Introduction
Osteoporosis, a widespread skeletal disease, is more
common with advancing age and remains a major public health problem
due to its association with low bone mineral density (BMD) and
pathological fractures (1).
Osteoporosis is defined as a BMD value of ≥2.5 standard deviations
below the average for young adult women by the World Health
Organization (2). The pathological
consequence of osteoporosis is fracture and annually, ~1,500,000
fractures are attributable to osteoporosis in the USA (3). Osteoporosis is more prevalent in the
East due to the Asian population having a lower body mass index
(BMI), shorter height, lack of physical activity and low dietary
calcium intake (4). The scientific
pathogenesis of osteoporosis and therapeutic management options for
osteoporosis have been investigated, with progress in studies of
antiresorptive or anabolic therapeutic drugs, however, the
underlying mechanisms and treatments of osteoporosis remain to be
elucidated.
An adequate supply of oxygen and nutrients is
essential for cell survival and metabolism. Hypoxia, which is
defined as the reduction or lack of oxygen in organs, tissues or
cells, has been suggested to contribute to a number of pathologies,
including rheumatoid arthritis, cancer and lung disease (5–7). For
cells, hypoxia is important in growth inhibition, oxidative stress
and apoptosis (8). The role of
hypoxia in cell proliferation, particularly in osteoblasts, remains
to be elucidated. The present study hypothesized that each tissue
and cell type may have a different ability to adapt to hypoxic
condition and have potential self-protection mechanisms. Low levels
of hypoxia may act as a signal to induce cell potential. The
activation and proliferation of osteoblasts is an important factor
for the treatment of osteoporosis. The present study aimed to
investigate the role of hypoxia on cell proliferation in MC3T3-E1
cells and examine the underlying molecular mechanism.
Materials and methods
Reagents
α-minimal essential medium (α-MEM),
penicillin-streptomycin, Triton X-100 and dimethyl sulfoxide
(DMSO), Hoechst 33342 and EDTA were purchased from Sigma-Aldrich
(St. Louis, MO, USA). Antibodies against extracellular
signal-regulated kinase [ERK; mouse monoclonal immunoglobulin
(Ig)G2a; no sc-514302], phosphorylated (p)-ERK (mouse monoclonal
IgG1; no. sc-377400), Akt (mouse monoclonal IgG1; sc-377457), p-Akt
(rabbit polyclonal IgG; sc-135650), proliferating cell nuclear
antigen (PCNA; mouse monoclonal IgG2a; sc-25280) and anti-β-actin
(mouse monoclonal antibody C4; sc-47778) were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA) and antibodies against
hypoxia-inducible factor (HIF)-1α (rabbit monoclonal IgG; 1:400
dilution; no. 14179) and cyclin D1 (rabbit monoclonal antibody; no.
2978) were purchased from Cell Signaling Technology, Inc. (Danvers,
MA, USA). All the reagents were trace element analysis grade and
water was glass distilled.
Cell culture and hypoxia culture
conditions
Osteoblastic MC3T3-E1 cells (cat. no. CRL-2593;
American Type Culture Collection, Manassas, VA, USA) were cultured
in α-MEM supplemented with 10% fetal bovine serum (FBS; Gibco-BRL,
Invitrogen Life Technologies, Carlsbad, CA, USA) and 1%
penicillin-streptomycin at 37°C in a 5% CO2 incubator.
The cells were subcultured every 3 days with 0.2% trypsin and 0.02%
EDTA. For experiments, the cells were cultured for 24 h to obtain
monolayers in 3 ml α-MEM with 10% FBS. The cells were incubated
under various O2 concentrations, 20% (normoxia), 10, 5
and 0% (hypoxia), in small individual culture chambers [BPN-240CRH
(UV); Shanghai Yiheng Scientific Instrument Co., Shanghai, China].
The chambers were filled with a non-standard gas mixture,
containing the indicated O2 level and 5% CO2
in a N2 base.
Assessment of cell viability
Cell viability was measured using an MTT assay in
96-well plates. Briefly, the cells were seeded at 5,000 cells/well
(100 µl) and cultured with 20, 10, 5 or 0% O2 for
0–72 h. Subsequently, 10 µl MTT solution (5 mg/ml) was added
to the medium and incubated at 37°C for 3 h. The MTT solution was
removed and 100 µl DMSO was added to dissolve the colored
formazan crystals for 15 min. The absorbance was measured at 490 nm
using a Sunrise RC microplate reader (Tecan Group, Ltd., Männedorf,
Switzerland). The relative cellular growth was determined using a
ratio of the average absorbance in the treated cells, vs. the
average absorbance in the control cells.
