Introduction
Lidocaine, which is a common local anesthetic and
antiarrhythmic drug, may lead to transient neurological symptoms
and cauda equina syndrome following spinal anesthesia (1). Despite this, spinal anesthesia with
lidocaine is a relatively widely used technique. Several studies
have demonstrated that the cytotoxicity of lidocaine is dose- and
time-dependent in rats and mice (2,3), and
it has been shown in vitro that lidocaine induces apoptosis
(4). This apoptotis-inducing
effect is due to mitochondrial injury (5). Apoptosis is controlled by caspases,
which are activated by two major signaling pathways, the extrinsic
death receptor and intrinsic mitochondrial pathways. Werdehausen
et al (6) reported that the
apoptosis induced by lidocaine is triggered by the intrinsic
mitochondrial death pathway rather than by death receptors.
Therefore, B-cell lymphoma-2 (BCL-2) and caspase-3 are useful
indicators of lidocaine-induced apoptosis.
Ginsenosides are found in the popular herb Panax
ginseng, which is widely used in traditional Chinese medicine
(7). Ginseng has been demonstrated
to exert pharmacological effects on the central nervous,
cardiovascular, endocrine and immune systems (8). For a long time the curative effects
of ginseng were attributed to the mixed results from Panax
ginseng rather than to the individual biologically active
compounds. However, several studies have since identified
ginsenosides, including Rb, Rc, Re, Rg and Rh, as the key
ingredient for the pharmacological actions (9,10).
Among these ginsenosides, ginsenoside Rg1 (Rg1) is considered to be
one of the most active and abundant steroid saponins, and acts as
an antioxidant (11). Rg1 is
crucial in the modulation of neurotransmission and the prevention
of scopolamine-induced memory deficits, and acts by increasing
cholinergic activity (12). Rg1
has also been demonstrated to increase humoral and cell-mediated
immune responses (13).
To date, lidocaine-induced cytotoxicity, which may
result in transient neurological symptoms and cauda equina
syndrome, has been widely accepted (14,15).
Thus, a novel strategy is required to protect against the
cytotoxicity of lidocaine. In the present study the effect of Rg1
on lidocaine-induced apoptosis was assessed in human Jurkat
T-lymphoma cells. In addition, the expression levels of the
regulators of lidcaine-induced apoptosis, including BCL-2 and
caspase-3, were examined. It was hypothesized that Rg1 is a
potential drug for lidocaine-induced cytotoxicity. The present
study may provide novel insights into therapeutic options for
transient neurological symptoms and cauda equina syndrome following
spinal anesthesia.
Materials and methods
Cell culture and treatment
An acute human T lymphoma Jurkat cell line was
provided by the Laboratory of Molecular Biology, Xiangya Medical
College of Central South University (Changsha, China). The cells
were maintained in RPMI-1640 medium (Life Technologies, Inc.,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Life
Technologies, Inc.), 50 U/ml penicillin, 50 μg/ml streptomycin and
2 mM glutamine at 37°C in a humidified incubator containing 5%
CO2. Having been cultured for five days, the cells were
seeded in a 96-well microtiter plate at a density of
2×104 cells/well. The cells were divided into five
groups: Control, 3 mM lidocaine without Rg1 pretreatment, 6 mM
lidocaine without Rg1 pretreatment, 3 mM lidocaine with Rg1
pretreatment and 6 mM lidocaine with Rg1 pretreatment. The 3 mM
lidocaine with Rg1 pretreatment and 6 mM lidocaine with Rg1
pretreatment groups were incubated with 50 mg/l Rg1 for 2 h prior
to lidocaine treatment. All groups, except the control group, were
then incubated with the corresponding lidocaine concentration for
16 h, ready for further assays.
Apoptosis analysis using flow cytometry
and terminal deoxynucleotidyl transferase-mediated dUTP nick end
labeling (TUNEL) assay
To investigate the effect of Rg1 on cell apoptosis
in Jurkat cells stimulated by lidocaine, a flow cytometry assay was
performed. All fluorescence signals of labeled cells were analyzed
using the FACScan™ flow cytometer (Becton-Dickinson, San Jose, CA,
USA). The measurement of phosphatidylserine redistribution on the
plasma membrane was conducted using an Annexin V-fluorescein
isothiocyanate (FITC) Apoptosis Detection kit (Becton Dickinson)
according to the manufacturer’s instructions.
DNA fragmentation was evaluated using a
fluorescein-TUNEL assay with an Apo-Direct kit (BD, Pharmingen, San
Diego, CA, USA) according to the manufacturer’s instructions.
Positive and negative controls provided by the manufacturer and
internal controls (specimens with known DNA damage) were included
for each run. Following washing in phosphate-buffered saline (PBS)
to remove ethanol, the cell pellets were resuspended in 50 μl
freshly prepared staining solution for 60 min at 37°C. The staining
solution contained terminal deoxytransferase (TdT) enzyme, TdT
reaction buffer, FITC-tagged dUTP nucleotides and distilled
water.
