Introduction
Metabolic syndrome caused by insulin resistance and
abnormal adipose tissue deposition is associated with various risk
factors, including coronary artery disease, stroke, fatty liver
development and type 2 diabetes (1,2).
This syndrome is also known as cardiometabolic syndrome, metabolic
syndrome X, syndrome X, insulin resistance syndrome and Reaven’s
syndrome. The majority of patients with this disorder are older,
obese and insulin-resistant, and have a sedentary lifestyle
(3). Clinical symptoms of
metabolic syndrome include hypertension, hyperglycemia,
hypertriglyceridemia, decreased high-density lipoprotein levels,
increased triglyceride levels and abdominal obesity (4,5);
however, the etiology of this syndrome is not yet clearly
understood.
Numerous mouse models, established through
spontaneous mutations or genetic modification, have been employed
to investigate metabolic syndrome. Leptin-deficient
[Lep(ob/ob)] (6),
leptin receptor-deficient [LepR(db/db)] (7), lethal yellow agouti (Ay/a)
(8), melanocortin 3
receptor-deficient (9),
melanocortin 4 receptor-deficient (10), low-density lipoprotein
receptor-deficient (11) and
apolipoprotein E-deficient mice have been generated (12,13).
These animal models develop clinical symptoms similar to those
observed in humans, including increased obesity and insulin
resistance (14,15).
11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1)
is an NADPH-dependent enzyme that is highly expressed in key
metabolic tissues, including liver, adipose tissue and the central
nervous system (16). In metabolic
tissue, including liver and adipose tissue, 11β-HSD1 converts
cortisone into the active hormone, cortisol (17,18).
11β-HSD1 transgenic mice develop abdominal obesity, hyperglycemia,
insulin resistance, hyperphagia, hyperleptinemia and increased
intra-adipose and portal levels, but not systemic corticosterone
levels (19). In addition, the
adipose tissue of obese humans has been observed to exhibit
elevated 11β-HSD1 activity (20).
Genetically modified mice are an attractive tool as
they are small in size, easy to handle and possess short generation
intervals. However, the phenotypic, genetic, physiological and
anatomical characteristics of mice are not similar to humans. As
pigs are physiologically similar to humans (21), in the present study, genetically
modified porcine transgenic fibroblasts were produced to induce
metabolic syndrome-like symptoms in a porcine model. An 11β-HSD1
cDNA construct conjugated to the pig adipose fatty acid-binding
protein (aP2) promoter was generated to facilitate 11β-HSD1 gene
expression in porcine adipose tissue. These transgenic fibroblasts
may serve as a source for somatic cell nuclear transfer (SCNT) to
produce a porcine model of metabolic syndrome.
Materials and methods
Cell culture
Mouse embryonic 3T3-L1 fibroblast adipose-like cells
were purchased from the Korean Cell Line Bank (Seoul, South Korea).
3T3-L1 preadipocytes were incubated, unless otherwise indicated, at
37°C and, unless otherwise indicated, all cell culture materials
were obtained from Invitrogen Life Technologies (Carlsbad, CA,
USA). On day 30 of pregnancy, porcine fibroblasts were obtained
from a miniature pig fetus (Yucatan pig; Optifarm Solution Inc.,
Gyeonggi-do, South Korea). Fibroblasts were cultured in Dulbecco’s
modified Eagle’s medium (DMEM) containing 10% fetal bovine serum
(FBS; WelGENE, Daegu, South Korea), 50 U/ml penicillin and 50 μg/ml
streptomycin in a humidified 5% CO2 atmosphere.
Genomic DNA extraction and PCR
Genomic DNA was isolated from the fibroblasts using
a G-DEX™ IIc genomic DNA extraction kit (Intron Biotechnology,
Seoul, South Korea). All experimental procedures and animal use
were approved by the Ethics Committee of the Chungbuk National
University. Genomic DNA (1 μg) was amplified in a 20 μl PCR
containing 1 unit LA-Taq polymerase (Takara Bio, Inc.,
Shiga, Japan) for long-range PCR or 1 unit i-star Taq
polymerase (Intron Biotechnology), 2 mM dNTPs (Takara Bio, Inc.)
and 10 pmol each specific primer. All primers are presented in
Table I. PCR conditions were as
follows: denaturation at 95°C for 30 sec, annealing at 62°C for 30
sec and extension at 72°C for 1 or 2 min. PCR products were
separated on a 0.7% agarose gel, stained with ethidium bromide,
imaged under UV illumination and processed for cloning. The gel
image was scanned using Gel Doc EQ (Bio-Rad, Hercules, CA,
USA).
