells. MARC and MPML effectively
inhibited the EGF-induced proliferation of EGFR+/+ cell
lines. Our data suggest that the dephosphorylation of EGFR may be
associated with MARC- and MPML-suppressed neoplastic cell
transformation and cell proliferation in JB6 P+ cells.
In conclusion, MARC and MPML have suppressed
EGF-induced transformation and proliferation through the
dephosphorylation of EGFR. Thus, MARC and MPML are capable of
inhibiting the growth of cancer cells through EGFR and may be
developed into effective anticancer agents against various types of
cancer that highly express EGFR.
Acknowledgements
This study was supported by the Basic Science
Research program through the National Research Foundation Korea
(NRF) funded by the Ministry of Education, Science and Technology
(2012-0003226), and the Next-Generation BioGreen 21 Program
(PJ008116062011), Rural Development Administration, Republic of
Korea.
Abbreviations:
|
ARC
|
Althaea rosea Cavanil
|
|
PML
|
Plantago major L.
|
|
EGF
|
epidermal growth factor
|
|
MARC
|
methanol extracts Althaea rosea
Cavanil
|
|
MPML
|
methanol extracts Plantago
major L.
|
|
EGFR
|
epidermal growth factor receptor
|
|
MEM
|
minimal essential medium
|
|
DMEM
|
Dulbecco’s modified essential
medium
|
|
MEFs
|
mouse embryonic fibroblasts
|
|
FBS
|
fetal bovine serum
|
|
MTS
|
(3-(4,5-dimethylthiazol-20yl)-(3-carboxymethoxyphenyl)-2-(4-sulphophenyl)-2H-tetrazolium)
|
|
BME
|
basal medium Eagle
|
References
|
1
|
Wang H, Zhou M, Shi B, et al:
Identification of an exon 4-deletion variant of epidermal growth
factor receptor with increased metastasis-promoting capacity.
Neoplasia. 13:461–471. 2011.PubMed/NCBI
|
|
2
|
Xue C, Wyckoff J, Liang F, et al:
Epidermal growth factor receptor overexpression results in
increased tumor cell motility in vivo coordinately with enhanced
intravasation and metastasis. Cancer Res. 66:192–197. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pandiella A, Lehvaslaiho H, Magni M,
Alitalo K and Meldolesi J: Activation of an EGFR/neu chimeric
receptor: early intracellular signals and cell proliferation
responses. Oncogene. 4:1299–1305. 1989.PubMed/NCBI
|
|
4
|
Liang K, Ang KK, Milas L, Hunter N and Fan
Z: The epidermal growth factor receptor mediates radioresistance.
Int J Radiat Oncol Biol Phys. 57:246–254. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ho R, Minturn JE, Hishiki T, et al:
Proliferation of human neuroblastomas mediated by the epidermal
growth factor receptor. Cancer Res. 65:9868–9875. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Modjtahedi H and Essapen S: Epidermal
growth factor receptor inhibitors in cancer treatment: advances,
challenges and opportunities. Anticancer Drugs. 20:851–855. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Engelman JA and Cantley LC: A sweet new
role for EGFR in cancer. Cancer Cell. 13:375–376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chaffanet M, Chauvin C, Laine M, et al:
EGF receptor amplification and expression in human brain tumours.
Eur J Cancer. 28:11–17. 1992. View Article : Google Scholar
|
|
9
|
Nishikawa R, Ji XD, Harmon RC, et al: A
mutant epidermal growth factor receptor common in human glioma
confers enhanced tumorigenicity. Proc Natl Acad Sci USA.
91:7727–7731. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang P, Steck PA and Yung WK: The
autocrine loop of TGF-alpha/EGFR and brain tumors. J Neurooncol.
35:303–314. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hwang MK, Bode AM, Byun S, et al:
Cocarcinogenic effect of capsaicin involves activation of EGFR
signaling but not TRPV1. Cancer Res. 70:6859–6869. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Newman DJ, Cragg GM and Snader KM: Natural
products as sources of new drugs over the period 1981–2002. J Nat
Prod. 66:1022–1037. 2003.PubMed/NCBI
|
|
13
|
Graham JG, Quinn ML, Fabricant DS and
Farnsworth NR: Plants used against cancer - an extension of the
work of Jonathan Hartwell. J Ethnopharmacol. 73:347–377. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Samuelsen AB: The traditional uses,
chemical constituents and biological activities of Plantago
major L. A review. J Ethnopharmacol. 71:1–21. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galvez M, Martin-Cordero C, Lopez-Lazaro
M, Cortes F and Ayuso MJ: Cytotoxic effect of Plantago spp.
on cancer cell lines. J Ethnopharmacol. 88:125–130. 2003.
|
|
16
|
Papiez MA: The influence of hollyhock
extract administration on testicular function in rats. J Mol
Histol. 35:733–740. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang NJ, Lee KW, Rogozin EA, et al: Equol,
a metabolite of the soybean isoflavone daidzein, inhibits
neoplastic cell transformation by targeting the
MEK/ERK/p90RSK/activator protein-1 pathway. J Biol Chem.
