Introduction
Adiponectin, derived from adipocytes, is a
circulating plasma protein. It has been observed to have a number
of effects on metabolism, including insulin sensitization and
vascular protective properties (1). Human studies have revealed that in
obese individuals plasma adiponectin levels are significantly
decreased (2), as is the case in
patients with insulin resistance (3) and Type 2 diabetes (4). Insulin resistance may be induced in
mice via targeted deletion of the adiponectin gene (5), whereas increasing the expression of
adiponectin resulted in a greater level of insulin sensitivity
(3,6). Such findings have led to the
hypothesis that adiponectin is an anti-inflammatory,
insulin-sensitizing adipokine, which may have protective effects in
metabolic diseases associated with obesity.
The adiponectin receptors 1 and 2 (AdipoR1 and
AdipoR2) mediate the physiological effects of adiponectin in the
liver and in the skeletal muscle, regarding the metabolism of
glucose and lipids (7). The
adiponectin receptors are 7-transmembrane proteins. The signaling
pathways responsible for the effects of adiponectin on metabolism
remain to be fully elucidated. The adiponectin-activated
AMP-activated protein kinase (AMPK), peroxisome
proliferator-activated receptor (PPAR)-α, and the p38
mitogen-activated protein kinase (MAPK) pathways (7) have all been revealed to involve
AdipoR1 and AdipoR2, and to mediate the metabolism of glucose and
lipids. However, the mechanism, which regulates the expression of
the adiponectin receptors remains to be elucidated.
MicroRNAs (miRNAs) are a class of 22 nucleotide
noncoding RNAs, which regulate gene expression at the
post-transcriptional level. miRNAs silence their target genes by
cleaving mRNA molecules or inhibiting their translation and thus
regulate various physiological and pathological processes (8). A previous study indicated that miRNAs
act as key regulators in insulin signaling and glucose metabolism
(9). For example, miR-375 was
observed to suppress glucose-induced insulin secretion (10). Other miRNAs, including Let-7
(11,12), miR-103, miR-107 (13) and miR-29a/b (14) were revealed to regulate insulin
sensitivity in the liver and peripheral tissues by controlling the
expression of numerous components of the insulin signaling pathway.
In the present study, it was observed that miR-218 inhibits
adiponectin signaling and glucose uptake by targeting AdipoR2.
Materials and methods
Cell culture
HepG2 cells (American Type Culture Collection,
Manassas, VA, USA) were maintained in Dulbecco’s modified Eagle’s
medium (DMEM; Invitrogen Life Technologies, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (Hyclone, Waltham, MA,
USA) at 37°C.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol reagent
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. Equal amount of total RNA (1 μg) were reverse
transcribed using a High-Capacity cDNA Reverse Transcription kit
(Applied Biosystems, Foster City, CA, USA). qPCR analysis was
performed in 10 μl reactions using the ABI7900 Real-Time PCR
system (Applied Biosystems) and SYBR Green PCR Master Mix (Applied
Biosystems). The cycling conditions were as follows: Initial
denaturation at 95°C for 30 sec, then 40 cycles of denaturation at
95°C for 10 sec and annealing and extension at 60°C for 30 sec. The
following primers (Sangon Biotech Co., Ltd., Shanghai, China) were
used: AdipoR2 forward, 5′-CCCAAGAAGTCCGAGACACG-3′ and reverse,
5′-TGTTGGCTCGTTCATGGGAT-3′; and β-actin forward,
5′-TTGCGTTACACCCTTTCTTG-3′ and reverse, 5′-CACCTTCACCGTTCCAGTTT-3′.
Expression of AdipoR2 was normalized relative to β-actin using the
2−ΔΔCT method.