Detection of the cell cycle
The MC3T3-E1 cells were cultured with 20, 10 or 0%
O2 for 48 h and the cell cycle phases were analyzed
using fluorescent activated cell sorting (FACS). The cells were
collected and fixed in 70% pre-cooled ethanol (4°C) overnight. The
cells were then washed and centrifuged in phosphate-buffered saline
(PBS) at 150 ×g for 5 min prior to resuspending in 0.5 ml 1% RNase
(Beyotime Institute of Biotechnology, Haimen, China) at 37°C for 30
min and staining with propidium iodide (50 µg/ml;
Sigma-Aldrich). Cell cycle analysis was performed to determine the
G1 phase and the cell cycle distribution. The DNA content and the
cell cycle were analyzed using a flow cytometer (FACSCalibur; BD
Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
The total cell extracts and nuclear extracts were
prepared, as described previously (9). The cellular proteins (50 µg)
were electrophoresed and blotted onto a polyvinylidene difluoride
membrane. The membrane was blocked with 5% nonfat milk in
Tris-buffered saline containing 25 mM Tris and 0.15 M NaCl (pH 7.4)
containing 0.1% Tween-20 (TBS-T) for 1 h, prior to incubating the
membrane with specific primary antibodies for 1 h. The following
primary antibodies were used: Anti-ERK (1:400), anti-p-ERK (1:400),
anti-Akt (1:400), anti-p-Akt (1:400), anti-PCNA (1:400),
anti-β-actin, anti-HIF-1α (1:400) and anti-cyclin D1 (1:400). The
membrane was washed three times with TBS-T for 10 min and was then
incubated with the secondary antibodies [horseradish peroxidase
(HRP)-conjugated goat anti-mouse IgG; sc-2005; or HRP-conjugated
goat anti-rabbit IgG; sc-2004; both from Santa Cruz Biotechnology,
Inc.]. Following another three washes with TBS-T for 10 min each,
the membranes were incubated with enhanced chemiluminescence
reagent for horseradish peroxidase (Super-Signal West Femto Maximum
Sensitivity Substrate; Pierce Biotechnology, Inc., Rockford, IL,
USA) for 30 sec and exposed to an autoradiography film for
visualization of the bands using the LAS-3000 luminescent image
analyzer (Fuji Photo Film Co., Ltd., Tokyo, Japan). The relative
quantities of the various proteins were analyzed and the results
were quantified using Quantity One software V4.62 (Bio-Rad,
Hercules, CA, USA).
Fluorescence microscopy
The MC3T3-E1 cells were seeded into six-well plates
at 100 000 cells/well (2,000 µl) and cultured with 20, 10 or
0% O2 for 48 h. The cells were then washed once with
ice-cold PBS (4°C) and fixed using 4% para-formaldehyde for 30 min.
The cells were washed with PBS three times and incubated with 1%
Triton X-100 for 10 min. The cells were blocked at non-specific
antibody binding sites by incubating with 10% goat serum in PBS
containing 0.3% Triton X-100 and 0.5% bovine serum albumin for 30
min at room temperature. The cells were then incubated with mouse
monoclonal antibody against HIF-1α (1:400 in PBS) overnight.
Tetramethylrhodamine-conjugated goat anti-mouse immunoglobulin G
(1:100 in PBS) was added for 30 min at room temperature and
Hoechst33342 was then added to the cells for 15 min. Subsequently,
the cells were washed three times with PBS and were visualized by
fluorescence microscopy (TCS SP5 II; Leica Microsystems, Wetzlar,
Germany).
HIF-1α overexpression plasmid
transfection and small interfering RNA (siRNA)
To induce the overexpression of HIF-1α, a plasmid
(HIF-1α-pcDNA3.1/myc-hisA) expressing a full-length HIF-1α cDNA
gene was transiently transfected using Lipofectamine 2000
(Invitrogen Life Technologies). siRNA targeting HIF-1α was prepared
by Invitrogen Life Technologies. The sequence of the siRNA,
targeting a specific sequence in ZnT7 mRNA, was
5′-AGTTCACCTGAGCCTAATA-3′ and the scrambled siRNA sequence was
5′-UUGGCUUCGUGACUAUGGCTT-3′. The target sequences of the HIF-1α
siRNA were identified using BLAST, searched against the GenBank
database (http://www.ncbi.nlm.nih.gov/genbank/; ID SI02664053).
All transfection experiments were performed in independent
triplicates according to the manufacturer's instructions for
Lipofectamine 2000. The cells were allowed to recover in α-MEM for
24 h following transfection.
