Introduction
Staphylococcus aureus is a significant human
pathogen causing healthcare-associated and community-acquired
infections (1). Antibiotics
effectively treat these infections, however, the emergence of
methicillin-resistant S. aureus (MRSA) currently presents a
challenge to healthcare systems worldwide (2). Globally, ~2,000,000,000 MRSA carriers
exist, of whom as many as 53,000,000 suffer from overt MRSA
infections. In addition, Staphylococcus aureus clones
resistant to the antibiotic vancomycin have been identified; and
vancomycin is the last known drug to which earlier strains had been
uniformly sensitive (3). These
organisms are termed vancomycin-intermediate-resistant
Staphylococcus aureus and vancomycin-resistant
Staphylococcus aureus (4,5).
Therefore, it is becoming difficult to treat staphylococcal
infections with current chemotherapeutic agents (6).
Honeybee (Apis mellifera L.) venom contains a
complex mixture of therapeutic compounds, including antimicrobial
peptides, allowing bees to defend their hives against predators and
external threats (7). Several
biological and pharmacological studies have examined bee venom
components for use as potential pain relievers and treatments for
inflammatory diseases (8–10). In addition, the antibacterial
activities of venom against several human and animal pathogens have
been evaluated (11). However, as
venom contains certain complex toxic components, its human
therapeutic applications have been limited. Previously, the
majority of bee venom components have been individually purified
and their specific pharmacological activities investigated.
The melittin peptide, the predominant component of
bee venom (40–48%, w/w), has been investigated substantially, and
exhibits potent cytolytic and antimicrobial activities (12). Potential actions against bacteria,
viruses and cancer cells have been extensively examined in
vitro, although the antimicrobial molecular mechanism remains
to be elucidated (13,14). However, to date, few investigations
of the in vivo antimicrobial activities of melittin have
been performed. The present study investigated the antimicrobial
activity of melittin from bee venom, and examined whether it can
inhibit MRSA infections in vitro and in vivo.
Materials and methods
Ethical statement
All animal investigations were performed in
accordance with the Guidelines for the Care and Use of Laboratory
Animals of the Ministry of Food and Drug Safety of Korea, and were
approved by the Animal Care and Use Committee of the Korea Atomic
Energy Research Institute (Jeongeup Si, Korea; IACUC protocol no.
2014–023).
Bacterial strains and reagents
The bacterial strains examined in the present study
are listed in Table I. The
streptococcal and staphylococcal strains were grown at 37°C in
Todd-Hewitt broth (BD Biosciences, Franklin Lakes, NJ, USA)
supplemented with 0.5% (w/v) yeast extract and Tryptic-soy broth
(BD Biosciences), respectively. Purified melittin was purchased
from Sigma-Aldrich (St. Louis, MO, USA). Synthetic melittin
(GIGAVLKVLTTGLPALISWIKRKRQQ) was chemically synthesised by
A&PEP Co., Inc. (DaeJeon, Korea).
 | Table IBacterial strains examined in the
present study. |
Table I
Bacterial strains examined in the
present study.
| Bacterial
strain | Description | Source |
|---|
| Streptococcus
agalactiae CNCTC 10/84 | Clinical isolate,
serotype V | (18) |
| Streptococcus
gordonii M99 | Endocarditis
clinical isolate | (21) |
| Streptococcus
pneumonia TIGR4 | Laboratory strain,
serotype IV | (22) |
| Streptococcus
epidermidis RP62a | Clinical
isolate | Present study |
| Streptococcus
bovis NEM760 | Clinical isolate,
biotype II | Present study |
| Staphylococcus
aureus USA300 (LAC) |
Methicillin-resistant clinical
isolate | (23) |
| Staphylococcus
aureus Newman |
Methicillin-resistant clinical
isolate | (23) |
| Staphylococcus
aureus MW2 |
Methicillin-resistant clinical
isolate | (23) |
| Staphylococcus
aureus MRSA1 |
Methicillin-resistant clinical
isolate | Present study |
| Staphylococcus
aureus MRSA2 |
Methicillin-resistant clinical
isolate | Present study |
| Staphylococcus
aureus ISP4790 | Clinical
isolate | (23) |
| Staphylococcus
aureus MU50 | Clinical
isolate | (23) |
Purification of bee venom
Controlled colonies of natural honeybees (Apis
mellifera L.) were maintained at room temperature at the
National Academy of Agricultural Science (Suwon, Korea). In brief,
a bee venom collector apparatus (Chunglin Biotech, Ansan, Korea)
was placed on the hive, and the bees that landed on the apparatus
were subjected to an electric shock sufficient to cause the bees to
'sting' a glass plate from which dried bee venom was harvested. The
collected venom was dissolved in distilled water, centrifuged at
12,000 × g for 10 min to remove insoluble materials, and stored in
a refrigerator until further use (15–17).
Bactericidal assay
Bacteria were harvested at the early log phase
(A600=0.5) and suspended in phosphate-buffered saline
(PBS) at ~108 to 1010 CFU/ml. Subsequently,
the bacterial samples were incubated with the indicated
concentrations of bee venom or melittin at 25°C for 30 min, and
surviving bacteria were evaluated using a plate counting method, as
described previously (18).
Briefly, samples were serially diluted in PBS and plated onto blood
agar (Kisan Bio, Suwon, Korea). Following a 16 h incubation at
37°C, the number of surviving bacteria was counted.
Determination of the minimum inhibitory
concentration
To determine the minimum inhibitory concentration
(MIC), the present study used a micro-dilution broth method,
according to the recommendations of the National Committee for
Clinical Laboratory Standards (19). In brief, the cells of the
experimental bacterial strains were collected in the logarithmic
phase of growth, suspended in 30 mM phosphate buffer (pH 7.0) with
60 mM NaCl, and adjusted to an A600 of 0.3 arbitrary
units (1×105 cells/ml). The bee venom and the melittin
samples were dissolved in 10 mM phosphate buffer (pH 6.0) with 130
mM NaCl and 0.2% (w/v) bovine serum albumin prior to serial
dilution. Sample aliquots (10 µl) were mixed with the
diluted bacterial suspensions (190 µl) followed by
incubation for 20 h at 37°C. Bacterial growth was determined by
measurement of the A650 levels using a VICTOR™ X3 ELISA
reader (PerkinElmer, Inc., Waltham MA, USA).
Cytotoxicity assays
The cytotoxic effects of bee venom and melittin on
cultured MCF7 cells were evaluated using a Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc., Gaithersburg, MD,
USA). The cells were seeded at a density of 5×103
cells/200 µl/well into wells of 96-well round-bottomed
plates and allowed to grow for 24 h at 37°C, followed by incubation
with bee venom or purified synthetic melittin for 6 h at 37°C. The
culture supernatants (100 µl quantities) were harvested and
mixed with 10 µl aliquots of CCK-8 solution. Following 3 h
incubation at 37°C, the optical densities at A450 were
measured using the VICTOR™ X3 ELISA reader (PerkinElmer, Inc.).