Plasmid construction
The wild-type 3′-untranslated region (3′-UTR) of the
AdipoR2 gene containing the predicted miR-218 target sites was
amplified using polymerase chain reaction (PCR) from HepG2 cDNA
using the following primer set (Sangon Biotech Co., Ltd.): AdipoR2,
forward 5′-GCTCTAGATACCTACCAGTCTCCAGG-3′ and reverse
5′-GCTCTAGATAGAAACTGAGAATACAGC-3′. The 3′-UTR fragment was cloned
downstream of the firefly luciferase coding region in the
XbaI site of the pGL3-control plasmid (Promega Corporation,
Madison, WI, USA).
Transfection
HepG2 cells were transiently transfected with 50 nM
of the chemically synthesized miR-218, 5′-UUGUGCUUGAUCUAACCAUGU-3′,
or negative control miRNA, 5′-UUCUCCGAACGUGUCACGU-3′ (Genepharma,
Shanghai, China), using Lipofectamine 2000 (Invitrogen Life
Technologies) according to the manufacturer’s instructions. After
24 h transfection, cells were used for subsequent
experimentation.
Dual-luciferase reporter assay
HepG2 cells were co-transfected with
pGL3-AdipoR2-3′UTR together with an RNA oligonucleotide. Equal
amount of renilla luciferase were co-expressed as the internal
control. The cell extracts were prepared 24 h after transfection
and the luciferase activity was measured using the dual-luciferase
reporter assay system (Promega Corporation). The firefly luciferase
activity was normalized to Renilla luciferse activity
(15).
Immunoblotting
The cells were washed twice with ice-cold
phosphate-buffered saline (PBS) and lysed in ice-cold lysis buffer
(100 mM Tris-HCl, pH 7.5; 300 mM NaCl, 1% NP-40 and 1 mM DTT)
containing protease inhibitor cocktail tablets (Cell Signaling
Technology, Beverly, MA, USA). The cell lysates were resuspended in
SDS loading buffer, heated at 95°C for 5 min and separated using
SDS-PAGE. Immunocomplexes were analyzed via immunoblotting using a
polyclonal goat anti-AdipoR2 antibody (sc-46751; 1:1,000; Cell
Signaling Technology), followed by a polyclonal donkey anti-goat
immunoglobulin G conjugated with horseradish peroxidase (sc-2020;
1:1,000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA).
Immunodetection was performed with an enhanced chemiluminescence
kit (Cell Signaling Technology).
Glucose uptake assay
HepG2 cells cultured on 96-well black plates were
transfected with miR-218 or negative control siRNA. At 48 h after
transfection, HepG2 cells were washed with DMEM three times and
incubated with or without 10 μg/ml recombinant adiponectin
(R&D Systems, Inc. Minneapolis, MN, USA) for 30 min. Following
adiponectin incubation, cells were incubated with 50 μM
2-(N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl) amino)-2-deoxyglucose
(2-NBDG; Invitrogen Life Technologies) in PBS for 15 min and then
washed with additional PBS to remove excess 2-NBDG. The levels of
fluorescence in the cells were measured at an excitation wavelength
of 485 nm and an emission wavelength of 535 nm with Tecan infinite
M200 (Tecan Group Ltd., Seestrasse, Switzerland). The fluorescence
of the control group transfected with negative control and not
treated with adiponectin was normalized to 1.0. The fluorescence of
other groups was compared with the control group.
Statistical analysis
The data are presented as the mean ± standard
deviation. Statistical differences between groups were determined
using Student’s t-test. All statistical analyses were performed
using Microsoft Excel 2010 (Microsoft Corp., Redmond, WA, USA).
P<0.01 was considered to indicate a statistically significant
difference.
Results
miR-218 targets AdipoR2 mRNA
To identify potential AdipoR2-targeting miRNAs, an
in silico search was initially performed using publicly
available algorithms, including TargetScan (www.targetscan.org) and miRanda (www. microrna.org). miR-218 was identified as a
candidate miRNA for targeting of AdipoR2. Phylogenetic analysis
revealed that miR-218 is well-conserved among different species,
ranging from Canis familiaris to Homo sapiens (Fig. 1A). A total of seven nucleotides in
the 3′UTR of the human AdipoR2 mRNA are perfectly complementary to
nucleotides 1–7 of miR-218 and this sequence is highly conserved in
mRNA orthologs of different species (Fig. 1B).