RNA extraction and reverse transcription
quantitative polymerase chain reaction (RT-qPCR)
The total RNA was isolated from the cells using
TRIzol reagent (Invitrogen Life Technologies) according to the
manufacturer's instructions. A Protein-Nucleic Acid Analyzer
(GeneQuant Pro DNA/RNA; GE Healthcare, Little Chalfont, UK) was
used to determine the concentration and quality of total RNA
extracted. cDNA synthesis and PCR detection were performed using a
PCR instrument (PTC-200; Bio-Rad) according to the manufacturer's
instructions of the PrimeScript™ RT-PCR kit (Takara Bio, Inc.,
Otsu, Japan). The sequences of the HIF-1α and GAPDH genes were
obtained from the GenBank database and specific primers were
designed over an exon-exon junction using Primer Premier 5.0
software (Premier Biosoft, Palo Alto, CA, USA). The following
primers were used: GAPDH forward, 5′-AGCAGTCCCGTACACTGGCAAAC-3′ and
reverse, 5′-AGCAGTCCCGTACACTGGCAAAC-3′; HIF-1α forward,
5′-GACAATAGCTTCGCAGAATGC-3′ and reverse,
5′-TCGTAACTGGTCAGCTGTGG-3′. RT-qPCR was performed using
gene-specific oligonucleotides under conditions of linear
amplification. The reaction conditions were as follows: 94°C for 4
min; 94°C for 40 sec, 50°C for 45 sec and 72°C for 45 sec, for 35
cycles; and followed by a final extension at 72°C for 10 min.
Electrophoresis (2% agarose gel; 110 V for 30 min) was performed to
separate the PCR products. The RT-qPCR products were stained with
ethidium bromide and visualized with ultraviolet light. A gel
imaging system (FluorChem FC2; Alpha Innotech, Palo Alto, CA, USA)
was used to capture images.
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. Statistical comparisons were made by analysis of variance
and individual comparisons were made using Tukey's multiple
comparison test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Low levels of hypoxia promotes the
proliferation of MC3T3-E1 cells
To investigate the role of hypoxia on cell
proliferation, the MC3T3-E1 cells were cultured with various
concentrations of O2 for 1–72 h and the cell viability
was detected using an MTT assay. As shown in Fig. 1A, a low level of hypoxia (10 or 5%
O2) promoted cell proliferation in a time-dependent
manner, while severe hypoxia (1 or 0% O2) inhibited cell
proliferation or induced apoptosis in the cells. The promotion of
proliferation under 10% O2 was more marked. To determine
whether the effect on cell proliferation was associated with cell
cycle regulation, the cell cycle progression of the MC3T3-E1 cells
was investigated. Notably, the number of cells in the S phase of
the cell cycle under low levels of hypoxia (10% O2) was
significantly increased compared with the normoxic group (20%
O2) and severe hypoxia (0% O2) significantly
reduced the number of cells in the S phase. Light microscopy
revealed that low levels of hypoxia promoted cell proliferation and
severe hypoxia resulted in apoptotic morphological changes
(Fig. 1C). To assess whether low
levels of hypoxia were involved in cell proliferation, the levels
of proliferation-associated protein, PCNA, and cell cycle protein,
cyclin D1, were detected by western blot analysis under different
O2 concentrations. As shown in Fig. 1D, low levels of hypoxia upregulated
the expression levels of PCNA and cyclin D1. Conversely, the
proteins were markedly decreased in the severe hypoxia cells
compared with the control group. In conclusion, low levels of
hypoxia were not harmful to the cells and, to a certain extent, may
assist in promoting the proliferation of MC3T3-E1 cells.
 | Figure 1Hypoxia promotes MC3T3-E1 cell
proliferation. (A) MC3T3-E1 cells were cultured in 20, 10, 5 or 0%
O2 for 0–72 h and cell viability was analyzed using a
3-(4,5-dimethylthiazol-2-y)-2,5-diphenyltetrazolium bromide assay.
