Introduction
Y-box binding protein 1 (YB-1), a member of the
highly evolutionarily conserved Y-box family, has an important role
in cell physiological activities, including gene transcription
(1–3), translation (4,5),
damage repair and cancer drug resistance (6,7). It
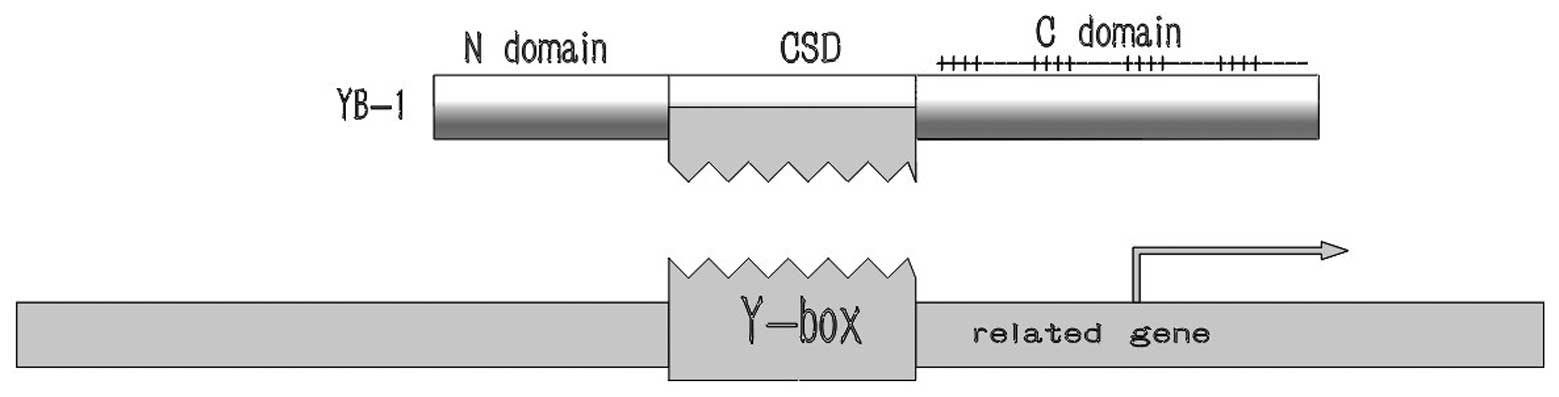
consists of three principal domains (Fig. 1): i) The N-terminal domain, also
termed the A/P domain, is rich in alanine and proline and involved
in transcriptional regulation. ii) The middle part is the cold
shock domain (CSD), a highly conserved sequence, which performs the
core function, i.e., it binds to the Y-box, an inverted CCAAT box
in the promoter of numerous specific genes, including major
histo-compatibility complex class II, multi-drug resistance gene 1
as well as cyclin A and B1 (8–10).
iii) An elongated C-terminal domain, containing alternating
clusters of positively and negatively charged amino-acid residues,
which is implicated in protein-protein interactions and serves as a
pivotal signaling peptide during nuclear translocation (11).
Several studies have reported that CSD is implicated
in the regulation of the transcription of the α1- and α2-strands of
collagen I [COLα1(I) and COLα2(I)], which are intrinsically linked
to the development of hepatic fibrosis (12). Type I collagen is a heterotrimer
composed of two α1-strands and one α2-strand. YB-1 was reported to
suppress the transcription of the COLα1(I) gene via binding to
collagen Y-box element or transforming growth factor (TGF)-β
response element in the promoter of COLα1(I) (13,14).
Furthermore, YB-1 was confirmed to be involved in interferon-γ
(IFN-γ)-induced down-regulation of the COLα2(I) gene (15) through binding to the
IFN-γ-responsive element in the promoter of COLα2(I) (16).
YB-1 regulates pro-collagen I expression in the
nucleus only; therefore, nuclear translocation of YB-1 is essential
for it to exert its function. Agents with the ability to promote
the nuclear translocation of YB-1 may have beneficial effects on
hepatic fibrosis. In previous studies, p53 (17), ultraviolet irradiation (18) and the small molecule HSc025
(19) were demonstrated to enhance
the nuclear translocation of YB-1.
Oxymatrine (OMT) is the major active component of
the Chinese medicinal herb Kushen, which is the dried root of
Sophora flavescens. The anti-viral (20), antioxidative (21,22),
anti-fibrotic (23) and
immunoregulatory effects (24,25)
of OMT have been demonstrated in numerous studies (20,21,23–25).
At present, OMT is widely applied in the clinic to protect the
liver from fibrosis.
The present study assessed whether OMT may exert its
long-known effects against hepatic fibrosis via promoting the
nuclear translocation of YB-1 to suppress the production of
pro-collagen I. The present study considerably enhanced the current
understanding of the underlying mechanism of the anti-fibrotic
effects of OMT.
Materials and methods
Cell culture
The LX-2 human hepatic stellate cell (HSC) line
(Shanghai Fuxiang Biotechnology Co., Ltd, Shanghai, China), was
cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS; Invitrogen) and kept in an incubator
containing 5% CO2 at 37°C. The cells were sub-cultured
every 3–4 days.
Drug treatment
OMT (BOC Sciences, New York, NY, USA) was dissolved
in complete medium to produce a stock solution. Prior to use in the
assays, cells were synchronized by serum-starvation for 24 h. Cells
(5×105) were then seeded into 60 mm dishes.