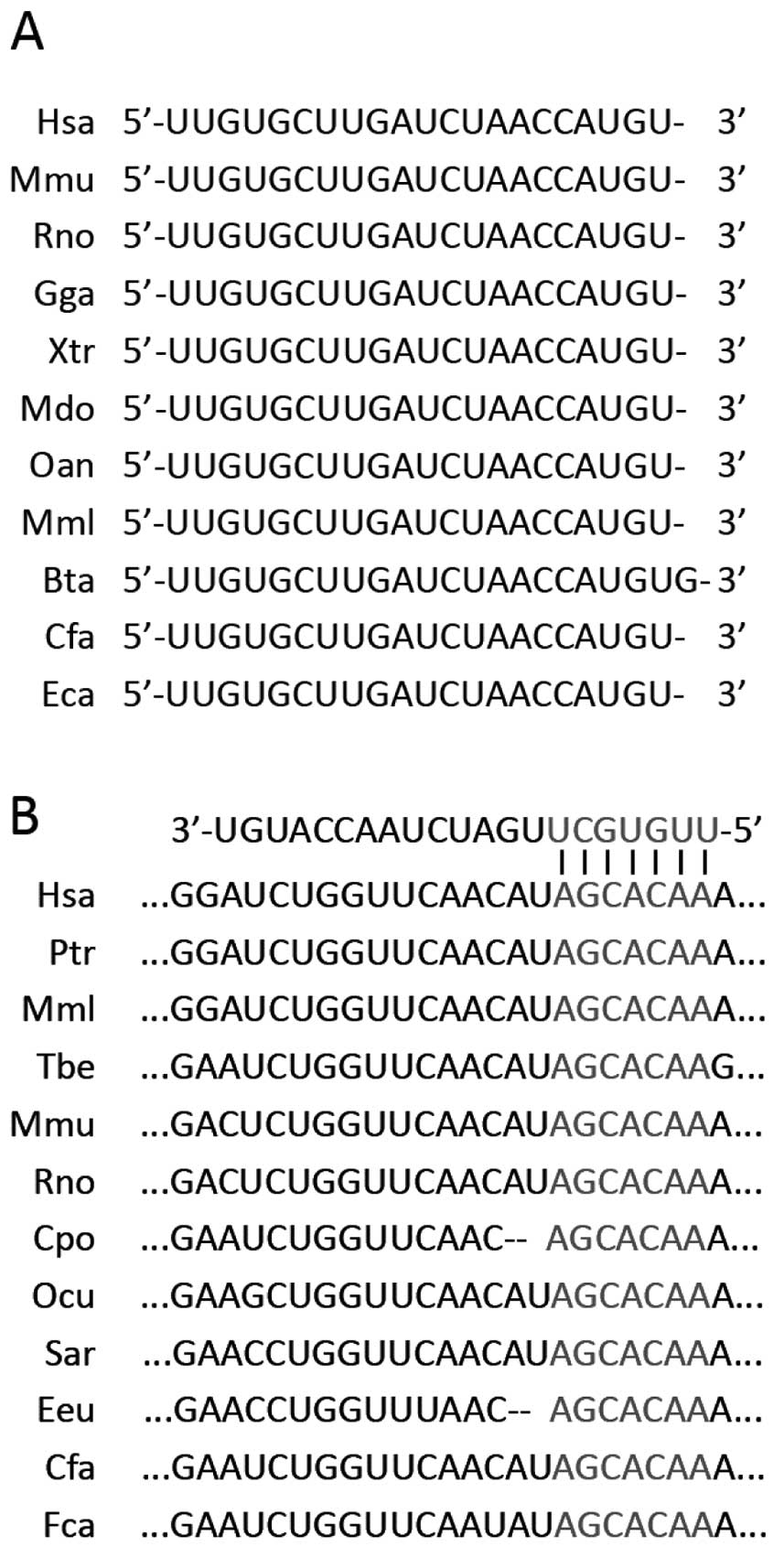 | Figure 1Conservation of miR-218 and its
predicted binding site within AdipoR1 3′ untranslated region. (A)
Evolutionary conservation of miR-218. (B) Predicted binding site of
miR-218 within the AdipoR2 3′ untranslated region of humans and
other species. miR, microRNA; AdipoR, adiponectin receptor. Hsa,
Homo sapiens; Mmu, Mus musculus; Rno, Rattus
norvegicus; Gga, Gallus gallus; Xtr, Xenopus
