Introduction
Gastric cancer is one of the most common types of
malignant tumor in China (1). At
present, the rate of gastric cancer-associated mortality is
increasing, accounting for almost a quarter of the total cases of
cancer-associated mortality worldwide (2). In addition, the number of patients
diagnosed with gastric cancer has increased (2,3).
Thus, it is clear that gastric cancer presents a serious threat to
public health. Gastric cancer can occur at any age, with the
majority of patients diagnosed between the ages of 40 and 60 years
(3). At present, no single cause
has been identified; however, associated factors have been reported
to include diet, lifestyle, environmental factors, genetic
predisposition and Helicobacter pylori infection (4). At present, comprehensive therapy,
including chemotherapy, is the main method for the treatment method
used for treating advanced gastric cancer (5). Chemotherapeutic drugs do not only
cause tumor cell death, they also damage normal tissue cells,
therefore, the overall survival rate of patients with gastric
cancer is not significantly improved by chemotherapy alone
(6). Thus, in order to maximize
the effect of eradicating tumor cells in malignant tumor tissues,
greater importance has been placed upon the identification of
pharmaceutical drugs exhibiting the lowest toxicity towards normal
cells (7).
The nuclear factor-κB (NF-κB) transcriptional factor
family consists of five subunits: Rel (cRel), p65 (RelA; NF-κB3),
RelB and p50 (NF-κB1) and p52 (NF-κB2) (8). The two most common dimers of NF-κB
include p65 and p50. In resting cells, IκB, the inhibitory unit of
NF-κB, combines with NF-κB, resulting in inactivation of the
cytoplasmic form. When the cells are stimulated by extracellular
signals, the IκB kinase complex (IκB kinase; IKK) phosphorylates
IκB, and the nuclear localization sites of NF-κB are exposed.
Subsequently, the free NF-κB rapidly translocates into the nucleus
and combines with specific κB sequences that induce gene
transcription (9). Previous
histological studies have indicated the importance of local
inflammation in nasopharyngeal carcinoma tumorigenesis (10). As a key inflammatory signaling
pathway, NF-κB has been demonstrated to be constitutively active in
tumors by immunohistochemical staining (10). The constitutive activation of NF-κB
commonly results in malignant carcinoma cell proliferation in
various cancer cells and tissues, due to the fact that the NF-κB
signaling pathway regulates a series of target genes involved in
cellular proliferation, apoptosis, immune response and
transcription (11,12).
MicroRNAs (miRNAs) are small, non-coding RNAs of
20–25 nucleotides in length. miRNAs negatively regulate gene
expression via incomplete complementarity to the 3′-untranslated
regions (UTR) of the target genes (13). In previous years, the aberrant
expression of miRNAs has been increasingly linked with various
types of human cancer (14). In
addition, as important mediators, miRNAs are known to be key
modulators or effectors of the NF-κB signaling pathway (15). For example, miR-146a and miR-146b
negatively interact with interleukin-1 receptor-associated kinase 1
and tumor necrosis factor (TNF) receptor-associated factor 6
protein levels, resulting in the activation of NF-κB (16). In addition, miR-199a has been
demonstrated to suppress IKKβ, which reduces the activity of NF-κB
signaling (17).
The present study aimed to investigate the relative
expression levels of miR-19a in human gastric carcinoma. The
effects of increased miR-19a levels on gastric carcinoma cell
proliferation and migration were also examined. These
investigations aimed to determine whether miR-19a affected gastric
cell proliferation and migration through the NF-κB signaling
pathway, thereby preventing gastric cancer progression.
Materials and methods
Cell culture and treatments
The GES-1, MGC-803, BGC-823 and SGC-7901 human
gastric carcinoma cell lines as well as HEK293T cells were
purchased from American Type Culture Collection (Manassas, VA, USA)
and were cultured in Dulbecco's modified Eagle's medium (DMEM)/F12
(GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS; GE Healthcare Life Sciences), 100 U/ml
penicillin and streptomycin (Invitrogen Life Technologies,
Carlsbad, CA, USA) in a 25 cm2 culture flask at 37°C in
a humidified atmosphere of 5% CO2. Cells were treated
with 10 ng/μl TNF-α for 48 h at 60% confluence.
Transient transfections
Prior to transfection, 1.5×105 cells/well
were seeded into a 6-well plate in 2 ml DMEM culture medium
containing FBS and antibiotics. Prior to transfection, the cells
were incubated under normal growth conditions (37°C and 5%
CO2). Subsequently, the cells were transfected with
miR-19a mimics, miR-19a inhibitors or miR negative controls for 48
h (Shanghai GenePharma Co., Ltd., Shanghai, China), which were
pre-incubated with HiPerFect transfection reagent (Qiagen, Hilden,
Germany), with a final concentration of miRNA analogues at 100
nmol/l.
RNA extraction
Total RNA was extracted from the cell lines (5 mg)
using TRIzol reagent (Invitrogen Life Technologies), according to
the manufacturer's instructions.