 | Table IPrimer sequences and restriction
enzymes. |
Table I
Primer sequences and restriction
enzymes.
| Name | Restriction
enzyme | Direction | Sequences (5′ to
-3′) |
|---|
| Pig aP2 promoter
(−4,424) | MluI | Forward |
ACGCGTTATGGGAAGTATGTTTTGGA |
| Pig aP2 promoter
(−2,826) | MluI | Forward |
ACGCGTGGACTTTAATGGACACCTCACC |
| Pig aP2 promoter
(−658) | MluI | Forward |
ACGCGTTACAACCCAACAGCAAAA AAGCC |
| Pig aP2 promoter
(+51) | XhoI | Reverse |
CTCGAGCCTTCAGGAAGGTGCAATGAC |
| Pig 11β-HSD cDNA | BglII | Forward |
AGATCTATGGCTTTTATGAAAAAATATCTCCTCC |
| Pig 11β-HSD cDNA | XbaI | Reverse |
TCTAGACTAGTTGTTTGTAAACCTTTCCATATTA |
| EGFP cDNA | EcoRV | Forward |
GATATCCACAACCATGGTGAGCAAGGGCGA |
| EGFP cDNA | BamHI | Reverse |
GGATCCTTACTTGTACAGCTCGTCCATGCC |
| Confirming primer
a | | Forward |
CCATGATAATAAGCCTGCTCTACTCCA |
| Confirming primer
b | | Reverse |
GGAAGTCATGAAGGCCTGGGTGATG |
| Confirming primer
c | | Forward |
CATGAAGCAGCACGACTTCT |
| Confirming primer
d | | Reverse |
CCTAGGAATGCTCGTCAAGA |
| 1A | | Forward |
CCAGGATTTGGAATTATTTC |
| 1A | | Reverse |
GAAAATAAAGCCTAAGGCTC |
| Pig 11β-HSD | | Forward |
CAACGTGTCAATCACGCTCT |
| Pig 11β-HSD | | Reverse |
TTCCTGGATTTTCCAACAGG |
RNA preparation and semi-quantitative
PCR
Total RNA was extracted from 3T3-L1 cells or pig
liver (Yucatan pig; Optifarm Solution Inc.) using TRIzol reagent
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. Total RNA concentration was determined by measuring
absorbance at 260 nm (Epoch microplate spectrophotometer; BioTek,
Winooski, VT, USA). First strand cDNA was prepared by subjecting
total RNA (1 μg) to reverse transcription using M-MLV reverse
transcriptase (Invitrogen Life Technologies) and random primers
(9-mers; Takara Bio Inc.). Optimal conditions for logarithmic phase
PCR amplification of the target cDNA were determined by amplifying
aliquots of total cDNA (1 μg) using a different number of cycles.
The cytochrome c oxidase subunit 1 (1A) gene was used as an
internal control to eliminate the possibility of RNA degradation
and to account for variation in mRNA concentration. A linear
correlation between the PCR product band visibility and the number
of amplification cycles was observed in the target mRNA products.
The 1A gene and target gene, insulin, were quantified using 28 and
30 cycles, respectively. PCR conditions were as follows:
denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec and
extension at 72°C for 30 sec. PCR products were separated in a 2.3%
agarose gel, stained with ethidium bromide and imaged under UV
illumination. The image was scanned and band densities were
analyzed using Gel Doc EQ (Bio-Rad).
Vector construction
Restriction enzymes were obtained from Takara Bio,
Inc. Specific regions of the aP2 promoter were prepared by
long-range PCR using porcine genomic DNA as a template and specific
primers containing restriction enzyme sites (MluI at 5′ end
or BglII at 3′ end). Amplified fragments were digested with
MluI and BglII and ligated into the promoterless
pGL3-Basic luciferase expression plasmid (Promega Corporation,
Madison, WI, USA). The 11β-HSD1 expression cassette plasmid was
produced in several steps. 11β-HSD1 cDNA was prepared by PCR using
cDNA from pig liver as a template. The amplified fragments were
inserted into the recombinant pGL3 construct containing the pig aP2
promoter region (−2,826 to +51 nt). For the selection cassettes,
the enhanced green fluorescent protein (EGFP) gene was amplified
from the pIRES2-EGFP plasmid (Clontech Laboratories Inc., Mountain
View, CA, USA), digested with EcoRV and BamHI and
inserted into the pIRESneo plasmid (Clontech Laboratories Inc.).
The EGFP and neomycin-resistant (Neor) genes were amplified by PCR,
digested with SalI and ligated into the recombinant pGL3
vector encoding pig 11β-HSD1 cDNA controlled by the porcine aP2
promoter. Finally, sequences of the targeting vector were confirmed
by nucleotide sequencing (Genotech Corporation, Daejeon, South
Korea).
