Introduction
Malignant glioma is the most common primary tumor of
the brain and the most aggressive human tumor (1,2).
Despite intense efforts to find effective treatments, high-grade
malignant gliomas, particularly glioblastoma (GBM), are currently
incurable. Temozolomide (TMZ), a first-line chemotherapeutic agent,
in addition to radiotherapy and surgical resection, improved the
overall survival rate and progression-free survival of patients
with newly diagnosed GBM (3,4).
However, numerous GBM cases gradually develop resistance to TMZ
treatment despite the initial benefit. Intrinsic or acquired
chemoresistance is a major cause of TMZ treatment failure in GBM
patients (5). TMZ is a
DNA-alkylating agent that causes DNA damage predominantly by
methylating the O6-position of guanine, eventually
inducing cell death (6). Previous
studies have suggested a significant inverse association between
cellular ability to repair O-alkyl guanine and sensitivity to
methylating agents (7–10). O6-methylguanine-DNA
methyl-transferase (MGMT) is a DNA repair protein, which restores
the structural integrity of O6-alkylated guanine bases
by direct transfer of the alkyl group to an internal cysteine
residue (Cys145) within its active site (5,7). The
level of MGMT expression positively correlates with in vitro
and in vivo glioma resistance to TMZ and
bis-chloroethylnitrosourea (11,12).
Previously, evidence in certain malignancies has
supported the theory that various types of tumor are organized in a
hierarchy of heterogeneous cell populations (13,14).
The capability to sustain tumor formation and growth is exclusively
due to a small proportion of tumor cells termed cancer stem cells
or tumor-initiating cells, which are termed glioblastoma stem cells
(GSCs) in GBM (15). In addition,
a number of studies suggest that GSCs are closely associated with
resistance to radiotherapy and chemotherapy although the underlying
mechanism remains to be elucidated (16–23).
Resistance to apoptosis is a fundamental part of
carcinogenesis and is critical for chemotherapeutic drug resistance
(24). It is well established that
the p53 pathway is critical in detecting DNA damage and regulating
the signaling pathways required to mediate apoptosis. p53
upregulated modulator of apoptosis (PUMA) was identified as a
principal mediator of p53-dependent and independent apoptotic
pathways (25). PUMA is a B-cell
lymphoma 2 (Bcl-2) homology 3 protein and a potent pro-apoptotic
Bcl-2 family member (26). A
previous study demonstrated that PUMA was able to induce apoptosis
of glioma cells and overexpression of PUMA induces activation of
caspases and cytochrome c release (27).
It has been the focus of ongoing preclinical and
clinical research to understand the mechanisms underlying TMZ
resistance in human glioma and develop more effective strategies to
overcome chemotherapy resistance (28). This suggested that a reduction of
PUMA may be responsible for TMZ resistance in U251R GSCs.
Therefore, the present study aimed to examine whether the
introduction of PUMA into the TMZ resistant CD133+ U251R
cells may reverse the drug resistance of U251R GSCs cells in
response to TMZ treatment.
Materials and methods
Cell culture and treatments
The human glioma cell line, U251MG, with partial TMZ
sensitivity was purchased from the Chinese Academy of Sciences Cell
Bank (Shanghai, China). U251MG cells were cultured in the following
complete medium: Dulbecco’s modified Eagle’s medium (DMEM;
Invitrogen Life Technologies, Carlsbad, CA, USA), 10 mM HEPES
(Invitrogen Life Technologies), 10% heat-inactivated fetal bovine
serum (Irvine Scientific, Santa Ana, CA, USA), 100 U/ml penicillin
and 100 μg/ml streptomycin. Over a period of 6 months,
TMZ-resistant U251MG cells, termed U251R, were obtained by
sequentially increasing the concentration of TMZ. Subsequently,
U251R cells were divided into two subgroups, CD133+
U251R cells and CD133− U251R cells using positive
magnetic activated cell separation. The medium was replaced with
stem cell-permissive medium containing DMEM/F-12 with high glucose,
20 ng/ml basic fibroblast growth factor (Peprotech, Rocky Hill, NJ,
USA), 20 ng/ml of epidermal growth factor (Peprotech), 4
μg/ml B27 supplement (Gibco-BRL, Carlsbad, CA, USA), 0.1
mg/ml Glutamax (Invitrogen Life Technologies), 100 U/ml penicillin
G and 100 μg/ml streptomycin (Gibco-BRL). In all
experiments, cells were maintained at 37°C in a humidified
atmosphere with 5% CO2. The cells were observed and
photoed with an Olympus IX71 inverted fluorescence microscope
(Olympus, Tokyo, Japan). TMZ was obtained from Schering Corporation
(Kenilworth, NJ, USA) and dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich, St. Louis, MO, USA) to produce a 150 mmol/l stock
solution. The stock solution was diluted to appropriate
concentrations with cell culture media (DMSO≤0.3%, v/v). For
combination treatments, cells were infected with adenoviruses for
16–18 h prior to TMZ treatment.
Recombinant adenoviruses
Adenoviruses expressing wild-type PUMA (Ad-PUMA)
bound to DsRed were constructed using the Ad-Easy system from
Qbiogene (Irvine Scientific). High-titer viruses were produced in
HEK-293 cells and purified by a series of CsCl gradient
ultracentrifugation, as previously described (29). The cells were infected with
Ad-DsRed-PUMA, diluted in cell culture medium in the absence of
serum or antibiotics at the indicated multiplicity of infection
(MOI) for 2 h at 37°C.
Magnetic cell sorting and flow
cytometry
U251R cells were dissociated and resuspended in
phosphate-buffered saline (PBS) containing 0.5% bovine serum
albumin and 2 mmol/l EDTA. Subsequently, U251R cells were labeled
with 1 μl CD133/l microbeads per 1 million cells using the
CD133 Cell Isolation kit (MACS; Miltenyi Biotec, Bergisch-Gladbach,
Germany). Following separation, CD133+ and
CD133− cells were seeded into the same stem
cell-permissive medium. The efficiency of magnetic separation was
evaluated by flow cytometry with a fluorescence-activated cell
sorting (FACS) Calibur machine (BD Biosciences, Franklin Lakes, NJ,
USA) when the cells were incubated at 4°C for an additional 30 min
with 10 μl phycoerythrin (PE)-conjugated anti-CD133/2
(Miltenyi Biotec).
MTT assay
CD133+ neurospheres or adherent
non-sphere forming cells were prepared into single cell suspension
and seeded in 96-well plates at 1,000 cells/well in 200 μl
stem cell-permissive medium. MTT (20 μl) solution (final
concentration: 0.5 mg/ml; Sigma-Aldrich) was added at 0, 24, 48, 72
and 96 h following substitution of the medium in the presence or
absence of TMZ. Subsequently, the cells were incubated for 4 h and
the medium was replaced with 150 μl DMSO. The plates were
agitated for 15 min and the optical density of the solution in the
wells was measured at 490 nm on a Versamax microplate reader
(Bio-Rad, Hercules, CA, USA).