The data are expressed as the mean ± standard deviation of three
independent experiments. (B) The cells were cultured in 20, 10 or
0% O2 for 48 h and cell cycle phases were analyzed using
fluorescent activated cell sorting. (C) Cells were treated, as
described above, and cell morphology was observed by microscopy
(magnification, ×400). (D) Cells were cultured in 20, 10, 5 or 0%
O2 for 48 h and the expression levels of PCNA and cyclin
D1 were detected using western blot analysis. The results represent
three independent experiments. β-actin was used as a loading
control. Values are expressed as the mean ± standard error of the
mean. **P<0.01 and #P<0.01, vs. 20%
O2 group. OD, optical density; PCNA, proliferating cell
nuclear antigen. |
Hypoxia upregulates the expression of
HIF-1α in MC3T3-E1 cells
In order to investigate whether hypoxia affected the
expression of HIF-1, the MC3T3-E1 cells were cultured with 20, 10,
5 or 0% O2 for 48 h and the protein expression of HIF-1α was
determined by western blot analysis. As shown in Fig. 2A, the protein expression of HIF-1α
was significantly induced under hypoxia (10% O2) and
reduced under severe hypoxia (0% O2). Additionally, an
increase in the expression of HIF-1α at 10% O2 and a
decrease at 0% O2 were observed by fluorescent
microscopy (Fig. 2B). To determine
whether the hypoxia-dependent expression of HIF-1α occurred in a
time-dependent manner, the cells were cultured with 10%
O2 for 0, 12, 24, 36, 48, 60 and 72 h, and the
expression of HIF-1α was detected using western blot analysis. As
shown in Fig. 2C and D, the
expression of HIF-1α increased in hypoxia (10% O2),
peaked at 48 h and decreased after 48 h, suggesting an internal
mechanism inhibited the further increase in the expression of
HIF-1α. However, hypoxia was important in the upregulation of the
overall expression of HIF-1α.
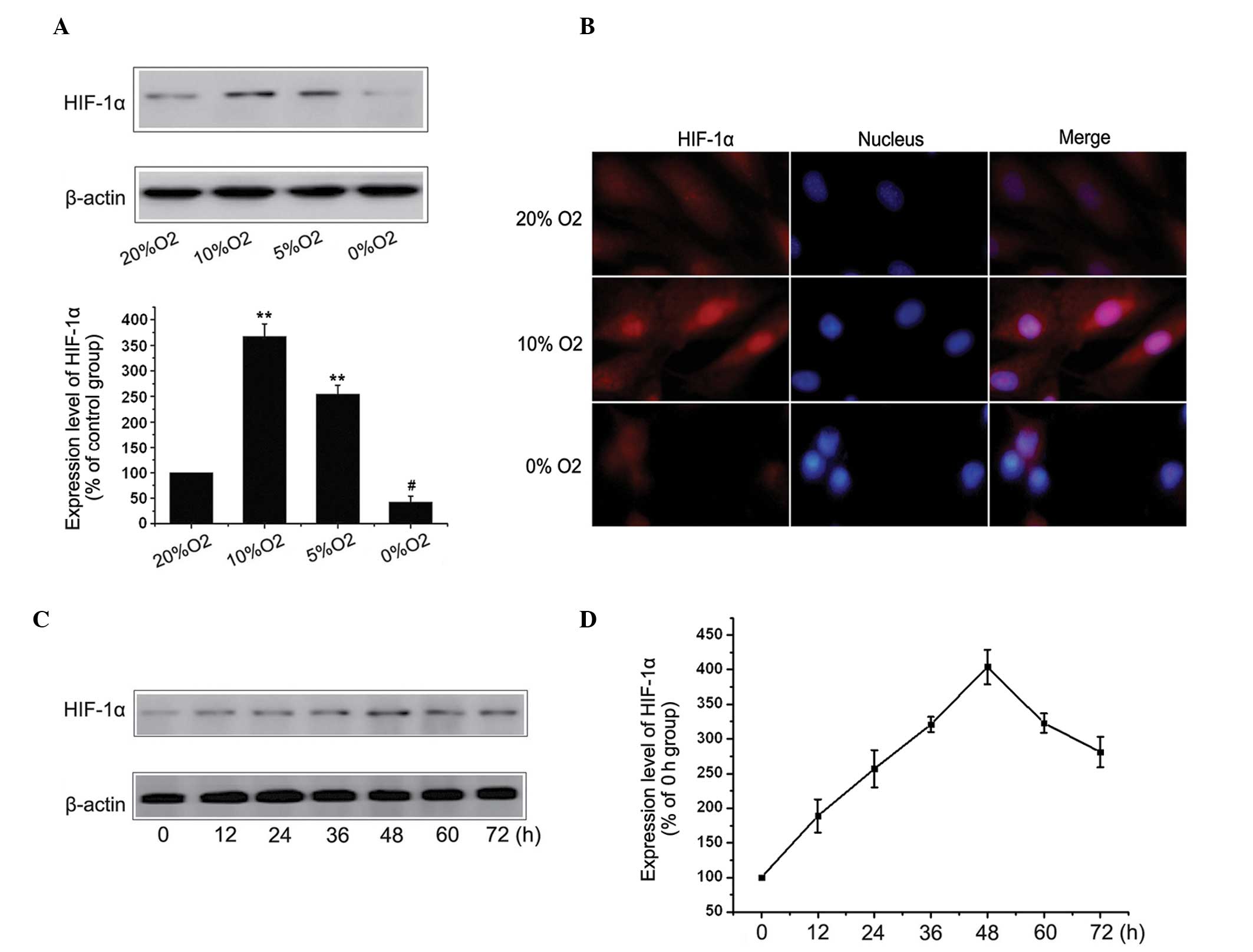 | Figure 2Hypoxia increases the expression of
HIF-1α in MC3T3-E1 cells. (A) MC3T3-E1 cells were cultured in 20,
10, 5 or 0% O2 for 48 h and the expression of HIF-1α was
detected by western blot analysis. The results represent three
independent experiments. β-actin was used as loading control.