tropicalis; Mdo, Monodelphis domestica; Oan,
Ornithorhynchus anatinus; Mml, Macaca mulatta; Bta,
Bos taurus; Cfa, Canis familiaris; Eca, Equus
caballus; Ptr, Pan troglodytes; Tbe, Tupaia
belangeri; Cpo, Cavia porcellus; Ocu, Oryctolagus
cuniculus; Sar, Sorex araneus; Eeu, Erinaceus
europaeus; Fca, Felis catus. |
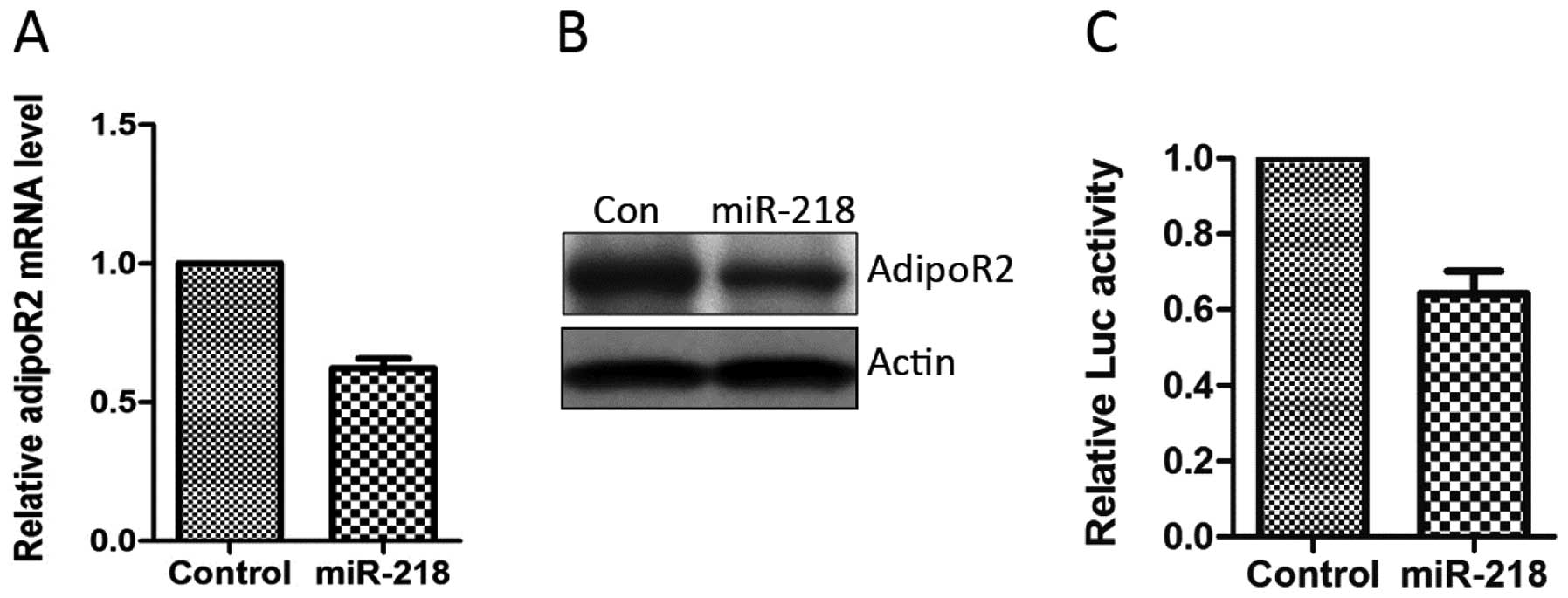
Several approaches were subsequently employed to
determine whether miR-218 regulates AdipoR2 expression.
