Introduction
Sevoflurane is widely used in the clinic,
particularly in pediatric anesthesia. However, infants brains are
at a stage at which important changes are occurring, including
synapse formation and axon and dendrite growth; this renders them
strongly susceptible to environmental influences, such as general
anesthetics. A number of studies have demonstrated that early
exposure to a variety of anesthetics can cause widespread
neurodegeneration in the developing brain and persistent learning
deficits (1,2). Loepke et al (3) observed that isoflurane led to
apoptotic neurodegeneration, and Head et al (4) elucidated the relevant signaling
pathways. Satomoto et al (5) demonstrated that exposing neonatal
mice to sevoflurane results in learning deficits, in addition to
autism-like abnormal social behavior. These studies together
demonstrate that sevoflurane is harmful to the developing
brain.
Methyl-CpG island binding protein 2 (MeCP2) is a
transcriptional repressor and is important in neuron maturation and
normal brain function (6). Studies
have suggested that MeCP2 is closely associated with NMDA receptors
in the brain (7,8). MeCP2 is a nuclear protein that
selectively binds methylated DNA, then recruits other proteins to
form an inhibition complex, thereby inhibiting the expression of
various genes (9,10). MeCP2 gene mutations can cause the
neurodevelopmental disorder Rett syndrome (11), which can lead to cognitive
impairment, motor disability and repetitive stereotyped hand
movements. Up to 80% of those affected with Rett syndrome
experience seizures. A number of studies have hypothesized that
these symptoms may be due to defects in experience-dependent
synapse maturation (12,13), where normal synapse connections
fail to establish in the critical period. These studies have
demonstrated that MeCP2 is critical in the maturation of the
nervous system and the normal functioning of nerve cells.
cAMP response element-binding protein (CREB) is a
nuclear regulatory factor that may regulate gene transcription
following phosphorylation of serine 133, thus enhancing the
expression of multiple target genes. One study indicated that CREB
is the major transcription regulatory factor of brain-derived
neurotrophic factor (BDNF), and thus increases BDNF expression.
This is a crucial process in the survival and differentiation of
neurons (14). Another study
demonstrated that isoflurane anesthesia induces neuronal apoptosis
by affecting the formation of neuronal synapses in the
developmental stages, potentially via the BDNF-p75-RhoA signaling
pathway (4).
Memantine is a derivative of amantadine, and acts on
the glutamatergic system by inhibiting NMDA receptors. It is
involved in neuroprotection and acts to preserve normal synaptic
function. It is approved for use in moderate-to-severe Alzheimer’s
disease, by the FDA and the European Medicines Agency, and has been
demonstrated to produce positive effects on cognition, mood and
behavior (15,16).
To investigate the molecular mechanism of
neurodegeneration induced by sevoflurane, neonatal mice were
exposed to sevoflurane and the protein expression levels of CREB
and BDNF were assessed in the developing hippocampus. Protein
expression levels of MeCP2 and the effect of pre-injected memantine
were also examined, in order to determine whether memantine is able
to reverse the neurodegeneration.
Materials and methods
Animals and sevoflurane exposure
All animal experiments were performed using
protocols approved by the institutional animal use and care
committee of the Zhongshan Hospital, Fudan University (Shanghai,
China). At postnatal day 7 (P7), male C57BL/6 mice (weight, 3–5 g;
Shanghai Laboratory Animal Center, Shanghai, China) were randomly
divided into a sevoflurane-treated group (n=6) and an air-treated
control group (n=6) for analysis of the effects of sevoflurane on
CREB phosphorylation, BDNF expression and MeCP2 phosphorylation
levels. Mice were placed in a plastic container and continuously
exposed to 1.5% sevoflurane (Maruishi Pharmaceutical Co., Osaka,
Japan) in air, or to air alone for 2 h, with a gas flow of 2 l/min.
For further experiments, male C57BL/6 mice at postnatal day 7 (P7)
were randomly divided into four groups: The sevoflurane-saline
group (sevo group, n=7); the air-saline group (control group, n=6);
the sevoflurane-memantine group (sevo+mem group, n=7); and the
air-memantine group (mem group, n=6). Mice received 1 mg/kg saline
or memantine intraperitoneally prior to sevoflurane or air
treatment. The mice were then placed in a plastic container and
continuously exposed to 1.5% sevoflurane in air or to air alone for
2 h, with a gas flow of 2 l/min. During exposure to sevoflurane or
air, the container was heated to 37°C with a heating pad. The
concentrations of sevoflurane, oxygen and carbon dioxide in the
container were monitored with a gas monitor (Datex Cardiocap II,
Datex-Ohmeda, Madison, WI, USA). Following exposure to sevoflurane
or air, the mice were returned to their cages. The mice were housed
six per cage and maintained on a 12 h light/dark cycle with access
to food and water ad libitum. Two hours post-exposure the mice were
sacrificed by decapitation, and their hippocampi were removed.