Transient transfection and reporter gene
assay
Transient transfection was performed using
Lipofectamine™ 2000 (Invitrogen Life Technologies) according to the
manufacturer’s instructions. To account for varied transfection
efficiencies of the various luciferase constructs, the Rous sarcoma
virus (RSV)-lacZ plasmid was co-transfected with the luciferase
constructs containing the pig aP2 promoter, as previously described
(22,23). Briefly, 3×105 cells were
seeded in 6-well tissue culture plates (Invitrogen Life
Technologies) 1 d prior to transfection. Constructs (4 μg)
containing the aP2 promoter and RSV-lacZ plasmid (0.5 μg) were
co-transfected into the cells with DMEM. Following an incubation
period of 4 h, the media was replaced with DMEM containing 10% FBS
and glucose (0.1 or 4 mM) and the cells were incubated for an
additional 48 h at 37°C. Cell lysates were assayed for luciferase
activity using a luciferase assay system (Promega Corporation).
Luminescence was measured using a GloMax 20/20 Luminometer (Promega
Corporation). β-galactosidase activity was measured using a
β-galactosidase enzyme assay system (Promega Corporation). Relative
luciferase activity was calculated as luciferase
activity/β-galactosidase activity (%).
Establishment of transgenic cell
lines
Porcine fibroblasts were transfected with the
linearized targeting vector using Lipofectamine 2000. Following 24
h transfection, the medium was replaced with DMEM supplemented with
10% FBS and 250 μg/ml G-418 (Roche Diagnostics, Indianapolis, IN,
USA) for 4 weeks. Antibiotic-resistant colonies were further
selected according to EGFP expression observed with a fluorescence
microscope (Nikon, Tokyo, Japan). Antibiotic and visually selected
colonies were subjected to PCR-based genotyping and stored until
required for SCNT.