Overexpression of miR-218 mimics reduced the AdipoR2 mRNA level to
~60% that of the control group (Fig.
2A). The AdipoR2 protein level was also significantly reduced
when cells were transfected with miR-218 mimics (Fig. 2B). Luciferase assays were then
performed to further confirm the regulatory role of miR-218 in the
expression of AdipoR2. The 3′UTR of the AdipoR2 gene was cloned
downstream of the coding sequence of luciferase. This construct was
co-transfected into HepG2 cells with miR-218 or control. The data
demonstrated that miR-218 specifically decreased the luciferase
levels (Fig. 2C). These results
suggested that miR-218 directly binds to the 3′UTR of AdipoR2 mRNA
and reduces AdipoR2 expression.
miR-218 inhibits AMPK and p38 MAPK
pathways
AdipoR2 has been revealed to be involved in the
adiponectin-activated AMP-activated protein kinase and p38
mitogen-activated protein kinase pathways (7), and further mediates the effect of
adiponectin on glucose metabolism. The inhibition of AdipoR2 by
miR-218 prompted the hypothesis that miR-218 may affect AMPK and
p38 MAPK activation. Therefore, cells were transfected with miR-218
or negative control and cells were treated with adiponectin. The
data revealed that phosphorylation of p38 MAPK and AMPK decreased
in miR-218 expressing cells (Fig.
3), suggesting that miR-218 inhibited the adiponectin-activated
AMPK and p38 MAPK pathways.
miR-218 inhibits adiponecin-induced
glucose uptake
Adiponectin enhances glucose uptake, therefore, it
was proposed that miR-218 may regulate adiponectin-induced glucose
metabolism. To assess this hypothesis, HepG2 cells transfected with
miR-218 were then incubated with adiponectin and the uptake of
2-NBDG, a metabolizable fluorescent derivative of glucose, was
measured. As expected, adiponectin treatment was observed to
markedly increase glucose uptake by ~1.67±0.15 fold more than that
of the untreated group. Adiponectin-induced glucose uptake was
reduced by 0.97±0.21 fold when miR-218 was expressed (Fig. 4), indicating that miR-218 inhibits
adiponectin-induced glucose uptake.
Discussion
As adiponectin has been observed to exhibit a
significant antidiabetic function, elucidating the adiponectin
signaling pathway is essential to harness the therapeutic potential
of this hormone. Adiponectin receptors may have important roles in
adiponectin signaling transduction. However, the mechanism
regulating the expression of adiponectin receptors remains to be
elucidated. In the present study, it was demonstrated that miR-218
directly targets AdipoR2. In addition, it was demonstrated that
miR-218 inhibits adiponectin signaling and glucose uptake by
repressing AdipoR2 expression.
The expression levels of AdipoR1 and AdipoR2 are
closely associated with insulin sensitivity. AdipoR1 expression was
decreased in the skeletal muscle of Type 2 diabetic patients
(16). The expression of AdipoR1
and AdipoR2 in skeletal muscle and the liver is significantly
increased in fasted mice and decreased in refed mice (17). Insulin deficiency induced by
streptozotocin increased the expression of AdipoR1 and AdipoR2, and
insulin replenishment reduced the expression of AdipoR1 and AdipoR2
in vivo (17). Insulin also
reduced the expression of AdipoR1 and AdipoR2 in hepatocytes or
myocytes via the phosphoinositide 3-kinase/Foxo1-dependent pathway
in vitro (17). However,
other studies have demonstrated that insulin may also decrease the
expression of AdipoR1, but not AdipoR2 in primary cultured neonatal
rat ventricular myocytes (18) and
L6 myoblasts (19). The
inconsistency in the expression of AdipoRs suggested that the
expression regulation of AdipoR1 and AdipoR2 is complicated and not
restricted to the transcriptional level. The present data indicated
that miR-218 regulates AdipoR2 expression at the
posttranscriptional level, nevertheless the physiological and
pathological significance of miR-218-repressed AdipoR2 expression
requires further investigation.