Mouse intraperitoneal infection
Mouse infection with Staphylococcus aureus
was performed, as described previously (20). Bacteria of the USA300 strain
(American Type Culture Collection, Manassas, VA, USA) were
spectrophotometrically (OPTIZEN POP; Mecasys Co., Ltd., Daejeon,
Korea) adjusted to the desired concentration prior to injection,
and bacterial numbers were confirmed via serial dilution and
Tryptic soy agar plating. The cultured USA300 bacteria were
pelleted, washed and suspended in PBS at 0.5×108 CFU/ml.
Mice (7-week-old males) of the CD1 strain were obtained from
Oriental Bio, Inc. (Seongnam, Korea), with 10 animals per treatment
group. The mice were infected with the USA300 strain (200
µl) via intraperitoneal (i.p.) injection, followed by i.p.
injection of 100 µl bee venom or purified melittin 1 h
later. The infected animals were monitored every 3 h for up to 36
h. The mice were housed in controlled conditions: Temperature,
23±2°C; humidity 55±10%; light between 07:00 and 19:00. Each group
was housed seperately. All animal experiments in the present study
adhered to institutional guidelines upon review of the experimental
protocol, and were approved by the Institutional Biosafety
Committee and the Institutional Animal Care and Use Committee of
Korea Atomic Energy Research Institute.
Mouse skin infection
CD1 mice (7-week old; 3 mice/group) were used to
examine skin infection. Following the induction of general
anesthesia, the dorsal hair was electrically shaved and the skin
was cleaned with 70% (v/v) ethanol. Skin infection was induced via
subcutaneous inoculation of 50 µl volumes of USA300
suspension (106 CFU/ml) in PBS. Subsequently, bee venom,
melittin (purified or synthetic; 100 µg in 80 µl
PBS), or sterile PBS was applied once daily to each surface lesion.
Lesion progression was monitored at 24 h intervals for 10 days by
measuring the lesion dimensions with callipers (Jeung Do B&P
Co., Ltd., Seoul, Korea), and capturing images using a digital
camera (WB5500; Samsung, Seoul, Korea).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analysis was conducted using GraphPad InStat software
version 5 (GraphPad Software, Inc., La Jolla, CA, USA). The
statistical significance of between-group differences was evaluated
using two-tailed Student's t-test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Bee venom exhibits a broad specrtum of
antimicrobial activity
The present study examined the antibacterial
activities of bee venom against the Streptococcus agalactiae,
Streptococcus gordonii, Streptococcus pneumonia, Streptococcus
epidermidis, Streptococcus bovis and Staphylococcus aureus
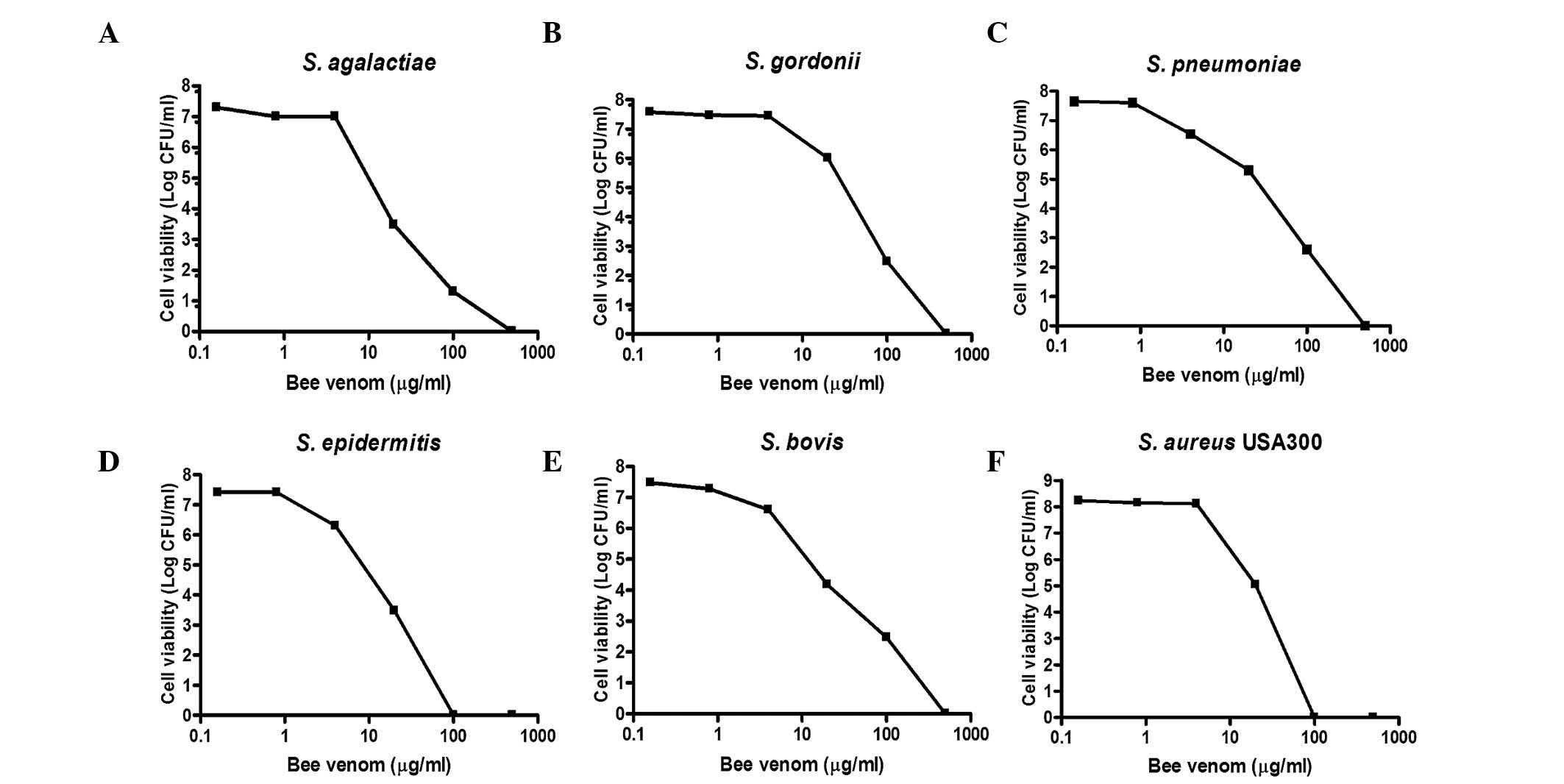
Gram-positive bacteria. As shown in Fig. 1, when all the bacterial strains
were treated with the indicated concentrations of bee venom for 30
min, concentration-dependent death of the bacteria was evident. At
venom concentrations between 1.25 and 12.5 µg/ml, bacterial
viability decreased by >90%. The MIC values of the bee venom
ranged between 1.56 and 12.5 µg/ml (Table II). Notably, the USA300
antibiotic-resistant Staphylococcus strain had the lowest observed
MIC (1.56 µg/ml).
| Table IIMIC of bee venom towards bacterial
strains. |
Table II
MIC of bee venom towards bacterial
strains.