Quantitative polymerase chain reaction
(PCR)
Quantitative PCR was used to measure the RNA
transcripts. Total RNA was isolated from cultured cells of
different treatment groups using TRIzol reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA) according to the manufacturer’s
instructions. Total RNA (1 μg) was digested with DNase I
(Fermentas, Vilnius, Lithuania) to eliminate residual DNA and
transcribed to cDNA with oligo (dT)12–18 primer using a
reverse transcription system (First-Strand cDNA kit; Fermentas).
The single-stranded cDNA was amplified by quantitative PCR with
BCL-2 and caspase-3 primer pairs in a Prism 7500 Sequence Detection
system (Applied Biosystems, Foster City, CA, USA) (Table I). β-actin was used as an
endogenous control (Table I). The
reaction conditions were as follows: 50°C for 5 min and 95°C for 10
min followed by 40 cycles at 95°C for 15 sec and 60°C for 45 sec.
Dissociation curves were then used to examine whether the PCR
products were specific. The results were corrected according to PCR
efficiency, tested for significance by a pair-wise fixed
reallocation randomization test (iterations=10,000) and plotted
using standard error estimation via a complex Taylor algorithm.
 | Table IPrimers used for quantitative
polymerase chaine reaction. |
Table I
Primers used for quantitative
polymerase chaine reaction.
| Gene | Forward primer (5′ to
3′) | Reverse primer (5′ to
3′) |
|---|
| Caspase-3 |
TTCATTATTCAGGCCTGCCGAGG |
TTCTGACAGGCCATGTCATCCTCA |
| BCL-2 |
CATGCCAAGAGGGAAACACCAGAA |
GTGCTTTGCATTCTTGGATGAGGG |
Western blot analysis
Following incubation, the cells of each group were
collected by centrifugation at 200 × g for 5 min (Eppendorf,
Hamburg, Germany) at 4°C and washed three times with ice-cold PBS,
pH 7.4, prior to undergoing centrifugation at 200 × g for 5 min.
The cell pellets were resuspended in radioimmunoprecipitation assay
(RIPA) lysis buffer (Pierce, Rockford, IL, USA). To obtain the
solubilized cellular proteins, lysates were centrifuged at 10,000 ×
g for 15 min at 4°C. Proteins were separated using 15%
polyacrylammide gels and elecrotransferred to polyvinylidene
diflouride membranes. The membranes were blocked and incubated with
BCL-2 (monoclonal, mouse, ab692; Abcam, Cambridge, MA, USA) and
caspase-3 (monoclonal, mouse, ab2171; Abcam) antibodies, followed
by horseradish peroxidase-conjugated second antibody (monoclonal,
rabbit, ab97046; Abcam). Subsequent to washing with PBS three
times, the signals were detected by Enhanced Chemiluminescence
(ECL) Reagent (Amersham, Little Chalfont, UK). For an internal
control, β-actin was used to normalize the target bands.
Immunofluorescence
Human T lymphoma Jurkat cells were grown on 24×24 mm
cover glasses and fixed for 30 min in 4% paraformaldehyde solution
in phosphate buffer, prior to 30 min incubation with blocking
reagent (5% fetal bovine serum in PBS). Primary antibodies of BCL-2
and caspase-3 were incubated for ≥1 h at room temperature prior to
washing. Immunofluorescence staining was performed with secondary
antibodies and 4′,6-diamidino-2-phenylindole. A conventional
fluorescence microscope (Axioskop 20;Carl Zeiss Inc., Thornwood,
NY, USA) was used for visualization.