Arterial blood gas analyses
Arterial blood samples were obtained from the left
cardiac ventricle of the mice immediately after exposure to
sevoflurane, and were transferred to heparinized glass capillary
tubes. Blood pH, partial pressure of carbon dioxide in mmHg
(PaCO2), partial pressure of oxygen in mmHg
(PaO2), lactate (Lac), and bicarbonate (HCO3)
were analyzed immediately after blood collection using a GEM
Premier 3000 analyzer (Instrumentation Laboratory, Lexington, MA,
USA).
Protein extraction and western blot
analysis
Resected hippocampi were placed into 1.5-ml
centrifuge tubes and preserved in liquid nitrogen. All methods were
conducted on ice. An NE-PER Nuclear and Cytoplasmic Extraction kit
(cat no.78835; Thermo Fisher Scientific, Waltham, MA, USA) was used
to extract protein samples. All steps were conducted according to
the manufacturer’s instructions. Sodium dodecyl sulphate (SDS) was
added to the samples prior to boiling for 10 min at 100°C. Equal
quantities of protein (15 μg) were used to detect the expression of
the proteins of interest. Samples were electrophoresed on 10 or 15%
SDS polyacrylamide gel, blotted onto polyvinylidine fluoride
membranes (Bio-Rad Laboratories, Hercules, CA, USA) and then
incubated with the following antibodies overnight at 4°C:
Anti-phospho-CREB (ser133), (cat no. 06-519, EMD Millipore,
Billerica, MA, USA) 1:4,000 dilution in 5% non-fat milk; anti-CREB
(cat no. MAB5432, Millipore) 1:5,000 dilution in 5% non-fat milk;
anti-BDNF (cat no. AB1779SP, Millipore) 1:1,000 dilution in 5%
non-fat milk; anti-MeCP2 (cat no. 3456P, Cell Signaling Technology,
Danvers, MA, USA) 1:4,000 dilution in 5% non-fat milk;
anti-phospho-MeCP2-S421 (cat no. AP3693a, Abgent Biotech, Suzhou,
China) 1:2,000 dilution in 5% non-fat milk); and anti-actin (cat
no. A5441, Sigma-Aldrich, St. Louis, MO, USA) 1:10,000 dilution in
5% non-fat milk. The following day, the blots were incubated for 1
h at room temperature with horseradish peroxidase-conjugated
secondary goat anti-rabbit or goat anti-mouse immunoglobulin G
(Kangchen, Shanghai, China), 1:5,000 dilution in 5% non-fat milk.
Immunoreactive bands were visualized using Amersham ECL Prime
Western Blotting Detection kit (cat NO.RPN2232; GE Healthcare,
Chalfont St. Giles, UK). The protein signals were quantified using
Quantity One software and a GS-800 Calibrated Imaging Densitometer
(Bio-Rad Laboratories) and normalized to a corresponding internal
reference: CREB for the exression of p-CREB-S133, MeCP2 for
P-MeCP2-S421 and actin for BDNF.
Statistical analysis
All data are presented as the mean ± standard error.
Data were analyzed using the unpaired Student’s t-test in Origin
software, version 7.5 (OriginLab, Northampton, MA, USA). P<0.05
was considered to represent a statistically significant
difference.