Subsequently, OMT solution was added to generate a series
concentrations of OMT (240, 480 and 960 mg/l). Furthermore, a
control group without OMT treatment was established. The cells were
incubated for 3–5 days prior to analysis.
Reverse-transcription quantitative
polymerase chain reaction (RT-qPCR) analysis
RNA from LX2 cells was extracted by using TRI
Reagent® (Sigma-Aldrich, St. Louis, MO, USA) according
to the manufacturer's instructions. cDNA was synthesized from 1
μg RNA using the FastQuant RT kit (Tiangen, Beijing, China)
according to the manufacturer's protocol. Real Time PCR mixtures
(10 μl) were then prepared in triplicate, each containing 1
μl cDNA template, 5 μl of SYBR® Premix Ex
Taq (TaKara, Japan), 0.2 ul of ROX Reference Dye II (Takara
Bio, Inc., Otsu, Japan), 0.2 μl primer (10 mM; Genewiz,
Inc., Suzhou, China), and 3.6 ml DEPC-treated water. Primers were
as follows: COLα1(I), 5'-CATGTTCAGCTTTGTGGACC-3' (forward) and
5'-TTCTGTACGCAGGTGATTGG-3' (reverse); COLα2(I),
5'-TGCTGCTCAGTATGATGGAAA-3' (forward) and
5'-CAGGTCCTTGGAAACCTTGA-3' (reverse); YB-1,
5'-TCGCCAAAGACAGCCTAGAGA-3' (forward) and 5'-TCT
GCGTCGGTAATTGAAGTTG-3' (reverse); α-SMA, 5'-AGC
AGTTGTAGCTACCCGCC-3' (forward) and 5'-CACGAT GGACGGGAACAC-3'
(reverse). GAPDH, 5'-GAAGGTGAA GGTCGGAGTC-3' (forward) and
5'-GAAGATGGTGAT GGGATTTC-3' (reverse). cDNA samples were amplified
in an Applied Biosystems 7500 Real-Time PCR system (Thermo Fisher
Scientific). The thermocycling conditions included initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec and 60°C for 34 sec. Finally all quantified values were
normalized to the endogenous GAPDH control. Gene expression levels
were analyzed using the ΔΔCt method (26).
Transient transfection
LX2 cells in 60 mm cell culture dishes without serum
were cultured to ~50% confluence on the second day. The cells were
transiently transfected with 300 pmol small interfering (si)RNA
(Gene Pharma, Shanghai, China) targeting YB-1 (siYB-1) or negative
control siRNA (NC) mixed with 15 μl Lipofectamine 2000
(Invitrogen) at 6 h prior to treatment with 960 mg/l OMT. Cells
were harvested after incubation for four days. The siYB-1 sequence
was UGA CACCAAGGAAGAUGUATT and the NC sequence was
UUCUCCGAACGUGUCACGUTT.
Nuclear/cytoplasmic extraction and
western blot analysis
Nuclear and cytoplasmic proteins were separated
using the Nuclear and Cytoplasmic Protein Extraction kit (Beyotime
Institute of Biotechnology, Shanghai, China). The separation
procedure was performed with caution to avoid contamination of the
nuclear protein in the pellet with cytoplasmic extract.
Cytoplasmic and nuclear extracts and total protein
samples were quantified using a Bichinchoninic Acid Protein assay
kit (Beyotime Institute of Biotechnology, Shanghai, China) and then
mixed with 4X Protein SDS-PAGE loading buffer (Takara Bio, Inc.) at
100°C for 20 min. Proteins were separated on 10–15% SDS-PAGE
(Invitrogen; Thermo Fisher Scientific) and transferred onto
polyvinylidene Fluoride membranes (EMD Millipore, Billerica, MA,
USA). Following blocking with Tris-Buffered Saline and Tween 20
(TBST) with 5% fat-free milk, the membranes were incubated with the
following primary antibodies overnight at 4°C: Anti-α-smooth muscle
actin (α-SMA; cat. no. ab32575), anti-extracellular
signal-regulated kinase 1 and 2 (ERK1/2; cat. no. ab17942),
anti-YB-1 (cat. no. ab12148; all 1:1,000; Epitomics, Burlingame,
CA, USA), as well as anti-GAPDH (cat. no. ab2302) and anti-histone3
(cat. no. 2514256; both 1:1,000; Millipore, Billerica, MA, USA).
The membranes were then washed with TBST for 30 min and incubated
with secondary antibodies conjugated with horseradish peroxidase
for 2 h at room temperature. Finally the images were captured using
a gel imaging analysis system (Tanon 4100, Tanon Science and
Technology Co., Ltd., Shanghai, China).