Bioinformatic analysis
TargetScan (http://www.targetscan.org/) was used to predict the
target gene of miR-19a.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
In order to detect and quantify mature microRNA-19a,
a TaqMan MicroRNA Reverse Transcription kit and TaqMan MicroRNA
assay were used, according to the manufacturer's instructions
(Applied Biosystems, Life Technologies, Foster City, CA, USA). U6
RNA was used for normalization. To quantify the miRNA levels, 10 ng
total RNA was reverse-transcribed using the Taq-Man MicroRNA
Reverse Transcription kit, using specific primers for miR-19a and
U6. Nucleotide primers used for reverse transcription were as
follows (5′-3′): miR-19a,
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGAGCA; U6,
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAATATG. The primers
used for real-time PCR were as follows (5′-3′): miR-291b-3p
forward, GGCAAACAGCAAAAC; U6 forward, GCGCGTCGTGAAGCGTTC; Universal
reverse primer, GTGCAGGGTCCGAGGT. Subsequently, PCR amplifications
were performed in 20 μl reaction volumes, containing 10
μg TaqMan 2X Universal PCR Master Mix, 1 μl 20X
TaqMan MicroRNA Assay mix (Applied Biosystems, Life Technologies)
and 1.33 μl template cDNA in the same system used for mRNA
quantitation (ABI 2720; Applied Biosystems). The thermal cycling
conditions were as follows: 95°C for 10 min, followed by 40 cycles
at 95°C for 15 sec and at 60°C for 1 min. The relative miRNA
expression of miR-19a was normalized against the U6 RNA endogenous
control using the 2ΔΔCT method. Bio-Rad CFX Manager
software (v 1.6; Bio-Rad Laboratories, Inc., Hercules, CA, USA) was
used for the quantitative analysis of mRNA and miRNA.
Protein extraction, western blotting and
antibody incubation
The cellular proteins were extracted from the cells
using radioimmunoprecipitation buffer, containing 50 mM Tris/HCl
(pH 7.4), 150 mM NaCl and 1% (v/v) NP-40, and 0.1% (w/v) SDS
(Beijing SolarBio Science & Technology Co., Ltd., Beijing,
China), containing 1% (v/v) phenylmethanesulfonylfluoride (Beijing
SolarBio Science & Technology Co., Ltd.), 0.3% (v/v) protease
inhibitor (Sigma-Aldrich, St. Louis, MO, USA) and 0.1% (v/v)
phosphorylated proteinase inhibitor (Sigma-Aldrich). The lysates
were centrifuged at 13,000 ×g at 4°C for 15 min and the supernatant
was collected for total protein analysis. A bicinchoninic protein
assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA) was used
to determine the protein concentration. Equal quantities of protein
(15 μg) were separated on an SDS-PAGE gel (10% (v/v)
polyacrylamide) and transferred onto a polyvinylidene difluoride
membrane (EMD Millipore, Billerica, MA, USA). Nonspecific binding
was blocked using 8% (w/v) milk in Tris-buffered saline with 1%
Tween-20 (TBST; Beijing SolarBio Science & Technology Co.,
Ltd.) for 2 h at room temperature. The membranes were then
incubated with primary antibodies against β-actin (13E5) rabbit
monoclonal antibody (mAb) (cat no. 4970; Cell Signaling Technology
Inc., Beverly, MA, USA), NF-κB p65 (L8F6) mouse mAb (cat no. 6956;
Cell Signaling Technology), NF-κB p65 (D14E12) XP®
rabbit mAb #9609 (Cell Signaling Technology Inc.); VCAM-1 (E1E8X)
rabbit mAb #13662 (Cell Signaling Technology Inc.) ICAM-1 rabbit
mAb #4915 (Cell Signaling Technology Inc.); MCP-1 rabbit mAb #2027
(Cell Signaling Technology Inc.) overnight at 4°C. Following
several washes with TBST, the membranes were incubated in
horseradish peroxidase (HRP)-conjugated goat anti-rabbit and
anti-mouse immunoglobulin (Ig)G or HRP-conjugated mouse anti-goat
IgG (all at a 1:5,000 dilution) for 2 h at room temperature and
were then washed (five times with TBST for 10 min each). The target
proteins were visualized using enhanced chemiluminescence (EMD
Millipore), according to the manufacturer's instructions, and were
quantified using density analysis normalized against β-actin,
according to the manufacturer's instructions, with values expressed
as the fold-changes, compared with the control.
Inhibition of NF-κB by RNA
interference
NF-κB-specific small interfering (si)RNA (siHuR) and
negative control were purchased from Shanghai Genepharma.
1×105 cells per well in a six-well plate were
transfected with 50 nM siHuR or negative control for 48 h using
HiperFect transfection reagent (Qiagen) as described above.
Luciferase target assay
For the luciferase assay, the 3′ UTR of IκBα,
including the binding site for miR-19a, was amplified from the
MGC-803 cells using the following primers: IκBα, forward
5′-AAGGAGGAGGGCAGAATCAT-3′ and reverse, 5′-ATCTGCATGGTGATGTTGGA-3′.
The PCR product was then digested with XbaI (New England Biolabs,
Beverly, MA, USA) and cloned into the pGL3 reporter plasmid
(Promega Corporation, Madison, WI, USA), downstream of the
luciferase reporter gene.
The modified firefly luciferase vector (500
ng/μl) was transfected into HEK293 cells (2×105
cells/ml), as described previously, and firefly and Renilla
luciferase activities were measured 48 h after transfection using a
Dual-Luciferase Reporter Assay system (Promega Corporation).
Firefly activity was normalized to Renilla activity to control the
transfection efficiency.