RNA isolation and reverse transcription
polymerase chain reaction (RT-PCR)
For RNA isolation, Invisorb Spin Tissue RNA Mini kit
250 (Invitek, Berlin, Germany) was used according to the
manufacturer’s instructions. The quantity and purity of the RNA was
estimated spectrophotometrically (UV-2450; Shimadzu Corp., Kyoto,
Japan) and the integrity was assessed by electrophoresis on a 1%
agarose gel. RNA was reverse transcribed using the Omniscript RT
kit (Qiagen, Hilden, Germany). The following primers were used:
MGMT, forward 5′-GTGAAATGAAACGCACCACAC-3′ and reverse
5′-GGAACTCTTCGATAGCCTCGG-3′; Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) gene (served as an internal control), forward
5′-ACACCCACTCCTCCACCTTT-3′ and reverse
5′-TAGCCAAATTCGTTGTCATACC-3′. The amplicon size for GAPDH was 225
bp and MGMT was 120 bp. The PCR conditions for MGMT genes were 29
cycles at 94°C for 45 sec, 54°C for 30 sec followed by 72°C for 45
sec. The PCR conditions for GAPDH were 30 cycles at 94°C for 30
sec, 54°C for 30 sec followed by 72°C for 1 min. Amplified PCR
products were electrophoresed on a 1.5% (w/v) agarose gel
(Sigma-Aldrich) containing 0.5 μg/ml ethidium bromide.
Relative levels of the target genes were quantified using a
computer-aided imaging analysis system, AlphaEaseFC™ (Alpha
Innotech Corp., San Leandro, CA, USA).
Western blot analysis
Cultured cells were washed with PBS and protein was
extracted using a lysis buffer (50 mM HEPES pH 8.0, 50 mM NaCl, 1%
Triton X-100, 10% glycerol, 1 mM MgCl2, 1.5 mM EDTA, 20
mM β-glycerophosphate, 50 mM NaF, 1 mM Na3VO4, 10 μg/ml
aprotinin, 1 μM pepstatin A, 1 mM phenylmethylsulphonyl
fluoride). The protein concentrations were determined using a DC
protein assay kit (Bio-Rad). Whole cell lysates containing 50
μg of protein were fractionated using 12% SDS-PAGE (Bio-Rad)
and transferred onto a nitrocellulose membrane (Bio-Rad). The
membrane was blocked with Tris-buffered saline plus 0.1% Tween 20
(Sigma-Aldrich) containing 5% non-fat milk for 1 h at room
temperature followed by incubation overnight with the following
primary antibodies: Mouse anti-human monoclonal anti-MGMT (1:500),
rabbit anti-human monoclonal anti-PUMA (1:500), rabbit anti-human
polyclonal anti-Bcl-2 (1:1,000), rabbit anti-human polyclonal
anti-Bax (1:1,000) and mouse anti-human monoclonal anti-GAPDH,
which were all purchased from Santa Cruz Biotechnology, Inc. (Santa
Cruz, CA, USA) at 4°C. The membrane was washed three times and
incubated with horseradish peroxidase-conjugated monoclonal goat
anti-rabbit immunoglobulin G (1:1,000) or rabbit anti-mouse
(1:1,000) secondary antibodies (Cell Signaling Technology, Inc.,
Beverly, MA, USA) for 1 h. Immunoreactive protein was detected
using chemiluminescence with Kodak X-AR film (Eastman Kodak,
Rochester, NY, USA).
Apoptosis assay
Trypsin was purchased from Sigma-Aldrich and the
concentration of trypsin was 0.25%. The cells were digested with
0.25% trypsin for 3–5 min at 37°C. Following trypsin enzyme
digestion, adherent and floating cells were harvested and washed
twice in ice-cold PBS. Subsequently, cells were resuspended at a
density of 1×106 cells/ml in 1X binding buffer
(Sigma-Aldrich) followed by the addition of 5 μl of Annexin
V-fluorescein isothiocyanate and 5 μl of propidium iodide
(Sigma-Aldrich). Following this, cells were incubated in the dark
at room temperature for 15 min, cell death was determined using a
flow cytometer (FACSCalibur; BD Biosciences). Data were analyzed
using CellQuest Pro software, version 6 (BD Biosciences).
Tumor xenografts
The present study was approved by the ethics
committee of the Institutional Animal Care and Use Committee of
Shanxi Medical University (Taiyuan, China) and all procedures were
conducted in accordance with the criteria outlined in the
international guidelines for the care and treatment of laboratory
animals. A total of 24, 4-week-old female BALB/C-nude mice weighing
18–20 g were purchased from Hunan Silaikejingda Laboratory Animal
Technology Co., Ltd. (Changsha, China) and housed in pathogen-free
conditions. The mice were subjected to subcutaneous inoculation
into the bilateral axillae with a total of 4×106 (200
μl) U251R cells per mouse. Treatment was initiated when the
tumors reached a mean volume of 30–40 mm3. The nude mice
were randomly divided into the following four groups (n=6): i) PBS
control; ii) TMZ; iii) Ad-PUMA and iv) TMZ+Ad-PUMA. A total of 50
μl PBS alone or Ad-PUMA (5×108 PFU/ml) in 50
μl PBS was administered by direct injection into the tumor
and TMZ (150 mg/m2/day) was administered by gastric
catheter. Subsequently, treatment was administered once every 3
days and was provided five times in total. Tumor growth was
monitored every other day. Tumor volume (V) was calculated as
(LxW2)/2, where L=length (mm) and W=width (mm) as
described previously (30). On day
40, animals were euthanized and tumors were excised and weighed.
The tumor specimens of nude mice were fixed in formalin, embedded
in paraffin and cut 4.0 μm in thickness. Sections were
stained with hematoxylin and eosin and subjected to terminal
deoxynucleotidyl transferase biotin-dUTP nick end labeling (TUNEL)
staining for detection of apoptotic cells using the TUNEL apoptotic
detection kit (Upstate Biotechnology, Inc., Lake Placid, NY, USA)
according to the manufacturer’s instructions.
Statistical analysis
All experiments were repeated at least three times.
The data are expressed as the mean ± standard deviation.
Statistical analysis was performed using IBM SPSS Statistics 19.0
software (SPSS, Inc., Chicago, IL, USA). Student’s t test or
one-way analysis of variance were performed to determine
significant differences. P<0.05 was considered to indicate a
statistically significant difference.