| Bacterial
strain | MIC
(µg/ml) |
|---|
| Streptococcus
agalactiae CNCTC 10/84 | 6.25 |
| Streptococcus
gordonii M99 | 6.25 |
| Streptococcus
pneumonia TIGR4 | 3.12 |
| Streptococcus
epidermidis RP62a | 0.78 |
| Streptococcus
bovis NEM760 | 1.56 |
| Staphylococcus
aureus USA300 (LAC) | 0.78 |
| Staphylococcus
aureus Newman | 0.78 |
| Staphylococcus
aureus MW2 | 1.56 |
| Staphylococcus
aureus MRSA1 | 3.12 |
| Staphylococcus
aureus MRSA2 | 1.56 |
| Staphylococcus
aureus ISP4790 | 6.25 |
| Staphylococcus
aureus MU50 | 6.25 |
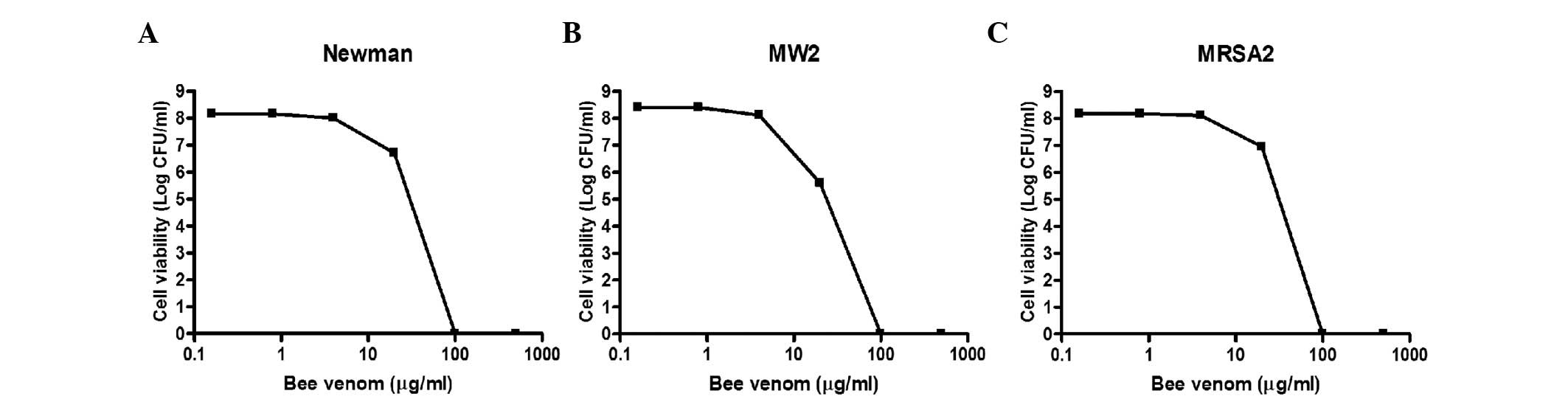
The present study further examined the antibacterial
activities of bee venom against three MRSA clinical isolates. As
shown in Fig. 2, the viabilities
of all three strains decreased markedly upon treatment with bee
venom for 30 min, and no bacteria survived incubation with 100
µg/ml venom. The MIC values for the three MRSA strains
ranged between 0.78 and 3.13 µg/ml (Table II). Notably, the
methicillin-sensitive Staphylococcus aureus strains (Mu50,
ISP479C, PS735, PS736 and PS737) were less susceptible to bee venom
(MIC=3.13–12.5 µg/ml), compared with the MRSA strains
(Table II), suggesting that bee
venom contains antimicrobial molecules, which specifically target
MRSA strains.
Bee venom protects against staphylococcal
infection
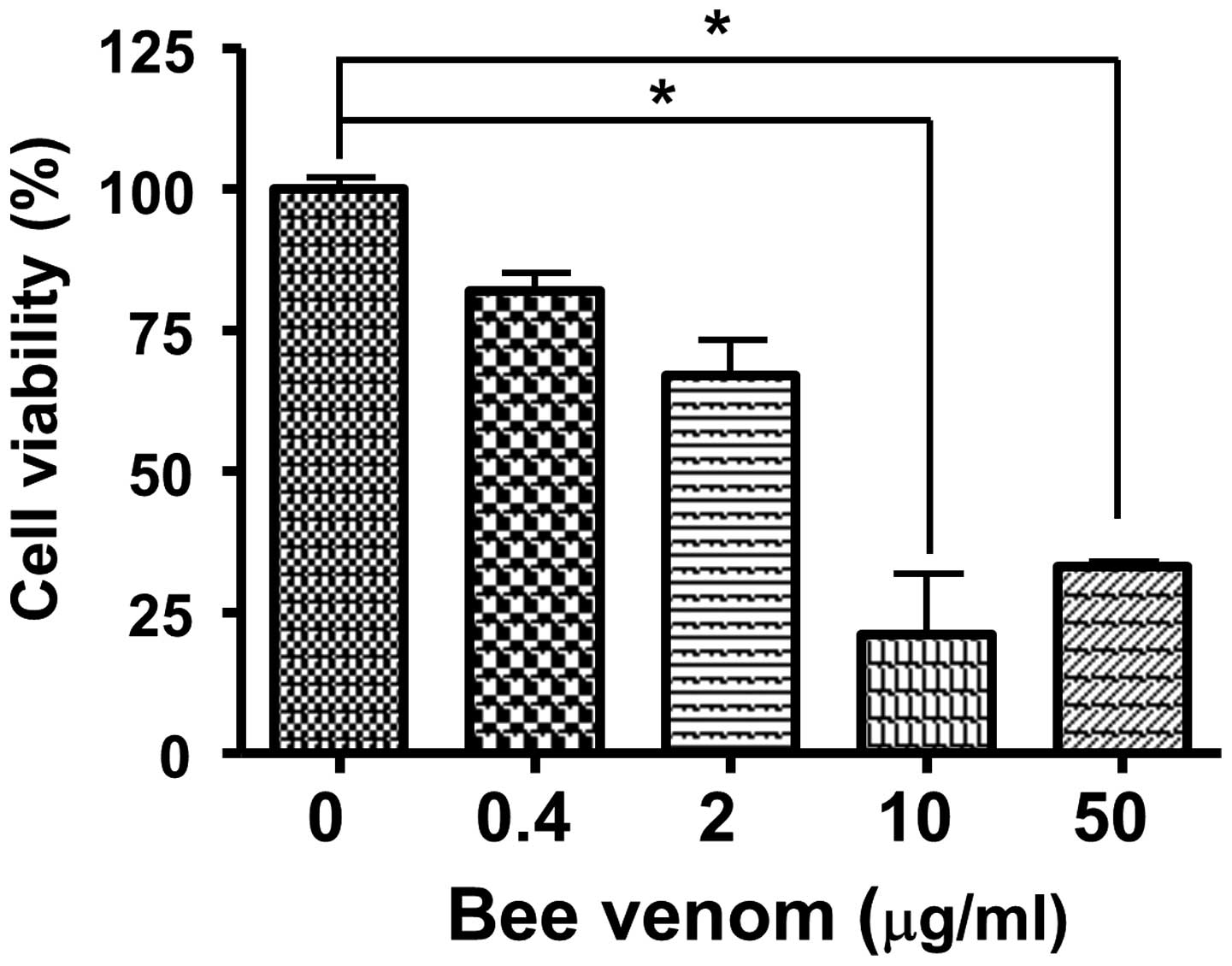
To measure the cytotoxicity of bee venom, human
epithelial cells were incubated with venom for 24 h and cell
viabilities were measured using an MTT assay. As shown in Fig. 3, bee venom was not cellulotoxic at
a concentration of 0.4 µg/ml. In addition, the
administration of bee venom in vivo at up to 20 mg/kg i.p.,
caused no signs or symptoms of toxicity in the CD1 mice (data not
shown).
