Introduction
Endometriosis is a common gynecological disease
affecting 6–10% of women of reproductive age (1). It is characterized by the presence of
endometrial glands and/or stroma outside of the uterine cavity,
including the ovaries, ligaments of the uterus, fallopian tubes,
cervical vaginal area, peritoneum, umbilicus and urinary tract
(2,3). The primary symptoms of endometriosis
are pelvic pain and infertility, although other symptoms, including
ovarian masses, dysmenorrhea, dyspareunia, irregular uterine
bleeding and dysuria, are also common (1,4).
Various hypotheses have been postulated to explain
the pathogenesis of endometriosis, including stem cell-,
metaplasia- and implantation-based theories (3,5,6).
However, the implantation theory is the most accepted one, which
proposes that ectopic endometrioid lesions develop via the
implantation of endometrial glands and/or stroma that are
retrogradely transported into the pelvic cavity through menstrual
blood. That is, the pathogenesis of endometriosis involves the
adhesion, proliferation and invasion of endometrial cells and
angiogenesis (7). Cytokines,
angiogenic factors and adhesion promotion factors, including
interleukin (IL)-6, IL-8, tumor growth factor-β and vascular
endothelial growth factor, contribute to cell attachment,
proliferation and invasion, and neovascularization (8,9). It
is also considered that various factors, including inflammatory,
genetic, epigenetic, hormonal, immune, anatomic and lifestyle
factors, are associated with the etiology of endometriosis
(6,10,11).
However, despite the various hypotheses, the exact pathogenesis of
endometriosis remains unclear.
Endometriosis is a chronic disease with a high risk
of recurrence and histological analysis is the only definitive
method of diagnosis (12,13). Hormone therapy, medication and
surgery can only alleviate the symptoms (14). As efficient diagnostic methods and
treatments are lacking, patients with endometriosis typically
experience severe symptoms and high medical expenses, which
considerably influence their quality of life (12,13,15).
Thus, it is important to explore the pathological mechanisms and
identify more efficient biomarkers for the diagnosis and drug
treatment of endometriosis.
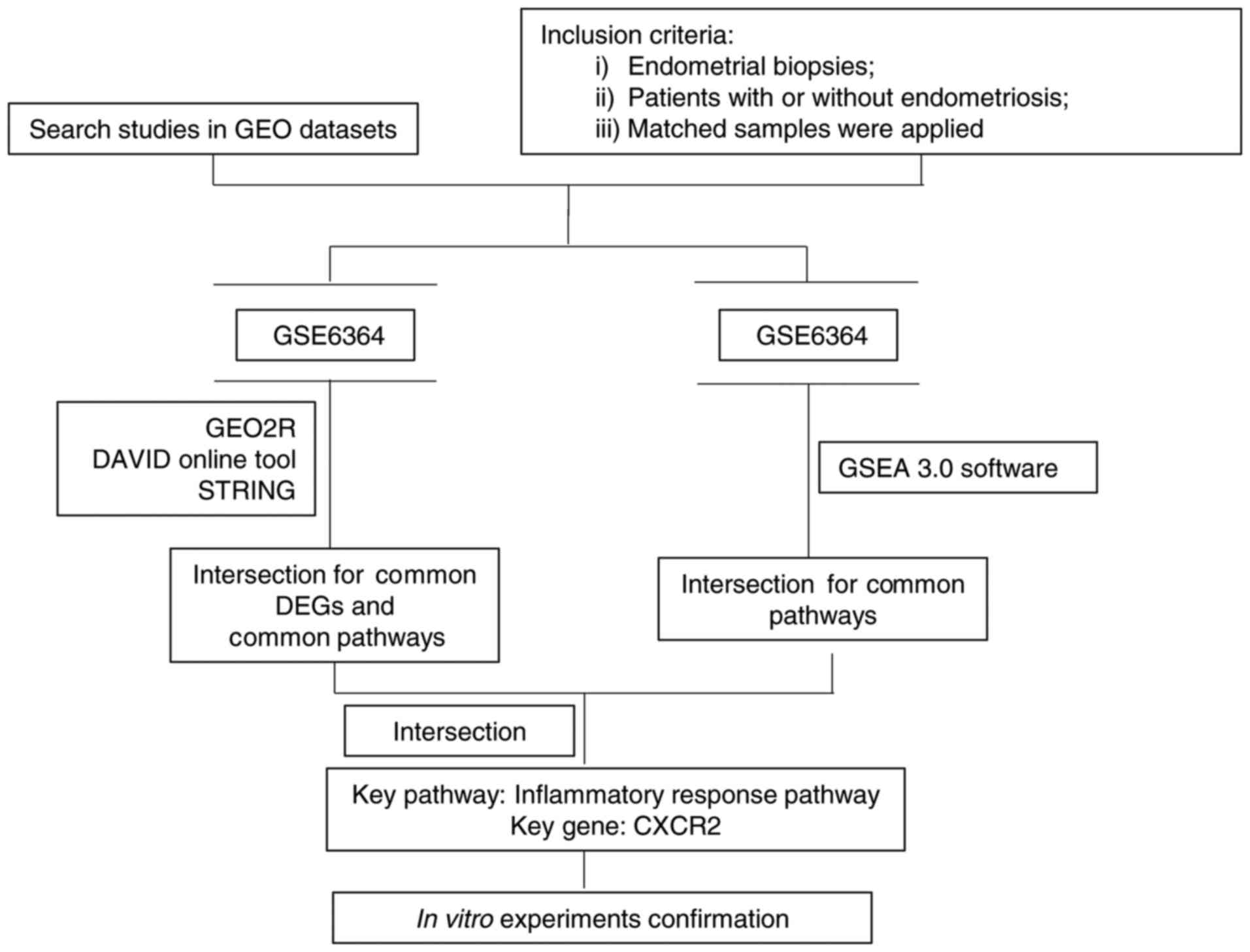
In the present study, a bioinformatics approach was
applied to systematically identify the pathways involved in
endometriosis. Gene expression profiling analysis is a useful
method to distinguish differentially expressed genes (DEGs) in a
particular condition (16,17). Based on Sampson's implantation
theory that ectopic endometrial tissues contribute to the
pathogenesis of endometriosis (18), the gene expression profiles from
endometrial biopsies from women with or without endometriosis were
investigated using the Gene Expression Omnibus (GEO) database.