Statistical analysis
Data are expressed as the mean ± standard deviation
(SD). Statistical analysis of the data was performed using the
Duncan’s test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Protective effect of Rg1 on
lidocaine-induced apoptosis
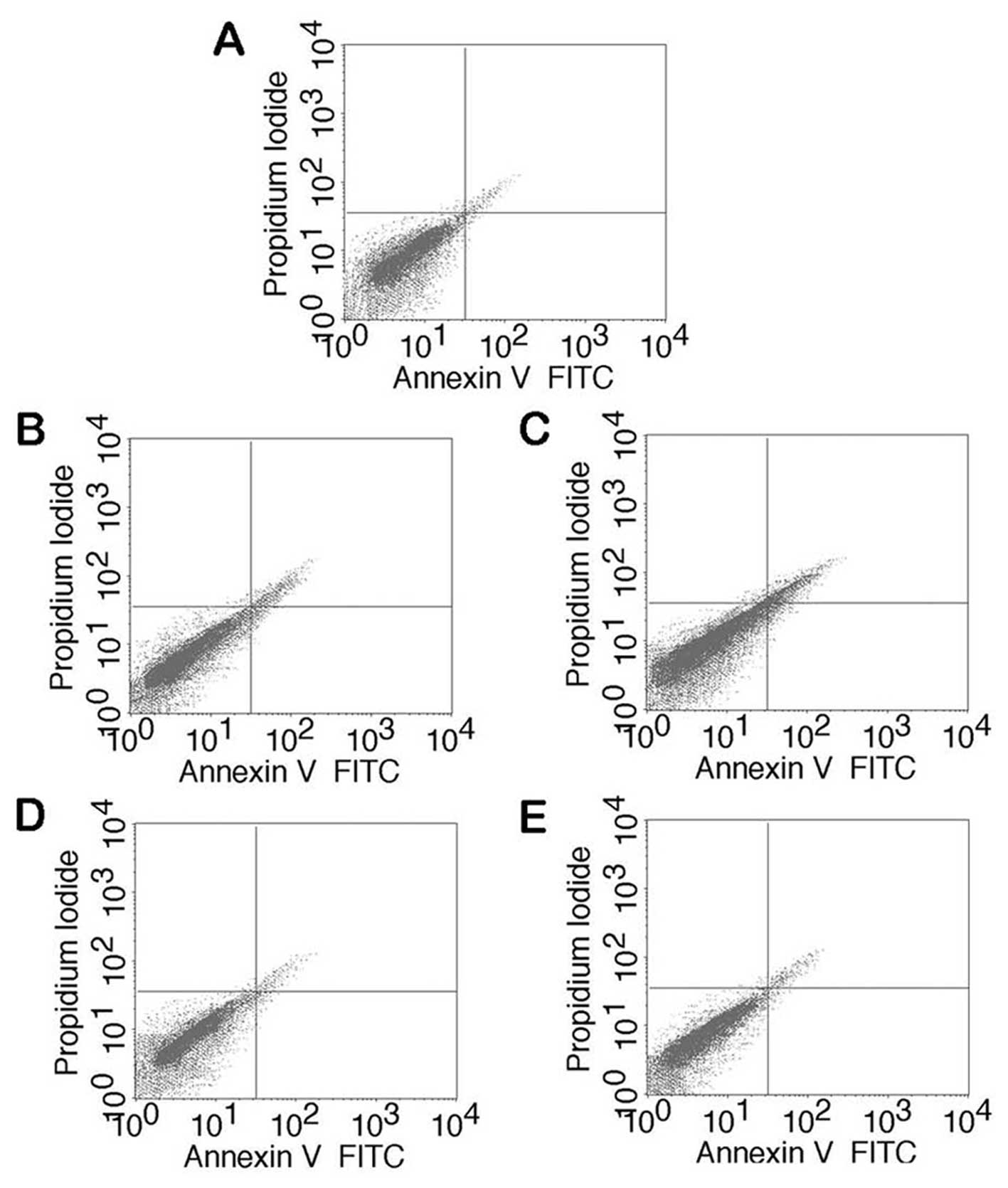
The effect of Rg1 on lidocaine-induced apoptosis was
determined using flow cytometry and TUNEL assays. The results from
the flow cytometry assay revealed that lidocaine significantly
increased the number of early- [Annexin V+/propidium
iodide (PI−)] and late-stage (Annexin
V+/PI+) apoptotic bone marrow stromal cells
(BMSCs) compared with the negative control group. The
Rg1-pretreated groups, however, exhibited decreased
lidocaine-induced apoptosis in BMSCs, including early- and
late-stage apoptosis, compared with the groups without Rg1
pretreatment (Fig. 1).
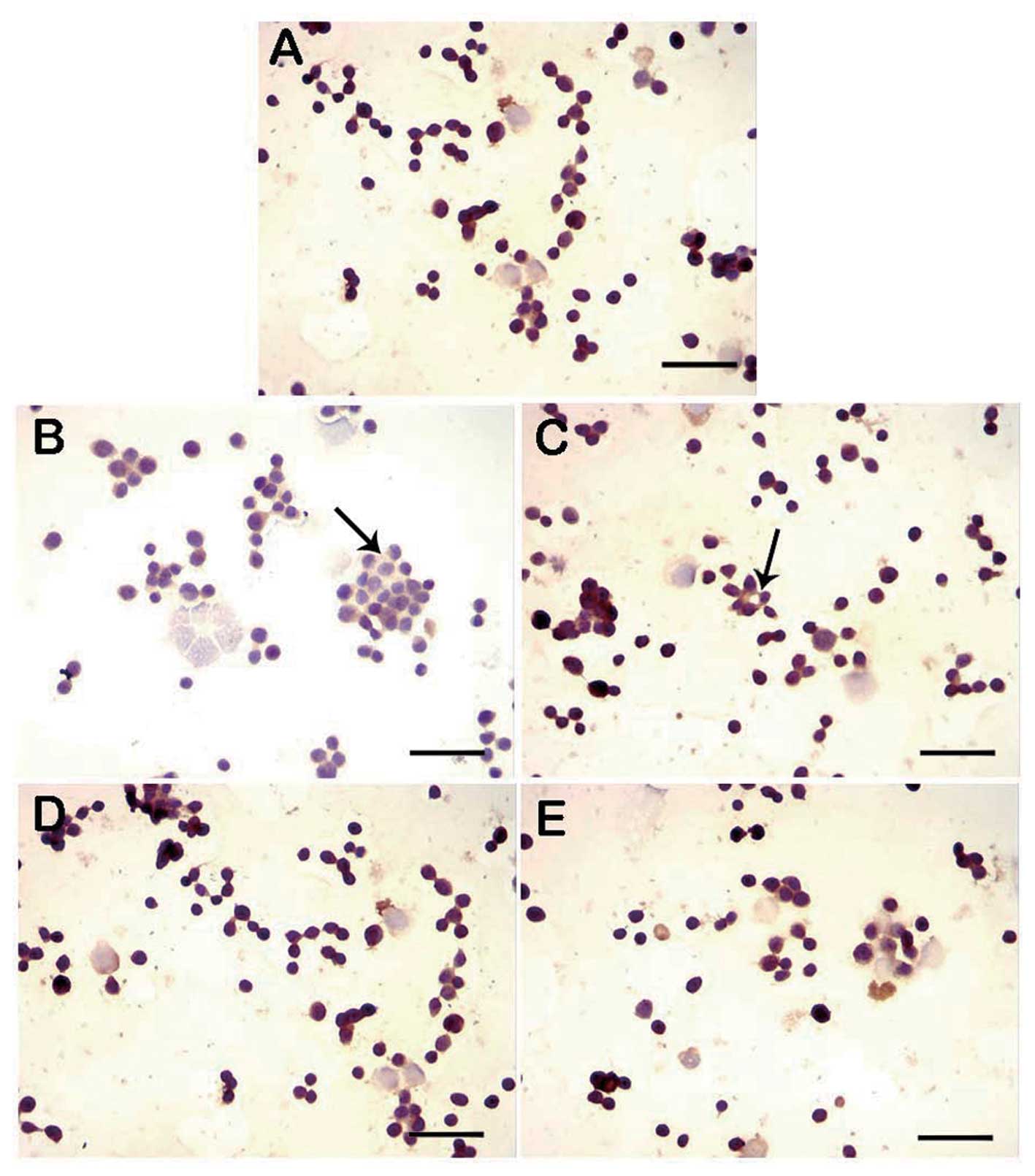
DNA fragmentation was assessed in cultures from the
control, lidocaine-treated and lidocaine with Rg1 pretreatment
groups using the TUNEL assay. Compared with the control group,
lidocaine significantly increased apoptosis in Jurkat cells. The
lidocaine with Rg1 pretreatment groups, however, showed
intermediary apoptosis compared with the control and lidocaine
groups (Fig. 2).
Rg1 suppresses the expression of
caspase-3
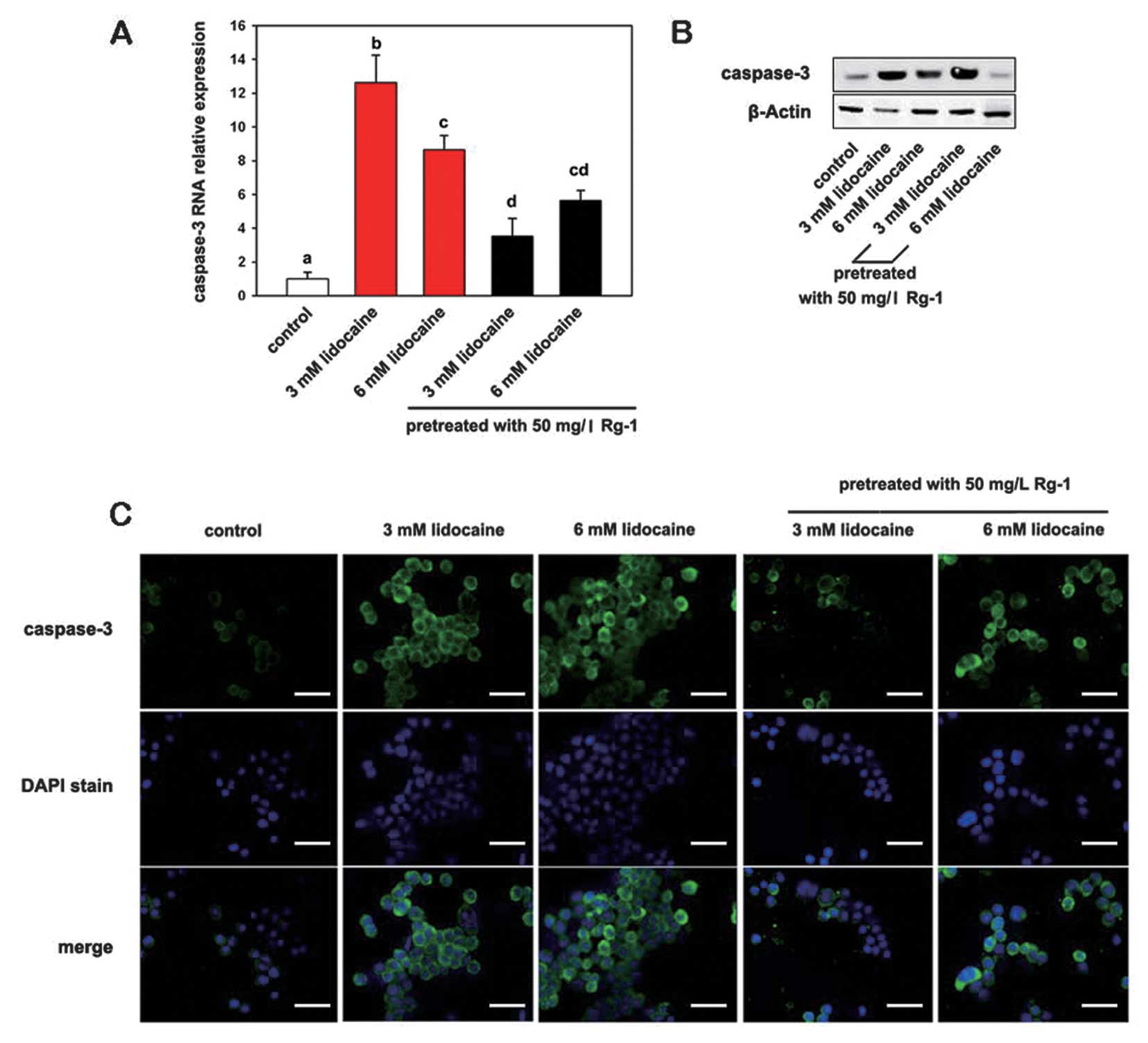
To assess the antiapoptotic effect of Rg1,
quantitative PCR was performed using specific primers for
