Introduction
Spermatogenesis is composed of three phases,
mitosis, meiosis and spermiogenesis, through which spermatogonial
stem cells (SSCs) self-renew and differentiate into sperm. To
maintain normal homeostasis, the balance between self-renewal and
differentiation of SSCs must be closely regulated. Increasing
evidence has revealed that the processes of maintenance and
self-renewal of SSCs and testicular development are regulated by a
variety of factors and involves the participation of multiple genes
(1,2).
Multinucleated cells are present in a variety of
sites and in association with several conditions. Coenocytes have
been identified in specific pathological processes, including cells
affected by chemicals, drug injury or exposure to adverse factors,
including thermal or mechanical damage (3–9).
However, the involvement of multinucleated cells in normal
testicular development requires further clarification. However, the
appearance of multinucleated cells in normal testicular development
processes has not been reported.
In the present study, the possible mechanism and
role of multinucleated cells in normal testicular development was
investigated. Results revealed that multinucleated cells occur
during the mouse testicular development process. To explore the
mechanism of the occurrence of multinucleated cells in testicular
development, the expression of cell proliferation- and
apoptosis-related proteins, including proliferating cell nuclear
antigen (PCNA), cyclin D1, caspase 3 and Bax was detected in
multinucleated cells of mouse testicle tissue (day 30 and 33
following birth).
Materials and methods
Animals
Experiments were conducted following the Guide for
Care and Use of Laboratory Animals (NIH Guide). Protocols for the
use of animals were approved by the Department of Laboratory Animal
Sciences (Kunming Medical University, Kunming, China). Experimental
healthy male Kunming mice were purchased from the Laboratory Animal
Center of Kunming Medical University and were divided into the
following groups: postnatal day 1, 4, 8, 11, 15, 20, 23, 27, 30,
33, 36, 40, 43, 47, 50, 55 and 57 (n=5).
Reagents
In situ cell apoptosis detection kit and
antibodies against PCNA, cyclin D1, caspase 3 and Bax were
purchased from Fuzhou new biotechnology company (Fuzhou Maxim,
Fuzhou, China). All other reagents were from Sigma-Aldrich (St.
Louis, MO, USA) unless otherwise stated.
Histological analysis
Testes were obtained from mice sacrificed by
cervical dislocation and fixed in 10% formaldehyde, dehydrated in
ascending concentrations of ethanol (70–100%), embedded in paraffin
and cut into 3-μm sections using a microtome (RM2125RT, Leica
Microsystems Trading Ltd., Shanghai, China), mounted and stained
with hematoxylin and eosin (H&E).
TUNEL analysis
Sections of postnatal days 30 and 33 testes were
incubated at 60°C in the oven for 40 min. Following dewaxing with
two washes of xylene, 1 ml 100% ethanol was added to remove
residual xylene. The sections were then rehydrated with 95, 85, 70
and 50% ethanol for 3 min at room temperature and then rinsed with
PBS. Sections were incubated with pepsin for 15 min at room
temperature. Following two 2-min PBS Tween-20 rinses, sections were
incubated in 50 μl alkaline phosphatase antibody at 37°C for 30
min. Sections were washed with PBS three times and incubated for 20
min at room temperature with 5-bromo-4-chloro-3-indolyl
phosphate/nitro blue tetrazolium solution. Following rinsing with
PBS three times, sections were labeled with nuclear fast red for 15
min. These sections were visualized under an Olympus inverted
microscope (Olympus Corporation, Tokyo, Japan) and representative
images were captured. In situ detection of
apoptosis-positive cells appeared purple or dark purple and
negative cells appeared pink.
Immunohistochemistry
Sections of postnatal day 30 and 33 testes were
incubated at 60°C in the oven for 40 min. Following dewaxing with
two washes of xylene, 1 ml 100% ethanol was added to remove
residual xylene. The sections were rehydrated with 95, 85, 70 and
50% ethanol for 3 min at room temperature and rinsed with PBS.
Tissue sections were soaked in distilled water to prepare for use.
Following antigen retrieval using citrate buffer at high
temperature and high pressure, sections were incubated with 50 μl
3% H2O2 solution at room temperature for 10
min to block endogenous peroxidase activity. Sections were then
rinsed with PBS three times and non-specific binding was blocked
with normal goat serum. Incubation with the primary antibodies was
performed at room temperature for 60 min. Sections were washed with
PBS three times, incubated with MaxVisionTM rapid
immunochemistry reagent (Fuzhou Maxim) for 15 min at room
temperature, washed again and incubated in 0.03% (w/v)
3,3′-diaminobenzidine with 0.003% (v/v) hydrogen peroxide until a
brown reaction product was observed.
Quantification of multinucleated cell
variety in mouse age groups
Five slides were counted from each group under a
microscope. To quantify the percentage of seminiferous tubules
containing multinucleated cells, six fields of vision were randomly
selected under magnification, ×200. Following this, 20 seminiferous
tubules containing multinucleated cells were selected and the
number of multinucleated cells was counted under magnification,
×400. Results were presented as mean ± SEM.