Results
Sevoflurane does not induce metabolic or
respiratory deterioration
Blood gas analyses indicated that there was no
deterioration in respiration or metabolism in the animals following
a 2-h sevoflurane exposure. All parameters were tested, including
PH, PaCO2, PaO2, oxygen saturation, Lac and
HCO3. No significant differences in any of the
parameters were detected between the sevoflurane and the control
group (P>0.05; Table I).
 | Table IArterial blood gas analysis. |
Table I
Arterial blood gas analysis.
| Arterial blood
gas | Control (n=6) | Sevoflurane
(n=6) |
|---|
| pH | 7.4±0.05 | 7.38±0.04 |
| PaCO2
(mmHg) | 28.4±3.0 | 30.1±4.0 |
| PaO2
(mmHg) | 91.0±6.8 | 88.8±5.6 |
| SaO2
(%) | 93.6±2.6 | 92.7±2.5 |
| Lac (mmol/l) | 1.1±0.5 | 1.2±0.4 |
| HCO3
(mmol/l) | 23.8±1.6 | 24.1±1.5 |
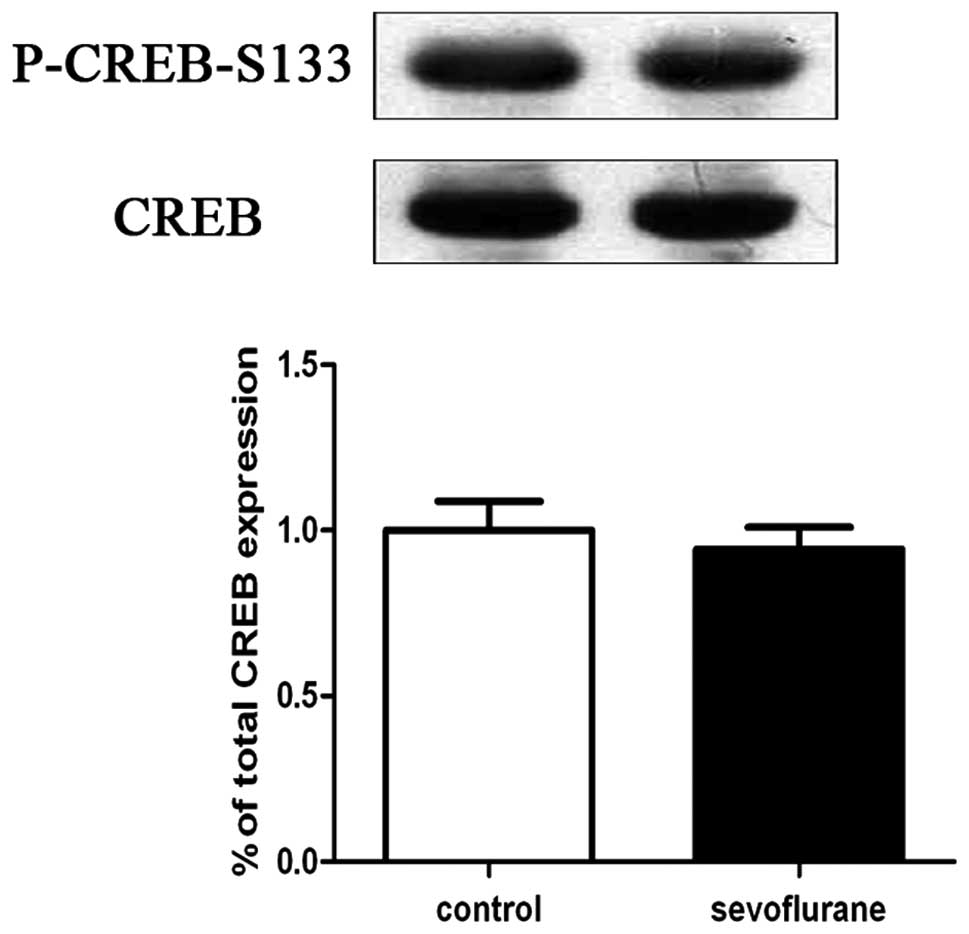
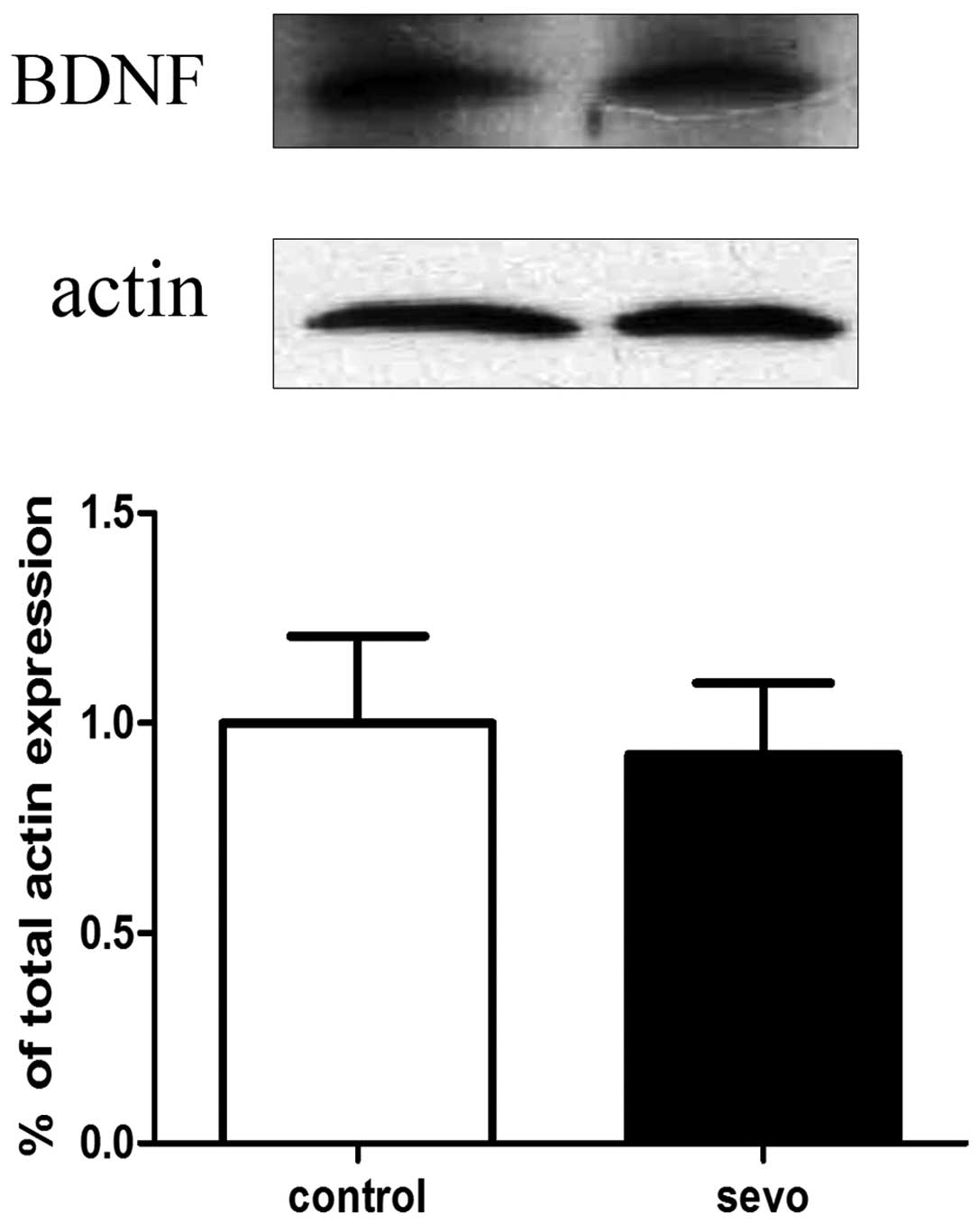
Sevoflurane does not cause changes in
CREB phosphorylation and BDNF expression in the hippocampus
Western blot analyses of hippocampal CREB
phosphorylation and BDNF expression levels were performed 2 h
following exposure to sevoflurane or air. The results indicated
that there were no significant differences in levels of CREB
phosphorylation (Fig. 1) and BDNF
(Fig. 2) expression between the
sevoflurane-treated (n=6) and air-treated (n=6) groups.
Sevoflurane treatment increases MeCP2
phosphorylation at the serine 421 loci in the hippocampus
Western blot analyses of hippocampal MeCP2
phosphorylation in the sevoflurane-treated and control groups were
performed 2 h following exposure to sevoflurane or air. The results
indicated that sevoflurane-treated mice exhibited an increase in
hippocampal MeCP2 phosphorylation at serine 421 loci
(P-MeCP2-S421), compared with phosphorylation in the hippocampi of
the control mice. There expression level of P-MeCP2-S421 was
increased in the sevoflurane-treated group (n=6) compared with the
control group(n=6) (P<0.05; Fig.
3).