(**P<0.01, vs. 20% O2 group). (B) Cells
were cultured in 20, 10 or 0% O2 for 48 h and the HIF-1α
protein was stained and observed by fluorescent microscopy
(magnification, ×400). (C) MC3T3-E1 cells were cultured in 10%
O2 for 0, 12, 24, 36, 48, 60 and 72 h and the expression
of HIF-1α was detected using western blot analysis. (D) Results
represent three independent experiments. β-actin was used as a
loading control. Values are expressed as the mean ± standard error
of the mean. HIF, hypoxia-inducible factor. |
Hypoxia promotes cells proliferation via
the HIF-1α pathway
Low levels of hypoxia not only increased cell
proliferation, but also upregulated the expression of HIF-1α in the
MC3T3-E1 cells. The present study investigated the effect of HIF-1α
on cell proliferation by siRNA knockdown and overexpression of
HIF-1α. The effects of HIF-1α knockdown and the overexpression of
HIF-1α were most marked 48 h after transient transfection (Fig. 3A-C). The cells in the control,
HIF-1α-overexpression and HIF-1α-knockdown groups were cultured in
20 or 10% O2 and the expression levels of the
proliferation-associated protein, PCNA, and cell cycle protein,
cyclin D1, were detected by western blot analysis. As shown in
Fig. 3C and D, the overexpression
of HIF-1α increased the effect of hypoxia-induced cell
proliferation and this activation by hypoxia was significantly
reversed by HIF-1α-knockdown, indicating that low levels of hypoxia
promoted cell proliferation via the HIF-1α pathway.
Hypoxia promotes the expression of HIF-1α
via the mitogen-activated protein kinase (MAPK)/ERK and
phosphoinositide 3-kinase (PI3K)/Akt pathways
Previous studies have demonstrated that the MAPK/ERK
and PI3K/Akt pathways are important in regulating the expression of
HIF-1α (10,11). The present study investigated
whether these signaling pathways were involved in regulating the
expression of HIF-1α in the MC3T3-E1 cells under hypoxic
conditions. The MC3T3-E1 cells were cultured under different
O2 concentrations and the expression levels of ERK,
p-ERK, Akt and p-Akt were detected by western blot analysis. The
total levels of Akt and ERK remained unchanged under different
O2 concentrations. Based on the western blot analysis
results, a low level of hypoxia significantly increased the
activation of p-Akt and p-ERK, while severe hypoxia inhibited the
PI3K/Akt and MAPK/ERK pathways. To assess whether the low level
hypoxia-induced activation of ERK or Akt specifically regulated the
expression of HIF-1α, the ERK pathway was suppressed using PD98059,
an inhibitor of the upstream ERK regulator, MEK1/2. The PI3K/Akt
pathway was inhibited using an LY294002 PI3K inhibitor. The
LY294002 (10 µM) (12)
markedly decreased the activation of p-Akt and the expression of
HIF-1α (Fig. 4C) and PD98059 (10
µM) (13), had a similar
effect on the expression of HIF-1α, indicating that the
hypoxia-induced increased expression of HIF-1α in the MC3T3-E1
cells may be mediated by the PI3K/Akt and MAPK/ERK pathways.