Statistical analysis
Blots were representative of three independent
experiments performed in triplicate. Statistical analysis of the
data was performed using Student's t-test and analyzed using SPSS
software version 13.0 (SPSS, Inc., Chicago, IL, USA). Data were
expressed as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
OMT reduces the expression of endogenic
type I pro-collagen
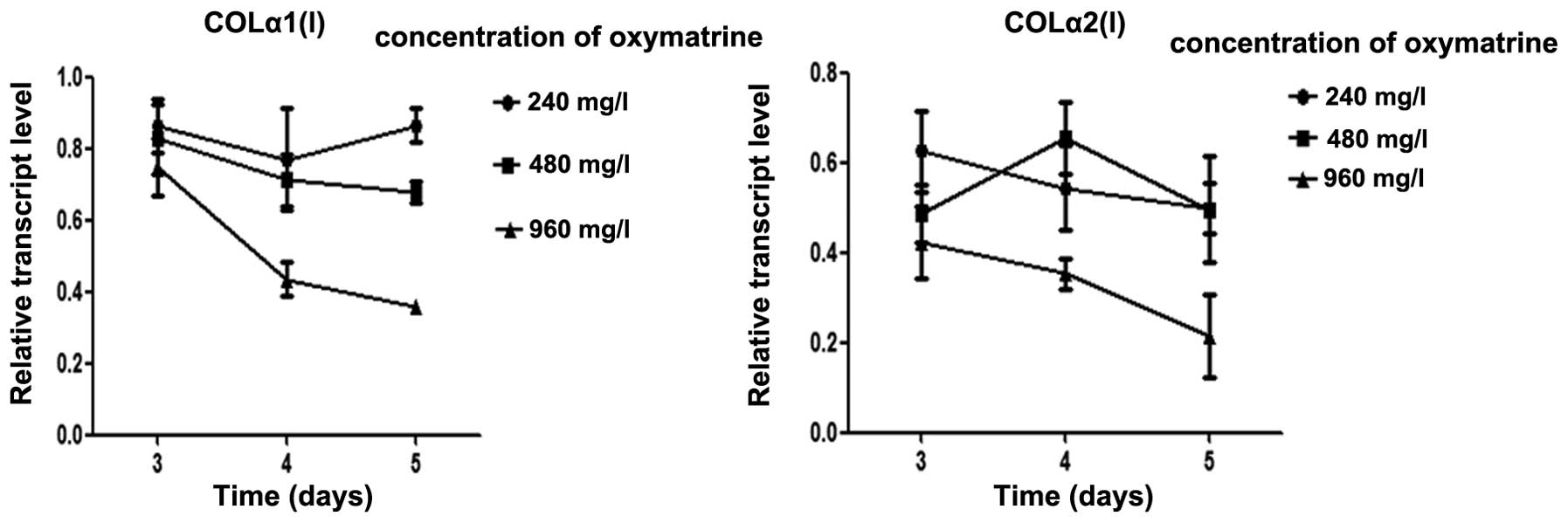
RT-qPCR analysis indicated a marked decrease in
COLα1(I) and COLα2(I) gene expression following incubation with OMT
(240, 480 and 960 mg/l for 3–5 days) in a dose- and time-dependent
manner (Fig. 2). The high
concentration of OMT (960 mg/l) was then used in the subsequent
experiments.
OMT accelerates nuclear translocation of
YB-1
Transcription factor YB-1 is known to inhibit
pro-collagen I production (13–16).
However, factors modulating the activity of YB-1 have remained to
be elucidated.
The present study hypothesized that OMT inhibits the
expression of pro-collagen I via facilitating the nuclear
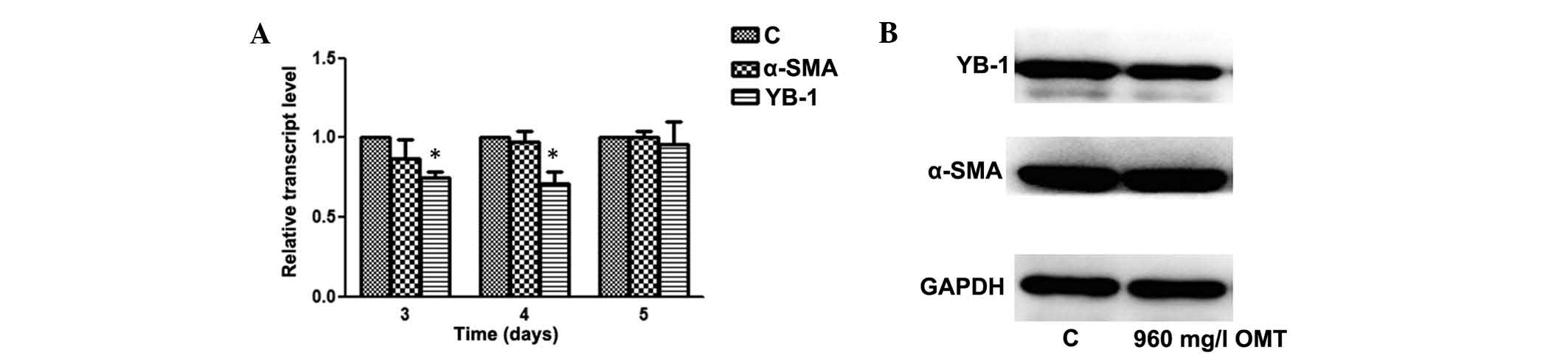
translocation of YB-1. In order to test this hypothesis, LX-2 cells
were treated with OMT (960 mg/l), which resulted in a reduction in
YB-1 expression at the transcriptional and translational level,
particularly on the fourth day of incubation, while α-SMA
expression was not affected (Fig.