Immunofluorescence
MGC-803 cells were cultured on six-well chamber
slides and fixed with 4% paraformaldehyde for 10 min at −20°C. The
slides were washed in PBS three times and incubated with a
polyclonal antibody against NF-κB (1:50 diluted in PBS with 1% BSA;
50 μl/slide) for 2 h at room temperature. After washing with
PBS three times (5 min per time), the slides were incubated with
tetramethylrhodamine-conjugated anti-rabbit IgG (Beijing Zhongshan
Jinqiao Biotechnology Co., Ltd, Beijing, China; diluted 1:100 in
PBS with 1% BSA; 50 μl/slide) for 1 h at room temperature.
Three times after washing the slides in PBS, the slides were
incubated with Hoechst 33258 (10 μg/ml) for 5 min. The
slides were then washed again and examined using a fluorescence
microscope (Leica CM3000; Leica Microsystems GmbH, Buffalo Grove,
IL, USA).
MTT assay
In order to evaluate the effect of miR-19a on cell
proliferation, the cells were seeded at 5,000 cells/well in 100
μl medium in 96-well plates and were transfected with
miR-19a mimics or inhibitors (50 nM) or negative control-miRNA
mimics (50 nM), as described above. Following transfection, 20
μl MTT reagent (Beijing SolarBio Science & Technology
Co., Ltd.) was added to the wells after 24 h and incubated for 4 h
at 37°C. Subsequent to removal of the medium, 200 μl
dimethyl sulfoxide was added to dissolve the formazan, and the
absorbance was measured at 550 nm using a SpectraMax® M3
(Molecular Devices Inc., Sunnyvale, CA, USA). Wells containing only
MGC-803 cells served as blank controls.
Statistical analysis
Data are presented as the mean ± standard error of
three independent experiments. ImageJ software (National Institutes
of Health, Bethesda, MD, USA) was used for density analysis.
GraphPad Prism (GraphPad, Inc., La Jolla, CA, USA) was used for
statistical analyses. Student's t-test was used to assess
differences between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-19a is upregulated in human gastric
carcinoma cells
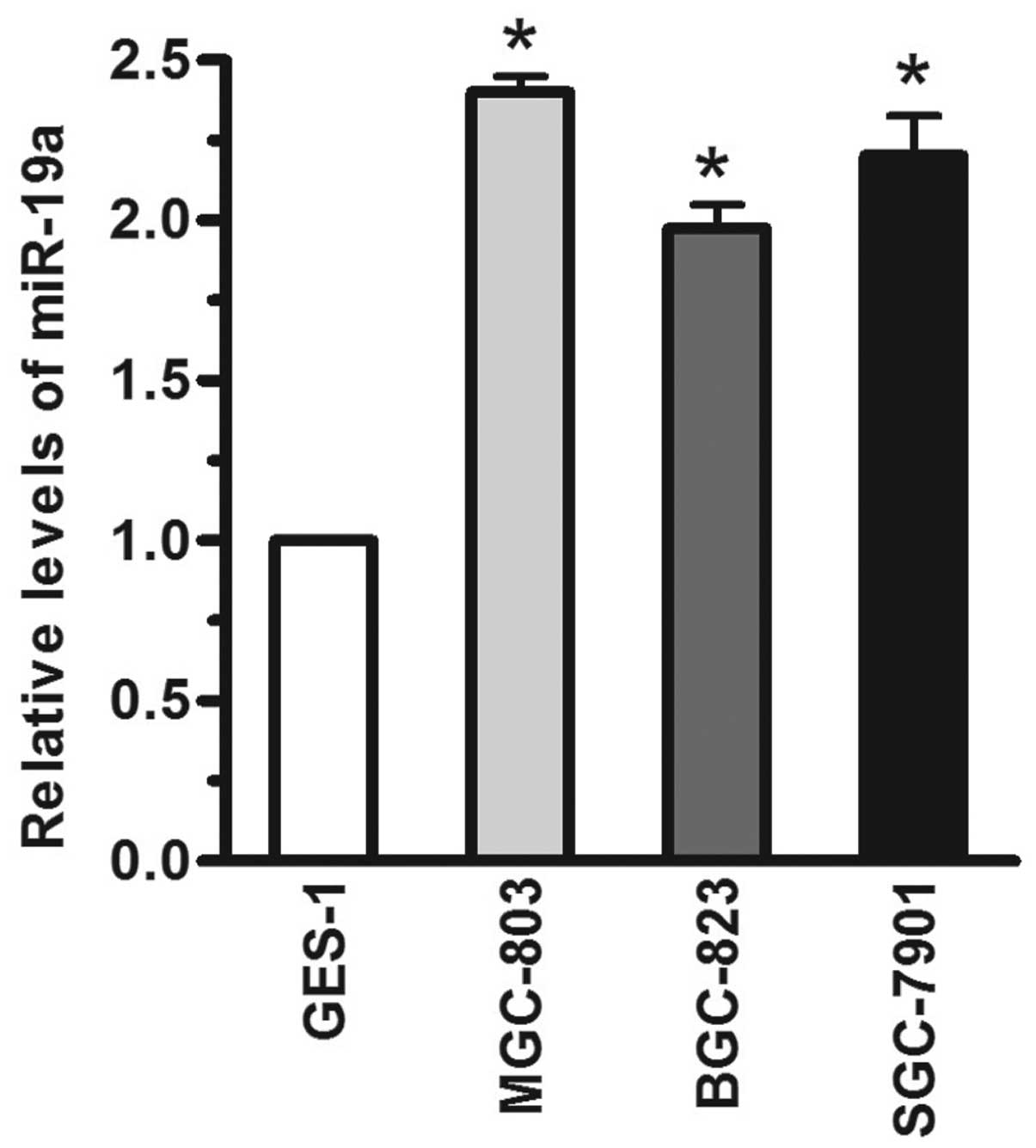
The relative levels of miR-19a in human gastric
carcinoma cells were detected using RT-qPCR. Compared with the
GES-1 immortalized gastric epithelial cell line, the miR-19a levels
were significantly increased (>1-fold) in the MGC-803, BGC-823
and SGC-7901 human gastric carcinoma cell lines, when the miR-19a
expression level was normalized to U6 (Fig. 1; P<.05). Based on these results,
the levels of miR-19a were significantly increased in human gastric
carcinoma cells.