It has been observed that miR-218 is a
multifunctional microRNA. It acts as a tumor suppressor in numerous
types of cancer, including oral cancer (20), mesenchymal glioblastoma (21) and medulloblastoma (22). miR-218 inhibits the expression of
the mammalian target of rapamycin component Rictor and therefore
inhibits AKT phosphorylation and cell proliferation (20). Through targeting the receptor
tyrosine kinase signaling pathway, miR-218 inhibits the activation
of hypoxia-inducible factor and thereby inhibits glioblastoma
multiforme cell survival and angiogenesis (21). In addition to its tumor-suppressing
effects, miR-218 was also observed to be involved in cell
differentiation. Through downregulation of Wnt signaling
inhibitors, miR-218 may activate Wnt signaling to promote the
differentiation of osteoblasts (23) or osteogenic differentiation of
human adipose tissue-derived stem cells (24). In the present study, the role of
miR-218 in glucose metabolism was revealed, for the first time to
the best of our knowledge, as demonstrated by the observation that
overexpression of miR-218 inhibits the AMPK and p38 MAPK pathways
(Fig. 3) and adiponectin-induced
glucose uptake (Fig. 4). Further
studies using mouse models may assist in elucidating the function
of miR-218 in glucose metabolism and the development of
diabetes.
In conclusion, AdipoR2 has been identified as a
direct target of miR-218. Furthermore, miR-218 was revealed to
inhibit adiponectin-induced AMPK and p38 MAPK activation and
glucose uptake in HepG2 cells. These results indicated that miR-218
targets AdipoR2 to inhibit the adiponectin signaling pathway. This
data may also provide a potential target for diabetes therapy
through inhibiting miR-218.
Acknowledgments
The present study was supported by grants from the
Natural Science Foundation of Zhejiang Province (grant nos.
Y14H040003 and Y14H040004), the Medical Foundation of Zhejiang
Province (grant no. 2013KYA207), and the Shaoxing Science and
Technology Bureau Program (grant nos. 2013B70079 and 2013B70060).
Dr Tao Zhang was awarded a grant by the China Postdoctoral Science
Foundation.
Abbreviations:
|
MAPK
|
mitogen-activated protein kinase
|
|
AdipoR
|
adiponectin receptor
|
|
AMPK
|
AMP-activated protein kinase
|
|
PPAR
|
peroxisome proliferator-activated
receptor
|
References
|
1
|
Gil-Campos M, Cañete RR and Gil A:
Adiponectin, the missing link in insulin resistance and obesity.
Clin Nutr. 23:963–974. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ukkola O and Santaniemi M: Adiponectin: a
link between excess adiposity and associated comorbidities? J Mol
Med (Berl). 80:696–702. 2002. View Article : Google Scholar
|
|
3
|
Yamauchi T, Kamon J, Waki H, et al: The
fat-derived hormone adiponectin reverses insulin resistance
associated with both lipoatrophy and obesity. Nat Med. 7:941–946.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kadowaki T, Yamauchi T, Kubota N, et al:
Adiponectin and adiponectin receptors in insulin resistance,
diabetes, and the metabolic syndrome. J Clin Invest. 116:1784–1792.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maeda N, Shimomura I, Kishida K, et al:
Diet-induced insulin resistance in mice lacking adiponectin/ACRP30.