Statistical analysis
Data are presented as the mean ± SEM. A statistical
analysis was performed by Student’s t-test for two-pair
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Functional analysis of the pig aP2
promoter
Prior to the generation of the 11β-HSD1 cDNA
construct, a pig aP2 promoter that was hypothesized to be activated
in adipocytes was produced. The aP2 gene is typically expressed in
adipose tissue and the maximum promoter activity of aP2 in the
adipose tissue was previously confirmed (24). A series of pig aP2 gene promoters
with sequential deletions were inserted into the luciferase (Luc)
gene in a promoterless pGL3-basic vector. The aP2 promoter
constructs were introduced into the mouse embryonic
fibroblast-adipose 3T3-L1 cells and the promoter activities were
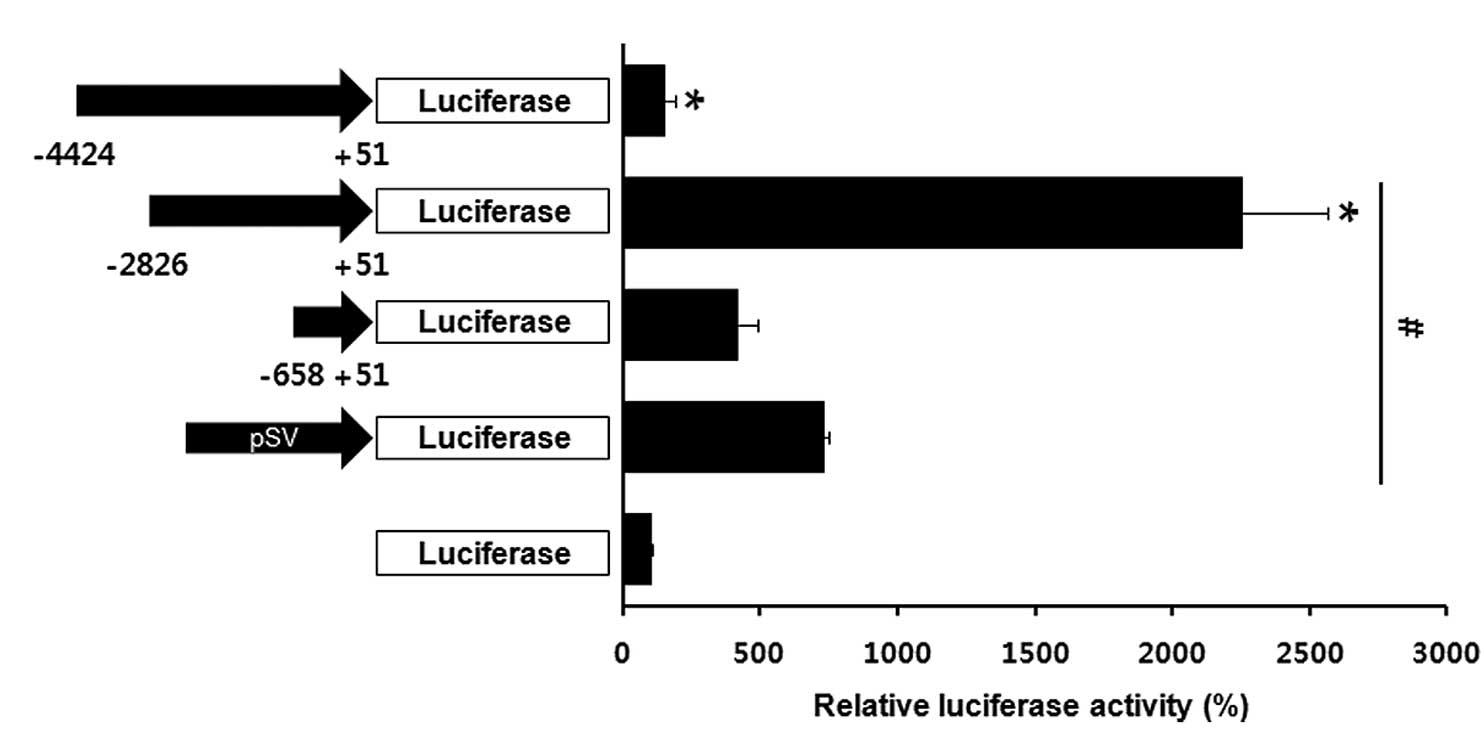
evaluated according to Luc expression. As presented in Fig. 1, the promoter region between −2,826
and +15 nt resulted in an ~20-fold increase of Luc activity
compared with the promoterless construct. These observations
indicate that −2,826 to +15 nt of the aP2 promoter was the best
candidate adipocyte-specific expression promoter.
Overexpression of recombinant pig
11β-HSD1 in 3T3-L1 cells
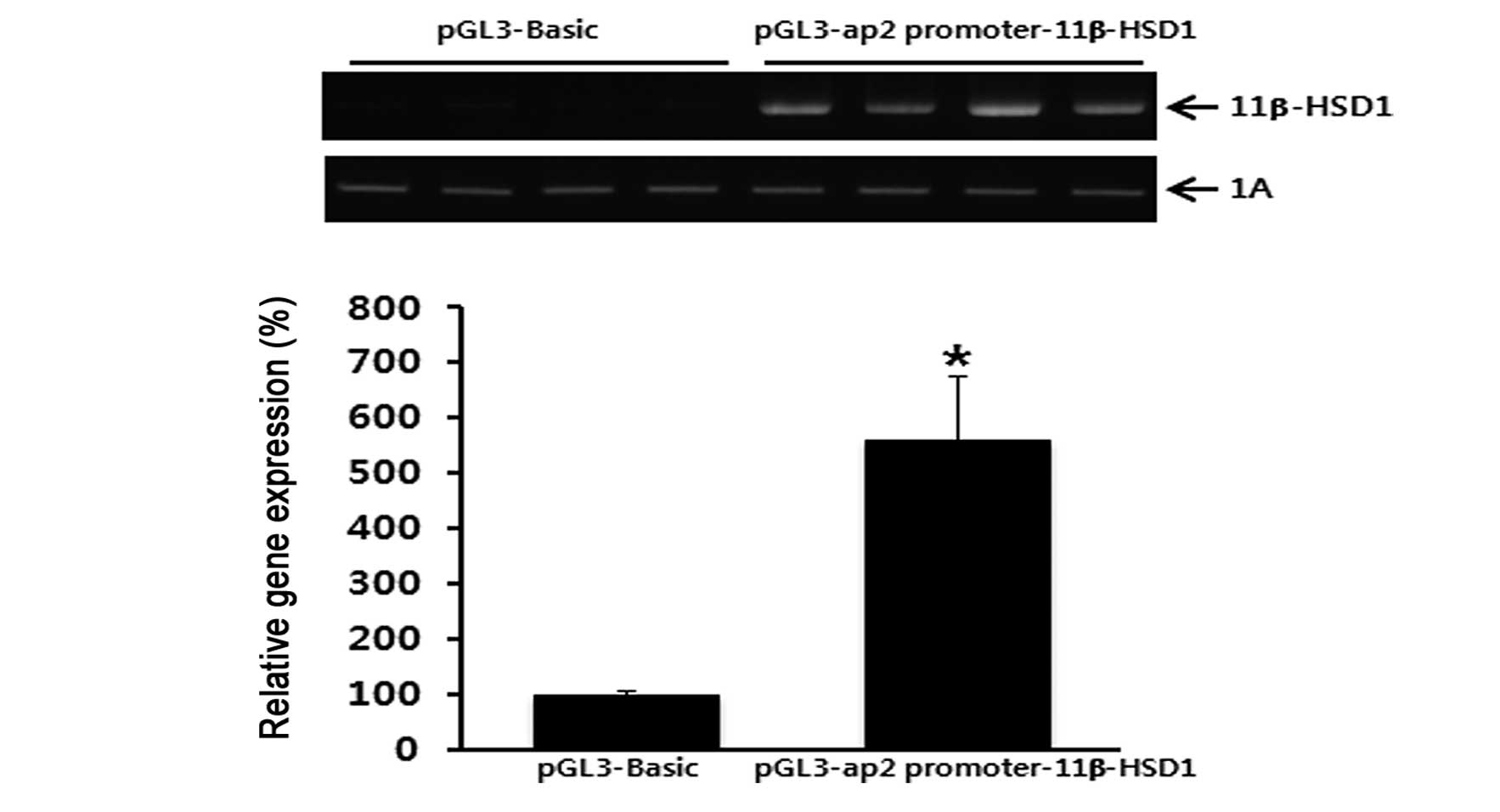
Following verification that the aP2 promoter
construct containing −2,826 to +15 nt had maximal activity, the
construct was subcloned with porcine 11β-HSD1 cDNA. The resulting
aP2/11β-HSD1 construct was capable of expressing 11β-HSD1 mRNA in
an adipocyte-specific manner. 11β-HSD1 mRNA levels increased in the
transiently-transfected 3T3-L1 cells (Fig. 2). These observations indicate that
the recombinant aP2/11β-HSD1 construct was appropriate for
expressing pig 11β-HSD1 in an adipose tissue-specific manner.