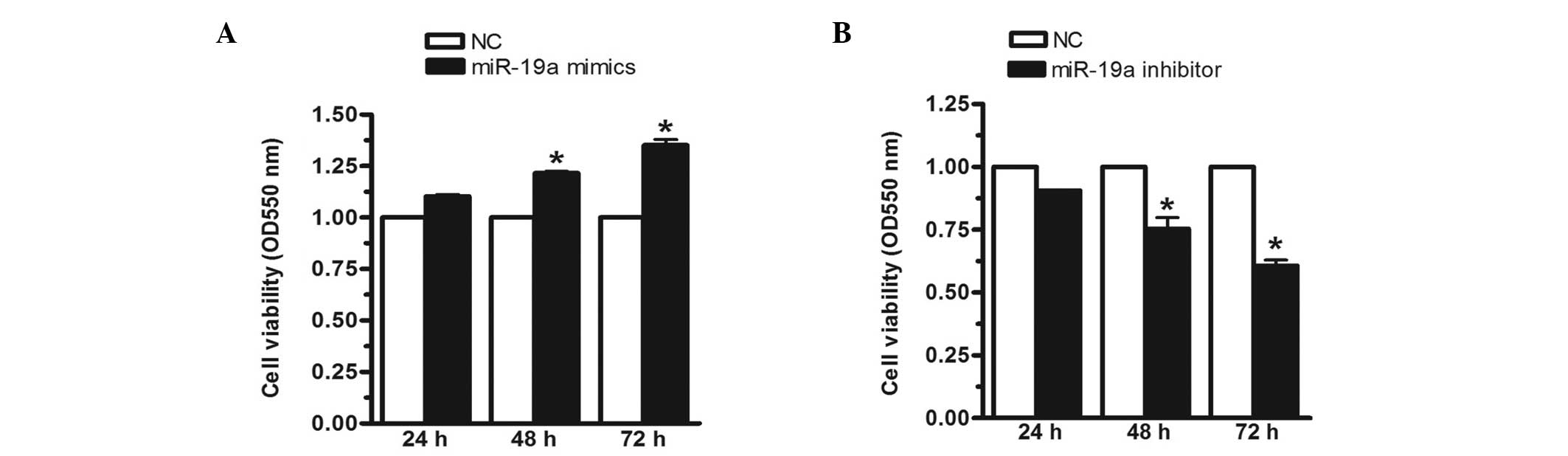
Upregulation of miR-19a increases MGC-803
cell viability
To investigate the effect of miR-19a on MGC-803 cell
viability, the MGC-803 cells were transfected with miR-19a mimics,
inhibitors or negative controls for 24, 48 or 72 h, respectively.
In this investigation, the mimics were analogues that enhanced the
expression of miR-19a, whereas the inhibitors were analogues that
reduced the expression of miR-19a. The MTT assay demonstrated that,
when the miR-19a mimics were transfected into the MGC-803 human
gastric carcinoma cell line, cell viability was significantly
increased by 23 and 35% at 48 and 72 h, respectively (Fig. 2A). Conversely, when the expression
of miR-19a was inhibited, cell viability was reduced by 17 and 36%
at 48 and 72 h, respectively (Fig.
2B). These results indicated that miR-19a increased MGC-803
cell viability.
miR-19a activates the NF-κB signaling
pathway
In gastric carcinoma, the NF-κB signaling pathway is
constitutively activated. In previous studies, several miRNAs have
been reported to activate NF-κB. For example, miR-301a has been
demonstrated to target NF-κB-repressing factor and to correlate
with abnormal NF-κB activation (17). In the present study, the MGC-803
gastric carcinoma cells were treated with 10 ng/μl TNF-α for
48 h, following which the relative levels of miR-19a were analyzed.