282:32856–32866. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ehmann LM, Heinemann V and Wollenberg A:
New tyrosine kinase and EGFR inhibitors in cancer therapy: Cardiac
and skin toxicity as relevant side effects. Part B: skin. Internist
(Berl). 52:1359–1364. 2011.(In German).
|
|
19
|
Cheung CW, Gibbons N, Johnson DW and Nicol
DL: Silibinin - a promising new treatment for cancer. Anticancer
Agents Med Chem. 10:186–195. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fahey JW, Haristoy X, Dolan PM, et al:
Sulforaphane inhibits extracellular, intracellular, and
antibiotic-resistant strains of Helicobacter pylori and
prevents benzo[a]pyrene-induced stomach tumors. Proc Natl Acad Sci
USA. 99:7610–7615. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yanaka A, Fahey JW, Fukumoto A, et al:
Dietary sulforaphane-rich broccoli sprouts reduce colonization and
attenuate gastritis in Helicobacter pylori-infected mice and
humans. Cancer Prev Res (Phila). 2:353–360. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Butler MS: The role of natural product
chemistry in drug discovery. J Nat Prod. 67:2141–2153. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shokrzadeh M, Azadbakht M, Ahangar N,
Hashemi A and Saeedi Saravi SS: Cytotoxicity of hydro-alcoholic
extracts of Cucurbitapepo and Solanum nigrum on HepG2 and CT26
cancer cell lines. Pharmacogn Mag. 6:176–179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Etkin NL: A Hausa herbal pharmacopoeia:
biomedical evaluation of commonly used plant medicines. J
Ethnopharmacol. 4:75–98. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Misumi N, Hiraike M, Nawata F, et al:
[Protective effects of d-chlorpheniramine maleate pre-treatment
against acute side effects of Irinotecan(CPT- 11)]. Gan To Kagaku
Ryoho. 38:1149–1153. 2011.(In Japanese).
|
|
27
|
Sun X, Zhang J, Gupta R, Macgibbon AK,
Kuhn-Sherlock B and Krissansen GW: Dairy milk fat augments
paclitaxel therapy to suppress tumour metastasis in mice, and
protects against the side-effects of chemotherapy. Clin Exp
Metastasis. 28:675–688. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang DF, Shang JY and Yu QH: Analgesic and
anti-inflammatory effects of the flower of Althaea rosea (L.) Cav.
Zhongguo Zhong Yao Za Zhi. 14(46–48): 641989.(In Chinese).
|
|
29
|
Furuse J: Growth factors as therapeutic
targets in HCC. Crit Rev Oncol Hematol. 67:8–15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Khalil MY, Grandis JR and Shin DM:
Targeting epidermal growth factor receptor: novel therapeutics in
the management of cancer. Expert Rev Anticancer Ther. 3:367–380.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mimeault M, Pommery N and Henichart JP:
New advances on prostate carcinogenesis and therapies: involvement
of EGF-EGFR transduction system. Growth Factors. 21:1–14. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Amstad PA, Liu H, Ichimiya M, Berezesky IK
and Trump BF: Manganese superoxide dismutase expression inhibits
soft agar growth in JB6 clone41 mouse epidermal cells.
Carcinogenesis. 18:479–484. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang NJ, Lee KW, Kwon JY, et al:
Delphinidin attenuates neoplastic transformation in JB6 Cl41 mouse
epidermal cells by blocking Raf/mitogen-activated protein kinase
kinase/extracellular signal-regulated kinase signaling. Cancer Prev
Res (Phila). 1:522–531. 2008. View Article : Google Scholar
|
|
34
|
Dong Z and Bode AM: Proceedings -
targeting carcinogenesis: transduction, transcription, translation.
Mol Carcinog. 45:353–354. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nguyen-Ba G and Vasseur P: Epigenetic
events during the process of cell transformation induced by
carcinogens (Review). Oncol Rep. 6:925–932. 1999.PubMed/NCBI
|
|
36
|
Lee KW, Kang NJ, Rogozin EA, et al:
Myricetin is a novel natural inhibitor of neoplastic cell
transformation and MEK1. Carcinogenesis. 28:1918–1927. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ciardiello F and Tortora G: EGFR
antagonists in cancer treatment. N Engl J Med. 358:1160–1174. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gschwind A, Zwick E, Prenzel N, Leserer M
and Ullrich A: Cell communication networks: epidermal growth factor
receptor transactivation as the paradigm for interreceptor signal
transmission. Oncogene. 20:1594–1600. 2001. View Article : Google Scholar
|
|
39
|
Fichera A, Little N, Jagadeeswaran S, et
al: Epidermal growth factor receptor signaling is required for
microadenoma formation in the mouse azoxymethane model of colonic
carcinogenesis. Cancer Res. 67:827–835. 2007. View Article : Google Scholar
|
|
40
|
Wakeling AE, Guy SP, Woodburn JR, et al:
ZD1839 (Iressa): an orally active inhibitor of epidermal growth
factor signaling with potential for cancer therapy. Cancer Res.
62:5749–5754. 2002.PubMed/NCBI
|