caspase-3. In addition, protein expression of caspase-3 was
detected using western blot analysis and immunofluorescence. The
results from the RNA assay showed that lidocaine significantly
induced caspase-3 expression (P>0.05); however, following
pretreatment with Rg1 the induced caspase-3 expression was
significantly decreased (P≤0.05) (Fig.
3A). Similarly, there was a significant increase in caspase-3
protein expression following treatment with lidocaine, while
pretreatment with Rg1 decreased lidocaine-induced caspase-3 protein
expression (Fig. 3B). Caspase
expression appears high in the 3 mM lidocain with Rg1 pretreatment
group in Fig. 3B due to the
reduced cellular apoptosis. Immunofluorescence showed strong,
punctuate staining for caspase-3 in lidocaine-treated groups
compared with the control and Rg1-pretreated groups (Fig. 3C).
Rg1 increases the expression of
BCL-2
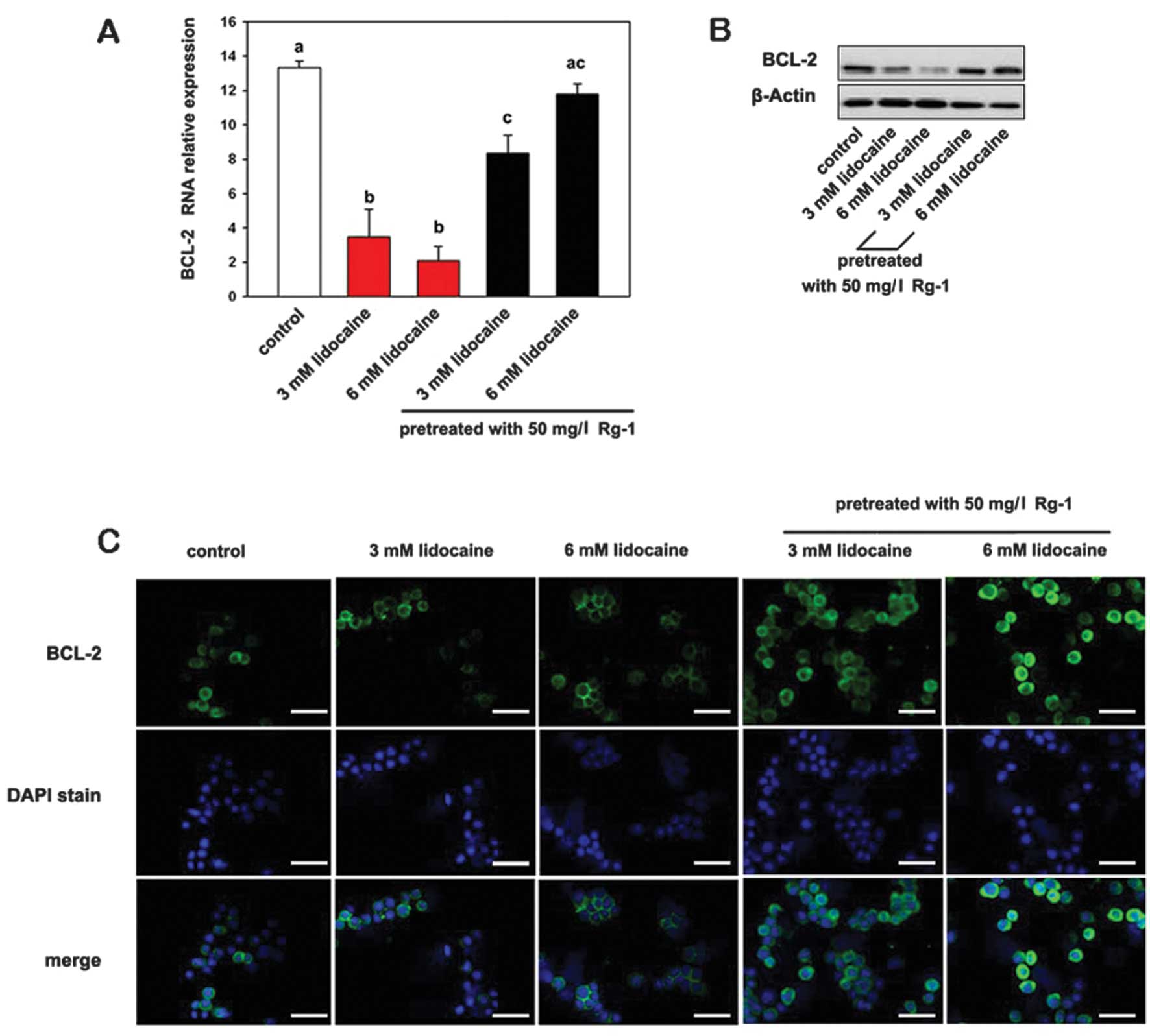
The expression of BCL-2 mRNA and protein was
decreased in Jurkat cells cultivated with lidocaine compared with
the control cells. However, cells pretreated with Rg1 showed
significantly higher BCL-2 mRNA and protein expression levels
(Fig. 4). Rg1 thus exerts a
cytoprotective effect against lidocaine-induced apoptosis in
cultured Jurkat cells. Immunofluorescence showed strong, punctuate
staining for BCL-2 in control and Rg1 pretreatment groups, compared
with the groups treated only with lidocaine.