Through analysis of the biological functions and pathways of the
identified genes, the present study aimed to identify critical
pathways and genes contributing to endometriosis, and potential
biomarkers for its diagnosis, prognosis and therapy.
Materials and methods
Bioinformatics approach
Microarray data
The GSE6364 dataset was downloaded from the GEO
DataSets database (www.ncbi.nlm.nih.gov/gds). GSE6364 contained the
processed and normalized gene expression profiles of endometrial
biopsies from women with normal endometrial pathology and no
history of endometriosis, and from women with laparoscopy-proven
moderate-to-severe stage endometriosis from an Affymetrix GPL570
platform (Affymetrix Human Genome U133 Plus 2.0 Array; Affymetrix;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) (19). In the present study, proliferative
phases (day 8–14 in regular menstruation) samples were selected,
including six samples (GSM150190-150195) from patients with
endometriosis and five normal samples (GSM150196-150201).
Identification of DEGs
DEG analysis of the dataset was performed using
GEO2R (www.ncbi.nlm.nih.gov/geo/geo2r/), which performed
comparisons using the GEO query and limma R packages of the
Bioconductor project (20).
P-values of the DEGs were determined using Student's t-test. An
adjusted P<0.05 and a fold-change in expression ≥2 were set as
thresholds to identify statistically significant DEGs. Hierarchical
clustering analysis was applied to categorize the data, and a
heatmap was produced with HemI 1.0 software (21).
Gene ontology (GO) and pathway
enrichment analyses of DEGs
GO analysis is commonly used for gene annotation,
including the molecular function (MF), biological process (BP) and
cellular component (CC) categories (22). The DAVID online tool (david.ncifcrf.gov/) was applied to analyze the
functional level, including GO enrichment of the DEGs (23). Gene Set Enrichment Analysis (GSEA)
was conducted using the software GSEA v3.0 (www.broadinstitute.org/gsea/) (24,25).
Construction of a protein-protein
interaction (PPI) network
The Search Tool for the Retrieval of Interacting
Genes (STRING version 10.5; string-db.org/) database is an online tool used for
the evaluation of the PPIs (26).
To identify interactions among the DEGs, STRING was applied to map
the DEGs. Experimentally validated interactions were included,
whereas single nodes without interactions were excluded. PPI
networks visualization was achieved with the Cytoscape software
(version 3.5.1) (27).
Validation of key genes
Ethics statement
Endometrial tissue in the present study was
collected from patients from September to November 2017 in Renmin
Hospital of Wuhan University (Renmin, China) and they provided
written informed consent. The present study was approved by the
Ethics Committee of Renmin Hospital of Wuhan University in
September 2017.