Results
Histological analysis
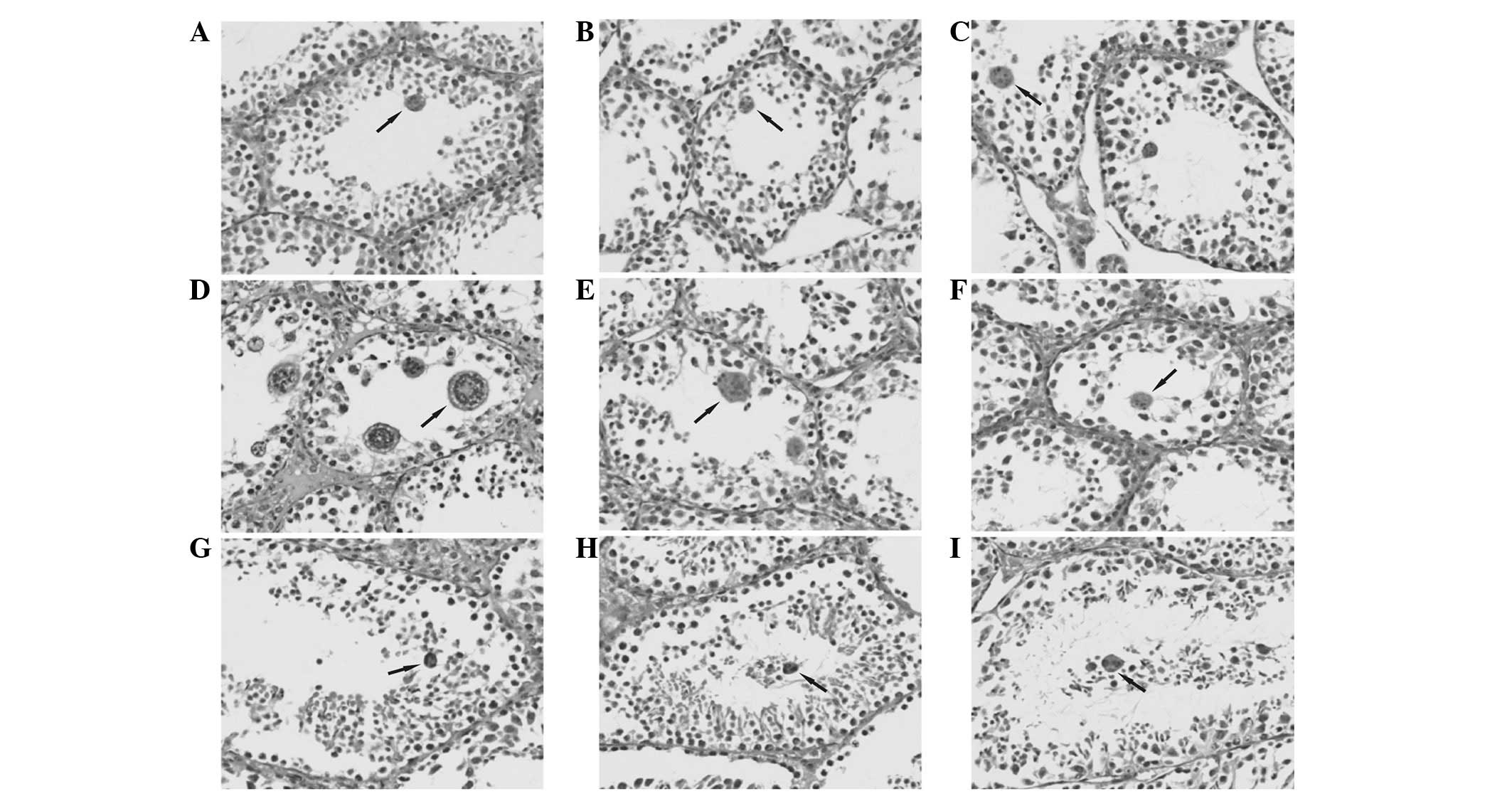
Multinucleated cells were not present in the
seminiferous tubules at postnatal days 1, 4, 8, 11, 15, 20 and 43
in mouse testicular tissue. In seminiferous sections of tubules in
mouse testicular tissues at day 23, 27, 30, 33, 36, 40, 47, 50 and
54, multinucleated cells were observed by a collection of multiple
nuclei close to the lumen. The numbers ranged between several and
one dozen and the nuclei were as small as the secondary
spermatocytes and sperm nucleus and arranged in piles or as a
flower ring (Fig. 1).