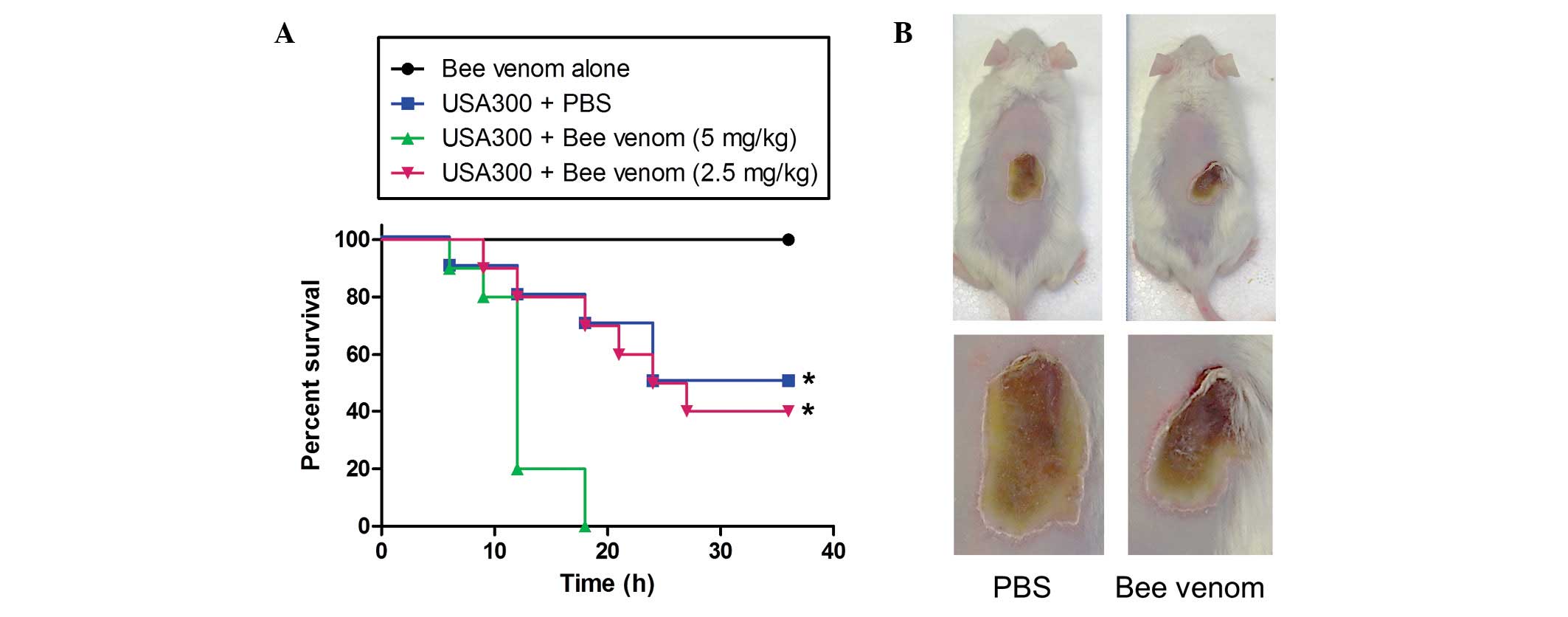
The i.p injection of 1×108 CFU of the
USA300 strain into mice caused bacteraemia and mortality rates of
100% within 18 h. When the USA300-infected mice were administered
with 1.25 or 2.5 mg/kg bee venom at the time of infection, no
protective effect was evident (data not shown). A low dose of
USA300 (1×107 CFU per mouse) was injected 1 h following
the administration of PBS or bee venom. Notably, all the mice died
18 h following the injection of USA300 with bee venom, whereas only
five mice of the control group had died by 24 h post-infection
(Fig. 4A). These data demonstrated
that, although bee venom exhibited a marked antimicrobial effect
in vitro, in vivo administration enhanced MRSA
propagation and infection.
In addition, the present study examined the
protective effect of bee venom in a staphylococcal skin infection
model (Fig. 4B). When USA300 was
inoculated intradermally and the areas of infected skin treated
with PBS or bee venom (10 µg) once daily, the abscesses
formed by USA300 were 21.3±4.8 and 18.8±6.8 mm in diameter in the
PBS and bee venom groups, respectively, by day 5, and no
significant difference was observed even following 10 days of venom
treatment.