Cell isolation and culture
Primary endometrial stromal cells (HESCs) were
isolated as described previously (28). Patients, ranging from 24–45-years
old, had regular menstrual cycles, and were documented not to be
pregnant at the time of surgery were enrolled in the present study;
however, patients using any form of hormonal treatment within 3
months of biopsy were excluded. Briefly, endometrial tissue from 8
patients with endometriosis was finely minced and the cells
dispersed by incubation in Hank's balanced salt solution (25
mmol/ml) containing collagenase (1 mg/ml, 15 U/mg), 1%
penicillin/streptomycin and deoxyribonuclease (0.1 mg/ml, 1,500
U/mg) at 37°C for 60 min in a water bath. Following filtration
through a 40-µm cell strainer (Falcon®; Corning
Incorporated, Corning, NY, USA), the cells were seeded in 75
cm2 Falcon tissue culture flasks (BD Biosciences, San
Jose, CA, USA) and suspended in Ham's F12:Dulbecco's modified eagle
medium (1:1) containing 10% fetal bovine serum (FBS; HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) and antibiotics (100
IU/ml penicillin, 100 µg/ml streptomycin and 0.25 µg/ml
amphotericin B) in a 37°C incubator with 5% CO2. The
purity of HESCs were determined by vimentin immunohistochemical
staining. Briefly, following 10% formalin fixation in room
temperature for 30 min, cells were immersed in 3% hydrogen peroxide
solution for 10 min at room temperature. Then, incubation with an
anti-vimentin primary antibody (1:400; ab8978) at 37°C for 1 h was
conducted and followed by a second antibody goat anti-mouse Alexa
Fluor® 488 IgG (1:3,000; ab150117; both Abcam,
Cambridge, MA, USA) at 37°C for 20 min. Subsequently, staining with
diaminobenzidine (GK6007, Gene Tech Co., Ltd., Shanghai, China)
according to manufacturer recommendations and then counterstained
with hematoxylin at room temperature for 10 min, followed by
analysis with an inverted light microscope (magnification, ×400,
Nikon Corporation, Tokyo, Japan). Cultured HESCs were used for
further analysis following 3–5 passages.
Cell treatments
Overexpression of C-X-C motif chemokine receptor 2
(CXCR2) was achieved using a pLKO lentiviral vector targeting CXCR2
constructed by Thermo Fisher Scientific, Inc. Lentivirus stocks
were obtained using the ViraPower™ Lentiviral Packing Mix and 293FT
cell line according to the manufacturer's protocol (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). HESCs at 50%
confluency were cultured in a 1:1 dilution of virus:media with 5
µg/ml polybrene for 24 h at 37°C, followed by selection of stable
cell lines. The knockdown of CXCR2 in HESCs was achieved using
small interfering RNA (siRNA). Cells were transfected with
CXCR2-siRNA at a final concentration of 50 nM using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Cells
transfected with a final concentration of 50 nM scramble sequences
were the control group. Sequences of the CXCR2 primers and
CXCR2-siRNAs used are demonstrated in Table I. Following 24 h, cells were
collected for the subsequent analyses.
 | Table I.siRNA sequences and primers. |
Table I.
siRNA sequences and primers.
| Name | Sense or forward
sequences (5′-3′) | Anti-sense or
reverse sequences (5′-3′) |
|---|
| CXCR2 siRNA |
AGCGACCCAGUCAGGAUUUTT |
AAAUCCUGACUGGGUCGCUTT |
| CXCR2 siRNA
scramble |
AGCAGCUCAAUGCGUCAGUTT |
ACUGACGCAUUGAGCUGCUTT |
| CXCR2 primers |
AGCTGAGAATATGCAGCCGTT |
CATAGCAGGCTGGGCTAACA |
| GAPDH primers |
TTGATGGCAACAATCTCCAC |
CGTCCCGTAGACAAAATGGT |
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted with TRIzol reagent (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The PrimeScript™ RT Reagent kit was purchased from Takara
Biotechnology Co., Ltd. (Dalian, China). Details of the RNA
isolation, cDNA conversion and RT-qPCR assays have been described
previously (29). To examine the
mRNA expression levels of CXCR2, RT-qPCR was conducted in 96-well
reaction plates using the ABI Step One Plus™ real-time PCR System
(Thermo Fisher Scientific, Inc.). Each well contained 1 µl cDNA
template, 0.2 µl each primer, 3.6 µl diethyl
pyrocarbonate-H2O and 5 µl SYBR Green dye. CXCR2 and
GAPDH primers were designed and synthesized by Sangon Biotech Co.,
Ltd. (Shanghai, China). The qPCR cycling conditions were as
follows: Pre-denaturation at 95°C for 30 sec, followed by 40 cycles
of denaturation at 95°C for 5 sec, annealing at 60°C for 10 sec and
extension at 72°C for 30 sec. GAPDH was used to normalize the
relative mRNA expression of CXCR2. The 2−ΔΔCq method was
applied to calculate the relative expression level of the target
amplicon (30).