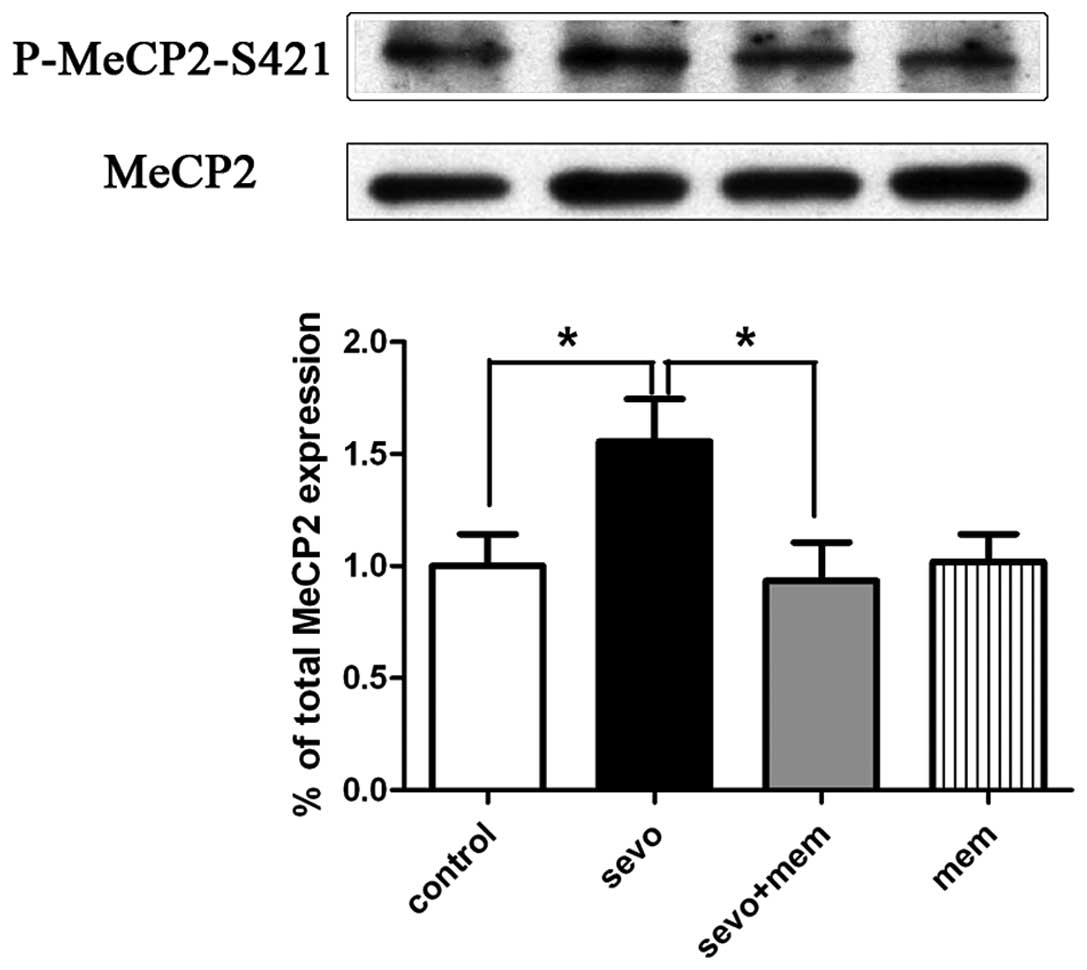
Sevoflurane increases the MeCP2
phosphorylation at the serine 421 loci in the hippocampus, and
pre-injection of memantine reverses this phenomenon
Western blot analyses of hippocampal MeCP2
phosphorylation were performed 2 h following exposure to
sevoflurane or air. Compared with the control group (air and
saline, n=6), the sevo group (sevoflurane and saline, n=7)
exhibited a significant increase in hippocampal P-MeCP2-S421
expression levels (P<0.05; Fig.
4). Compared with the sevo group, the sevo+mem group exhibited
a significant decrease in hippocampal P-MeCP2-S421 levels
(P<0.05; Fig. 4). No
significant difference was detected between the mem group
(memantine and saline, n=6) and the control group (n=6).
Discussion
In the present study, it was demonstrated that
P-MeCP2-S421 expression levels in hippocampi resected at P7
increased in mice exposed to 1.5% sevoflurane for 2 h, and that
memantine pre-injection was able to reverse this increase. However,
sevoflurane did not cause significant changes in CREB
phosphorylation and BDNF expression levels in the hippocampus.
A dose of 1.5% sevoflurane, which did not inhibit
respiration and circulation in mouse pups, was selected for the
current study. Arterial blood analyses confirmed that none of the
mice experienced hypoxemia or hypercapnia during the 2-h
sevoflurane exposure; there were no significant differences in any
of the tested parameters between the sevoflurane group and the
control group. These results exclude the possibility of hypoxemia
and hypercapnia affecting the outcome of the following
experiments.