Immunohistochemistry
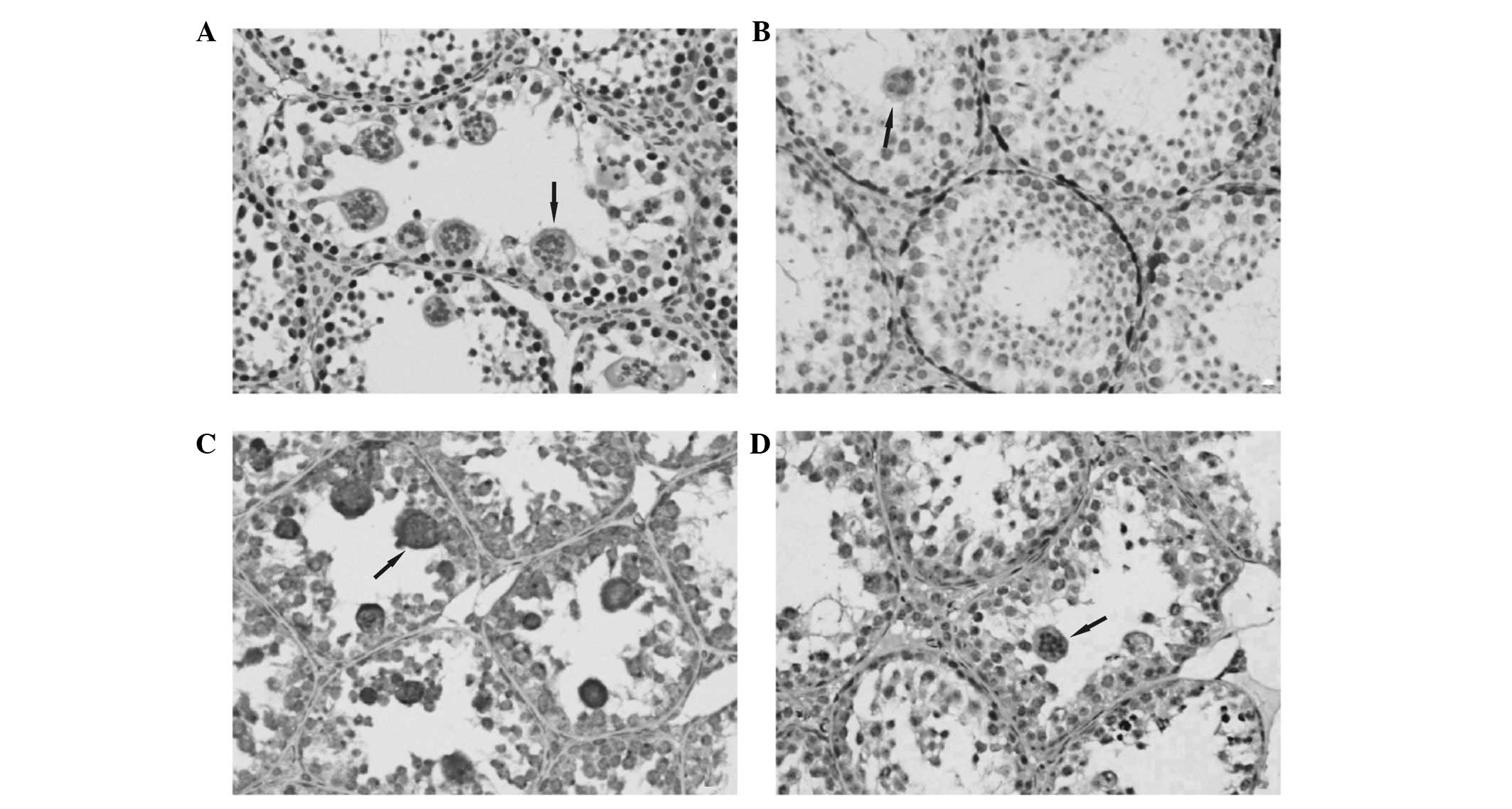
PCNA protein immunopositive signal was detected in
all seminiferous tubules in postnatal day 30 and 33 mouse testis.
PCNA-positive cells were found on the first and second layers of
the seminiferous tubules wall. Combined with the results of H&E
staining and judged by cell morphology, the positive cells were
reported to be spermatogonia and primary spermatocytes. No positive
signals were identified in multinucleated cells (Fig. 2A).
Cyclin D1 protein immunopositive signal was detected
in a small number of seminiferous tubules of day 30 and 33 mouse
testis. Cyclin D1-positive cells in the seminiferous tubules were
located in the first layer of the wall and the positive cells were
part of spermatogonial cells. No positive signals were found in
multinucleated cells (Fig.
2B).
A marked immunopositive signal of Bax protein was
detected in all seminiferous tubules of day 30 and 33 mouse testis.
Bax-positive cells located between layers 1 and 6 had strong
immunoreactive signals and were also found in multinucleated cells.
The positive cells may be spermatogonia, primary spermatocytes,
secondary spermatocytes, round spermatids and multinucleated cells
(Fig. 2C).
Caspase 3 protein immunoreactive signal was detected
in a small number of seminiferous tubules in day 30 and 33 mouse
testis. Caspase 3-positive cells were scattered in each layer of
the seminiferous tubules and a marked immunoreactive signal was
detected in multinucleated cells. Cell morphology revealed that the
positive cells were multinucleated cells, spermatogonia, primary
spermatocytes, secondary spermatocytes and round spermatids
(Fig. 2D).