Cell proliferation and Transwell
assays
For the cell proliferation assay, transfected HESCs
were suspended in Ham's F12/DMEM for 3 days in 96-well plates
(5×104 cells/well). A Cell Counting kit-8 (CCK-8) assay
was subsequently conducted. CCK-8 (10 µl; WST-8, Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was mixed with the cells,
followed by incubation at 37°C for 2 h. The absorbance at 450 nm
was measured using an ELISA reader (Tecan Group, Ltd., Mannedorf,
Switzerland) to detect cell viability. For the cell migration and
invasion assays, Transwell permeable supports (Corning
Incorporated) and Matrigel (BD Biosciences) were used. Transfected
cells were cultured in 200 µl in Ham's F12:Dulbecco's modified
eagle medium prior to being transferred onto the upper chambers of
24-well plates (5×105 cells/well) with or without a
Matrigel coating. A total of 800 µl medium with 10% FBS was added
to the lower chamber. Following 24 h incubation, cells in the lower
chamber were fixed with absolute methanol and stained with 0.1%
crystal violet solution (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 10 min at room temperature. Cell counting was
performed at medium magnification (×100) under an inverted light
microscope from thee randomly selected fields. For each condition,
three independent experiments were conducted. Image analysis was
performed using Image J software version 1.49 (National Institutes
of Health, Bethesda, MD, USA).
Statistical analysis
Statistical analysis was performed using the Stata
13.0 software (StataCorp LP, College Station, TX, USA). One-way
analysis of variance and the Scheffe post hoc test were applied.
Data are presented as the mean ± standard error. P<0.05 was
considered to indicate a statistically significant difference.
Results
Overview of the GEO microarray data
and identification of DEGs
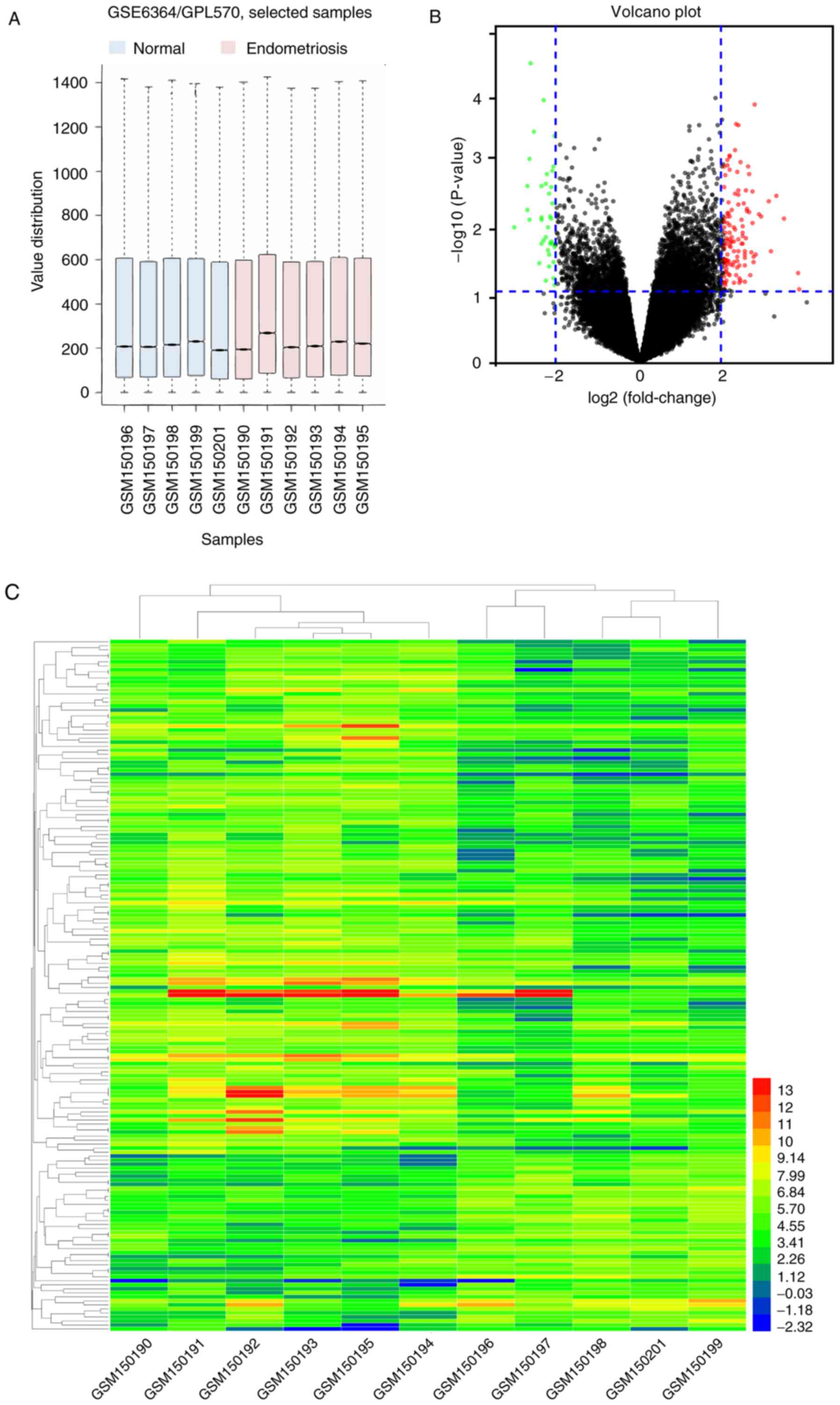
A box plot graph of the value distribution in the
GSE6364 dataset was generated to assess whether the distributions
of values across samples were median-centered and of conformity.