As shown in Fig. 3A, the relative
levels of miR-19a increased ~4-fold following TNF-α treatment. In
addition, western blotting was performed to ascertain whether TNF-α
treatment significantly activated the NF-κB signaling pathway. To
address whether miR-19a contributes to NF-κB activation, the
MGC-803 gastric carcinoma cells were transfected with miR-19a
mimics. Based on the results of the western blot analysis, it was
concluded that when miR-19a was overexpressed, NF-κB was
significantly activated. As shown in Fig. 3A, compared with the negative
control, the phosphorylated-NF-κB/NF-κB ratio was increased by
>1-fold. Vascular cell adhesion molecule (VCAM), intercellular
adhesion molecule (ICAM) and monocyte chemoattractant protein-1
(MCP-1) are among the important adhesion molecules that are
aberrantly expressed in various types of cancer (18). To validate the abnormal NF-κB
activation, the protein levels of VCAM, ICAM and MCP-1 were
measured. As shown in Fig. 3B,
their relative levels were all increased ~1-fold. Based on the
above results, it was concluded that miR-19a contributed to
TNF-α-stimulated NF-κB activation. In order to investigate the
effect of NF-κB on cellular proliferation, a small interfering
(si)RNA-targeting NF-κB was selected. As shown in Fig. 3C, following knockdown of NF-κB,
cell viability was significantly reduced, even in the cells
transfected with the miR-19a mimics. The MTT assay demonstrated
that cell viability was significantly reduced in the cells
transfected with si-NF-κB. These data suggested that miR-19a
enhanced cellular proliferation through activation of the NF-κB
signaling pathway.
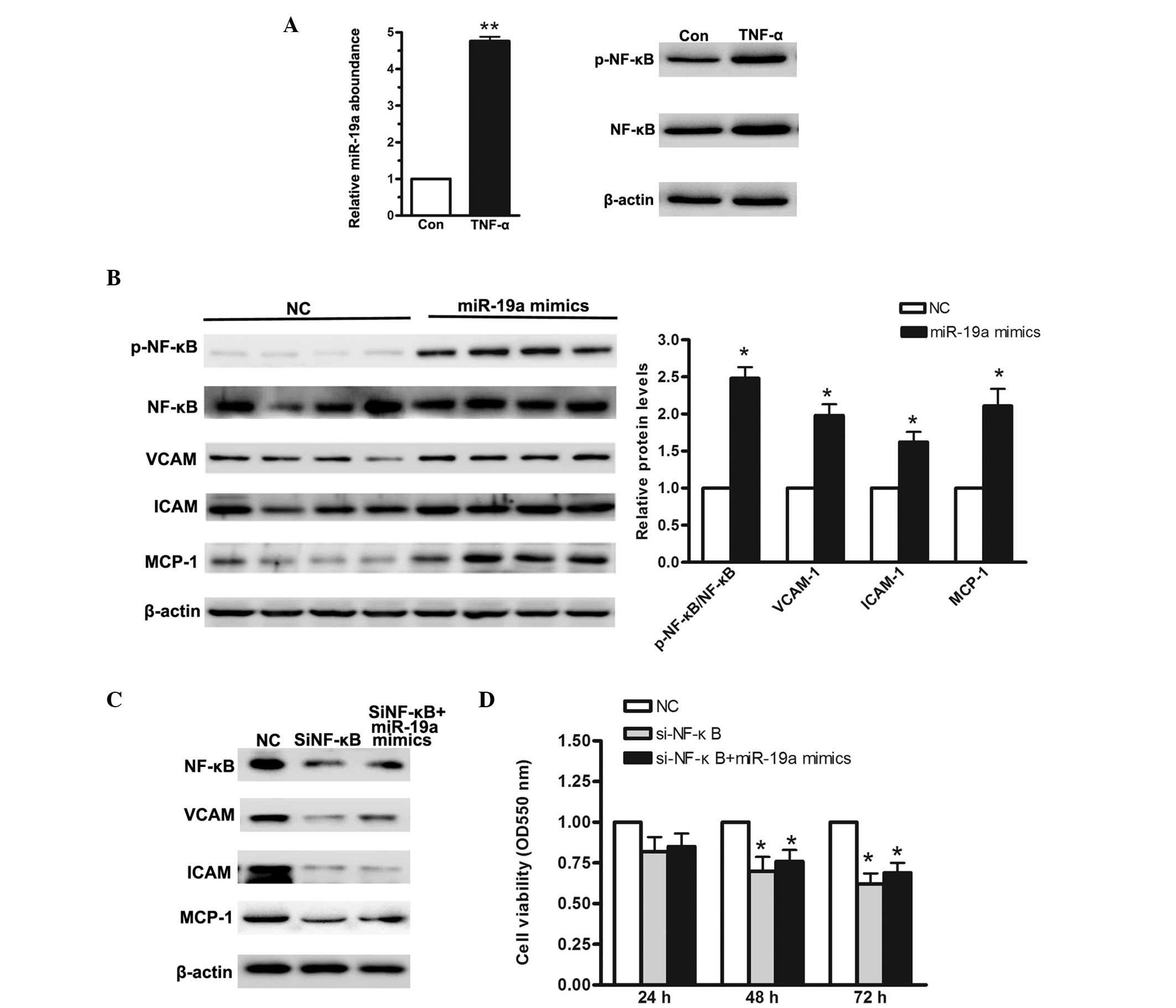 | Figure 3NF-κB signaling pathway is activated
when miR-19a is overexpressed in the MGC-803 human gastric
carcinoma cell line. (A) Reverse transcription-quantitative
polymerase chain reaction was used to determine the relative levels
of miR-19a when MGC-803 cells were treated with 10 ng/μl
TNF-α for 48 h. (B) Western blot analysis of NF-κB activation and
its downstream regulators when miR-19a was overexpressed. (C)
Western blot analysis of siRNA targeting NF-κB. (D) MTT assay
demonstrated a low cellular proliferation rate in cells
cotransfected with si-NF-κB and miR-19a mimics. Data are presented
as the mean ± standard error of three independent experiments.