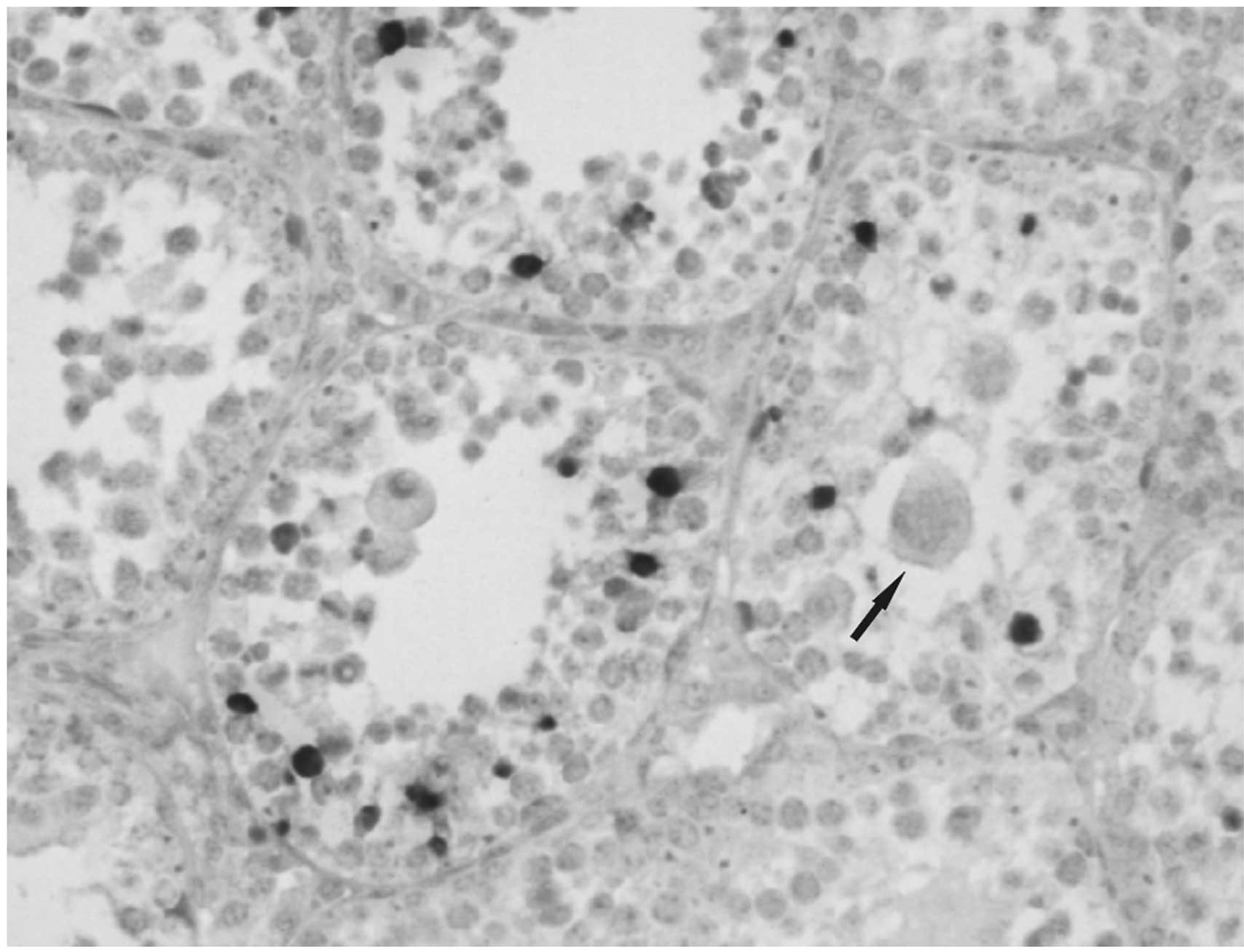
In situ cell apoptosis detection
In day 33 testicular tissue, spermatogonia, primary
spermatocytes, secondary spermatocytes and sperm cells exhibited a
strong apoptotic signal and were detected in seminiferous tubules.
Multinucleated cells did not reveal a positive apoptotic signal
(Fig. 3).