The results indicated that the data of GSE6364 were generally
normalized and cross-comparable (Fig.
1A). An adjusted P<0.05 and a fold-change ≥2 were set to
filter for statistically significant DEGs. A total of 172 genes
were identified, as demonstrated in the volcano plot (Fig. 1B) and a hierarchical cluster
analysis was performed on the DEGs, as indicated in the heat map
(Fig. 1C).
Pathway enrichment analysis
The results of the GO analysis of the DEGs indicated
that ‘inflammatory response’, ‘innate immune response’ and
‘chemokine production’ were the most significant terms in the BP
category. ‘Extracellular space’, ‘extracellular exosome’ and
‘membrane’ were the most significant in CC. The significant MF
terms were ‘Toll-like receptor 4 binding’, ‘arachidonic acid
binding’, ‘signaling pattern recognition receptor activity’ and
‘CXCR chemokine receptor binding’ (Table II).
| Table II.GO analysis of the differentially
expressed genes (P<0.05). |
Table II.
GO analysis of the differentially
expressed genes (P<0.05).
| A,
GOTERM_BP_DIRECT |
|
| Term | Count | P-value | Fold
enrichment |
|---|
| GO:0006954
inflammatory response | 9 |
3.70×10−4 | 5.05 |
| GO:0045087 innate
immune response | 9 |
8.48×10−4 | 4.45 |
| GO:0002523
leukocyte migration involved in inflammatory response | 3 |
1.14×10−3 | 57.97 |
| GO:0030593
neutrophil chemotaxis | 4 |
3.58×10−3 | 12.88 |
| GO:0042742 defense
response to bacterium | 5 |
4.64×10−3 | 7.33 |
| GO:0002227 innate
immune response in mucosa | 3 |
5.96×10−3 | 25.51 |
| GO:0006910
phagocytosis, recognition | 3 |
7.45×10−3 | 22.77 |
| GO:0070488
neutrophil aggregation | 2 |
9.27×10−3 | 212.56 |
| GO:0016337 single
organismal cell-cell adhesion | 4 |
1.16×10−2 | 8.42 |
| GO:0032602
chemokine production | 2 |
1.39×10−2 | 141.70 |
| GO:0019731
antibacterial humoral response | 3 |
1.78×10−2 | 14.49 |
| GO:0032119