3). As it is well known that YB-1 regulates gene transcription
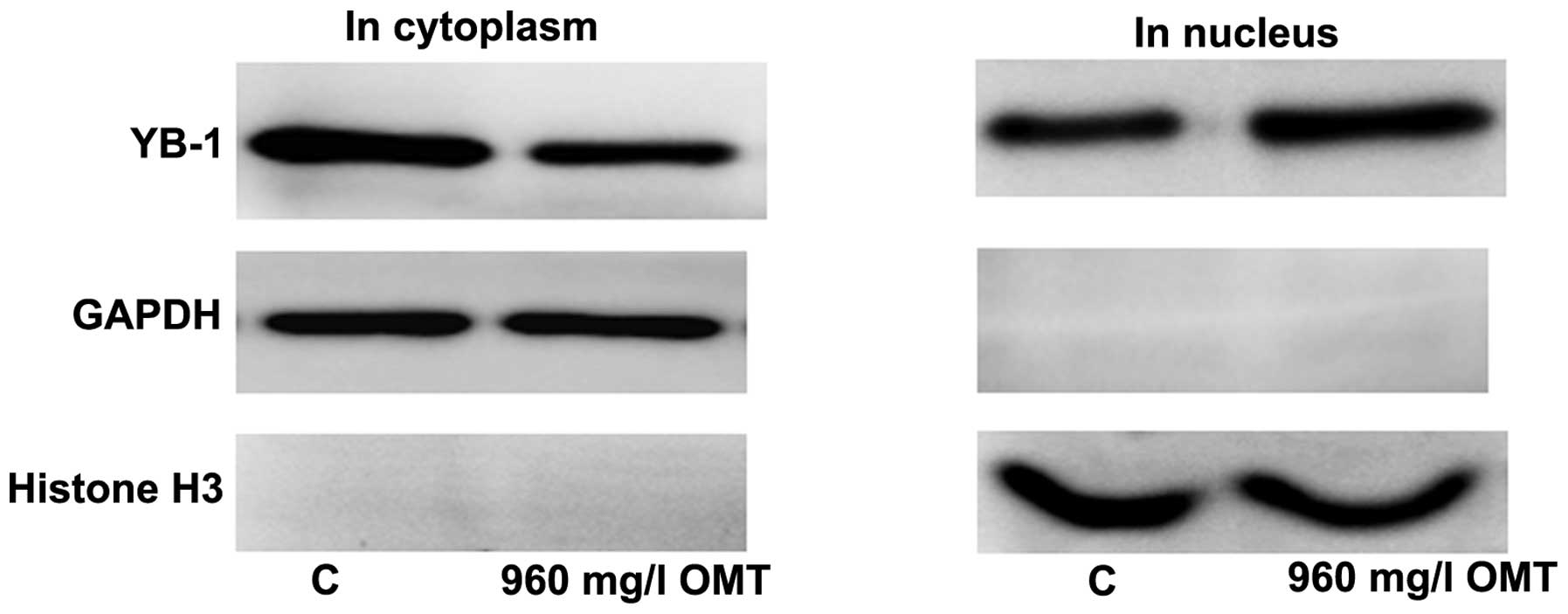
in the nucleus only, the present study examined the expression of
YB-1 in the cytoplasm and nucleus separately. The results showed
that LX-2 cells treated with 960 mg/l OMT for four days exhibited a
decrease of YB-1 in the cytoplasm and an increase of YB-1 in the
nucleus as compared with that in the control group (Fig. 4).
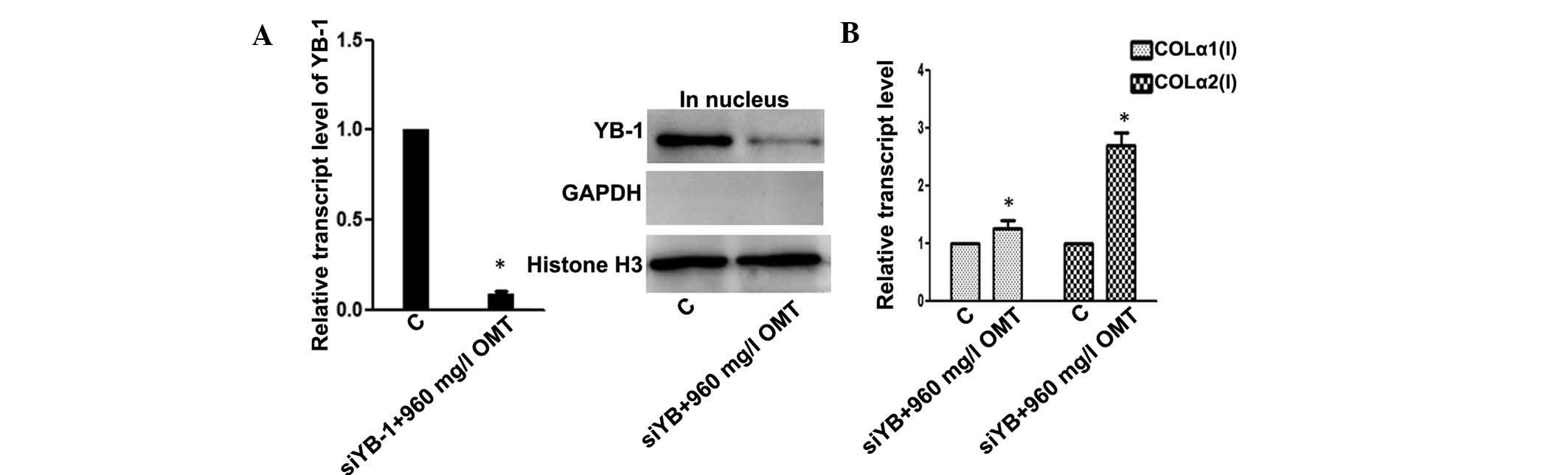
Depletion of YB-1 reverses the inhibitory
effects of OMT on pro-collagen I production
In the abovementioned experiments, OMT restrained
the expression of pro-collagen I, at least in part, by enhancing
the nuclear translocation of YB-1. In order to further verify this
mechanism, the effects of YB-1 knockdown on the expression levels
of pro-collagen I were assessed. As shown in Fig. 5A, transient transfection with
siYB-1 successfully knocked down YB-1. As expected, siYB-1
abrogated the inhibitory effects of OMT (960 mg/l) on pro-collagen
I synthesis, as indicated by significantly elevated mRNA levels of
COLα1(I) and an even greater elevation of COLα2(I) mRNA expression
(Fig. 5B).