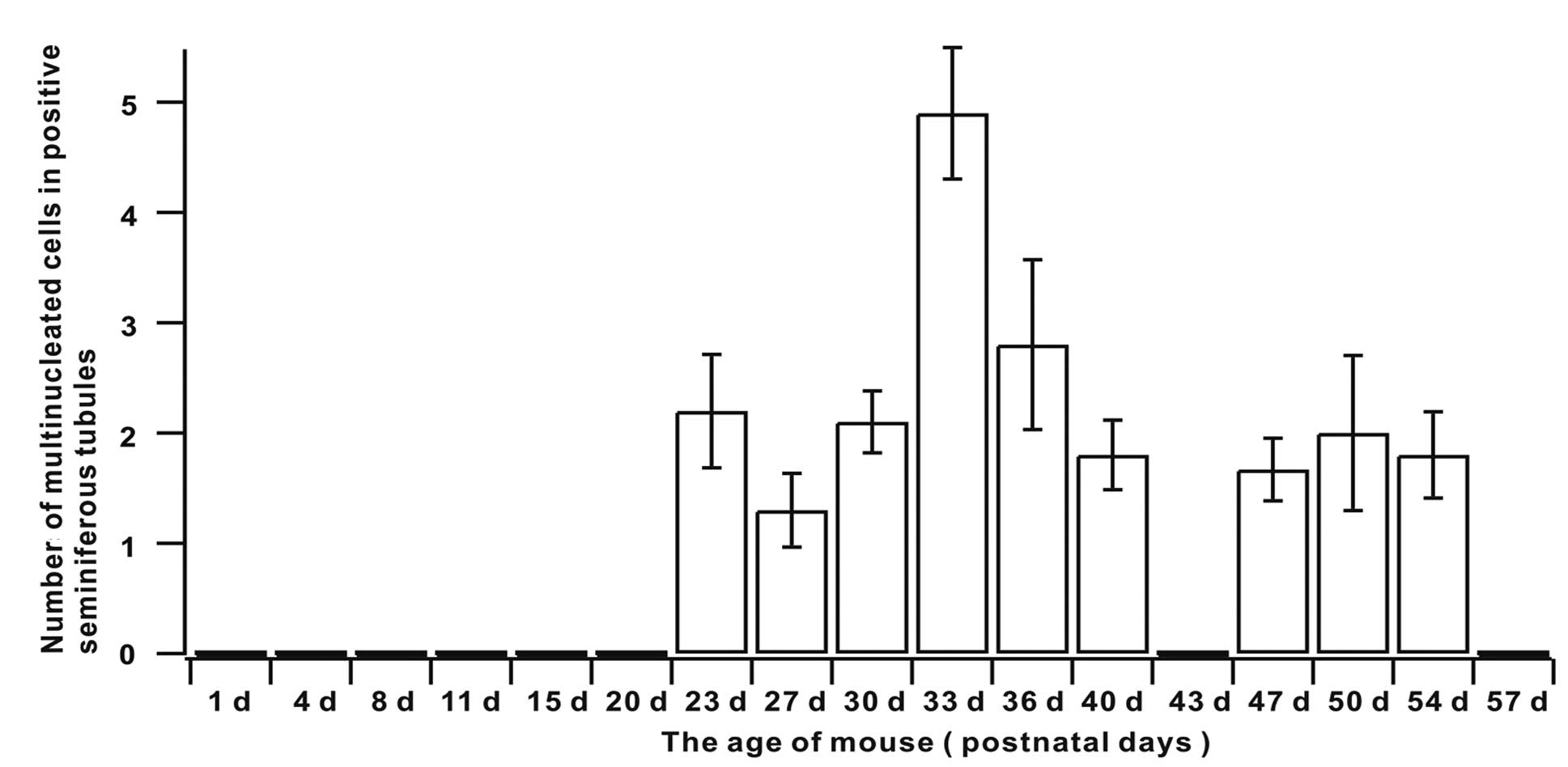
Quantification of multinucleated
cells
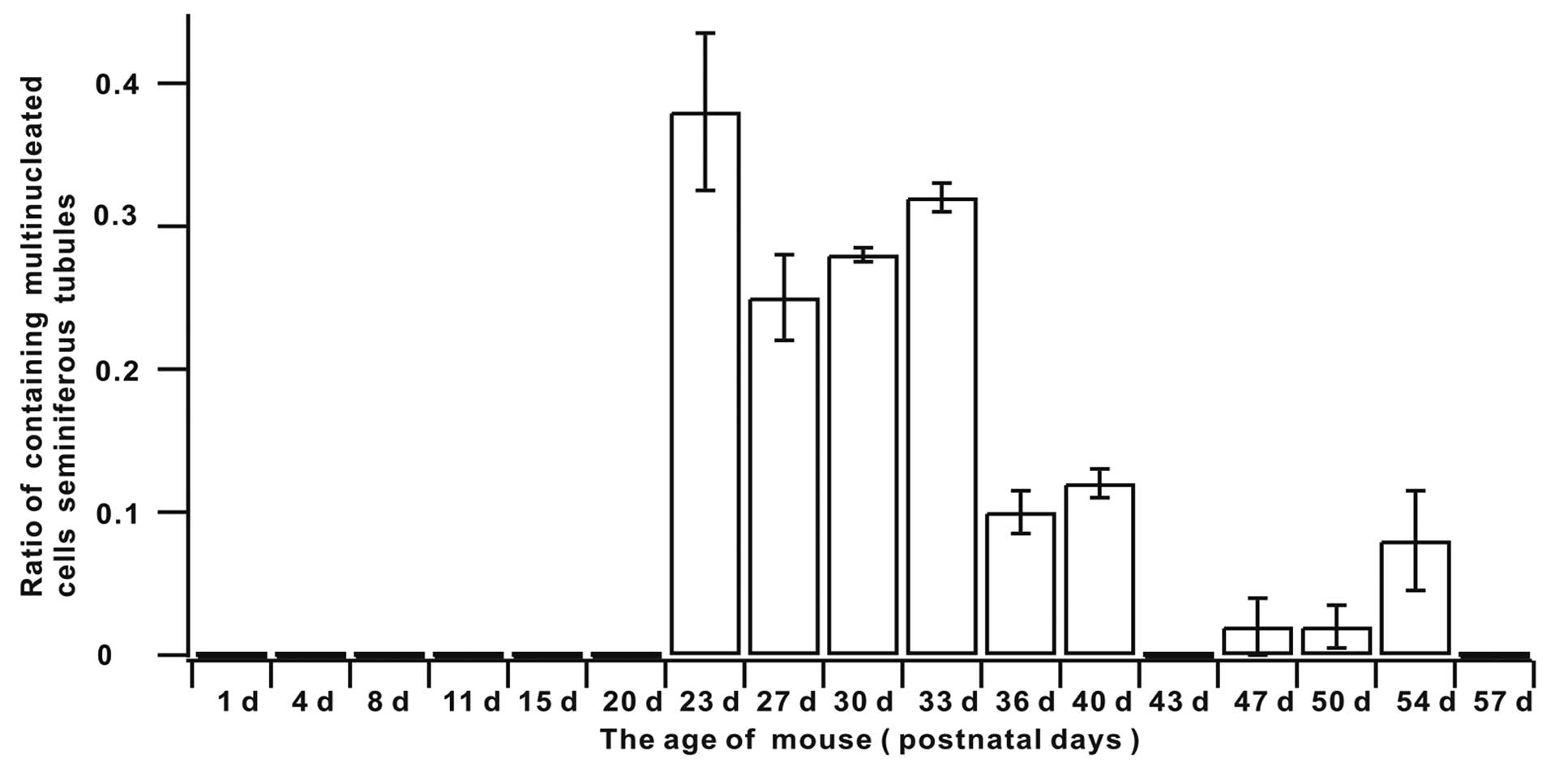
The ratio of the seminiferous tubules containing
multinucleated cells in mouse testis indicated that the rates from
days 1 to 20 were 0 (Table I and
Fig. 4). The appearance of
multinucleated cells in mouse testis seminiferous tubules began at
day 23 and the rate was maintained at an elevated level between
days 23 and 33. It began to decline at day 36 and remained at a
lower level in the cell phases that followed (Table I and Fig. 4). Quantification of multinucleated
cells in the seminiferous tubules indicated that multinucleated
cell numbers peaked at day 33 and were maintained at a relatively
stable state in the remaining phases (Table I and Fig. 5).
 | Table IQuantification of multinucleated cells
of mouse testis from various age groups. |
Table I
Quantification of multinucleated cells
of mouse testis from various age groups.
| Age of the
mouse(postnatal days) | Ratio of seminiferous
tubules containing multinucleated cells (mean ± SEM) | Ratio of number of
multinucleated cells in seminiferous tubules (mean ± SEM) |
|---|
| 1 | 0 | 0 |
| 4 | 0 | 0 |
| 8 | 0 | 0 |
| 11 | 0 | 0 |
| 15 | 0 | 0 |
| 20 | 0 | 0 |
| 23 | 0.38±0.055 | 2.20±0.515 |
| 27 | 0.25±0.030 | 1.30±0.335 |
| 30 | 0.28±0.005 | 2.10±0.280 |
| 33 | 0.32±0.010 | 4.90±0.595 |
| 36 | 0.10±0.015 | 2.80±0.770 |
| 40 | 0.12±0.010 | 1.80±0.315 |
| 43 | 0 | 0 |
| 47 | 0.02±0.020 | 1.67±0.285 |
| 50 | 0.02±0.015 | 2.00±0.705 |
| 54 | 0.08±0.035 | 1.80±0.390 |
| 57 | 0 | 0 |