sequestering of zinc ion | 2 |
1.85×10−2 | 106.28 |
| GO:0002793 positive
regulation of peptide secretion | 2 |
1.85×10−2 | 106.28 |
| GO:0032870 cellular
response to hormone stimulus | 3 |
1.85×10−2 | 14.17 |
| GO:0007267
cell-cell signaling | 5 |
3.05×10−2 | 4.18 |
| GO:0006260 DNA
replication | 4 |
3.55×10−2 | 5.49 |
| GO:0002221 pattern
recognition receptor signaling pathway | 2 |
3.66×10−2 | 53.14 |
| GO:0032496 response
to lipopolysaccharide | 4 |
4.09×10−2 | 5.18 |
| GO:0002544 chronic
inflammatory response | 2 |
4.10×10−2 | 47.23 |
| GO:0070098
chemokine-mediated signaling pathway | 3 |
4.31×10−2 | 8.98 |
| GO:0006955 immune
response | 6 |
4.61×10−2 | 3.03 |
|
| B,
GOTERM_CC_DIRECT |
|
| Term | Count | P-value | Fold
enrichment |
|
| GO:0005615
extracellular space | 20 |
1.30×10−5 | 3.11 |
| GO:0005576
extracellular region | 20 |
1.48×10−4 | 2.60 |
| GO:0070062
extracellular exosome | 23 |
9.88×10−3 | 1.71 |
| GO:0005886 plasma
membrane | 29 |
2.18×10−2 | 1.47 |
| GO:0016020
membrane | 18 |
2.68×10−2 | 1.71 |
| GO:0009986 cell
surface | 7 |
4.30×10−2 | 2.71 |
|
| C,
GOTERM_MF_DIRECT |
|
| Term | Count | P-value | Fold
enrichment |
|
| GO:0035662
Toll-like receptor 4 binding | 2 |
1.86×10−2 | 105.51 |
| GO:0050544
arachidonic acid binding | 2 |
2.32×10−2 | 84.41 |
| GO:0008329
signaling pattern recognition receptor activity | 2 |
3.23×10−2 | 60.29 |
| GO:0045236 CXCR
chemokine receptor binding | 2 |
4.13×10−2 | 46.89 |
Key pathways and genes
identification
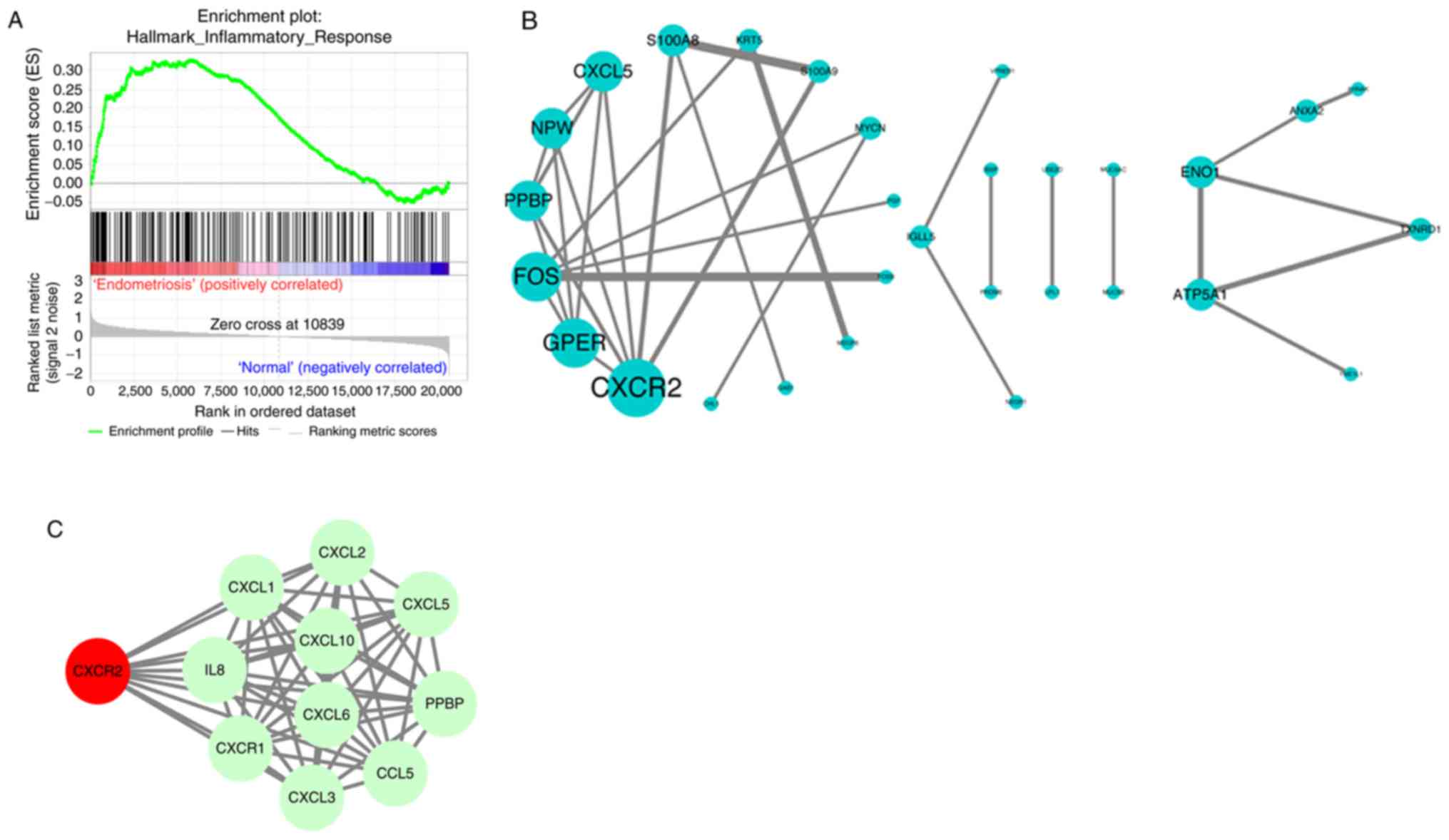
To further identify the key pathways, GSEA was
conducted on the GSE6364 dataset. ‘INFLAMMATORY RESPONSE’ was
identified, in accordance with the results of GO analysis. The
enrichment score for this signaling pathway was positively
correlated to endometriosis samples, while the enrichment score was
lower in normal samples (Fig. 2A).