Melittin is the major antimicrobial
component of bee venom
Bee venom is a complex mixture of proteins, peptides
and low-molecular-weight materials. The principal components of the
venom are phospholipase A2 (PLA2; 10–12%, w/w) and the melittin
peptide (40–48%, w/w). The results of the present study confirmed
and extended the previous results, demonstrating that melittin and
PLA2 induced death in a broad range of bacteria, including MRSA
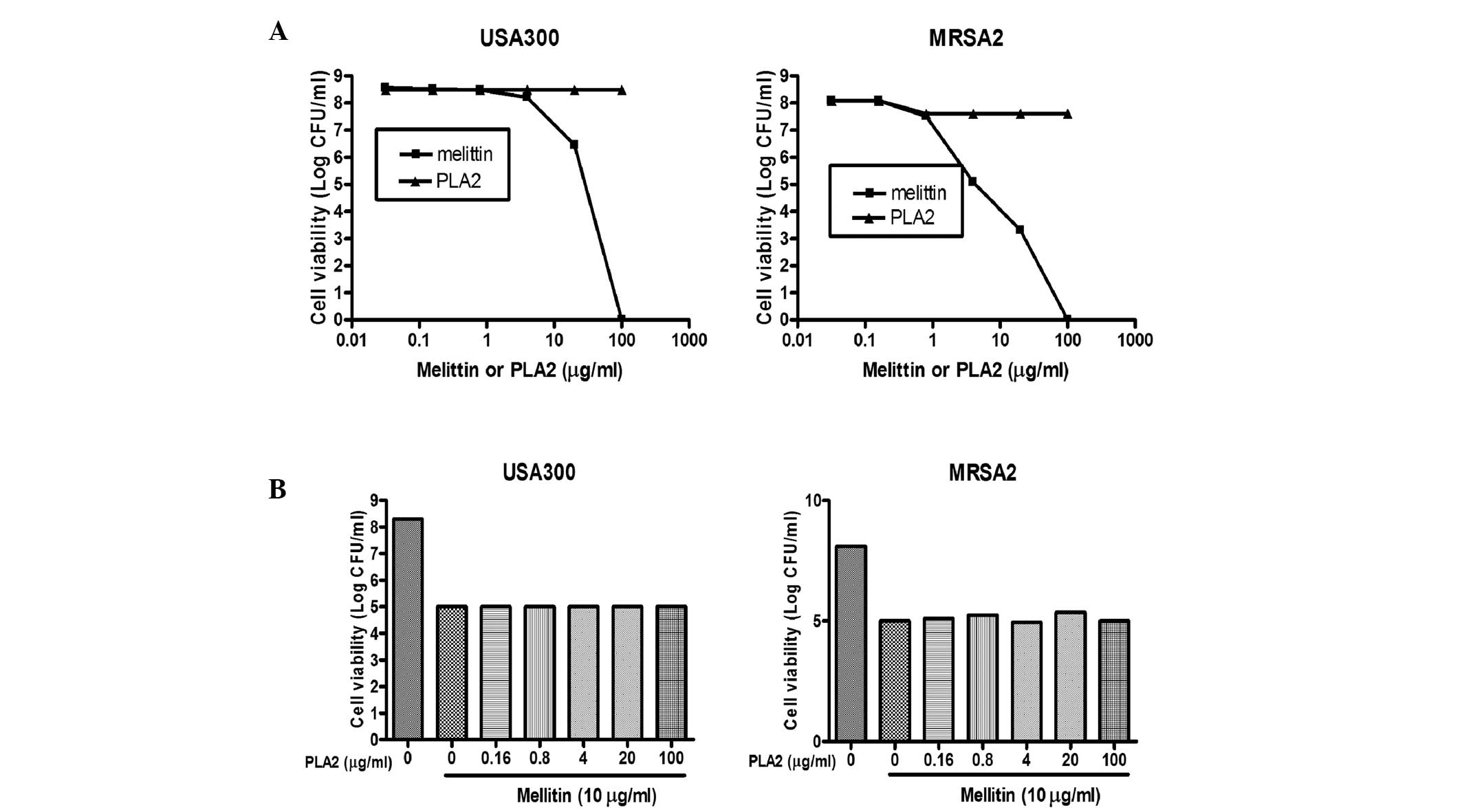
strains. As shown in Fig. 5A,
treatment of the USA300 and MRSA2 strains with PLA2 did not affect
cell viability, whereas the viabilities of the MRSA strains treated
with purified melittin decreased to levels comparable to those
observed when bee venom was used. To examine whether melittin and
PLA2 acted synergistically, two MRSA strains were treated with
melittin admixed with PLA2 at various concentrations. When the
USA300 and MRSA2 strains were treated with melittin alone (25
µg/ml), the total number of bacteria decreased by ~2.5–3 log
CFU (Fig. 5B). However, when the
cells were treated with melittin (25 µg/ml) in combination
with various concentrations of PLA2, similar results were observed,
indicating that PLA2 did not act synergistically with melittin to
cause bacterial cell death.
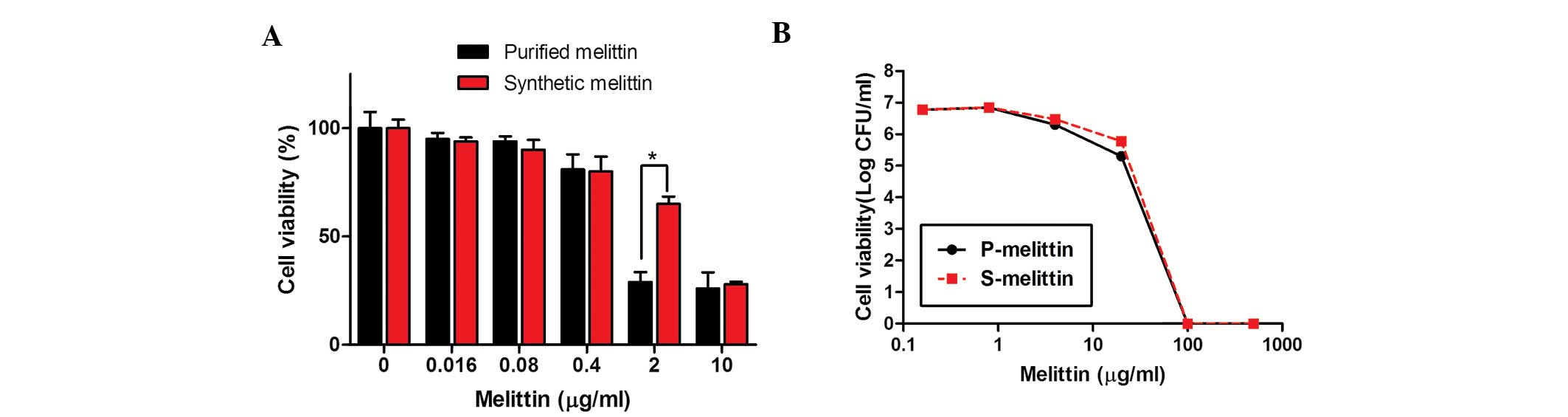
Subsequently, the present study confirmed that
synthetic melittin exhibited an antimicrobial activity similar to
that of purified melittin. Initially, the toxicities of the two
forms of melittin towards human epithelial cells were determined,
as described above. As shown in Fig.
6A, synthetic melittin (99.2% pure) was ~25% less toxic than
the 'purified' melittin (93% pure). However, the antibacterial
activities of the two preparations against the MRSAs were
comparable (Fig. 6B).