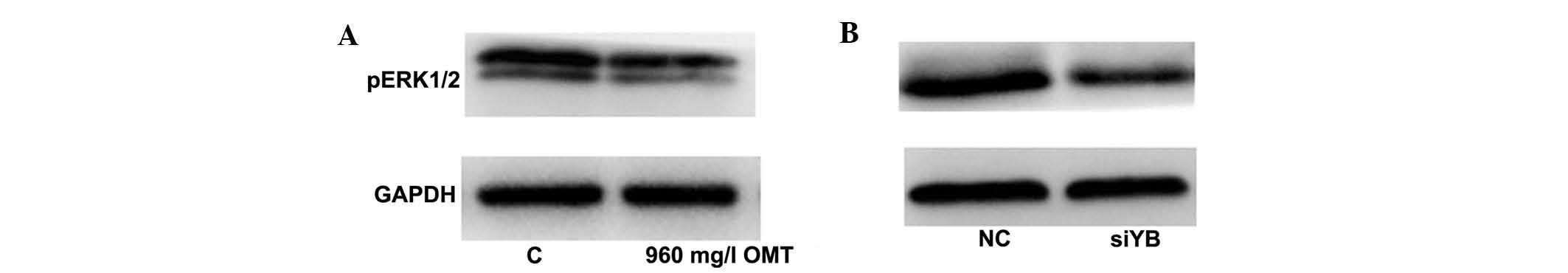
The ERK1/2 signaling pathway is involved
in the anti-fibrotic effects of YB-1
Since YB-1 participates in the negative regulation
of fibrosis, the present study investigated the associated
signaling pathways. It was found that the phosphorylation level of
ERK1/2 proteins was positively associated with the changes in YB-1
expression. In the cells treated with OMT (960 mg/l),
phosphorylated ERK1/2 proteins and YB-1 were declined compared to
those in the control group. Furthermore, the levels of
phosphorylated ERK1/2 proteins were also decreased as a result of a
knockdown of YB-1 in LX-2 cells, indicating that the ERK1/2
signaling pathway have a significant role in OMT-mediated
inhibition of pro-collagen I expression (Fig. 6).
Discussion
It is thought that patients with hepatocellular
carcinoma (HCC) present with hepatitis or cirrhosis prior to
developing carcinoma, with fibrosis having a central role in the
pathology of all of these conditions (27). Furthermore, overexpression of YB-1,
a multifunctional transcription factor, was observed in HCC, which
made it a promising prognostic biomarker used in the clinic
(28,29).
However, excess YB-1 can repress type I pro-collagen
expression, resulting in a reduction of fibrosis. It is well known
that the TGF-β/Smad signaling pathway is implicated in pro-collagen
I transcription. Furthermore, YB-1 was confirmed to be an
antagonist of TGF-β, not only by inducing overexpression of Smad7,
but also by inhibiting the p300-mediated activation of Smad3 and
its binding to TGF-β-responsive element in the promoter of COLα2(I)
(16).
Although the association between OMT and
downregulation of type I collagen is well established (30–32),
the precise mechanism has largely remained elusive. The findings of
the present study suggested that OMT stimulated the nuclear
translocation of YB-1, thereby inhibiting the endogenic expression
of type I pro-collagen.
It was effectively verified that OMT suppressed the
expression of COLα1(I) and COLα2(I) genes in a dose- nd
time-dependent manner. Furthermore, α-SMA, which is expressed in
activated HSCs only, was not significantly affected, indicating
that OMT did not cause the recovery of the HSCs to reach the
quiescent state (LX-2 is an active cell line).
Of note, the phosphorylation level of ERK1/2
proteins was positively correlated with the changes in YB-1 levels
following OMT treatment. ERK1/2, members of the mitogen-activated
protein kinase family, regulate diverse cellular functions,
including proliferation, differentiation and cell cycle progression
(33,34). Studies have indicated that YB-1
activates the physiological features of certain types of cancer
cell via the ERK1/2 pathway (35–37);
however, its roles in fibrotic cells, such as HSCs, have remained
elusive.
In addition, ERK1/2 may have interacted with the
TGF-β signaling pathway as an alternative mechanism. The functions
of Smad3, the key protein in the TGF-β signaling pathway, were
shown to be ERK-dependent in certain cell types (38). Therefore, the present study
hypothesized that a link exists between the YB-1-mediated
regulation of the TGF-β signaling pathway and ERK1/2.
In the present study, the total protein levels of
YB-1 were decreased after OMT treatment, which was in parallel to
the phosphorylation of ERK1/2. A similar result was observed after
YB-1 knockdown in LX-2 cells. The appearance of only the pERK2 (44
KDa) band in the western blot may be due to the cell status during
transfection.