*P<0.05, vs. control. TNF-α, tumor necrosis factor-α;
NF-κB, nuclear factor-κB; VCAM, vascular cell adhesion molecule;
ICAM, intercellular adhesion molecule; MCP, monocyte
chemoattractant protein; miR, microRNA; si, small interfering; p-,
phosphorylated; NC, negative control; Con, control. |
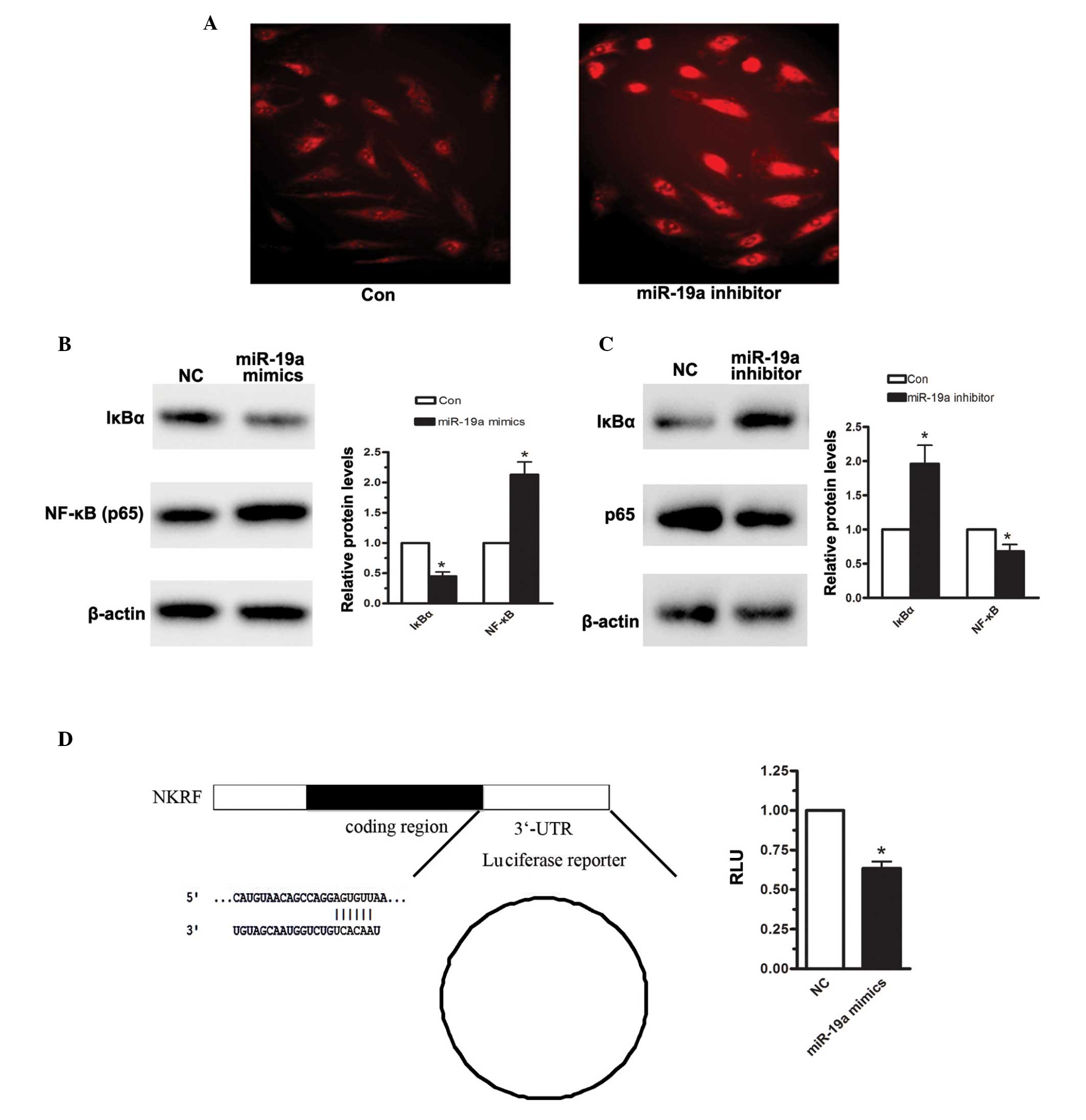
IκBα is the host gene of miR-19a
To further confirm that NF-κB was activated upon
miR-19a overexpression, immunofluorescence was measured. As shown
in Fig. 4A, enhanced NF-κB (p65)
protein levels were detected when the MGC-803 cells were
transfected with miR-19a mimics. According to TargetScan, miR-19a
was predicted to target IκBα. IκB is an NF-κB inhibitory protein
(19). In the resting state, IκB-α
combines with p65, P50, resulting in the inactivation of NF-κB in
the cytoplasm. Through the activation of IKK, IκBα is degraded,
which drives the two subunits of NF-κB to translocate between the
cytoplasm and the nucleus, particularly the p65 subunit, inducing a
downstream signaling pathway. Therefore, the effects of miR-19a on
the expression of IκBα were assessed. When the MGC-803 human
gastric carcinoma cell line was transfected with miR-19a mimics for
48 h, the density of IκBα was significantly reduced, compared with
the negative control (Fig. 4B).