Nat Med. 8:731–737. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li S, Shin HJ, Ding EL, et al: Adiponectin
levels and risk of type 2 diabetes: a systematic review and
meta-analysis. JAMA. 302:179–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yamauchi T and Kadowaki T: Adiponectin
receptor as a key player in healthy longevity and obesity-related
diseases. Cell Metab. 17:185–196. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fernández-Hernando C, Ramírez CM, Goedeke
L and Suárez Y: MicroRNAs in metabolic disease. Arterioscler Thromb
Vasc Biol. 33:178–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Poy MN, Eliasson L, Krutzfeldt J, et al: A
pancreatic islet-specific microRNA regulates insulin secretion.
Nature. 432:226–230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Frost RJ and Olson EN: Control of glucose
homeostasis and insulin sensitivity by the Let-7 family of
microRNAs. Proc Natl Acad Sci USA. 108:21075–21080. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu H, Shyh-Chang N, Segrè AV, et al: The
Lin28/let-7 axis regulates glucose metabolism. Cell. 147:81–94.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trajkovski M, Hausser J, Soutschek J, et
al: MicroRNAs 103 and 107 regulate insulin sensitivity. Nature.
474:649–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pullen TJ, da Silva Xavier G, Kelsey G and
Rutter GA: miR-29a and miR-29b contribute to pancreatic
beta-cell-specific silencing of monocarboxylate transporter 1
(Mct1). Mol Cell Biol. 31:3182–3194. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao X, Dong H, Lin C, et al: Reduction of
AUF1-mediated follistatin mRNA decay during glucose starvation
protects cells from apoptosis. Nucleic Acids Res. 42:10720–10730.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Debard C, Laville M, Berbe V, et al:
Expression of key genes of fatty acid oxidation, including
adiponectin receptors, in skeletal muscle of Type 2 diabetic
patients. Diabetologia. 47:917–925. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsuchida A, Yamauchi T, Ito Y, et al:
Insulin/Foxo1 pathway regulates expression levels of adiponectin
receptors and adipo-nectin sensitivity. J Biol Chem.
279:30817–30822. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui XB, Wang C, Li L, et al: Insulin
decreases myocardial adipo-nectin receptor 1 expression via
PI3K/Akt and FoxO1 pathway. Cardiovasc Res. 93:69–78. 2012.
View Article : Google Scholar
|
|
19
|
Fang X, Palanivel R, Zhou X, et al:
Hyperglycemia- and hyperinsulinemia-induced alteration of
adiponectin receptor expression and adiponectin effects in L6
myoblasts. J Mol Endocrinol. 35:465–476. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Uesugi A, Kozaki K, Tsuruta T, et al: The
tumor suppressive microRNA miR-218 targets the mTOR component
Rictor and inhibits AKT phosphorylation in oral cancer. Cancer Res.
71:5765–5778. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mathew LK, Skuli N, Mucaj V, et al:
mir-218 opposes a critical RTK-HIF pathway in mesenchymal
glioblastoma. Proc Natl Acad Sci USA. 111:291–296. 2014. View Article : Google Scholar :
|
|
22
|
Venkataraman S, Birks DK, Balakrishnan I,
et al: MicroRNA 218 acts as a tumor suppressor by targeting
multiple cancer phenotype-associated genes in medulloblastoma. J
Biol Chem. 288:1918–1928. 2013. View Article : Google Scholar :
|
|
23
|
Hassan MQ, Maeda Y, Taipaleenmaki H, et
al: mir-218 directs a wnt signaling circuit to promote
differentiation of osteoblasts and osteomimicry of metastatic
cancer cells. J Biol Chem. 287:42084–42092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang WB, Zhong WJ and Wang L: A
signal-amplification circuit between miR-218 and Wnt/beta-catenin
signal promotes human adipose tissue-derived stem cells osteogenic
differentiation. Bone. 58:59–66. 2014. View Article : Google Scholar
|