Subsequently, the PPI among the DEGs were evaluated by STRING and
PPI network visualization was performed using Cytoscape. CXCR2, G
protein-coupled estrogen receptor 1, Fos proto-oncogene, C-X-C
motif chemokine ligand 5 and others were preserved according to the
inclusion criterion (Fig. 2B). As
the most significant DEG, CXCR2 was uploaded into STRING to search
its potential PPI with possible proteins. All the interacting
proteins were involved in the inflammatory response, which was
consistent with the other findings (Fig. 2C). Therefore, CXCR2 was considered
the most important identified gene.
Validation of the function of CXCR2 in
HESCs
To validate the critical role of CXCR2 in the
development of endometriosis, HESCs were used. Overexpression and
knockdown of CXCR2 in cells were achieved via transfection with
pLKO lentiviral vectors and siRNA, respectively. Cells with
scramble sequences were set as the control group. RT-qPCR was
applied to confirm the efficacy of transfection (Fig. 3A). In the analysis of cell
proliferation by CCK-8 assay, the overexpression of CXCR2 clearly
increased the proliferation capacity of HESCs, while the capacity
markedly decreased following the knockdown of CXCR2 (Fig. 3B). For the analysis of cell
migration and invasion, Transwell assays were performed. As
demonstrated in Fig. 3C, the
migration and invasion capacities of HESCs were significantly
promoted in the pLKO- CXCR2 group, but clearly suppressed in the
si-CXCR2 group, compared with the control.
Discussion
Endometriosis is characterized by the morphological
and biological properties of the endometrium, including the
proliferation, invasiveness, and attachment to the extracellular
matrix, which are greater in ectopic endometrial cells (3), and results in a severe decline in the
quality of life and psychological well-being, as well as it is a
heavy economic burden, for those affected (10,31).
Although various potential causative factors have been
investigated, the exact mechanisms underlying the aberrant
implantation of ectopic endometrial cells remain unclear. In the
present study, a bioinformatics approach was applied to reveal the
possible dysregulated pathways and genes in endometriosis,
specifically through gene expression analysis (Fig. 4). The gene expression profiles of
endometrial biopsies from women with or without endometriosis
(GSE6364) were selected. A total of 172 DEGs were extracted. Gene
expression profiling analysis through functional annotation and
gene set enrichment analysis as well as a PPI network, demonstrated
that genes associated with the inflammatory response were
upregulated, and CXCR2 was one of the most important genes.
Furthermore, the in vitro experiments confirmed that CXCR2
promoted proliferation, migration and invasion of HESCs, suggesting
that CXCR2 may serve a critical role in the development of
endometriosis.
According to Sampson's implantation theory that
ectopic endometrial tissues contribute to the pathogenesis of
endometriosis (18), the present
study compared the gene expression profiles of endometriosis
patients with that of normal subjects. The present results
demonstrated that inflammatory response genes were significantly
upregulated. Previous studies have indicated that inflammation
serves a key role in the pathogenesis of endometriosis (32,33).
Alterations in inflammatory factors may promote fibrotic adhesion
contributing to the growth and spread of endometrial tissue
(34). It has been reported that
the peritoneal fluid of patients with endometriosis exhibits an
increased level of activated macrophages and pro-inflammatory
cytokines compared with that in normal individuals, and that the
capacities for proliferation, invasion and adhesion to the
extracellular matrix are greater in ectopic endometrial cells than
in eutopic cells (3,35). The dynamic interactions between
cytokines may serve a pivotal role in the formation of a
microenvironment that favors the ectopic implantation of
endometrial tissues and the progression of endometriosis (36). The presence and role of cytokines,
such as IL-8, have been extensively investigated (37). IL-8 contributes to reproductive
pathological processes, such as endometriosis, through affecting
the proliferation and invasion of endometrial stroma cells (ESCs)
in the ectopic endometrial tissue (36,38–40).