Establishment of porcine fibroblast cell
lines that overexpress 11β-HSD1
Our constructs were composed of two parts, an
11β-HSD1 expression cassette and a selection cassette (Fig. 3A). The 11β-HSD1 expression cassette
contained pig 11β-HSD1 cDNA with the porcine aP2 promoter region
(−2,826 to +15 nt). The selection cassette contained EGFP and Neor
genes, whose expression was regulated by a cytomegalic virus
promoter. Expression of the EGFP and Neor genes enabled selection
of the cells of interest using antibiotics and visual
screening.
The targeting construct was linearized and used to
transfect porcine fibroblasts isolated from Yucatan pigs.
Fibroblasts were incubated in medium containing G-418 (250 μg/ml)
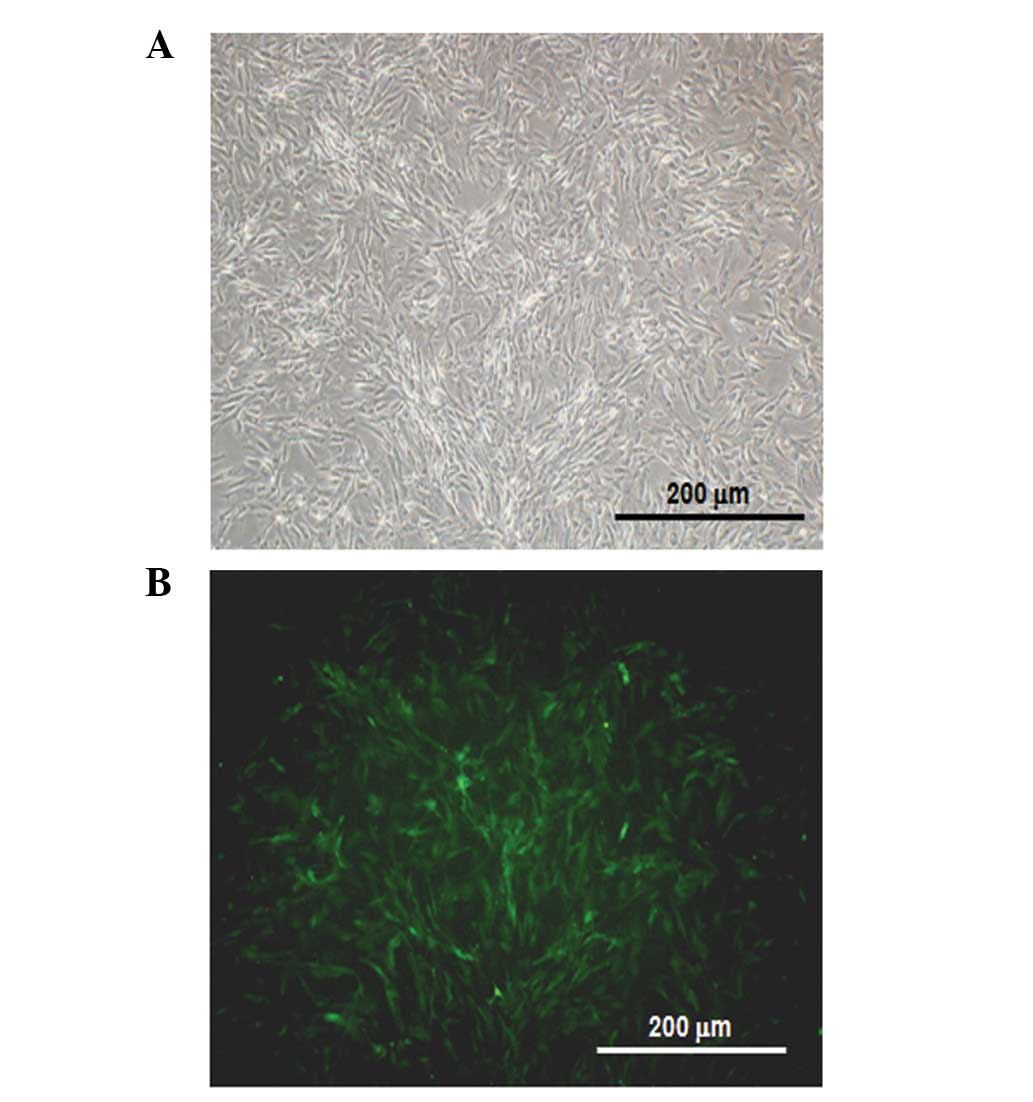
for 4 weeks to select for the stable transfectants. Identification
of Neor colonies as transgenic fibroblasts was confirmed by EGFP
expression (Fig. 4). The clones
were further identified by PCR-based methods using primers specific
for the targeting constructs. Genomic DNA from clones expressing
porcine 11β-HSD1 was analyzed with an a-b primer set (amplicon
size, 740 bp; Fig. 3B) or with c-d
primers (amplicon size, 1 kb; Fig.
3C). Transfection efficiency was ~89% (Table II). These observations indicate
that the cell model may be useful for establishing a transgenic
fibroblast system for creating porcine metabolic syndrome models by
expressing adipose tissue-specific 11β-HSD1.
| Table IITransfection efficiencies of the pig
fibroblasts. |
Table II
Transfection efficiencies of the pig
fibroblasts.
| Transfection
trials, n | G418-resistant
colonies, n | EGFP-positive
colonies, n | PCR-positive
colonies, n |
|---|
| 11 | 37 | 33 | 33 |
Discussion
Tissue-specific promoters generally contain
cis-acting elements to which tissue-specific transcription