Discussion
The current study evaluated multinucleated cell
appearance and potential roles during normal testicular
development. Multinucleated cells have been described as being
present primarily in pathological processes (10). To the best of our knowledge, the
present study has, for the first time, shown that during normal
development of the mouse testis, multinucleated cells appeared in
sections of the seminiferous tubules at postnatal days 23, 27, 30,
33, 36, 40, 47, 50 and 54, suggesting that multinucleated cells
exist in various stages of the normal developmental process in
mouse testis. Given this phenomenon, it is hypothesized that
multinucleated cells appear in pathological processes and in normal
testicular development.
Multinucleated cells were identified between
postnatal days 23 and 33 and peaked at day 33. The first
development and maturation time of the Kunming mouse spermatogenic
cell occurs ~5-weeks following birth (11), suggesting that the peak of the
multinucleated cells occurred prior to the first maturity period of
the sperm cell. Multinucleated cells were first observed at
postnatal day 23 in the center of the seminiferous tube cavity with
similar nucleus size as secondary spermatocytes and spermatids. Our
previous study revealed that secondary spermatocytes and a small
number of round spermatids may first be observed on postnatal day
23 in the seminiferous tubules of the mouse (12). Therefore, given the time, location
and nuclear morphology, these multinucleated cells may be derived
from the secondary spermatocytes and spermatids.