Protection from staphylococcal infection
by melittin
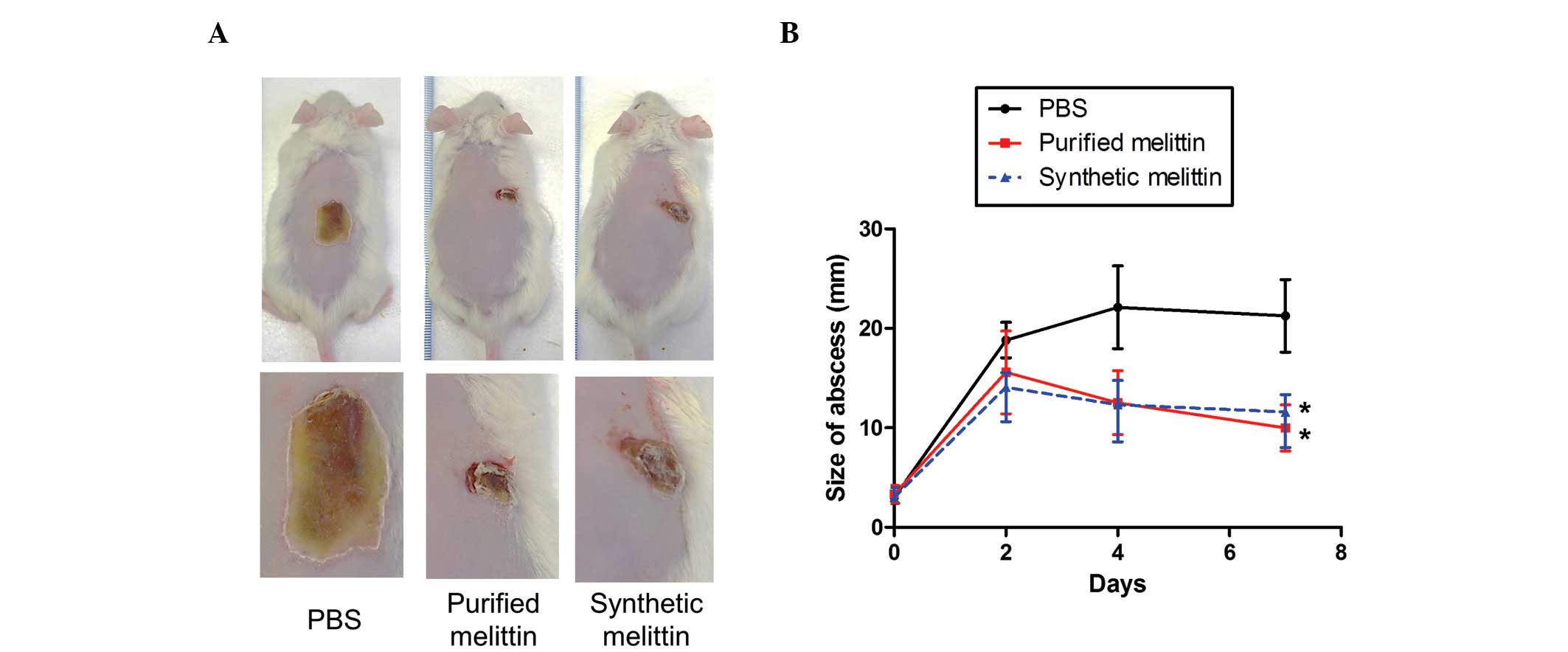
The present study also investigated whether melittin
can protect against MRSA skin infections. USA300 bacteria
(1×107 CFU/mouse) were injected intradermally into CD1
mice, which were administered with either PBS, or purified or
synthetic melittin (10 µg) 1 h post-infection. As shown in
Fig. 7, abscesses in the
PBS-treated group gradually increased in size to attain a diameter
of 22±6.3 mm by day 5. When the infected areas were treated with
purified or synthetic melittin once daily for 4 days, the diameters
of the abscesses were significantly lower than those measured in
the control group.
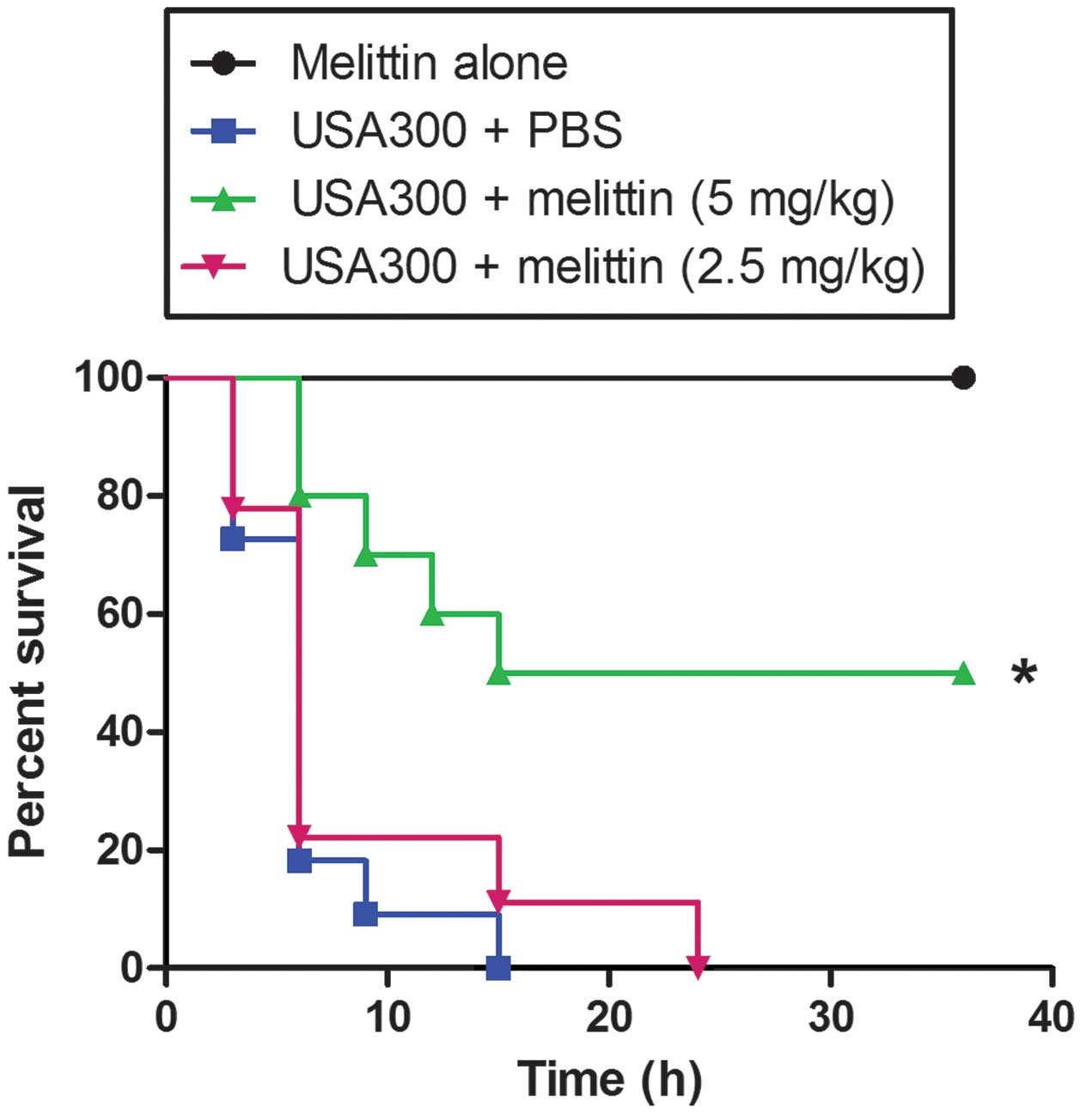
In addition, the protective effect of melittin was
investigated in a model of MRSA bacteraemia (Fig. 8). When a high dose of USA300 was
injected i.p., all the mice died following treatment with either
PBS or 2.5 mg/kg melittin after 24 h. However, when the infected
mice were injected with 5 mg/kg melittin 1 h post-infection, 50% of
the mice survived >24 h.
Discussion
Staphylococcus aureus is an important human
pathogen, which is responsible for the majority of bacterial soft
skin tissue infections and life-threatening infections, including
pneumonia, abscesses, endocarditis and infections of surgical sites
(2). The rapid spread of MRSA
strains is cause for alarm. The rates of MSRA infections are
increasing, and MRSA has become the leading cause of invasive
illness, resulting in a high rate of mortality worldwide (24–26).