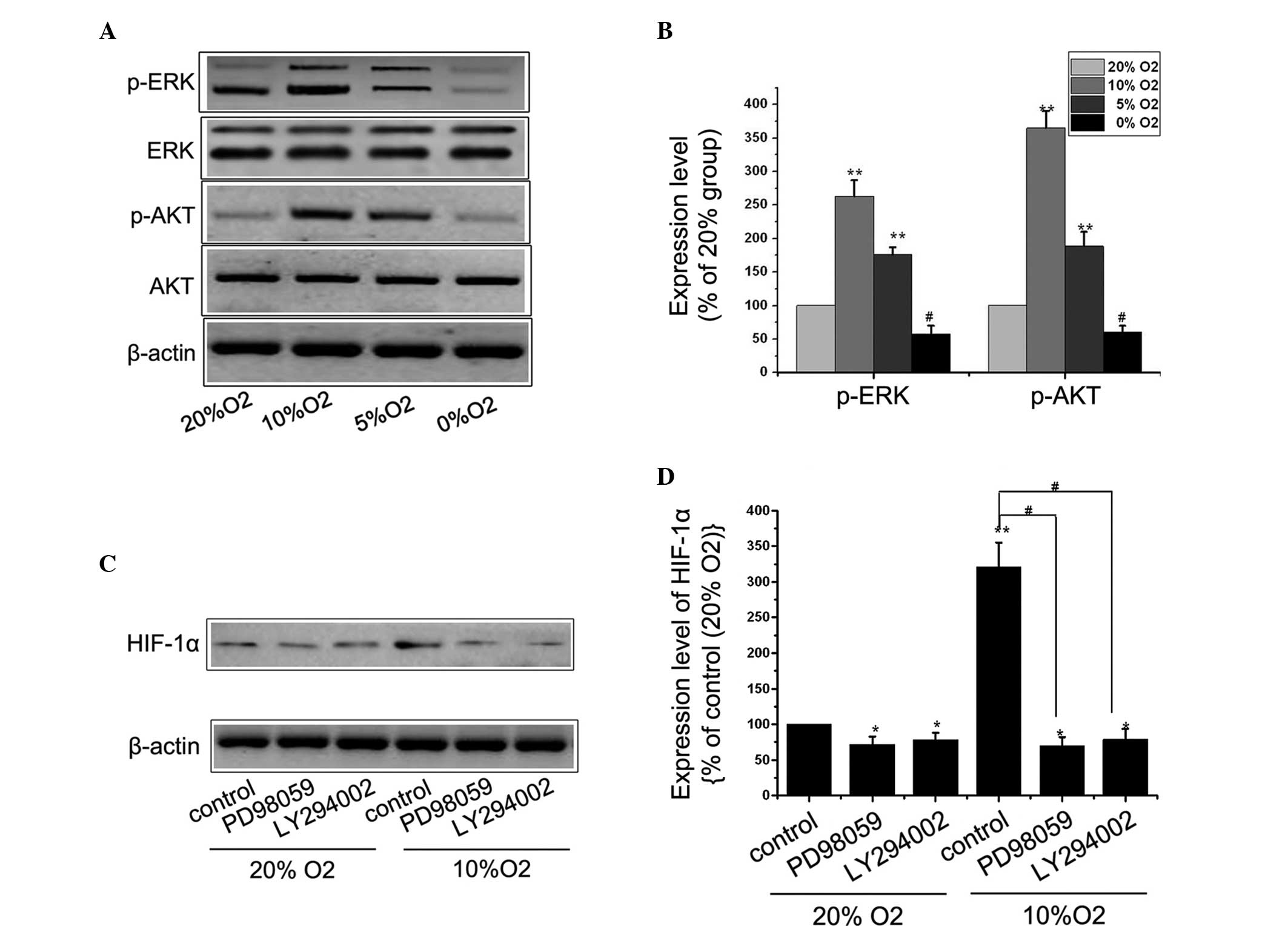 | Figure 4Mitogen activated protein kinase/ERK
and phosphoinositide 3-kinase/Akt pathways promote the expression
of HIF-1α under hypoxic conditions. (A) MC3T3-E1 cells were
cultured in 20, 10, 5 or 0% O2 for 48 h and the
expression levels of ERK, p-ERK, Akt and p-Akt were determined by
western blot analysis. (B) Results represent three independent
experiments. β-actin was used as loading control.
(**P<0.01 and #P<0.01, vs. 20%
O2 group). (C) Cells were cultured with or without 10
µM PD98059 or 10 µM LY294002 under 20 or 10%
O2, respectively, and the expression of HIF-1α was
determined by western blot analysis. (D) Results represent three
independent experiments. β-actin was used as loading control.
Values are expressed as the mean ± standard error of the mean.