A previous study demonstrated that early exposure to
sevoflurane causes widespread neurodegeneration in the developing
brain (17). However, the exact
mechanism of action underlying the effect of sevoflurane remains
unknown. The results of the present study may provide a possible
explanation for sevoflurane-mediated neurodegeneration, as they
suggested that MeCP2 may be important in neuronal degeneration
following neonatal sevoflurane exposure.
The γ-aminobutyric acid type A (GABA) and
N-methyl-D-aspartate glutamate (NMDA) receptors are essential for
the development of an ordered neural map (18,19),
and are important in the alteration of synaptic transmission.
Neurotransmitters or compounds that act on them may contribute to
the impairment of brain development and synaptogenesis (20,21).
MeCP2 links closely with NMDA receptors in the brain, and NMDA
receptors (particularly NR2A) are essential for visual cortical
function in the absence of MeCP2 (22). The activity-dependent expression of
another NMDA subunit (NR2B) is mediated by MeCP2-dependent
epigenetic regulation (23). Thus,
as indicated in the present study, MeCP2 may regulate NMDA
receptors, leading to various effects on brain function in mice.
MeCP2 phosphorylation at serine 421 loci is a key signal, which may
cause downstream changes in the signaling pathway and influence the
central nervous system. Previous studies indicated that CDKL5 is a
target of MeCP2 in the brain, and is regulated by DNA methylation
(24). MeCP2 can also interact
with some microRNAs to regulate brain function (25). Further study of these aspects is
required in order to figure out whether and how microRNAs are
involved in the regulation of neuroplasticity in the brain.
Memantine is an amantadine derivative and NMDA
receptor inhibitor. It has been used to treat Alzheimer’s disease
and is accepted to be safer than other NMDA receptor inhibitors, as
a therapeutic dose of memantine has a greater affinity for synaptic
NMDA receptors than extrasynaptic NMDA receptors. Thus, it exhibits
an important role in neuroprotection, while preserving normal
synaptic function (26). In the
present study, memantine was able to reverse the increase in MeCP2
phosphorylation in the hippocampus following sevoflurane exposure.
This demonstrates that memantine may have a protective effect
against neurodegeneration induced by sevoflurane exposure.
In conclusion, the results of the present study
demonstrated that P-MeCP2-S421 expression levels in the hippocampus
increased in P7 mice exposed to 1.5% sevoflurane for 2 h, and
pre-injected memantine reversed this increase. Sevoflurane did not
cause changes in CREB phosphorylation or BDNF expression levels in
the hippocampus. Future investigation of MeCP2 and NMDA receptors
is required in order to further investigate their effects on the
central nervous system during its development.
Acknowledgements
The current study was supported by grants from the
National Natural Science Foundation of China (grant no.
81100796).
References
|
1
|
Jevtovic-Todorovic V, Hartman RE, Izumi Y,
et al: Early exposure to common anesthetic agents causes widespread
neurodegeneration in the developing rat brain and persistent
learning deficits. J Neurosci. 23:876–882. 2003.
|
|
2
|
Liang G, Ward C, Peng J, Zhao Y, Huang B
and Wei H: Isoflurane causes greater neurodegeneration than an
equivalent exposure of sevoflurane in the developing brain of
neonatal mice. Anesthesiology. 112:1325–1334. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Loepke AW, Istaphanous GK, McAuliffe JR,
et al: The effects of neonatal isoflurane exposure in mice on brain
cell viability, adult behavior, learning, and memory. Anesth Analg.
108:90–104. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Head BP, Patel HH, Niesman IR, Drummond
JC, Roth DM and Patel PM: Inhibition of p75 neurotrophin receptor
attenuates isoflurane-mediated neuronal apoptosis in the neonatal
central nervous system. Anesthesiology. 110:813–825. 2009.
View Article : Google Scholar
|
|
5
|
Satomoto M, Satoh Y, Terui K, et al:
Neonatal exposure to sevoflurane induces abnormal social behaviors
and deficits in fear conditioning in mice. Anesthesiology.