Discussion
Lidocaine, as an anti-inflammatory, anesthetic and
antiarrhythmic substance, is often used for surface and spinal
anesthesia (16). However, studies
have shown that lidocaine is capable of inducing transient
neurological symptoms and cauda equina syndrome (17–19).
It has been suggested that lidocaine promotes apoptosis in
T-lymphoma cells, gingival fibroblasts, human chondrocytes and
leukemia cells (6), and that
lidocaine induces apoptosis intrathecally in a dose-dependent
manner (5). This effect may be
inhibited by overexpression of the cellular antiapoptotic protein
BCL-2 or by caspase-9 deficiency (6). These results indicate that lidocaine
may be dangerous for patients; however, lidocaine is still widely
used for short-lasting regional anesthesia.
Ginsenoside Rg1 is a steroidal saponin abundantly
found in ginseng (20).
Ginsenosides are the most important active compounds identified in
all species of ginseng. There are two predominant classes of
ginsenosides, which are derived either from protopanaxatriol (Rg1,
Rg2, Re and Rf) or protopanaxadiol (Rb1, Rb2, Rc and Rd) (21). In the present study, the effects of
Rg1 on lidocaine-induced apoptosis were assessed in Jurkat cells
using flow cytometry and TUNEL. The results showed that Rg1
pretreatment may protect Jurkat cells from lidocaine-induced
apoptosis, presumably by acting as a biological antiapoptotic
substance. Therefore, Rg1 may promote the recovery of Jurkat cells
under similar pathological conditions.
To understand how the apoptosis pathway changes in
response to Rg1, caspase-3 gene expression was examined. Caspases
are cystein proteases that have a key role in the execution phase
of apoptosis (22–24). Among the caspase family, caspase-3
has been widely studied, and has been proposed to have a crucial
role in the cell death process (25,26).
It has been shown that caspase-3 induces apoptosis through several
different pathways, including degrading antiapoptotic proteins, and
cleaving DNA repair molecules, extracellular matrix proteins,
cytoskeletal proteins and other associated molecules (27). The results of the present study
showed that expression of caspase-3 increased when Jurkat cells
were treated with lidocaine, and that this was positively
correlated with the rate of apoptosis. However, pretreatment with
Rg1 decreased the expression of caspase-3 and the rate of
apoptosis. These results suggested that apoptosis of Jurkat cells
induced by lidocaine is associated with the upregulation of the
expression of caspase-3, and that Rg1 reduces the rate of
apoptotisis by downregulating the expression of caspase-3. In this
study, lidocaine induced Jurkat cell apoptosis, as indicated by
increased DNA fragmentation and caspase-3 activation. However,
these apoptotic events were blocked by Rg1, indicating that Rg1
inhibits the mitochondrial apoptotic signaling pathway.