factors bind and regulate tissue-specific expression of genes.
Fatty acid-binding proteins are an intracellular hydrophobic
ligand-binding protein family (25). aP2 (also known as fatty
acid-binding protein 4) facilitates the intracellular
solubilization and trafficking of lipids (26). This factor has been implicated as a
strong candidate for adipose-specific genes representing the
accumulation of lipids in pigs and cows (27,28).
Previously, the aP2 promoter in humans and mice was analyzed to
identify fat-specific regulatory regions (29). This promoter contains conserved
transcription factor binding sites, including C/EBP, adapter
primer-1, CAAT box, TATA box, direct repeat 1-type PPAR responsive
element, short interspersed repetitive elements and another PPAR
responsive element in humans, mice, cows, pigs and dogs (24). The −5.4 kb region upstream of the
aP2 promoter is hypothesized to direct the adipose-specific
expression of transgenes in mice (30). The aP2 promoter was utilized to
overexpress 11β-HSD1 in adipose tissue. Activity of the porcine aP2
promoter in the 3T3-L1 adipose cells was examined. A specific
portion of the porcine aP2 promoter (between −2,826 and +15 nt) was
observed to possess the highest activity among the various tested
promoter regions.
The 11β-HSD1 gene is highly expressed in the adipose
tissue of obese humans (20). In
the present study, a porcine fibroblast cell line was established
expressing the pig 11β-HSD1 gene under the control of the porcine
aP2 promoter. In addition, two selection markers, EGFP and the Neor
gene, were used for genetic information of somatic cells for
further SCNT-mediated cloning. The Neor and EGFP genes are used to
select transgenic cell colonies to increase SCNT efficiency and
identify transgenic piglets (31–33).