Results
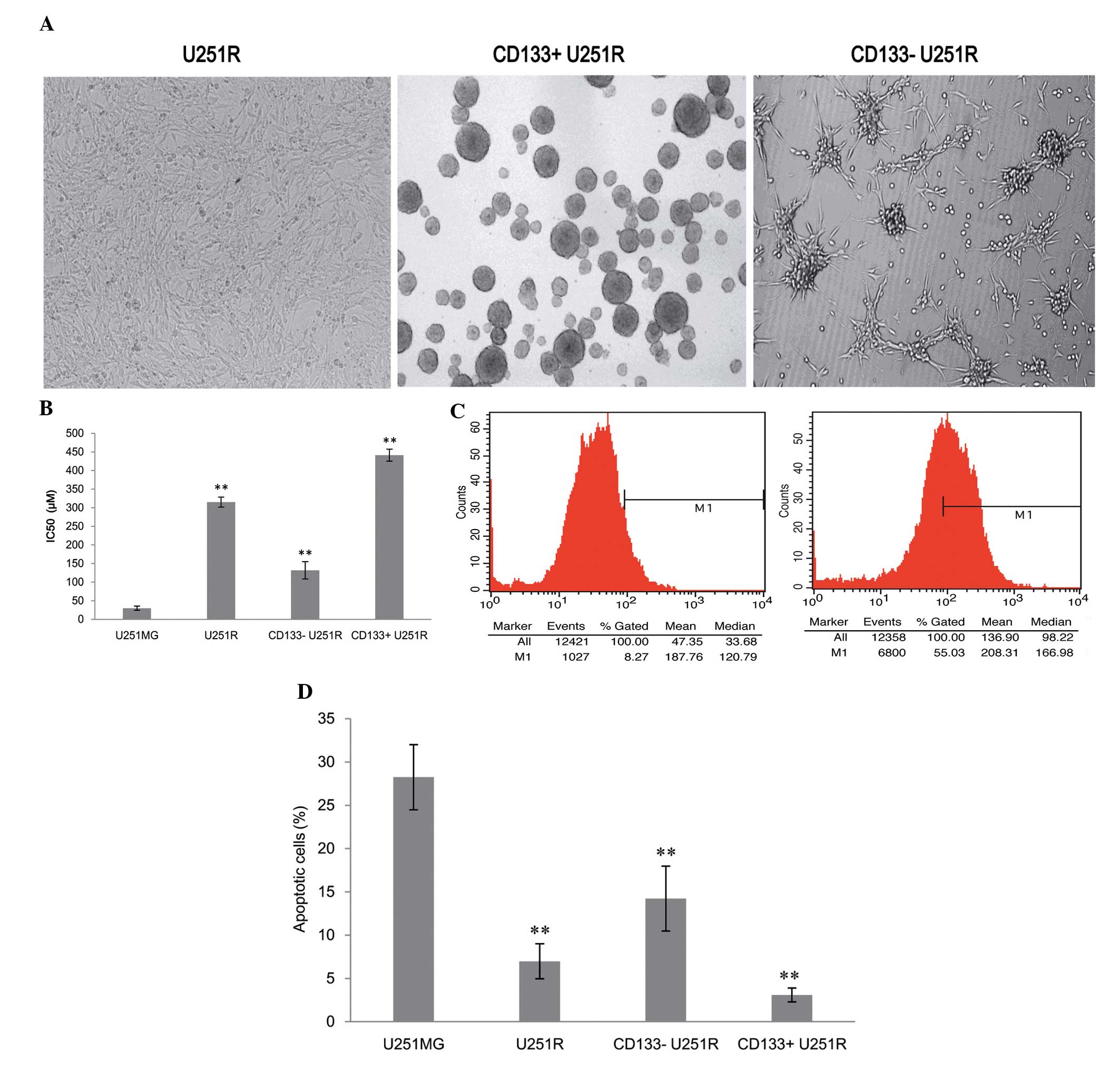
Drug-resistant GBM cell line, U251R may
be subclassified according to its CD133 immunophenotype
U251MG cells were exposed to TMZ from an initial
induction dose of 1.25 μmol/l to a final dose of 100
μmol/l. After 6 months, the stable TMZ-resistant variant was
obtained by progressively increasing the concentration of TMZ,
termed U251R. Subsequently, the U251R cells were isolated with a
magnetic activated cell sorting (MACS) kit based on the CD133
immunophenotype. The cells were divided into two groups,
CD133+ U251R cells and CD133− U251R cells.
The two groups of cells were cultured for ~2–3 days in stem cell
media. Floating tumor neurospheres were observed in
CD133+ U251R cells whereas CD133− U251R cells
were attached to the bottom of the wells with no tumor spheres
forming (Fig. 1A). Their
chemosensitivity was evaluated by MTT assay and U251R cells
exhibited apparent resistance to TMZ compared with parent U251MG
cells (P<0.05; Fig. 1B).
CD133+ U251R cells are more
resistant to TMZ-induced cell growth inhibition and apoptosis
The chemosensitivity of GBM cells to TMZ was
evaluated by MTT assay. The results revealed that compared with
parental U251MG cells (29.83±5.61 μM), the half maximal
inhibitory concentration (IC50) values of TMZ-treated cells
exhibited a 10.6-fold increase in U251R cells (315.20±13.38
μM), a 4.4-fold increase in CD133− U251R cells
(131.85±23.31 μM) and a 14.8-fold increase in
CD133+ U251R cells (441.24±16.27μM). This
indicated that CD133+ U251R cells possessed the
strongest drug resistance to TMZ-induced cell growth inhibition
(F=739.18; P<0.05; Fig. 1B).
This was further confirmed by flow cytometry analysis results
demonstrating that the percentage of CD133+ U251R cells
increased significantly (55.03%) compared with parental cells
U251MG (8.27%; P<0.05; Fig.
1C). Therefore, CD133+ cells or GSCs were enriched
in drug resistant U251R cells.
It is well established that resistance to apoptosis
is a hallmark of virtually all types of human cancer, including GBM
and is also involved in drug resistance. Therefore, in order to
analyze the effects of 200 μmol/l TMZ treatment for 48 h on
the apoptotic rates of U251MG, U251R, CD133+ U251R cells
or CD133− U251R cells, an annexin-V/propidium iodide
(PI) double-staining assay was performed. Compared with parent
U251MG cells, apoptotic rates were significantly decreased in the
other three groups (F=89.535; P<0.05). Apoptotic cells were
hardly detectable in CD133+ U251R (Fig. 1D).