Following this, the potential role of multinucleated
cells during mouse testicular development was investigated.
Spermatogenesis is the process by which SSCs self-renew and
differentiate into sperm. The role of multinucleated cells in SSC
proliferation and differentiation was analyzed. During the cell
cycle, cyclin D1 combines with cyclin-dependent kinase (CDK) 4 or
CDK6 to phosphorylate downstream retinoblastoma protein (RB) and
its associated proteins, P107 and P130. Phosphorylated RB may
relieve inhibition of the E2F transcription factor and initiate DNA
replication, driving the cell cycle from G1 to S phase (13,14).
In the present study, cyclin D1 was expressed in the spermatogonia.
Expression of cyclin D1 during active cell cycle is consistent with
its role in promoting the transition of the cell cycle from G1 to S
phase.
PCNA is a cofactor in eukaryotic DNA polymerase δ,
which plays a role in the S phase of DNA synthesis and repair and
indicates cell proliferation (15). In the current study, PCNA was
expressed in spermatogonia and primary spermatocytes, consistent
with a previous study by Zhang et al which demonstrated that
PCNA plays a role in spermatogonia and primary spermatocytes of the
DNA replication process (16).
Immunostaining revealed no positive immune signals
in multinucleated cells, indicating that cyclin D1 and PCNA was not
expressed in these cells. The observations indicated no
physiological and biochemical activities of the G1 or S-phase
appearing within the cells with no DNA replication occurring.
Therefore, it was hypothesized that multinucleated cells are not
involved in the cell cycle and the cell proliferation process.
In typical spermatogenesis, spermatids undergo
proliferation and degradation, the latter is achieved mainly
through apoptosis. Apoptosis of spermatogenic cells ensures the
quantity and quality of spermatogenic cell survival (17).
Bax is an apoptosis-inducing factor, mainly located
in the cytoplasm when apoptosis occurs. Bax inserts into the
mitochondrial membrane causing cytochrome c release from the
cytoplasm which induces apoptosis in spermatogenic cells (18,19).
Immunostaining revealed that the Bax protein immunoreactive signal
was detected in specific spermatogonia, primary spermatocytes,
secondary spermatocytes and round spermatids. This is consistent
with a previous study by Rodriguez et al which revealed that
prior to normal mouse testis maturation, a temporary elevated
expression stage of the Bax gene was noted (20). Findings of a study by Beumer et
al showed that all cell types expressed Bax protein in mouse
testes (21).
Caspase 3 is a important apoptosis effector in the
middle and later phases of apoptosis (22,23).
Immunostaining results of the present study revealed that caspase 3
was expressed in individual spermatogonia, primary spermatocytes,
secondary spermatocytes and round spermatids. In situ
apoptosis detection indicated that a small number of spermatogenic
cells appear to undergo apoptosis. This observation confirms that,
compared with Bax, caspase 3 expression is consistent with
apoptosis of spermatogenic cells in terms of time and space.
Immunostaining also revealed immunoreactive Bax and
caspase 3 signals in the nuclei of multinucleated cells, indicating
that Bax and caspase 3 are involved in the formation and apoptosis
of multinucleated cells. Thus, multinucleated cells may be
associated with apoptotic processes.
However, the observations from in situ
apoptosis detection, performed during the same period, failed to
produce a positive apoptotic signal in multinucleated cells,
indicating that apoptotic events did not arise in multinucleated
cells. Further investigation is required to determine whether
apoptosis occurs later following the expression of
apoptosis-associated genes or whether expression of these genes
does not result in apoptosis in these cells.
In summary, this study has to the best of our
knowledge demonstrated, for the first time, that multinucleated
cells are present during normal testicular development and may be
associated with SSC apoptosis. Therefore, multinucleated cells may
be important in the spermatogenesis process.
Acknowledgements
This study was supported by grants from the Applied
Basic Research Joint Foundation of Yunnan Provincial Science and
Technology department and Kunming medical university (Study on
mechanism of formation and clearance of multinucleated cells in
development in mouse testes, 2013, for Lan Luo), Research
Foundation of the Department of Education of Yunnan Province
(no.2011Y165), National Natural Science Foundation of China
(31260264, 81060034, 81060126 and 31260243). The study was also
supported by Chunhui Project (Z2011047), State Education Ministry
(SEM) and Program for New Century Excellent Talents in University
‘NCET’ for Dr Yin Wang.
References
|
1
|
Johnston DS, Wright WW, Dicandeloro P,
Wilston E, Kopf GS and Jelinsky SA: Stage-specific gene expression
is a fundamental characteristic of rat spermatogenic cells and
Sertoli cells. Proc Natl Acad Sci USA. 105:8315–8320. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kokkinaki M, Lee TL, He ZP, Jiang JJ,
Golestaneh N, Hofmann MC, Chan WY and Dym M: Age affects gene
expression in mouse spermatogonial stem/progenitor cells.