Furthermore, 48 h following transfection of the MGC-803 cells with
miR-19a, the expression level of IκBα was reduced by 56%.
Conversely, when miR-19a was inhibited in the MGC-803 cells, the
expression level of IκBα was increased by almost 1-fold (Fig. 4C). A luciferase assay was also
performed to detect the effect of miR-19a on the 3′-UTR region of
IκBα. As shown in Fig. 4D, miR-19a
significantly reduced IκBα-3′-UTR-luciferase reporter activity.
Based on the above analysis, it was suggested that miR-19a induced
NF-κB activation, predominantly by targeting IκBα, in the human
gastric carcinoma cells.
Discussion
miRNAs have been widely demonstrated to regulate
various cellular processes, particularly during cancer development
and progression (20). Gastric
carcinoma is common in males and females, and substantial efforts
being made to identify their metastatic characteristics. Several
previous studies have indicated abnormal miRNA levels in gastric
carcinoma (21,22), and the present study demonstrated
that miR-19a was abnormally upregulated in MGC-803 human gastric
carcinoma cells (23). This result
suggests an oncogenic role for miR-19a in the progression of
gastric carcinoma. The MGC-803 cell is an immortal cell line,
derived from human gastric carcinoma, which has been widely applied
in the investigation of gastric cancer. In the present study, the
BGC-823 and SGC-7901 human gastric cancer cell lines were also
investigated. In the present study, the MGC-803, BGC-823 and
SGC-7901 cells were first treated with miR-19a inhibitors.
According to the MTT assay, the inhibition of cell proliferation
demonstrated that tumor growth was significantly inhibited.
To analyze the correlation between miR-19a and human
gastric carcinoma, the relative levels of miR-19a were assessed in
human gastric carcinoma cells. The results demonstrated that
miR-19a was upregulated. In addition, miR-19a was found to activate
MGC-803 cell proliferation in the MTT assay. The data obtained in
the present study indicated that TNF-α treatment induced miR-19a
overexpression by ~4-fold in the MGC-803 human gastric carcinoma
cells. Notably, NF-κB was activated when the MGC-803 cells were
transfected with miR-19a mimics for 48 h, and the downstream
regulators of the NF-κB signaling pathway, including VCAM, ICAM and
MCP-1, were also upregulated. As all these molecules are
particularly important for tumor metastasis, the activation of the
NF-κB signaling pathway was further validated (24). As IΚB-α is considered an important
target gene for NF-κB, its relative levels were assessed using
western blotting and luciferase assays. These two sets of data
indicated that miR-19a induced NF-κB activation by targeting
IκBα.
Dysregulation of the NF-κB signaling pathway is
well-characterized in cancer cell proliferation, angiogenesis,
migration and invasion (25,26),
and the present study confirmed the abnormal activation of NF-κB in
human gastric carcinoma. In order to examine the possibility that
miR-19a is involved in NF-κB activation, western blot analysis of
NF-κB and its downstream regulator protein levels was performed.
The data suggested that the proteins examined were positively
regulated by miR-19a, and that IκBα was a target gene for miR-19a.
IκBα is considered to repress NF-κB translation by binding to
specific negative regulatory elements (27). However, the biological activity of
nucleolar IκBα remains to be fully elucidated (28).
In conclusion, the present study demonstrated that
miR-19a was overexpressed in human gastric carcinoma cells and that
miR-19a enhances human gastric carcinoma MGC-803 cell
proliferation. Furthermore, miR-19a activated the NF-κB signaling
pathway by targeting IκBα, a negative regulator.
References
|
1
|
Zhang W, Ha M, Gong Y, Xu Y, Dong N and
Yuan Y: Allicin induces apoptosis in gastric cancer cells through
activation of both extrinsic and intrinsic pathways. Oncol Rep.
24:1585–1592. 2010.PubMed/NCBI
|
|
2
|
Ha MW and Yuan Y: Allicin induced cell
cycle arrest in human gastric cancer cell lines. Zhonghua Zhong Liu
Za Zhi. 26:585–589. 2004.In Chinese.
|
|
3
|
Park SY, Cho SJ, Kwon HC, Lee KR, Rhee DK
and Pyo S: Caspase-independent cell death by allicin in human
epithelial carcinoma cells: Involvement of PKA. Cancer Lett.
224:123–132. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang YW, Eom SY, Yim DH, Song YJ, Yun HY,
Park JS, Youn SJ, Kim BS, Kim YD and Kim H: Evaluation of the
relationship between dietary factors, CagA-positive Helicobacter
pylori infection and RUNX3 promoter hypermethylation in gastric
cancer tissue. World J Gastroenterol. 19:1778–1787. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee J, Kim KM, Kang WK and Ou SH:
Innovative personalized medicine in gastric cancer: time to move
forward. Clin Genet. 86:37–43. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sato T, Kikuchi Y, Saito T, Hirano S and
Kouzuma T: Results of chemotherapy using new anti-cancer drugs
since S-1 for advanced or recurrent gastric cancer in our
institute. Gan To Kagaku Ryoho. 34:1819–1825. 2007.In Japanese.