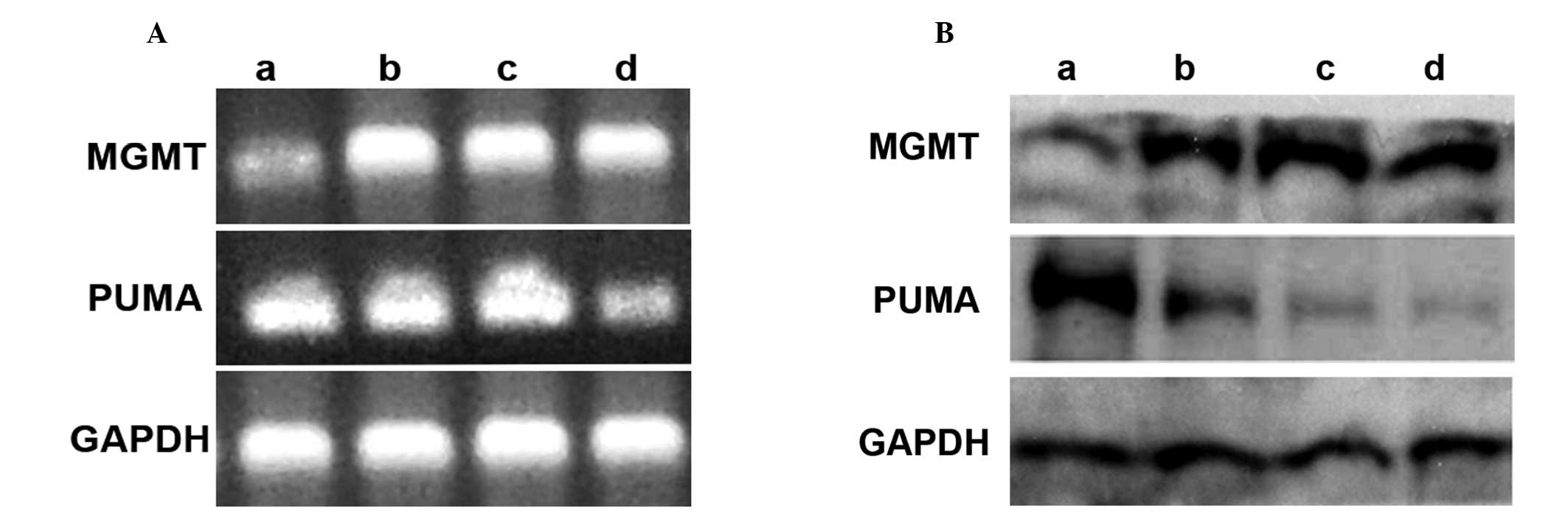
No significant differences are observed
in MGMT expression levels between CD133+ U251R and
CD133− U251R cells
Sensitivity of glioma cells to TMZ has been reported
to be significantly involved in the level of MGMT repair activity
in cells. Thus, the mRNA and protein expression of MGMT was
detected using RT-PCR and western blot analysis, respectively. The
results demonstrated that the mRNA and protein expression of MGMT
in U251MG was significantly lower than that in U251R,
CD133+ U251R or CD133− U251R cells
(P<0.05). However, no significant differences were identified in
the remaining three groups (P>0.05; Fig. 2). Considering that
CD133+ U251R cells are more resistant to TMZ-induced
cell growth inhibition and apoptosis, it was hypothesized that
there may be additional drug resistance mechanisms in
CD133+ U251R cells in addition to the mechanism of
MGMT-mediated resistance to TMZ.
Decreased PUMA expression may be in
accordance with drug-resistance of GSCs to TMZ
PUMA expression may be induced by p53 or
DNA-damaging agents. The expression of PUMA was analyzed and it was
observed that the three GBM drug-resistant variant cell lines
presented a reduced PUMA expression compared with their parental
U251MG cells (P<0.05). Notably, the PUMA protein or mRNA band of
the CD133+ U251R cell line group exhibited a lower
expression level than that of U251R and CD133− U251R
cells (P<0.05; Fig. 2). These
results suggested that PUMA-mediated apoptotic pathways may be
impaired in drug-resistant GSCs. Therefore, loss of PUMA may alter
the sensitivity of GBM cells to TMZ treatment.
Ad-PUMA markedly sensitizes
CD133+ U251R cells to TMZ through induction of
apoptosis
To investigate the hypothesis that exogenous
expression of PUMA may enhance sensitivity of CD133+
U251R cells to TMZ treatment, the recombinant adenovirus
Ad-DsRed-PUMA was used to treat the CD133+ U251R cells
at 100 MOI of the adenovirus. The infection efficacy (74.34%) of
Ad-PUMA to CD133+ U251R was assayed by FACS and red
color was observed in cells infected with Ad-DsRed-PUMA with
fluorescence microscopy, which indicated that the adenovirus may be
used in the treatment (Fig. 3A).
Western blot analysis results confirmed that PUMA expression level
was significantly upregulated in Ad-DsRed-PUMA infection groups
(P<0.05), while no statistical difference was identified between
the PBS control group and TMZ alone group (P>0.05; Fig. 3B). Following Ad-PUMA infection at
100 MOI for 2 h, another MTT assay was performed to investigate the
change in IC50 of CD133+ U251R cells to TMZ. The results
demonstrated that exogenous expression of PUMA decreases the IC50
by 3- to 8-fold compared with the IC50 of TMZ alone.