Reproduction. 139:1011–1020. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
D’Souza UJ: Tamoxifen induced
multinucleated cells (symplasts) and distortion of seminiferous
tubules in rat testis. Asian J Androl. 5:217–220. 2003.PubMed/NCBI
|
|
4
|
Boekelheide K, Kleymenova E, Liu KJ,
Swanson C and Gaido KW: Dose-dependent effects on cell
proliferation, seminiferous tubules and male germ cells in the
fetal rat testis following exposure to di(n-butyl) phthalate.
Microsc Res Tech. 72:629–638. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kondarewicz A, Kolasa A, Zawiślak B,
Baranowska-Bosiacka I, Marchlewicz M, Wenda-Różewicka L and
Wiszniewska B: Testis morphology in rats chronically treated with
letrozole, an aromatase inhibitor. Folia Histochem Cytobiol.
49:677–684. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramzan F and Qureshi IZ: Intraperitoneal
kisspeptin-10 administration induces dose-dependent degenerative
changes in maturing rat testes. Life Sci. 88:246–256. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Anton E: Arrested apoptosis without
nuclear fragmentation produced by efferent duct ligation in round
spermatids and multinucleated giant cells of rat testis.
Reproduction. 125:879–887. 2003. View Article : Google Scholar
|
|
8
|
Schofield JB and Evans DJ: Multinucleate
giant stromal cells in testicular atrophy following oestrogen
therapy. Histopathology. 16:200–201. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Coyne JD and Dervan PA: Multinucleated
stromal giant cells of testis. Histopathology. 31:381–383. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pitt MA, Roberts IS, Agbamu DA and Eyden
BP: The nature of atypical multinucleated stromal cells: a study of
37 cases from different sites. Histopathology. 23:137–145. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu J, Wan HJ, Cai ZM, Fang JZ, Zhang FT,
Ye J and Yin MJ: The histological observation of the stepwise
development of neonatal mouse testis in immunodeficient mice. Jie
Pou Xue Bao. 38:213–217. 2007.(In Chinese).
|
|
12
|
Luo L, Zhang Y and Yang F: The
histological observation in the testis of mice at different
development stage. Laboratory Animal Science. 27:10–13. 2010.(In
Chinese).
|
|
13
|
Beumer TL, Roepers-Gajadien HL, Gademan
IS, Kal HB and de Rooij DG: Involvement of the D-type cyclins in
germ cell proliferation and differentiation in the mouse. Biol
Reprod. 63:1893–1898. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baker GL, Landis MW and Hinds PW: Multiple
functions of D-type cyclins can antagonize pRb-mediated suppression
of proliferation. Cell Cycle. 4:330–338. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zusman I, Reifen R, Livni O, Smirnoff P,
Gurevich P, Sandler B, Nyska A, Gal R, Tendler Y and Madar Z: Role
of apoptosis, proliferating cell nuclear antigen protein in
chemically induced colon cancer in rats fed corncob fiber treated
with the fungus Pleurotus ostreatus. Anticancer Res.
17:2105–2113. 1997.PubMed/NCBI
|
|
16
|
Zhang J, Gao FL, Zhi HY and Duan XG:
Age-specific changes of proliferation and apoptosis in the
seminiferous epithelium of laboratory mice. Dong Wu Xue Bao.
47:209–214. 2001.(In Chinese).
|
|
17
|
de Rooij DG: Stem cells in the testis. Int
J Exp Pathol. 79:67–80. 1998.
|
|
18
|
Eskes R, Desagher S, Antonsson B and
Martinou BA: Bid induces the oligomerization and insertion of Bax
into outer mitochondrial membrane. Mol Cell Biol. 20:929–935. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Antonsson B, Montessuit S, Sanchez B and
Martinou JC: Bax is present as a high molecular weight
oligomer/complex in the mitochondrial membrane of apoptotic cells.
J Biol Chem. 276:11615–11623. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rodriguez I, Ody C, Araki K, Garcia I and
Vassalli P: An early and massive wave of germinal cell apoptosis is
required for the development of functional spermatogenesis. EMBO J.
16:2262–2270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beumer TL, Roepers-Gajadien HL, Gademan
IS, Lock TM, Kal HB and de Rooij DG: Apoptosis regulation in the
testis: involvement of Bcl-2 family members. Mol Reprod Dev.
56:353–359. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Almeida C, Cunha M, Ferraz L, Silva J,
Barros A and Sousa M: Caspase-3 detection in human testicular
spermatozoa from azoospermic and non-azoospermic patients. Inter J
Androl. 34:e407–e414. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Philchenkov AA: Caspases as regulators of
apoptosis and other cell functions. Biochemistry (Mosc).
68:365–376. 2003. View Article : Google Scholar : PubMed/NCBI
|