*P<0.05 and **P<0.01, vs. 20%
O2 control group; #P<0.01, vs. 10%
O2 control group. ERK, extracellular signal-regulated
kinase; p-, phosphorylated; HIF, hypoxia-inducible factor. |
Discussion
Oxygen is essential for life, however, there are
multiple physiological and pathological contexts in which cells
experience conditions of hypoxia. As a negative stimulus, hypoxia
can inhibit cell growth, promote apoptosis and promote tumor cell
invasion (14). Additionally,
hypoxia also initiates its own defense system. During hypoxia,
tissues undergo a series of physiological responses to defend
themselves against a reduced oxygen supply, including increased
angiogenesis, erythropoiesis and glucose uptake (15). Whether hypoxia acts as a positive
stimulus to promote cell proliferation remains to be elucidated.
The present study revealed that low levels of hypoxia (10 or 5%
O2) promoted cell proliferation and severe hypoxia (1 or
0% O2) induced apoptotic changes in the MC3T3-E1 cells.
Low levels of hypoxia (10% O2) increased the number of
cells in the S phase of the cell cycle compared with the normoxic
group and reduced the occurrence of G1 arrest. Thus, hypoxia also
had certain positive effects, similar to the results of a previous
study, which observed hypoxia enhanced the therapeutic effect of
mesenchymal stem cells in renal isch-emia/reperfusion injury by
promoting paracrine action and improving their anti-apoptotic
ability (16,17).
The effects of hypoxia in a number of tissue and
cell types are mediated primarily by HIF-1. HIF-1 is a heterodimer
comprised of HIF-α subunits and a stable HIF-1β subunit. Whereas
HIF-1β is expressed in cells, the expression of HIF-1α is regulated
by oxygen levels (18,19). HIF-1α is stimulated by low
intracellular oxygen or genetic alteration and has been extensively
investigated as an endogenous marker of hypoxia (20). The results from the present study
demonstrated that HIF-1α was involved in the proliferation of the
MC3T3-E1 cells under hypoxic conditions. Overexpression of HIF-1α
increased the effect of hypoxia-induced cell proliferation, while
HIF-1α-knockdown reversed the hypoxic effect. In addition, the
PI3K/Akt and MAPK/ERK pathways contributed to regulating the
expression of HIF-1α under hypoxic conditions. The present study
investigated how to effectively utilize the benefits of low
O2 concentrations, overcome the hazards of high
O2 concentrations (21,22)
and incorporate the positive role of HIF-1α, to provide a novel
direction in the treatment of osteoporosis.
In conclusion, the present study demonstrated that
hypoxia activates HIF-1α through the PI3K/Akt and MAPK/ERK pathways
and enhances cell proliferation in MC3T3-E1 cells. These findings
suggested that low levels of hypoxia may have a beneficial role in
osteoblast proliferation and that combined treatment with HIF-1
inhibitors under specific concentrations of oxygen, in a well
optimized manner, may provide an important, beneficial, therapeutic
approach in osteoporosis. Hypoxia, rather than acting as an
'on-off' switch for the hypoxic response as once hypothesized,
initiates a complex cellular response involving multiple factors,
including HIF-1, and requires further investigation.
Abbreviations:
|
α-MEM
|
α-minimal essential medium
|
|
PI
|
propidium iodide
|
|
DMSO
|
dimethylsulfoxide
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
FITC
|
fluorescein isothiocyanate
|
|
PBS
|
phosphate-buffered saline
|
|
HIF
|
hypoxia-inducible factor
|
|
TBS
|
Tris-buffered saline
|
|
PDF
|
peritoneal dialysis fluid
|
|
MAPK
|
mitogen-activated protein kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
References
|
1
|
Hadji P, Klein S, Gothe H, Häussler B,
Kless T, Schmidt T, Steinle T, Verheyen F and Linder R: The
epidemiology of osteoporosis - Bone Evaluation Study (BEST): an
analysis of routine health insurance data. Dtsch Arztebl Int.
110:52–57. 2013.PubMed/NCBI
|
|
2
|
Kanis JA, McCloskey EV, Johansson H, Oden
A, Melton LJ 3rd and Khaltaev N: A reference standard for the
description of osteoporosis. Bone. 42:467–475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Riggs BL and Melton LJ 3rd: The worldwide
problem of osteoporosis: Insights afforded by epidemiology. Bone.