The molecular mechanisms of the anti-fibrotic
effects of OMT by facilitating the nuclear translocation of YB-1
are illustrated in Fig. 7 and can
be summarized as follows: Besides directly regulating the promoter
of COLα1(I) and COLα2(I) genes, YB-1 decreases the production of
pro-collagen I through inhibiting the TGF-β signaling pathway by
ERK1/2.
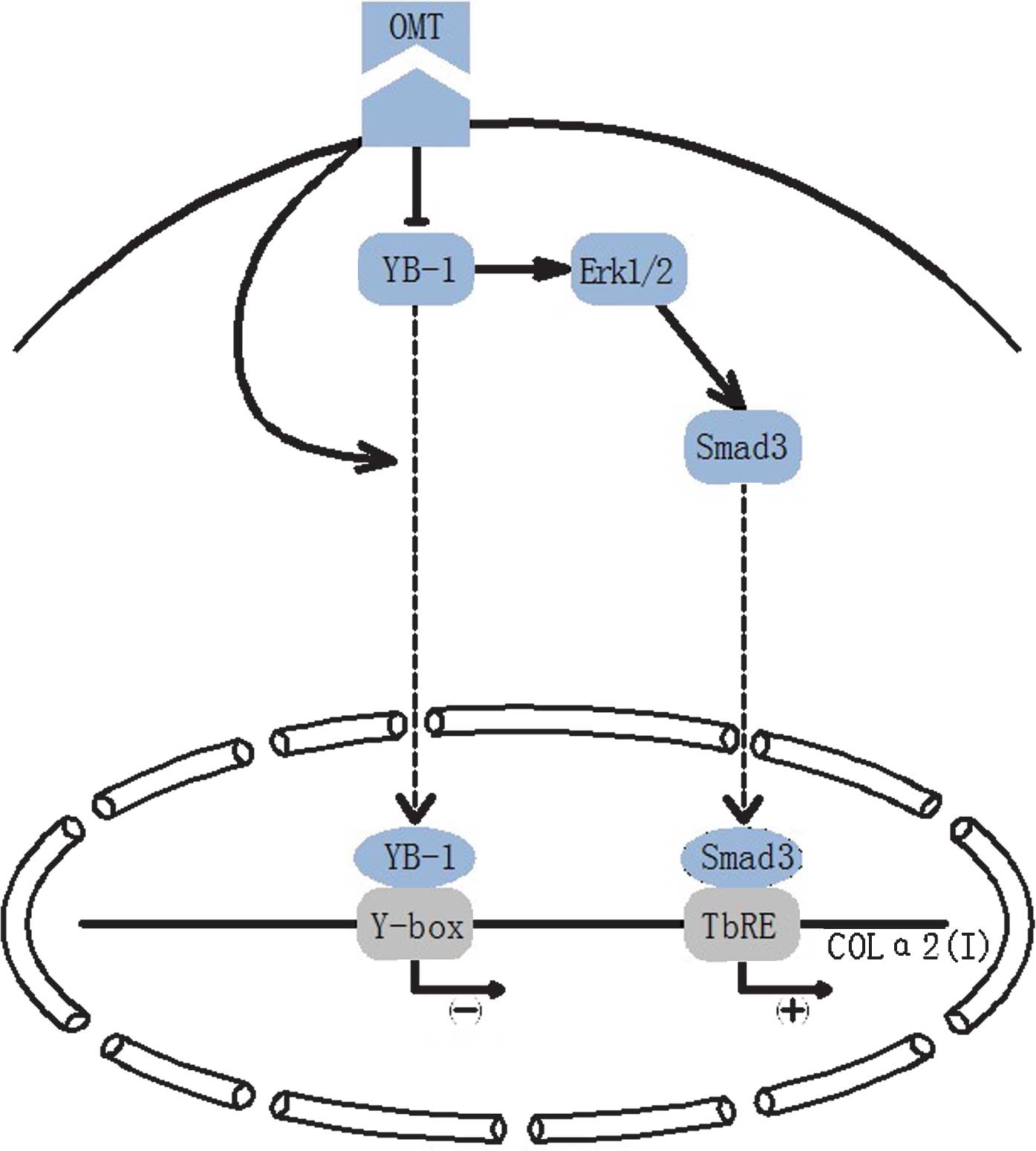 | Figure 7Attenuation of COLα2(I) expression by
OMT was, to a certain extent, mediated via nuclear translocation of
YB-1. In parallel, ERK1/2 was inhibited, which indirectly affected
the TGF-β signaling pathway. YB-1, Y-box binding protein 1; OMT,
oxymatrine; ERK, extracellular signal-regulated kinase; COLα1(I),
α1-strand of collagen I; TbRE, TGF-β-responsive element; TGF,
transforming growth factor. |
In conclusion, OMT attenuates pro-collagen I
expression, to a certain extent by increasing the levels of YB-1 in
the nucleus. Furthermore, OMT is capable of mediating the
inhibition of the TGF-β signaling pathway in the presence of
ERK1/2. Further study is required to completely elucidate the
mechanism of action of OMT against liver fibrosis.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81170405 and
81300327).
References
|
1
|
Krohn R, Raffetseder U, Bot I, Zernecke A,
Shagdarsuren E, Liehn EA, van Santbrink PJ, Nelson PJ, Biessen EA,
Mertens PR and Weber C: Y-box binding protein-1 controls CC
chemokine ligand-5 (CCL5) expression in smooth muscle cells and
contributes to neointima formation in atherosclerosis-prone mice.