PubMed/NCBI
|
|
7
|
Monjazeb AM and Blackstock AW: The impact
of multimodality therapy of distal esophageal and gastroesophageal
junction adenocarcinomas on treatment-related toxicity and
complications. Semin Radiat Oncol. 23:60–73. 2013. View Article : Google Scholar
|
|
8
|
Burkitt MD, Williams JM, Duckworth CA,
O'Hara A, Hanedi A, Varro A, Caamaño JH and Pritchard DM: Signaling
mediated by the NF-κB sub-units NF-κB1, NF-κB2 and c-Rel
differentially regulate Helicobacter felis-induced gastric
carcinogenesis in C57BL/6 mice. Oncogene. 32:5563–5573. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shostak K and Chariot A: EGFR and NF-κB:
partners in cancer. Trends Mol Med. 21:385–393. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chung GT, Lou WP, Chow C, To KF, Choy KW,
Leung AW, Tong CY, Yuen JW, Ko CW, Yip TT, et al: Constitutive
activation of distinct NF-κB signals in EBV-associated
nasopharyngeal carcinoma. J Pathol. 231:311–322. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
LaBarbera KE, Hyldahl RD, O'Fallon KS,
Clarkson PM and Witkowski S: Pericyte NF-κB activation enhances
endothelial cell proliferation and proangiogenic cytokine secretion
in vitro. Physiol Rep. 3:e123092015. View Article : Google Scholar
|
|
12
|
Colangelo T, Fucci A, Votino C, Sabatino
L, Pancione M, Laudanna C, Binaschi M, Bigioni M, Maggi CA, Parente
D, et al: MicroRNA-130b promotes tumor development and is
associated with poor prognosis in colorectal cancer. Neoplasia.
15:1086–1099. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jeong D, Kim J, Nam J, et al: MicroRNA-124
links p53 to the NF-κB pathway in B-cell lymphomas. Leukemia.
View Article : Google Scholar
|
|
14
|
Ni F, Guo C, Sun R, et al: MicroRNA
transcriptomes of distinct human NK cell populations identify
miR-362-5p as an essential regulator of NK cell function. Sci Rep.
5:99932015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Taganov KD, Boldin MP, Chang KJ and
Baltimore D: NF-kappaB-dependent induction of microRNA miR-146, an
inhibitor targeted to signaling proteins of innate immune
responses. Proc Natl Acad Sci USA. 103:12481–12486. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dai L, Gu L and Di W: MiR-199a attenuates
endometrial stromal cell invasiveness through suppression of the
IKKβ/NF-κB pathway and reduced interleukin-8 expression. Mol Hum
Reprod. 18:136–145. 2012. View Article : Google Scholar :
|
|
17
|
Lu Z and Li Y, Takwi A, Li B, Zhang J,
Conklin DJ, Young KH, Martin R and Li Y: miR-301a as an NF-κB
activator in pancreatic cancer cells. EMBO J. 30:57–67. 2011.
View Article : Google Scholar :
|
|
18
|
Astarci E, Sade A, Cimen I, Savas B and
Banerjee S: The NF-κB target genes ICAM-1 and VCAM-1 are
differentially regulated during spontaneous differentiation of
Caco-2 cells. FEBS J. 279:2966–2986. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Benzler J, Ganjam GK, Pretz D, et al:
Central inhibition of IKKβ/NF-κB signaling attenuates high-fat
diet-induced obesity and glucose intolerance. Diabetes. 64:2015–27.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin L, Jiang H, Huang M, et al: Depletion
of histone deacetylase 1 inhibits metastatic abilities of gastric
cancer cells by regulating the miR-34a/CD44 pathway. Oncol Rep.
View Article : Google Scholar
|
|
22
|
Du Y, Wang L, Wu H, et al: MicroRNA-141
inhibits migration of gastric cancer by targeting zinc finger
E-box-binding homeobox 2. Mol Med Rep. View Article : Google Scholar
|
|
23
|
Yu M, Gou WF, Zhao S, et al: Beclin 1
expression is an independent prognostic factor for gastric
carcinomas. Tumour Biol. 34:1071–1083. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu Y, Zhu X, Liang GX, et al: Apelin-APJ
induces ICAM-1, VCAM-1 and MCP-1 expression via NF-κB/JNK signal
pathway in human umbilical vein endothelial cells. Amino Acids.
43:2125–2136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen F, Yang D, Wang S, Che X, Wang J, Li
X, Zhang Z, Chen X and Song X: Livin regulates prostate cancer cell
invasion by impacting the NF-κB signaling pathway and the
expression of FN and CXCR4. IUBMB Life. 64:274–283. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dutta S, Wang FQ, Wu HS, Mukherjee TJ and
Fishman DA: The NF-κB pathway mediates lysophosphatidic acid
(LPA)-induced VEGF signaling and cell invasion in epithelial
ovarian cancer (EOC). Gynecol Oncol. 123:129–137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nourbakhsh M, Oumard A, Schwarzer M and
Hauser H: NRF, a nuclear inhibitor of NF-κB proteins silencing
interferon-beta promoter. Eur Cytokine Netw. 11:500–501. 2000.
|
|
28
|
Chen Z, Chen LY, Dai HY, Wang P, Gao S and
Wang K: miR-301a promotes pancreatic cancer cell proliferation by
directly inhibiting Bim expression. J Cell Biochem. 113:3229–3235.
2012. View Article : Google Scholar : PubMed/NCBI
|