17:505S–511S. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soleymanian A, Niknami S, Hajizadeh E,
Shojaeizadeh D and Montazeri A: Development and validation of a
health belief model based instrument for measuring factors
influencing exercise behaviors to prevent osteoporosis in
pre-menopausal women (HOPE). BMC Musculoskelet Disord. 15:612014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Muz B, Khan MN, Kiriakidis S and Paleolog
EM: Hypoxia. The role of hypoxia and HIF-dependent signalling
events in rheumatoid arthritis. Arthritis Res Ther. 11:2012009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Scherbakov AM, Stefanova LB, Yakushina IA
and Krasilnikov MA: Beta-catenin signaling pathway and the
tolerance of breast cancer cells to hypoxic conditions. Klin Lab
Diagn. 10:68–70. 2013.
|
|
7
|
Liu SS, Wang HY, Tang JM and Zhou XM:
Hypoxia-induced collagen synthesis of human lung fibroblasts by
activating the angiotensin system. Int J Mol Sci. 14:24029–24045.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Banasiak KJ and Haddad GG: Hypoxia-induced
apoptosis: effect of hypoxic severity and role of p53 in neuronal
cell death. Brain Res. 797:295–304. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Calzado MA, de la Vega L, Möller A,
Bowtell DD and Schmitz ML: An inducible autoregulatory loop between
HIPK2 and Siah2 at the apex of the hypoxic response. Nat Cell Biol.
11:85–91. 2009. View
Article : Google Scholar
|
|
10
|
Yang XM, Wang YS, Zhang J, Li Y, Xu JF,
Zhu J, Zhao W, Chu DK and Wiedemann P: Role of PI3K/Akt and MEK/ERK
in mediating hypoxia-induced expression of HIF-1alpha and VEGF in
laser-induced rat choroidal neovascularization. Invest Ophthalmol
Vis Sci. 50:1873–1879. 2009. View Article : Google Scholar
|
|
11
|
Moon EJ, Sonveaux P, Porporato PE, Danhier
P, Gallez B, Batinic-Haberle I, Nien YC, Schroeder T and Dewhirst
MW: NADPH oxidase-mediated reactive oxygen species production
activates hypoxia-inducible factor-1 (HIF-1) via the ERK pathway
after hyperthermia treatment. Proc Natl Acad Sci USA.
107:20477–20482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fujita T, Azuma Y, Fukuyama R, Hattori Y,
Yoshida C, Koida M, Ogita K and Komori T: Runx2 induces osteoblast
and chondrocyte differentiation and enhances their migration by
coupling with PI3K-Akt signaling. J Cell Biol. 166:85–95. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang LY, Zhou YY, Chen F, Wang B, Li J,
Deng YW, Liu WD, Wang ZG, Li YW, Li DZ, Lv GH and Yin BL: Taurine
inhibits serum deprivation-induced osteoblast apoptosis via the
taurine transporter/ERK signaling pathway. Braz J Med Biol Res.
44:618–623. 2011.PubMed/NCBI
|
|
14
|
He X, Brenchley PE, Jayson GC, Hampson L,
Davies J and Hampson IN: Hypoxia increases heparanase-dependent
tumor cell invasion, which can be inhibited by antiheparanase
antibodies. Cancer Res. 64:3928–3933. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shao Y and Zhao FQ: Emerging evidence of
the physiological role of hypoxia in mammary development and
lactation. J Anim Sci Biotechnol. 5:92014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ding X, Lu C, Liu H, Rao S, Cai J, Liu S,
Kriegel AJ, Greene AS, Liang M and Ding X: Hypoxic preconditioning
with cobalt of bone marrow mesenchymal stem cells improves cell
migration and enhances therapy for treatment of ischemic acute
kidney injury. PLoS One. 8:e627032013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu S, Yan G, Xu H, He W, Liu Z and Ma G:
Hypoxic preconditioning increases survival of cardiac progenitor
cells via the Pim-1 kinase-mediated anti-apoptotic effect. Circ J.
78:724–731. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Semenza GL: Hypoxia-inducible factors in
physiology and medicine. Cell. 148:399–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Ma Z, Zhao C, Wang Y, Wu G, Xiao J,
McClain CJ, Li X and Feng W: HIF-1α and HIF-2α are critically
involved in hypoxia-induced lipid accumulation in hepatocytes
through reducing PGC-1α-mediated fatty acid β-oxidation. Toxicol
Lett. 226:117–123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tsai YP and Wu KJ: Hypoxia-regulated
target genes implicated in tumor metastasis. J Biomed Sci.
19:1022012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Al-Motabagani MA: Histological changes in
the alveolar structure of the rat lung after exposure to hyperoxia.
Ital J Anat Embryol. 110:209–223. 2005.
|
|
22
|
Wang X, Wang Y, Kim HP, Nakahira K, Ryter
SW and Choi AM: Carbon monoxide protects against hyperoxia-induced
endothelial cell apoptosis by inhibiting reactive oxygen species
formation. J Biol Chem. 282:1718–1726. 2007. View Article : Google Scholar
|