Circulation. 116:1812–1820. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stratford AL, Habibi G, Astanehe A, Jiang
H, Hu K, Park E, Shadeo A, Buys TP, Lam W, Pugh T, et al: Epidermal
growth factor receptor (EGFR) is transcriptionally induced by the
Y-box binding protein-1 (YB-1) and can be inhibited with Iressa in
basal-like breast cancer, providing a potential target for therapy.
Breast Cancer Res. 9:R612007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harada M, Kotake Y, Ohhata T, Kitagawa K,
Niida H, Matsuura S, Funai K, Sugimura H, Suda T and Kitagawa M:
YB-1 promotes transcription of cyclin D1 in human non-small-cell
lung cancers. Genes Cells. 19:504–516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gaudreault I, Guay D and Lebel M: YB-1
promotes strand separation in vitro of duplex DNA containing either
mispaired bases or cisplatin modifications, exhibits
endonucleolytic activities and binds several DNA repair proteins.
Nucleic Acids Res. 32:316–327. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
En-Nia A, Yilmaz E, Klinge U, Lovett DH,
Stefanidis I and Mertens PR: Transcription factor YB-1 mediates DNA
polymerase alpha gene expression. J Biol Chem. 280:7702–7711. 2005.
View Article : Google Scholar
|
|
6
|
Stein U, Jürchott K, Walther W, Bergmann
S, Schlag PM and Royer HD: Hyperthermia-induced nuclear
translocation of transcription factor YB-1 leads to enhanced
expression of multidrug resistance-related ABC transporters. J Biol
Chem. 276:28562–28569. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sengupta S, Mantha AK, Mitra S and Bhakat
KK: Human AP endonuclease (APE1/Ref-1) and its acetylation regulate
YB-1-p300 recruitment and RNA polymerase II loading in the
drug-induced activation of multidrug resistance gene MDR1.
Oncogene. 30:482–493. 2011. View Article : Google Scholar
|
|
8
|
Didier DK, Schiffenbauer J, Woulfe SL,
Zacheis M and Schwartz BD: Characterization of the cDNA encoding a
protein binding to the major histocompatibility complex class II Y
box. Proc Natl Acad Sci USA. 85:7322–7326. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kuwano M, Oda Y, Izumi H, Yang SJ, Uchiumi
T, Iwamoto Y, Toi M, Fujii T, Yamana H, Kinoshita H, et al: The
role of nuclear Y-box binding protein 1 as a global marker in drug
resistance. Mol Cancer Ther. 3:1485–1492. 2004.PubMed/NCBI
|
|
10
|
Jurchott K, Bergmann S, Stein U, Walther
W, Janz M, Manni I, Piaggio G, Fietze E, Dietel M and Royer HD:
YB-1 as a cell cycle-regulated transcription factor facilitating
cyclin A and cyclin B1 gene expression. J Biol Chem.
278:27988–27996. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eliseeva IA, Kim ER, Guryanov SG,
Ovchinnikov LP and Lyabin DN: Y-box-binding protein 1 (YB-1) and
its functions. Biochemistry (Mosc). 76:1402–1433. 2011. View Article : Google Scholar
|
|
12
|
Ghiassi-Nejad Z, Hernandez-Gea V, Woodrell
C, Lang UE, Dumic K, Kwong A and Friedman SL: Reduced hepatic
stellate cell expression of Kruppel-like factor 6 tumor suppressor
isoforms amplifies fibrosis during acute and chronic rodent liver
injury. Hepatology. 57:786–796. 2013. View Article : Google Scholar
|
|
13
|
Sun W, Hou F, Panchenko MP and Smith BD: A
member of the Y-box protein family interacts with an upstream
element in the alpha1(I) collagen gene. Matrix Biol. 20:527–541.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Norman JT, Lindahl GE, Shakib K, En-Nia A,
Yilmaz E and Mertens PR: The Y-box binding protein YB-1 suppresses
collagen alpha 1(I) gene transcription via an evolutionarily
conserved regulatory element in the proximal promoter. J Biol Chem.
276:29880–29890. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Higashi K, Inagaki Y, Suzuki N, Mitsui S,
Mauviel A, Kaneko H and Nakatsuka I: Y-box-binding protein YB-1
mediates transcriptional repression of human alpha 2(I) collagen
gene expression by interferon-gamma. J Biol Chem. 278:5156–5162.
2003. View Article : Google Scholar
|
|
16
|
Inagaki Y, Higashiyama R and Higashi K:
Novel anti-fibrotic modalities for liver fibrosis: Molecular
targeting and regenerative medicine in fibrosis therapy. J
Gastroenterol Hepatol. 27(Suppl 2): S85–S88. 2012. View Article : Google Scholar
|
|
17
|
Homer C, Knight DA, Hananeia L, Sheard P,
Risk J, Lasham A, Royds JA and Braithwaite AW: Y-box factor YB1
controls p53 apoptotic function. Oncogene. 24:8314–8325. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koike K, Uchiumi T, Ohga T, Toh S, Wada M,
Kohno K and Kuwano M: Nuclear translocation of the Y-box binding
protein by ultraviolet irradiation. FEBS Lett. 417:390–394. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Higashi K, Tomigahara Y, Shiraki H, Miyata
K, Mikami T, Kimura T, Moro T, Inagaki Y and Kaneko H: A novel
small compound that promotes nuclear translocation of YB-1
ameliorates experimental hepatic fibrosis in mice. J Biol Chem.