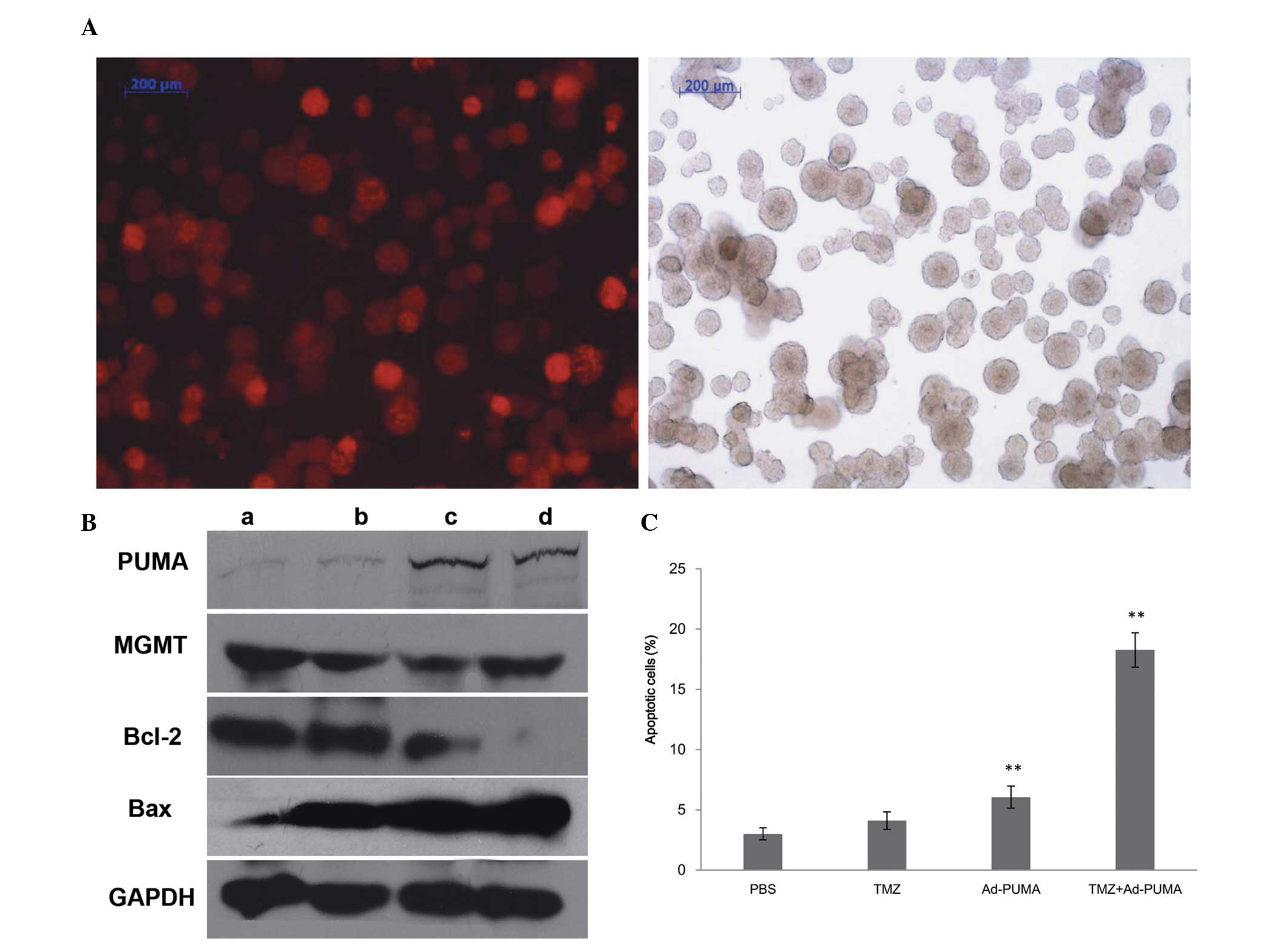 | Figure 3Ad-PUMA sensitizes CD133+
U251R cells to TMZ treatment through apoptosis induction. (A) In
the left picture, red fluorescence is observed in the
CD133+ U251R transfected with Ad-Dsred-PUMA. The right
picture shows the CD133+ U251R cells under normal white
light. (B) PUMA, MGMT, Bcl-2, Bax and internal control GAPDH
protein expression was analyzed by western blotting. (a) PBS
control; (b) 200 μmol/l TMZ treatment for 48 h; (c) 100
multiplicity of infection Ad-PUMA; (d) Ad-PUMA+TMZ. (C) Apoptosis
was quantitated by Annexin-V/propidium iodide assay following
different treatments. Values are presented as the mean ± standard
error of the mean. **P<0.05, compared with the PBS
control group. MGMT,
O6-methylguanine-DNA-methyltransferase; PUMA, p53
upregulated modulator of apoptosis; AD-PUMA, adenoviruses
expressing wild-type PUMA; PBS, phosphate-buffered saline; Bcl-2,
B-cell lymphoma 2; TMZ, temozolomide; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase; Bax, Bcl-2-associated X
protein. |
Furthermore, as shown in Fig. 3C, following TMZ treatment in the
presence or absence of Ad-PUMA, cells were harvested and analyzed
for levels of apoptotic cells using FACS. The result demonstrated
that few apoptotic cells were observed in CD133+ U251R
cells solely treated with TMZ or PBS. However, the proportion of
apoptotic cells was significantly increased in the Ad-PUMA group
and the Ad-PUMA in combination with TMZ group (F=329.433;
P<0.05).
PUMA significantly sensitizes GBM cells
to TMZ through elevated Bcl-2-associated X protein (Bax)/Bcl-2
ratio in a MGMT-independent manner
In order to further characterize the PUMA-induced
apoptosis in TMZ-resistant cells, the changes in three proteins,
MGMT, Bcl-2 and Bax, which were involved in the PUMA-induced
apoptotic process were examined. As shown in Fig. 3B, it was observed that when TMZ or
PBS alone was used to treat cells, the Bax/Bcl-2 ratio did not
alter in CD133+ U251R cells (P>0.05). However, when
TMZ combined with Ad-PUMA or Ad-PUMA alone was used to treat cells,
the Bax/Bcl-2 ratio was markedly increased, indicating that PUMA is
significantly associated with apoptosis when combined with TMZ
treatment (P<0.05). In addition, as shown in Fig. 3B, compared with control cells, the
expression level of MGMT was not statistically significant
(P>0.05). These data suggested that the chemoresistance of
CD133+ U251R cells to TMZ did not only result from the
deregulation of MGMT. In addition, the ability of PUMA to induce
apoptosis in malignant glioma cells may be independent of MGMT.
Possible mechanisms through which PUMA induces apoptosis or
sensitizes GBM cells to TMZ may be associated with an elevated
Bax/Bcl-2 ratio in an MGMT-independent manner.