Thus, the development of novel therapeutic methods is essential to
treat chronic wounds or systemic infections caused by MRSA. In the
present study, the in vitro anti-MRSA activities of the
natural antimicrobial components of bee venom were
investigated.
Bee venom contains several potential antibacterial
toxins, including melittin, PLA2, adolpanin, dopamine and
hyaluronidase (27). Each
component may exert selective and specific actions on human cells
and/or bacteria (16,28). Although the bee venom isolated in
the present study exhibited potential antimicrobial activities
against all the Gram-positive bacteria assessed in vitro, as
has been reported in several previous studies (9,11,29),
the i.p. administration of venom into MRSA-infected mice caused the
a higher mortality rate, compared with that observed in the
venom-free controls, suggesting that bee venom actually facilitated
MRSA infection. Notably, the PLA2 of bee venom is central to the
proinflammatory cascade by activating several physiological and
pathogenic immune activities (30,31).
In addition, certain hypervirulent bacteria produce and secrete
PLA2, which significantly potentiates early-stage infection and
inflammation (32–35). The present study also found that,
although PLA2 exhibited minimal antibacterial activity, i.p.
injection of MRSA-infected mice with PLA2 caused 100% mortality,
whereas only 50% mortality was observed in the control animals by
24 h, which was also true of the bee venom-treated mice (data not
shown). Thus, it is reasonable to suggest that PLA2 increased the
susceptibility of at-risk hosts to bacterial infection.
Melittin is the principal component (40–48%, w/w) of
honeybee venom (12), being a
small linear peptide of 26 amino acids forming an amphipathic helix
with a hydrophobic amino- and hydrophilic carboxyl-terminus. The
antibacterial effects of melittin have been widely investigated
in vitro (36). In the
present study, synthetic melittin exhibited anti-MRSA toxicity
in vitro, which was comparable to that of purified melittin.
However, the synthetic melittin was less toxic towards human
epithelial cells, suggesting that the purified melittin (93% pure)
in the present study contained an uncharacterized component, which
is either toxic and/or enhances the toxicity of melittin. Following
acquisition of these in vitro results, the present study
examined the protective effects of melittin in MRSA-infected mice.
Unlike bee venom, melittin exhibited significantly higher
protective effects in vivo in the models of bacteraemia and
skin infection. Although melittin directly affects microbes by
damaging or destabilising cell membranes, the material appears to
potentiate the innate immune and anti-inflammatory responses,
preventing the development of MRSA systemic infections and
facilitating wound healing around infected sites (14,37–39).
Melittin exerts anti-inflammatory effects on several types of cell
(38,40,41).
Melittin suppresses innate immune signaling, including that
mediated by nuclear factor-κB via Toll-like receptor and mitogen
activated protein kinase; the synthesis of cyclooxygenase-2; and
the expression of inducible nitric oxide synthase (38,39).
In addition, melittin stimulates pyrin domain-containing
inflammasomes to activate caspase-1 and interleukin1β, which
crucially recruit neutrophils to sites of expression (14,40,42).
Thus, melittin may inhibit MRSA infections by several mechanisms,
including the direct induction of MRSA cell death, the
downregulation of the innate immune response induced by MRSA and
the acceleration of neutrophil recruitment to sites of
infection.
Together, the results of the present study
demonstrated that bee venom, which is intrinsically toxic, exerts
negative effects when used as an anti-MRSA therapy. However, the
principal component of bee venom, melittin, exhibits antibacterial
effects with minimal toxicity in vitro and in vivo.
To the best of our knowledge, the present study is the first to
demonstrate that melittin may exert a possible therapeutic role in
the treatment of MRSA infections. The mechanism of this effect
requires further investigation.
Acknowledgments
This study was supported by grants from the Nuclear
R&D program of the Ministry of Science, ICT and Future planning
(grant no. 523330) to Dr Sangyong Lim and the Next BioGreen21
Program, Rural Development Administration, Republic of Korea
(grant. no. PJ009534) to Dr Joo-Hong Yeo.
References
|
1
|
Rasigade JP and Vandenesch F:
Staphylococcus aureus: A pathogen with still unresolved issues.
Infect Genet Evol. 21:510–514. 2014. View Article : Google Scholar
|
|
2
|
Taylor AR: Methicillin-resistant
Staphylococcus aureus infections. Prim Care. 40:637–654. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Limbago BM, Kallen AJ, Zhu W, Eggers P,
McDougal LK and Albrecht VS: Report of the 13th
vancomycin-resistant Staphylococcus aureus isolate from the United
States. J Clin Microbiol. 52:998–1002. 2014. View Article : Google Scholar :
|
|
4
|
Corey GR: Staphylococcus aureus
bloodstream infections: Definitions and treatment. Clin Infect Dis.
48(Suppl 4): S254–S259. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gould IM: VRSA-doomsday superbug or damp
squib? Lancet Infect Dis. 10:816–818. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bassetti M, Merelli M, Temperoni C and
Astilean A: New antibiotics for bad bugs: Where are we? Ann Clin
Microbiol Antimicrob. 12(22)2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Annila I: Bee venom allergy. Clin Exp
Allergy. 30:1682–1687. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Son DJ, Lee JW, Lee YH, Song HS, Lee CK
and Hong JT: Therapeutic application of anti-arthritis,
pain-releasing and anti-cancer effects of bee venom and its
constituent compounds. Pharmacol Ther. 115:246–270. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim JY, Lee WR, Kim KH, An HJ, Chang YC,
Han SM, Park YY, Pak SC and Park KK: Effects of bee venom against
Propionibacterium acnes-induced inflammation in human keratinocytes
and monocytes. Int J Mol Med. 35:1651–1656. 2015.PubMed/NCBI
|
|
10
|
Lee H, Lee EJ, Kim H, Lee G, Um EJ, Kim Y,
Lee BY and Bae H: Bee venom-associated Th1/Th2 immunoglobulin class
switching results in immune tolerance of NZB/W F1 murine lupus
nephritis. Am J Nephrol. 34:163–172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Perumal Samy R, Gopalakrishnakone P, Thwin
MM, Chow TK, Bow H, Yap EH and Thong TW: Antibacterial activity of
snake, scorpion and bee venoms: A comparison with purified venom
phospholipase A2 enzymes. J Appl Microbiol. 102:650–659. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gajski G and Garaj-Vrhovac V: Melittin: A
lytic peptide with anticancer properties. Environ Toxicol
Pharmacol. 36:697–705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Adade CM, Oliveira IR, Pais JA and
Souto-Padron T: Melittin peptide kills Trypanosoma cruzi parasites