286:4485–4492. 2011. View Article : Google Scholar :
|
|
20
|
Chen XS, Wang GJ, Cai X, Yu HY and Hu YP:
Inhibition of hepatitis B virus by oxymatrine in vivo. World J
Gastroenterol. 7:49–52. 2001.
|
|
21
|
Pei X, Wang W, Miao N, Xu M, Zhang C, Sun
M, Xu M and Liu Z: The protective effects of the combination of
sodium ferulate and oxymatrine on ethanol-induced liver damage in
mice. Environ Toxicol Pharmacol. 37:423–430. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang B, Niu W, Xu D, Li Y, Liu M, Wang Y,
Luo Y, Zhao P, Liu Y, Dong M, Sun R, et al: Oxymatrine prevents
hypoxia- and monocrotaline-induced pulmonary hypertension in rats.
Free Radic Biol Med. 69:198–207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang W, Zeng M, Fan Z, Mao Y, Song Y, Jia
Y, Lu L, Chen CW, Peng YS and Zhu HY: Prophylactic and therapeutic
effect of oxymatrine on D-galactosamine-induced rat liver fibrosis.
Chinese Journal of Hepatology. 10:193–196. 2002.In Chinese.
|
|
24
|
Zheng P, Niu FL, Liu WZ, Shi Y and Lu LG:
Anti-inflammatory mechanism of oxymatrine in dextran sulfate
sodium-induced colitis of rats. World J Gastroenterol.
11:4912–4915. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dong XQ, Du Q, Yu WH, Zhang ZY, Zhu Q, Che
ZH, Chen F, Wang H and Chen J: Anti-inflammatory effects of
oxymatrine through inhibition of nuclear factor-kappa B and
mitogen-activated protein kinase activation in
lipopolysac-charide-induced BV2 microglia cells. Iran J Pharm Res.
12:165–174. 2013.PubMed/NCBI
|
|
26
|
Zhang J, Wang M, Zhang Z, Luo Z, Liu F and
Liu J: Celecoxib derivative OSU-03012 inhibits the proliferation
and activation of hepatic stellate cells by inducing cell
senescence. Mol Med Rep. 11:3021–3026. 2015.
|
|
27
|
Chemin I and Zoulim F: Hepatitis B virus
induced hepatocellular carcinoma. Cancer Lett. 286:52–59. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tacke F, Kanig N, En-Nia A, Kaehne T,
Eberhardt CS, Shpacovitch V, Trautwein C and Mertens PR: Y-box
protein-1/p18 fragment identifies malignancies in patients with
chronic liver disease. BMC Cancer. 11:1852011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yasen M, Kajino K, Kano S, Tobita H,
Yamamoto J, Uchiumi T, Kon S, Maeda M, Obulhasim G, Arii S and Hino
O: The up-regulation of Y-box binding proteins (DNA binding protein
A and Y-box binding protein-1) as prognostic markers of
hepatocellular carcinoma. Clin Cancer Res. 11:7354–7361. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chai NL, Fu Q, Shi H, Cai CH, Wan J, Xu SP
and Wu BY: Oxymatrine liposome attenuates hepatic fibrosis via
targeting hepatic stellate cells. World J Gastroenterol.
18:4199–4206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen X, Sun R, Hu J, Mo Z, Yang Z, Liao D
and Zhong N: Attenuation of bleomycin-induced lung fibrosis by
oxymatrine is associated with regulation of fibroblast
proliferation and collagen production in primary culture. Basic
Clin Pharmacol Toxicol. 103:278–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang XY and Chen CX: Effect of
oxymatrine, the active component from Radix Sophorae flavescentis
(Kushen), on ventricular remodeling in spontaneously hypertensive
rats. Phytomedicine. 20:202–212. 2013. View Article : Google Scholar
|
|
33
|
Raman M, Chen W and Cobb MH: Differential
regulation and properties of MAPKs. Oncogene. 26:3100–3112. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoon S and Seger R: The extracellular
signal-regulated kinase: Multiple substrates regulate diverse
cellular functions. Growth Factors. 24:21–44. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Imada K, Shiota M, Kohashi K, Kuroiwa K,
Song Y, Sugimoto M, Naito S and Oda Y: Mutual regulation between
Raf/MEK/ERK signaling and Y-box-binding protein-1 promotes prostate
cancer progression. Clin Cancer Res. 19:4638–4650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee C, Dhillon J, Wang MY, Gao Y, Hu K,
Park E, Astanehe A, Hung MC, Eirew P, Eaves CJ and Dunn SE:
Targeting YB-1 in HER-2 overexpressing breast cancer cells induces
apoptosis via the mTOR/STAT3 pathway and suppresses tumor growth in
mice. Cancer Res. 68:8661–8666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao BX, Sun YB, Wang SQ, Duan L, Huo QL,
Ren F and Li GF: Grape seed procyanidin reversal of p-glycoprotein
associated multi-drug resistance via down-regulation of nf-κb and
MAPK/ERK mediated YB-1 activity in A2780/T cells. PLoS One.
8:e710712013. View Article : Google Scholar
|
|
38
|
Hayashida T, Decaestecker M and Schnaper
HW: Cross-talk between ERK MAP kinase and Smad signaling pathways
enhances TGF-beta-dependent responses in human mesangial cells.
FASEB J. 17:1576–1578. 2003.PubMed/NCBI
|