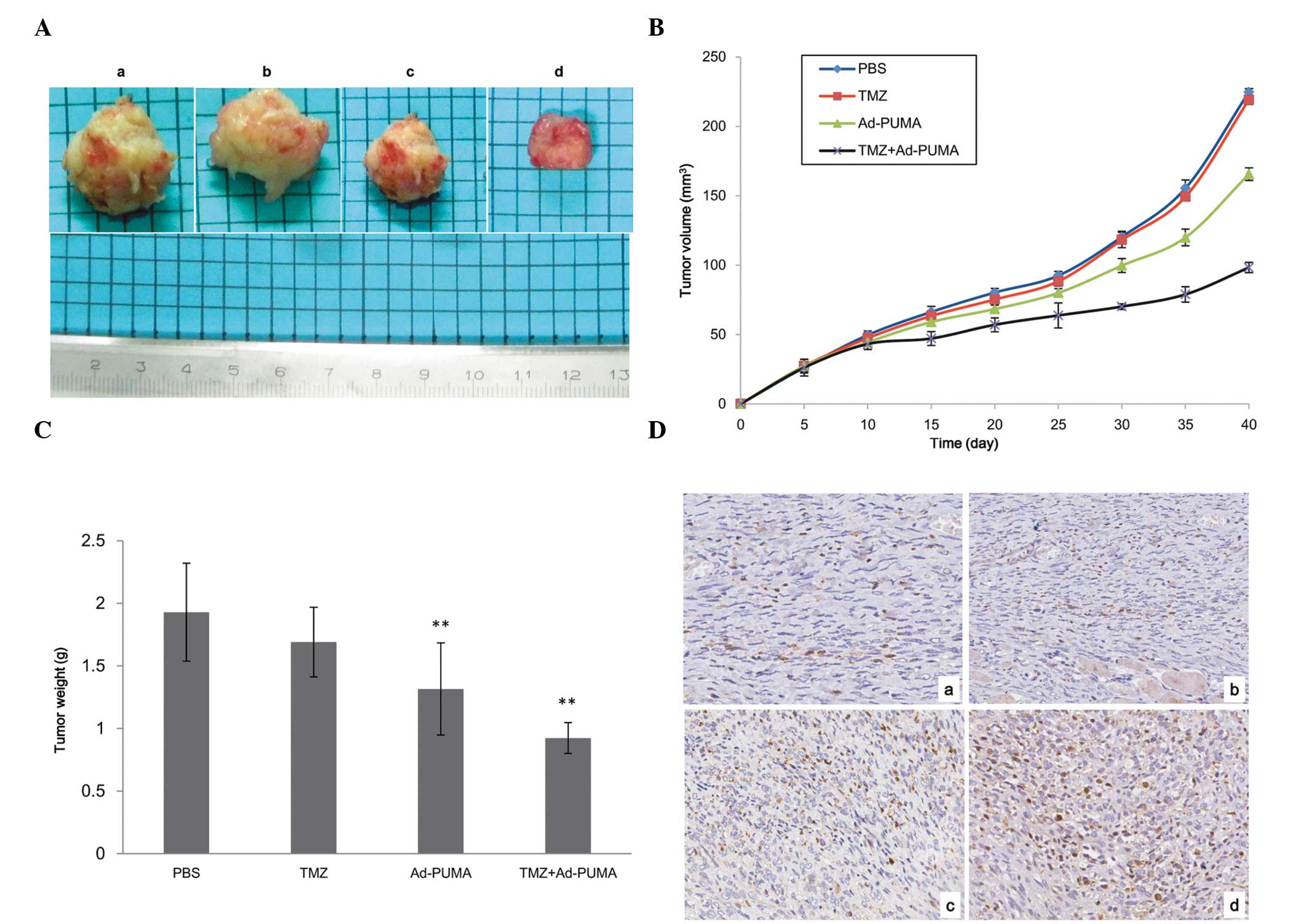
Effect of TMZ-based chemotherapy combined
with Ad-PUMA therapy on transplanted tumors induced by U251R in
vivo
Following on from in vitro experiments, which
revealed that Ad-PUMA sensitizes the drug resistant glioma cells to
TMZ treatment, it was further investigated whether this
sensitization effect may also be detected in vivo in tumor
xenograft animal models. U251R cells were injected subcutaneously
into the bilateral axillae of nude mice and secondary tumors were
observed in all injected mice following cell inoculation.
Subsequently, tumors initiated by U251R cells were treated with
PBS, TMZ alone, Ad-PUMA alone and combined TMZ plus Ad-PUMA,
respectively. As shown in Fig. 4A and
B, the average tumor volume in the Ad-PUMA+TMZ group and the
Ad-PUMA group 40 days after transplantation was smaller than the
other two groups (P<0.05). Ad-PUMA combined with TMZ suppressed
the growth of subcutaneous tumors more potently than Ad-PUMA alone.
Similarly, tumors treated with Ad-PUMA in combination with TMZ were
significantly lighter than the remaining three groups (P<0.05;
Fig. 4C). In addition, tumor
sections were stained using a TUNEL kit to evaluate the rates of
apoptosis. The results confirmed that Ad-PUMA may induce apoptosis
of xenograft tumors alone by enhanced apoptosis induced by TMZ
treatment. By contrast, apoptotic cells were almost undetectable in
tumors of PBS control treatment or the TMZ alone group (Fig. 4D). These results suggested that
U251R tumors possess significant drug resistance to TMZ and Ad-PUMA
may only partly reverse TMZ drug-resistance of tumors initiated by
U251R cells to TMZ. Once Ad-PUMA was used to infect the tumors,
which were simultaneously treated with TMZ, the tumor volume and
weight were markedly decreased, suggesting that the combination of
TMZ with Ad-PUMA was effective and its antitumor effect may be due
to induction of apoptosis.
Discussion
The alkylating agent, TMZ is currently the first
choice chemotherapy drug for GBM treatment with the best clinical
efficacy (31). However,
endogenous or acquired resistance to TMZ limits the clinical
outcome of GBM patients and is also an important cause of GBM
relapse (32). Therefore, in-depth
understanding of TMZ resistance mechanisms is key to overcoming TMZ
resistance and improving the prognosis of patients with GBM.
Through intervening in associated mechanisms, tumor cell
sensitivity to TMZ may be restored (33).
With the emergence of the cancer stem cell theory,
the aim of therapeutics shifted toward eradicating cancer stem
cells (34,35). Significant research efforts have
been devoted to understanding and overcoming this critical issue.
Therefore, understanding and exploiting the chemoresistance
mechanisms of GSCs to TMZ treatment in gliomas is an important
strategy for the development of new targeted therapies. In the
present study, drug resistant GBM cells, U251R, induced through
progressive increases in TMZ treatment for 6 months were developed.
Cells were separated into two subpopulations by MACS based on the
CD133 immunophenotype and subsequently cultured in stem
cell-permissive medium. It was observed that the growth rate of
CD133− U251R cells was slower than that of
CD133+ U251R cells. In addition, it was also noted that
the IC50 of CD133+ U251R cells was significantly higher
than CD133− U251R, U251 MG and U251R cells. FACS
analysis suggested that CD133+ cells are abundant in
U251R cells. Notably, no statistical differences in the expression
of MGMT were identified between CD133+ U251R and
CD133− U251R cells, although the two exhibited higher
expression levels than U251 MG cells. This demonstrated that the
resistance of CD133+ U251R against TMZ is not limited to
MGMT expression levels and there may be additional mechanisms. Liu
et al (22) also found that
CD133+ cells in GBM were more resistant to TMZ,
paclitaxel and VPl6 than the negative cells, which may be induced
by the higher expression levels of breakpoint cluster region
pseudogene 1 and MGMT, as well as the anti-apoptosis protein and
inhibitors of apoptosis protein families in CD133+
cells.
Prevention of apoptosis is a hallmark of cancer and
confounds the efficacy of cancer chemotherapy (24). TMZ exerts its antitumor effects
predominantly through cell death induced by DNA double-strand
breaks (6). Whether apoptosis
failed in CD133+/− U251R cells following TMZ treatment
was investigated. In the current study, when compared with parent
U251MG, the apoptotic rate of CD133+/− U251R cells was
significantly decreased. In order to illustrate this mechanism,
changes in the apoptosis-associated molecule, PUMA in U251MG,
U251R, CD133− U251R and CD133+ U251R were
further examined. Western blot analyses indicated that compared
with the parental U251MG cells, PUMA expression levels were
significantly decreased in CD133+/− U251R cells,
particularly in CD133+ U251R cells. This suggested that
CD133+ U251R cells or GSCs mainly contributed to the
resistance to TMZ, at least partly, through deregulated PUMA
induction.