Therefore, the use of these porcine fibroblasts cells is likely to
enable us to achieve high somatic cell cloning efficiencies in
mammals.
Although there are several animal models of
metabolic syndrome, the majority are rodents, including mice and
rats, which are relatively dissimilar to humans. A porcine model of
metabolic syndrome has several advantages, including the
physiological similarity between pigs and humans in terms of body
fat distribution and fat cell size. Therefore, insight into
lipogenesis, lipolysis and lipid mobilization gained from
evaluating a porcine model may be useful for studying human
obesity. In addition, numerous diagnostic and surgical techniques
developed for humans are applicable in pigs, further indicating
that the pig is a valuable model for investigating various human
diseases.
In conclusion, a porcine fibroblast cell line
containing constructs suitable for 11β-HSD1 overexpression was
established. To assess adipose-specific expression of the 11β-HSD1
gene, an aP2 promoter controlling expression in adipose tissue was
included in the construct. These porcine fibroblast cell lines may
be a useful source for SCNT procedures to generate a porcine model
of metabolic syndrome. Piglets derived from these cells may also
provide critical information for understanding the pathophysiology
of metabolic syndrome and developing novel diagnostic therapies for
combating human metabolic syndrome.
Acknowledgements
This study was supported by a grant from the
Next-Generation BioGreen 21 Program (no. PJ009563), Rural
Development Administration, Republic of Korea.
Reference
|
1
|
Olufadi R and Byrne CD: Clinical and
laboratory diagnosis of the metabolic syndrome. J Clin Pathol.
61:697–706. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rask-Madsen C and Kahn CR: Tissue-specific
insulin signaling, metabolic syndrome and cardiovascular disease.
Arterioscler Thromb Vasc Biol. 32:2052–2059. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Grundy SM, Brewer HB Jr, Cleeman JI, Smith
SC Jr and Lenfant C; National Heart, Lung and Blood Institute;
American Heart Association. Definition of metabolic syndrome:
report of the National Heart, Lung and Blood Institute/American
Heart Association conference on scientific issues related to
definition. Arterioscler Thromb Vasc Biol. 24:e13–e18. 2004.
View Article : Google Scholar
|
|
4
|
Roos CJ, Quax PH and Jukema JW:
Cardiovascular metabolic syndrome: mediators involved in the
pathophysiology from obesity to coronary heart disease. Biomark
Med. 6:35–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vogelzangs N and Penninx BW: Depressive
symptoms, cortisol, visceral fat and metabolic syndrome. Tijdschr
Psychiatr. 53:613–620. 2011.(In Dutch).
|
|
6
|
Nishina PM, Lowe S, Wang J and Paigen B:
Characterization of plasma lipids in genetically obese mice: the
mutants obese, diabetes, fat, tubby and lethal yellow. Metabolism.
43:549–553. 1994. View Article : Google Scholar
|
|
7
|
Hummel KP, Dickie MM and Coleman DL:
Diabetes, a new mutation in the mouse. Science. 153:1127–1128.
1966. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tschöp M and Heiman ML: Rodent obesity
models: an overview. Exp Clin Endocrinol Diabetes. 109:307–319.
2001.
|
|
9
|
Trevaskis JL, Gawronska-Kozak B, Sutton
GM, et al: Role of adiponectin and inflammation in insulin
resistance of Mc3r and Mc4r knockout mice. Obesity (Silver Spring).
15:2664–2672. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tallam LS, Stec DE, Willis MA, da Silva AA
and Hall JE: Melanocortin-4 receptor-deficient mice are not
hypertensive or salt-sensitive despite obesity, hyperinsulinemia
and hyperleptinemia. Hypertension. 46:326–332. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ishibashi S, Brown MS, Goldstein JL,
Gerard RD, Hammer RE and Herz J: Hypercholesterolemia in low
density lipoprotein receptor knockout mice and its reversal by
adenovirus-mediated gene delivery. J Clin Invest. 92:883–893. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Plump AS, Smith JD, Hayek T, et al: Severe
hypercholesterolemia and atherosclerosis in apolipoprotein
E-deficient mice created by homologous recombination in ES cells.
Cell. 71:343–353. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang SH, Reddick RL, Piedrahita JA and
Maeda N: Spontaneous hypercholesterolemia and arterial lesions in
mice lacking apolipoprotein E. Science. 258:468–471. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kennedy AJ, Ellacott KL, King VL and Hasty
AH: Mouse models of the metabolic syndrome. Dis Model Mech.