It has been shown that the release of proapoptotic
proteins from mitochondria is modulated by targeting BCL-2 family
members to the outer mitochondrial membrane (28). The BCL-2 family of proteins, which
consists of anti- and pro-apoptotic molecules, controls
mitochondrial function in apoptosis and constitutes a critical
intracellular checkpoint for apoptosis within a common cell death
pathway (29,30). The BCL-2 family includes proteins
that predispose cells to apoptosis, such as Bax and Bad, and
proteins that antagonize apoptosis, such as BCL-2 (31). The balance between these proteins
is important for the release of mitochondrial proapoptotic
proteins. BCL-2 serves as a critical regulator of pathways involved
in apoptosis, acting to inhibit cell death (32). Furthermore, the BCL-2 gene was
revealed to be upregulated in failing, as well as in aging, hearts
(33). The BCL-2 gene acts to
prevent programmed cell death of ventricular myocytes (34). Therefore, in the present study, the
effect of Rg1 on the expression of BCL-2 family genes was
investigated in Jurkat cells. Quantitative PCR and western blot
analysis revealed that lidocaine significantly increased the
expression of caspase-3, an apoptosis promoting BCL-2 family
member, and decreased antiapoptotic BCL-2 expression in Jurkat
cells. In combination, these data suggest that Rg1 is able to block
the mitochondrial apoptotic signaling pathway in Jurkat cells by
modulating the expression and activity of BCL-2 family proteins.
The identification of direct molecular targets of Rg1 that regulate
apoptotic signaling pathways is of interest and requires further
investigation (35,36).
Studies have demonstrated that lidocaine and other
local anesthetics are capable of inducing apoptosis in neuronal and
nonneuronal cells, and that apoptosis is capable of inducing
necrosis (6). Thus, understanding
the mechanism of lidocaine-induced apoptosis is key to finding the
resolution to this problem. Werdehausen et al (6) reported that the BCL-2 protein
overexpression or lack of caspase-9 expression abolishes apoptosis,
indicating that the intrinsic mitochondrial death pathway is
involved in lidocaine-induced apoptosis. Once the mechanism has
been fully elucidated, an effective therapy may be identified. In
the present study, it was observed that the ginsenoside Rg1
inhibits lidocaine-induced apoptosis, and this provides a novel
insight for the treatment of lidocaine-induced transient
neurological symptoms and cauda equina syndrome following spinal
anesthesia.
In conclusion, the present study demonstrated that
Rg1 provides protection against lidocaine-induced apoptosis in
cultured Jurkat cells. It was shown that Rg1 is a notable
antiapoptotic molecule that is capable of blocking the
caspase-dependent signaling cascade in Jurkat cells. The
mechanistic aspects of this study suggest that the protective
effect of Rg1 on lidocaine-induced apoptosis is mediated by
altering the level of BCL-2 family proteins and downregulating
caspase-3 expression. This in vitro study provides the basis
for understanding and evaluating the effect of Rg1 in the in
vivo treatment of lidocaine-induced transient neurological
symptoms and cauda equina syndrome.
References
|
1
|
Mazoit JX and Cao LS: Local anaesthetic
toxicity. Curr Opin Anaesthesiol. 8:409–413. 1995. View Article : Google Scholar
|
|
2
|
Del Valle LJ and Orihuela PA: Cleavage and
development in cultured preimplantation mouse embryos exposed to
lidocaine. Reprod Toxicol. 10:491–496. 1996.PubMed/NCBI
|
|
3
|
Saadé NE, Al Amin HA, Barchini J,
Tchachaghian S, et al: Brainstem injection of lidocaine releases
the descending pain-inhibitory mechanisms in a rat model of
mononeuropathy. Exp Neurol. 237:180–190. 2012.PubMed/NCBI
|
|
4
|
Friederich P and Schmitz T:
Lidocaine-induced cell death in a human model of neuronal
apoptosis. Eur J Anaesthesiol. 19:564–570. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnson ME, Uhl CB, Spittler KH, Wang H
and Gores GJ: Mitochondrial injury and caspase activation by the
local anesthetic lidocaine. Anesthesiology. 101:1184–1194. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Werdehausen R, Braun S, Essmann F,
Schulze-Osthoff K, et al: Lidocaine induces apoptosis via the
mitochondrial pathway independently of death receptor signaling.
Anesthesiology. 107:136–143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hon CC, Chow YC, Zeng FY and Leung FC:
Genetic authentication of ginseng and other traditional Chinese
medicine. Acta Pharmacol Sin. 24:841–846. 2003.PubMed/NCBI
|
|
8
|
Qi LW, Wang CZ and Yuan CS: Ginsenosides
from American ginseng: chemical and pharmacological diversity.