Wild type p53 protein is established as a potent
tumor suppressor (36). However,
p53 mutations are common in diverse types of human cancer,
including GBM (37). The mutations
not only result in p53 protein loss of wild-type function, in the
majority of cases, mutant p53 protein acquires an oncogenic gain of
function (38–41). PUMA, a p53 downstream molecule has
a pro-apoptotic role independent of p53 status. It may destroy
cancer cells through interacting with Bcl-2 family members, even
when the p53 pathway is dysfunctional (42). Yu et al (25) demonstrated that exogenous
expression of PUMA protein resulted in a more rapid and profound
apoptosis. Ito et al (27)
confirmed previous findings and revealed that, regardless of p53
status, PUMA expression was directly associated with tumor cell
apoptosis. Overexpression of PUMA may lead to a large number of
apoptotic cells, which was also accompanied by mitochondrial damage
and caspase activation. In a further study, Yu et al
(43) combined Ad-PUMA with
paclitaxel, fluorouracil, cisplatin and etoposide in the treatment
of human lung adenoma A549 cells. Ad-PUMA was demonstrated to
increase sensitivity of A549 cells to common chemotherapeutic drugs
and radiation (43). Ad-PUMA
appears to be selectively toxic to cancer cells and more efficient
than p53 in inhibiting the growth of cancer cells. It was therefore
hypothesized that exogenous PUMA may cause reversion of U251R cells
resistant to TMZ. In our subsequent experiments, the analysis of
apoptosis by FACS indicated that there were few or no apoptotic
cells in the CD133+ U251R cells treated with TMZ,
however, introduction of exogenous Ad-PUMA not only inhibited cell
growth, but also increased their apoptotic rates. It is worth
noting that, when combined with TMZ, Ad-PUMA may induce significant
growth inhibition and apoptotic responses. Furthermore, using a
nude mouse subcutaneous glioma model, the effect of Ad-PUMA
combined with TMZ on glioma growth was observed. Consistent with
in vitro results, Ad-PUMA and TMZ had a synergistic effect
on suppressing tumor growth in vivo. These results suggested
that Ad-PUMA has prospects for application in GBM for TMZ-based
chemotherapy.
Previous studies have demonstrated that TMZ
treatment alters the expression of pro-apoptotic Bax and
anti-apoptotic Bcl-2 involved in the mitochondrial pathway of
apoptosis (44,45). Additionally, the ratio of Bax/Bcl-2
correlated with chemotherapy resistance and shortened overall
survival (46). Western blot
analysis indicated that the ratio of Bax/Bcl-2 was partially
decreased in TMZ-resistant cells. However, once these cells were
infected with Ad-PUMA, the ratio of Bax/Bcl-2 was significantly
upregulated. This demonstrated that PUMA may induce apoptosis
through increasing the ratio of Bax/Bcl-2, as high levels of
Bax/Bcl-2 is established as an inhibitor of apoptosis (47,48).
In addition, this process had no association with MGMT as MGMT
protein expression levels did not change.
Together, the present results demonstrate that
CD133+ U251R cells were more resistant to TMZ treatment
than CD133− U251R cells although no significant
differences were observed in MGMT protein expression levels between
them, the major mediator of TMZ resistance in GBM. By contrast, it
was noted that PUMA expression in CD133+ U251R cells, a
crucial factor in the apoptosis of human glioma cells induced by
antitumor drugs, was significantly lower than the other cell lines
investigated. This explained, at least partly, why apoptosis was
defective in CD133+ U251R cells or GSCs. These results
suggest a requirement for new strategies against glioma resistance
to TMZ. CD133+ U251R cells may be a target for future
studies. In vitro and in vivo data suggested that the
combination of Ad-PUMA and TMZ is a promising strategy for treating
malignant gliomas resistant to chemotherapy. In addition, given the
complexity of the p53 pathophysiological network in apoptosis,
senescence, growth arrest and the high mutation rate in GBM,
approaches of gene therapy that involve increasing PUMA expression
may be an improved approach to reverse TMZ resistance in glioma
cells. This may induce glioma cell death regardless of p53 status
and possess the same efficacy for inducing apoptosis in
CD133+ tumor cells.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81201991 and 81470115), the
Science and Technology Research Program of Shanxi Province (grant
no. 20110313011-3), the Basic Research Program of Shanxi Province
(grant no. 2012021035-4) and the Doctoral Foundation of the First
Hospital of Shanxi Medical University (grant no. YB1109).
References
|
1
|
Furnari FB, Fenton T, Bachoo RM, Mukasa A,
Stommel JM, et al: Malignant astrocytic glioma: genetics, biology,
and paths to treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Omuro A and DeAngelis LM: Glioblastoma and
other malignant gliomas: a clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Hegi ME, Gilbert MR and
Chakravarti A: Chemoradiotherapy in malignant glioma: standard of
care and future directions. J Clin Oncol. 25:4127–4136. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, et al: Radiotherapy plus concomitant and adjuvant
temozolomide for glioblastoma. N Engl J Med. 352:987–996. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pegg AE: Repair of O(6)-alkylguanine by
alkyltransferases. Mutat Res. 462:83–100. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roos WP and Kaina B: DNA damage-induced
cell death: from specific DNA lesions to the DNA damage response
and apoptosis. Cancer Lett. 332:237–248. 2013. View Article : Google Scholar
|
|
7
|
Kaina B, Christmann M, Naumann S and Roos
WP: MGMT: key node in the battle against genotoxicity,
carcinogenicity and apoptosis induced by alkylating agents. DNA
Repair (Amst). 6:1079–1099. 2007. View Article : Google Scholar
|
|
8
|
Sabharwal A and Middleton MR: Exploiting
the role of O6-methylguanine-DNA-methyltransferase (MGMT) in cancer
therapy. Curr Opin Pharmacol. 6:355–363. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Day RS III, Ziolkowski CH, Scudiero DA,
Meyer SA, Lubiniecki AS, et al: Defective repair of alkylated DNA
by human tumour and SV40-transformed human cell strains. Nature.
288:724–7. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kondo N, Takahashi A, Ono K and Ohnishi T:
DNA damage induced by alkylating agents and repair pathways. J
Nucleic Acids. 2010:5435312010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fruehauf JP, Brem H, Brem S, Sloan A,
Barger G, et al: In vitro drug response and molecular markers
associated with drug resistance in malignant gliomas. Clin Cancer
Res. 12:4523–4532. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hermisson M, Klumpp A, Wick W, Wischhusen
J, Nagel G, et al: O6-methylguanine DNA methyltransferase and p53
status predict temozolomide sensitivity in human malignant glioma
cells. J Neurochem. 96:766–776. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dalerba P, Cho RW and Clarke MF: Cancer
stem cells: models and concepts. Annu Rev Med. 58:267–284. 2007.