110:628–637. 2009. View Article : Google Scholar
|
|
6
|
Kishi N and Macklis JD: MECP2 is
progressively expressed in post-migratory neurons and is involved
in neuronal maturation rather than cell fate decisions. Mol Cell
Neurosci. 27:306–321. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nguyen MV, Du F, Felice CA, et al: MeCP2
is critical for maintaining mature neuronal networks and global
brain anatomy during late stages of postnatal brain development and
in the mature adult brain. J Neurosci. 32:10021–10034. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kron M, Howell CJ, Adams IT, et al: Brain
activity mapping in Mecp2 mutant mice reveals functional deficits
in forebrain circuits, including key nodes in the default mode
network, that are reversed with ketamine treatment. J Neurosci.
32:13860–13872. 2012. View Article : Google Scholar
|
|
9
|
Cohen S, Gabel HW, Hemberg M, et al:
Genome-wide activity-dependent MeCP2 phosphorylation regulates
nervous system development and function. Neuron. 72:72–85. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fuks F, Hurd PJ, Wolf D, Nan X, Bird AP
and Kouzarides T: The methyl-CpG-binding protein MeCP2 links DNA
methylation to histone methylation. J Biol Chem. 278:4035–4040.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bienvenu T and Chelly J: Molecular
genetics of Rett syndrome: when DNA methylation goes unrecognized.
Nat Rev Genet. 7:415–426. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Z, Hong EJ, Cohen S, et al:
Brain-specific phosphorylation of MeCP2 regulates
activity-dependent Bdnf transcription, dendritic growth, and spine
maturation. Neuron. 52:255–269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Zhong X, Chau KF, Williams EC and
Chang Q: Loss of activity-induced phosphorylation of MeCP2 enhances
synaptogenesis, LTP and spatial memory. Nat Neurosci. 14:1001–1008.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tao X, Finkbeiner S, Arnold DB, Shaywitz
AJ and Greenberg ME: Ca2+ influx regulates BDNF
transcription by a CREB family transcription factor-dependent
mechanism. Neuron. 20:709–726. 1998.
|
|
15
|
Areosa Sastre ASF and McShane R: Memantine
for dementia. The Cochrane Library. 2006
|
|
16
|
Mount C and Downton C: Alzheimer disease:
progress or profit? Nat Med. 12:780–784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang X, Xue Z and Sun A: Subclinical
concentration of sevoflurane potentiates neuronal apoptosis in the
developing C57BL/6 mouse brain. Neurosci Lett. 447:109–114. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Simon DK, Prusky GT, O’Leary DD and
Constantine-Paton M: N-methyl-D-aspartate receptor antagonists
disrupt the formation of a mammalian neural map. Proc Natl Acad Sci
USA. 89:10593–10597. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hardingham GE, Fukunaga Y and Bading H:
Extrasynaptic NMDARs oppose synaptic NMDARs by triggering CREB
shut-off and cell death pathways. Nat Neurosci. 5:405–414.
2002.PubMed/NCBI
|
|
20
|
Johnson SA, Young C and Olney JW:
Isoflurane-induced neuroapoptosis in the developing brain of
nonhypoglycemic mice. J Neurosurg Anesthesiol. 20:21–28. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yon JH, Daniel-Johnson J, Carter LB and
Jevtovic-Todorovic V: Anesthesia induces neuronal cell death in the
developing rat brain via the intrinsic and extrinsic apoptotic
pathways. Neuroscience. 135:815–827. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Durand S, Patrizi A, Quast KB, et al: NMDA
receptor regulation prevents regression of visual cortical function
in the absence of Mecp2. Neuron. 76:1078–1090. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee S, Kim W, Ham BJ, Chen W, Bear MF and
Yoon BJ: Activity-dependent NR2B expression is mediated by
MeCP2-dependent epigenetic regulation. Biochem Biophys Res Commun.
377:930–934. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carouge D, Host L, Aunis D, Zwiller J and
Anglard P: CDKL5 is a brain MeCP2 target gene regulated by DNA
methylation. Neurobiol Dis. 38:414–424. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Im HI, Hollander JA, Bali P and Kenny PJ:
MeCP2 controls BDNF expression and cocaine intake through
homeostatic interactions with microRNA-212. Nat Neurosci.
13:1120–1127. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia P, Chen HS, Zhang D and Lipton SA:
Memantine preferentially blocks extrasynaptic over synaptic NMDA
receptor currents in hippocampal autapses. J Neurosci.
30:11246–11250. 2010. View Article : Google Scholar : PubMed/NCBI
|