Phytochemistry. 72:689–699. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Attele AS, Wu JA and Yuan CS: Ginseng
pharmacology: multiple constituents and multiple actions. Biochem
Pharmacol. 58:1685–1693. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tawab MA, Bahr U, Karas M, Wurglics M and
Schubert-Zsilavecz M: Degradation of ginsenosides in humans after
oral administration. Drug Metab Dispos. 31:1065–1071. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yi LN, Yin XY, Jiang YF and Liu QS:
Preparation and properties of ginsenoside Rg1 molecularly imprinted
polymers. Adv Mat Res. 550–553:1715–1718. 2012.PubMed/NCBI
|
|
12
|
Wu J, Pan Z, Wang Z, Zhu W, et al:
Ginsenoside Rg1 protection against β-amyloid peptide-induced
neuronal apoptosis via estrogen receptor α and glucocorticoid
receptor-dependent anti-protein nitration pathway.
Neuropharmacology. 63:349–361. 2012.
|
|
13
|
Zhang DF, Xu H, Sun BB, Li JQ, et al:
Adjuvant effect of ginsenoside-based nanoparticles (ginsomes) on
the recombinant vaccine against Eimeria tenella in chickens.
Parasitol Res. 110:2445–2453. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Drasner K: Local Anesthetic neurotoxicity
and cauda equina syndrome. Complications in regional anesthesia and
pain medicine. Neal and Rathmell JP: Lippincott, Williams and
Wilkins; Philadelphia, PA, USA: pp. 1252012
|
|
15
|
Vaghadia H, Neilson G and Lennox PH:
Selective spinal anesthesia for outpatient transurethral
prostatectomy (TURP): randomized controlled comparison of
chloroprocaine with lidocaine. Acta Anaesthesiol Scand. 56:217–223.
2012. View Article : Google Scholar
|
|
16
|
Hahnenkamp K, Herroeder S and Hollmann MW:
Regional anaesthesia, local anaesthetics and the surgical stress
response. Best Pract Res Clin Anaesthesiol. 18:509–527. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hampl KF, Schneider MC, Pargger H, Gut J,
et al: A similar incidence of transient neurologic symptoms after
spinal anesthesia with 2% and 5% lidocaine. Anesth Analg.
83:1051–1054. 1996.
|
|
18
|
Gerancher JC: Cauda equina syndrome
following a single spinal administration of 5% hyperbaric lidocaine
through a 25-gauge Whitacre needle. Anesthesiology. 87:687–689.
1997.
|
|
19
|
Zaric D, Christiansen C, Pace NL and
Punjasawadwong Y: Transient neurologic symptoms after spinal
anesthesia with lidocaine versus other local anesthetics: a
systematic review of randomized, controlled trials. Anesth Analg.
100:1811–1816. 2005. View Article : Google Scholar
|
|
20
|
Chan RY, Chen WF, Dong A, Guo D and Wong
MS: Estrogen-like activity of ginsenoside Rg1 derived from Panax
notoginseng. J Clin Endocrinol Metab. 87:3691–3695. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leung KW, Yung KK, Mak NK, Yue PY, et al:
Angiomodulatory and neurological effects of ginsenosides. Curr Med
Chem. 14:1371–1380. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Saraste A and Pulkki K: Morphologic and
biochemical hallmarks of apoptosis. Cardiovasc Res. 45:528–537.
2000. View Article : Google Scholar
|
|
23
|
Slee EA, Adrain C and Martin SJ:
Executioner caspase-3, -6 and -7 perform distinct, non-redundant
roles during the demolition phase of apoptosis. J Biol Chem.
276:7320–7326. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jänicke RU, Sprengart ML, Wati MR and
Porter AG: Caspase-3 is required for DNA fragmentation and
morphological changes associated with apoptosis. J Biol Chem.
273:9357–9360. 1998.
|
|
26
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dai ZJ, Gao J, Ji ZZ, Wang XJ, et al:
Matrine induces apoptosis in gastric carcinoma cells via alteration
of Fas/FasL and activation of caspase-3. J Ethnopharmacol.
123:91–96. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Danial NN and Korsmeyer SJ: Cell death:
critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng EH, Wei MC, Weiler S, Flavell RA, et
al: BCL-2, BCL-X(L) sequester BH3 domain-only molecules preventing
BAX- and BAK-mediated mitochondrial apoptosis. Mol Cell. 8:705–711.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reed JC: Bcl-2 family proteins. Oncogene.
17:3225–3236. 1998. View Article : Google Scholar
|
|
33
|
Li W, Ma N, Ong LL, Nesselmann C, et al:
Bcl-2 engineered MSCs inhibited apoptosis and improved heart
function. Stem Cells. 25:2118–2127. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
MacLellan WR and Schneider MD: Death by
design. Programmed cell death in cardiovascular biology and
disease. Circ Res. 81:137–144. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim EC, Yun BS, Ryoo IJ, Min JK, et al:
Complestatin prevents apoptotic cell death: inhibition of a
mitochondrial caspase pathway through AKT/PKB activation. Biochem
Biophys Res Commun. 313:193–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Min JK, Kim JH, Cho YL, Maeng YS, et al:
20(S)-Ginsenoside Rg3 prevents endothelial cell apoptosis via
inhibition of a mitochondrial caspase pathway. Biochem Biophys Res
Commun. 349:987–994. 2006. View Article : Google Scholar : PubMed/NCBI
|