by inducing different cell death pathways. Toxicon. 69:227–239.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jo M, Park MH, Kollipara PS, An BJ, Song
HS, Han SB, Kim JH, Song MJ and Hong JT: Anti-cancer effect of bee
venom toxin and melittin in ovarian cancer cells through induction
of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol
Appl Pharmacol. 258:72–81. 2012. View Article : Google Scholar
|
|
15
|
Han SM, Lee GG and Park KK: Acute dermal
toxicity study of bee venom (Apis mellifera L.) in rats. Toxicol
Res. 28:99–102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han SM, Lee KG, Park KK and Pak SC: Skin
sensitization study of bee venom (Apis mellifera L.) in guinea pigs
and rats. Cutan Ocul Toxicol. 32:27–30. 2013. View Article : Google Scholar
|
|
17
|
Han SM, Lee GG and Park KK: Skin
sensitization study of bee venom (Apis mellifera L.) in guinea
pigs. Toxicol Res. 28:1–4. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Seo HS, Mu R, Kim BJ, Doran KS and Sullam
PM: Binding of glycoprotein Srr1 of Streptococcus agalactiae to
fibrinogen promotes attachment to brain endothelium and the
eevelopment of meningitis. PLoS Pathog. 8:e10029472012. View Article : Google Scholar
|
|
19
|
Clinical and Laboratory Standards
Institute: M100-S16, Performance standards for antimicrobial
susceptibility testing; 16th informational supplement. Clinical and
Laboratory Standards Institute; Wayne, PA: 2007
|
|
20
|
Ganesh VK, Rivera JJ, Smeds E, Ko YP,
Bowden MG, Wann ER, Gurusiddappa S, Fitzgerald JR and Höök M: A
structural model of the Staphylococcus aureus ClfA-fibrinogen
interaction opens new avenues for the design of anti-staphylococcal
therapeutics. PLoS Pathog. 4:e10002262008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bensing BA, Gibson BW and Sullam PM: The
Streptococcus gordonii platelet binding protein GspB undergoes
glycosylation independently of export. J Bacteriol. 186:638–645.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Seo HS, Cartee RT, Pritchard DG and Nahm
MH: A new model of pneumococcal lipoteichoic acid structure
resolves biochemical, biosynthetic and serologic inconsistencies of
the current model. J Bacteriol. 190:2379–2387. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qian Z, Yin Y, Zhang Y, Lu L, Li Y and
Jiang Y: Genomic char-acterization of ribitol teichoic acid
synthesis in Staphylococcus aureus: Genes, genomic organization and
gene duplication. BMC Genomics. 7(74)2006. View Article : Google Scholar
|
|
24
|
Goldrick BA: MRSA, VRE, and VRSA: How do
we control them in nursing homes? Am J Nurs. 104:50–51. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hebert C and Weber SG: Common approaches
to the control of multidrug-resistant organisms other than
methicillin-resistant Staphylococcus aureus (MRSA). Infect Dis Clin
North Am. 25:181–200. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Todd B: Beyond MRSA: VISA and VRSA: What
will ward off these pathogens in health care facilities? Am J Nurs.
106:28–30. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park D, Jung JW, Lee MO, Lee SY, Kim B,
Jin HJ, Kim J, Ahn YJ, Lee KW, Song YS, et al: Functional
characterization of naturally occurring melittin peptide isoforms
in two honey bee species, Apis mellifera and Apis cerana. Peptides.
53:185–193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Palm NW and Medzhitov R: Role of the
inflammasome in defense against venoms. Proc Natl Acad Sci USA.
110:1809–1814. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fennell JF, Shipman WH and Cole LJ:
Antibacterial action of a bee venom fraction (melittin) against a
penicillin-resistant staphylococcus and other microorganisms.
USNRDL-TR-67-101. Res Dev Tech Rep. 5:1–13. 1967.
|
|
30
|
Putz T, Ramoner R, Gander H, Rahm A,
Bartsch G, Bernardo K, Ramsay S and Thurnher M: Bee venom secretory
phospholipase A2 and phosphatidylinositol-homologues cooperatively
disrupt membrane integrity, abrogate signal transduction and
inhibit proliferation of renal cancer cells. Cancer Immunol
Immunother. 56:627–640. 2007. View Article : Google Scholar
|
|
31
|
Carballido JM, Carballido-Perrig N,
Schwärzler C and Lametschwandtner G: Regulation of human T helper
cell differentiation by antigen-presenting cells: The bee venom
phospholipase A2 model. Chem Immunol Allergy. 91:147–158. 2006.
View Article : Google Scholar
|
|
32
|
Lapointe S, Brkovic A, Cloutier I, Tanguay
JF, Arm JP and Sirois MG: Group V secreted phospholipase A2
contributes to LPS-induced leukocyte recruitment. J Cell Physiol.
224:127–134. 2010.PubMed/NCBI
|
|
33
|
Sitkiewicz I, Stockbauer KE and Musser JM:
Secreted bacterial phospholipase A2 enzymes: Better living through
phospholipolysis. Trends Microbiol. 15:63–69. 2007. View Article : Google Scholar
|
|
34
|
Hunt CL, Nauseef WM and Weiss JP: Effect
of D-alanylation of (lipo) teichoic acids of Staphylococcus aureus
on host secretory phospholipase A2 action before and after
phagocytosis by human neutrophils. J Immunol. 176:4987–4994. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Koprivnjak T, Peschel A, Gelb MH, Liang NS
and Weiss JP: Role of charge properties of bacterial envelope in
bactericidal action of human group IIA phospholipase A2 against
Staphylococcus aureus. J Biol Chem. 277:47636–47644. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fennell JF, Shipman WH and Cole LJ:
Antibacterial action of melittin, a polypeptide from bee venom.
Proc Soc Exp Biol Med. 127:707–710. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Park JH, Kim KH, Lee WR, Han SM and Park
KK: Protective effect of melittin on inflammation and apoptosis in
acute liver failure. Apoptosis. 17:61–69. 2012. View Article : Google Scholar
|
|
38
|
Park HJ, Lee HJ, Choi MS, Son DJ, Song HS,
Song MJ, Lee JM, Han SB, Kim Y and Hong JT: JNK pathway is involved
in the inhibition of inflammatory target gene expression and
NF-kappaB activation by melittin. J Inflamm (Lond). 5:72008.
View Article : Google Scholar
|
|
39
|
Moon DO, Park SY, Choi YH, Kim ND, Lee C
and Kim GY: Melittin induces Bcl-2 and caspase-3-dependent
apoptosis through downregulation of Akt phosphorylation in human
leukemic U937 cells. Toxicon. 51:112–120. 2008. View Article : Google Scholar
|
|
40
|
Sommer A, Fries A, Cornelsen I, Speck N,
Koch-Nolte F, Gimpl G, Andrä J, Bhakdi S and Reiss K: Melittin
modulates keratinocyte function through P2 receptor-dependent ADAM
activation. J Biol Chem. 287:23678–23689. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dempsey CE: The actions of melittin on
membranes. Biochim Biophys Acta. 1031:143–161. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim SJ, Park JH, Kim KH, Lee WR, Kim KS
and Park KK: Melittin inhibits atherosclerosis in LPS/high-fat
treated mice through athero-protective actions. J Atheroscler
Thromb. 18:1117–1126. 2011. View Article : Google Scholar
|