3:156–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Asterholm IW, Halberg N and Scherer PE:
Mouse Models of Lipodystrophy Key reagents for the understanding of
the metabolic syndrome. Drug Discov Today Dis Models. 4:17–24.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Agarwal AK: Cortisol metabolism and
visceral obesity: role of 11beta-hydroxysteroid dehydrogenase type
I enzyme and reduced co-factor NADPH. Endocr Res. 29:411–418. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
White PC, Mune T and Agarwal AK: 11
beta-Hydroxysteroid dehydrogenase and the syndrome of apparent
mineralocorticoid excess. Endocr Rev. 18:135–156. 1997.PubMed/NCBI
|
|
18
|
White PC, Mune T, Rogerson FM, Kayes KM
and Agarwal AK: Molecular analysis of 11 beta-hydroxysteroid
dehydrogenase and its role in the syndrome of apparent
mineralocorticoid excess. Steroids. 62:83–88. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Masuzaki H, Paterson J, Shinyama H, et al:
A transgenic model of visceral obesity and the metabolic syndrome.
Science. 294:2166–2170. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rask E, Olsson T, Söderberg S, et al:
Tissue-specific dysregulation of cortisol metabolism in human
obesity. J Clin Endocrinol Metab. 86:1418–1421. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Groth CG: Xenotransplantation is the hope
of the future. Genetic modification of the donor, the pig, is
better than fighting organ rejection by submitting patients to
immunosuppression with frequent side effect. Lakartidningen.
99:252–254. 2002.(In Swedish).
|
|
22
|
Cheng CK, Hoo RL, Chow BK and Leung PC:
Functional cooperation between multiple regulatory elements in the
untranslated exon 1 stimulates the basal transcription of the human
GnRH-II gene. Mol Endocrinol. 17:1175–1191. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee GS, Choi KC, Han HJ and Jeung EB: The
classical and a non-classical pathways associated with NF-kappaB
are involved in estrogen-mediated regulation of calbindin-D9k gene
in rat pituitary cells. Mol Cell Endocrinol. 277:42–50. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shin J, Li B, Davis ME, Suh Y and Lee K:
Comparative analysis of fatty acid-binding protein 4 promoters:
conservation of peroxisome proliferator-activated receptor binding
sites. J Anim Sci. 87:3923–3934. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu Z, Bernlohr DA and Banaszak LJ: Crystal
structure of recombinant murine adipocyte lipid-binding protein.
Biochemistry. 31:3484–3492. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thompson BR, Mazurkiewicz-Muñoz AM,
Suttles J, Carter-Su C and Bernlohr DA: Interaction of adipocyte
fatty acid-binding protein (AFABP) and JAK2: AFABP/aP2 as a
regulator of JAK2 signaling. J Biol Chem. 284:13473–13480. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Estellé J, Perez-Enciso M, Mercadé A, et
al: Characterization of the porcine FABP5 gene and its association
with the FAT1 QTL in an Iberian by Landrace cross. Anim Genet.
37:589–591. 2006.PubMed/NCBI
|
|
28
|
Wibowo TA, Gaskins CT, Newberry RC,
Thorgaard GH, Michal JJ and Jiang Z: Genome assembly anchored QTL
map of bovine chromosome 14. Int J Biol Sci. 4:406–414. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rival Y, Stennevin A, Puech L, et al:
Human adipocyte fatty acid-binding protein (aP2) gene
promoter-driven reporter assay discriminates nonlipogenic
peroxisome proliferator-activated receptor gamma ligands. J
Pharmacol Exp Ther. 311:467–475. 2004. View Article : Google Scholar
|
|
30
|
Ross SR, Graves RA, Greenstein A, et al: A
fat-specific enhancer is the primary determinant of gene expression
for adipocyte P2 in vivo. Proc Natl Acad Sci USA. 87:9590–9594.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jung EM, Kim YK, Lee GS, Hyun SH, Hwang WS
and Jeung EB: Establishment of inducible cAMP early repressor
transgenic fibroblasts in a porcine model of human type 1 diabetes
mellitus. Mol Med Rep. 6:239–245. 2012.PubMed/NCBI
|
|
32
|
Kim YK, Lee GS, Jung EM, Hyun SH, Hwang WS
and Jeung EB: Generation of fibroblasts overexpressing
liver-specific PEPCK in a miniature pig model of human type 2
diabetes mellitus. Mol Med Rep. 6:45–50. 2012.PubMed/NCBI
|
|
33
|
Lee GS, Hyun SH, Kim HS, et al:
Improvement of a porcine somatic cell nuclear transfer technique by
optimizing donor cell and recipient oocyte preparations.
Theriogenology. 59:1949–1957. 2003. View Article : Google Scholar : PubMed/NCBI
|