View Article : Google Scholar
|
|
15
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, et al: Identification of a cancer stem cell in human
brain tumors. Cancer Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
16
|
Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, et
al: Glioma stem cells promote radioresistance by preferential
activation of the DNA damage response. Nature. 444:756–760. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Wakeman TP, Lathia JD, Hjelmeland
AB, Wang XF, et al: Notch promotes radioresistance of glioma stem
cells. Stem Cells. 28:17–28. 2010.
|
|
18
|
Hardee ME, Marciscano AE, Medina-Ramirez
CM, Zagzag D, Narayana A, et al: Resistance of
glioblastoma-initiating cells to radiation mediated by the tumor
microenvironment can be abolished by inhibiting transforming growth
factor-β. Cancer Res. 72:4119–4129. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deleyrolle LP, Harding A, Cato K,
Siebzehnrubl FA, Rahman M, et al: Evidence for label-retaining
tumour-initiating cells in human glioblastoma. Brain.
134:1331–1343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang MK and Kang SK: Tumorigenesis of
chemotherapeutic drug-resistant cancer stem-like cells in brain
glioma. Stem Cells Dev. 16:837–847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bleau AM, Huse JT and Holland EC: The
ABCG2 resistance network of glioblastoma. Cell Cycle. 8:2936–2944.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu G, Yuan X, Zeng Z, Tunici P, Ng H, et
al: Analysis of gene expression and chemoresistance of CD133+
cancer stem cells in glioblastoma. Mol Cancer. 5:672006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Li Y, Yu TS, McKay RM, Burns DK,
et al: A restricted cell population propagates glioblastoma growth
after chemotherapy. Nature. 488:522–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Martinez-Rivera M and Siddik ZH:
Resistance and gain-of-resistance phenotypes in cancers harboring
wild-type p53. Biochem Pharmacol. 83:1049–1062. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu J, Zhang L, Hwang PM, Kinzler KW and
Vogelstein B: PUMA induces the rapid apoptosis of colorectal cancer
cells. Mol Cell. 7:673–682. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu J and Zhang L: PUMA, a potent killer
with or without p53. Oncogene. 27(Suppl 1): S71–S83. 2001.
View Article : Google Scholar
|
|
27
|
Ito H, Kanzawa T, Miyoshi T, Hirohata S,
Kyo S, et al: Therapeutic efficacy of PUMA for malignant glioma
cells regardless of p53 status. Hum Gene Ther. 16:685–698. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nagane M, Kobayashi K, Ohnishi A, Shimizu
S and Shiokawa Y: Prognostic significance of O6-methylguanine-DNA
methyltransferase protein expression in patients with recurrent
glioblastoma treated with temozolomide. Jpn J Clin Oncol.
37:897–906. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He TC, Zhou S, da Costa LT, Yu J, Kinzler
KW and Vogelstein B: A simplified system for generating recombinant
adenoviruses. Proc Natl Acad Sci USA. 95:2509–2514. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mukai S, Kondo Y, Koga S, Komata T, Barna
BP, et al: 2-5A antisense telomerase RNA therapy for intracranial
malignant gliomas. Cancer Res. 60:4461–4467. 2000.PubMed/NCBI
|
|
31
|
Villano JL, Seery TE and Bressler LR:
Temozolomide in malignant gliomas: current use and future targets.
Cancer Chemother Pharmacol. 64:647–655. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beier D, Schulz JB and Beier CP:
Chemoresistance of glio-blastoma cancer stem cells - much more
complex than expected. Mol Cancer. 10:1282008. View Article : Google Scholar
|
|
33
|
Koritzer J, Boxhammer V, Schafer A,
Shimizu T, Klampfl TG, et al: Restoration of sensitivity in
chemo-resistant glioma cells by cold atmospheric plasma. PLoS One.
8:e644982013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Francipane MG, Chandler J and Lagasse E:
Cancer stem cells: A moving target. Curr Pathobiol Rep. 1:111–118.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Medema JP: Cancer stem cells: the
challenges ahead. Nat Cell Biol. 15:338–344. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Smith ND, Rubenstein JN, Eggener SE and
Kozlowski JM: The p53 tumor suppressor gene and nuclear protein:
basic science review and relevance in the management of bladder
cancer. J Urol. 169:1219–1228. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hollstein M, Sidransky D, Vogelstein B and
Harris CC: p53 mutations in human cancers. Science. 253:49–53.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Petitjean A, Mathe E, Kato S, Ishioka C,
Tavtigian SV, et al: Impact of mutant p53 functional properties on
TP53 mutation patterns and tumor phenotype: lessons from recent
developments in the IARC TP53 database. Hum Mutat. 28:622–629.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Goh AM, Coffill CR and Lane DP: The role
of mutant p53 in human cancer. J Pathol. 223:116–126. 2011.
View Article : Google Scholar
|
|
40
|
Brosh R and Rotter V: When mutants gain
new powers: news from the mutant p53 field. Nat Rev Cancer.
9:701–713. 2009.PubMed/NCBI
|
|
41
|
Oren M and Rotter V: Mutant p53
gain-of-function in cancer. Cold Spring Harb Perspect Biol.
2:a0011072010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Haupt S, Berger M, Goldberg Z and Haupt Y:
Apoptosis - the p53 network. J Cell Sci. 116:4077–4085. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yu J, Yue W, Wu B and Zhang L: PUMA
sensitizes lung cancer cells to chemotherapeutic agents and
irradiation. Clin Cancer Res. 12:2928–2936. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ma J, Murphy M, O’Dwyer PJ, Berman E, Reed
K, et al: Biochemical changes associated with a multidrug-resistant
phenotype of a human glioma cell line with temozolomide-acquired
resistance. Biochem Pharmacol. 63:1219–1228. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Manero F, Gautier F, Gallenne T, Cauquil
N, Gree D, et al: The small organic compound HA14-1 prevents Bcl-2
interaction with Bax to sensitize malignant glioma cells to
induction of cell death. Cancer Res. 66:2757–2764. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tothova E, Fricova M, Stecova N, Kafkova A
and Elbertova A: High expression of Bcl-2 protein in acute myeloid
leukemia cells is associated with poor response to chemotherapy.
Neoplasma. 49:141–144. 2002.PubMed/NCBI
|
|
47
|
Gielen PR, Aftab Q, Ma N, Chen VC, Hong X,
et al: Connexin43 confers Temozolomide resistance in human glioma
cells by modulating the mitochondrial apoptosis pathway.
Neuropharmacology. 75:539–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Eisele G and Weller M: Targeting apoptosis
pathways in glioblastoma. Cancer Lett. 332:335–345. 2013.
View Article : Google Scholar
|