CXCR2, a membrane receptor for IL-8, is a member of the
G-protein-coupled receptor family, and is involved in influencing
intracellular calcium concentration, the release of granular
enzymes and chemotaxis in response to IL-8 (36). Through the high-affinity binding of
IL8 to CXCR2, signals are transduced via a G protein-activated
second messenger system to promote the proliferation and invasion
of ESCs. The present results also demonstrated that CXCR2 promoted
the proliferation of HESCs. This may have been the result of CXCR2
high affinity for growth regulated protein-α, which has previously
been demonstrated to affect the proliferative capacity of
endometriosis cells (41,42). However, the contribution of
decreased proliferation to reductions in migration and invasion
cannot be excluded as migration/invasion of cells is generally
affected by the number of cells, which may pose as a limitation of
the present study. In addition, the expression of CXCR2 is very
weak in normal endometrium and detection of CXCR2 in endometrial
tissues through immunohistochemistry demonstrate that the
expression is higher in women with endometriosis than in those with
normal endometrium (36,42), which is consistent with the present
DEG analysis.
The diagnosis of endometriosis may be achieved by
detecting certain factors circulating in the serum or in the
peritoneal fluid of patients. For example, the expression of
soluble class-I and class-II molecules has been demonstrated to be
significantly reduced by >30% in patients with endometriosis
(43,44). The production of cytokines, which
are thought to regulate the processes involved in the progression
and development of endometriosis, largely depends on the immune
system status of each patient with endometriosis, detection of a
combination of these factors may allow endometriosis diagnosis more
rapidly than the surgical route. IL-8 is increased in the serum of
patients with endometriosis (45).
Thus, CXCR2 could serve as a novel biomarker for the diagnosis of
endometriosis; however, further studies are required to demonstrate
this.
To conclude, the application of bioinformatic
methods to reveal important biomarkers in endometriosis was of
importance of the present study. Overexpression of CXCR2 promotes
the proliferation, migration and invasion of endometrial cells,
which in situ may lead to the presence of endometrial glands
and/or stroma outside of the uterine cavity. CXCR2 may be
associated with the development of endometriosis via the
inflammatory response and it has potential as a biomarker for the
diagnosis of endometriosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Independent
Scientific Research Program of Wuhan University (grant no.
2042018kf0170).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AT conducted the experiments and wrote the
manuscript. RL conducted software-based analysis and modified the
English language of the manuscript. HL and ML collected the patient
data. PR made substantial contributions to the design of the
present study. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Endometrial tissue in the present study was
collected from patients from September to November 2017 and written
informed consent was provided. The present study was approved by
the Ethics Committee of Renmin Hospital of Wuhan University
(Renmin, China) in September 2017.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Eskenazi B and Warner ML: Epidemiology of
endometriosis. Obstet Gynecol Clin North Am. 24:235–258. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vetvicka V, Lagana AS, Salmeri FM, Triolo
O, Palmara VI, Vitale SG, Sofo V and Králíčková M: Regulation of
apoptotic pathways during endometriosis: From the molecular basis
to the future perspectives. Arch Gynecol Obstet. 294:897–904. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Laganà AS, Vitale SG, Salmeri FM, Triolo
O, Frangež Ban H, Vrtačnik-Bokal E, Stojanovska L, Apostolopoulos
V, Granese R and Sofo V: Unus pro omnibus, omnes pro uno: A novel,
evidence-based, unifying theory for the pathogenesis of
endometriosis. Med Hypotheses. 103:10–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Louis Buck GM, Hediger ML, Peterson CM,
Croughan M, Sundaram R, Stanford J, Chen Z, Fujimoto VY, Varner MW,
Trumble A, et al: Incidence of endometriosis by study population
and diagnostic method: The ENDO study. Fertil Steril. 96:360–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sourial S, Tempest N and Hapangama DK:
Theories on the pathogenesis of endometriosis. Int J Reprod Med.
2014:1795152014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Laganà AS, Salmeri FM, Vitale SG, Triolo O
and Götte M: Stem cell trafficking during endometriosis: May
epigenetics play a pivotal role? Reprod Sci. 25:978–979. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta S, Agarwal A, Krajcir N and Alvarez
JG: Role of oxidative stress in endometriosis. Reprod Biomed
Online. 13:126–134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gazvani R and Templeton A: Peritoneal
environment, cytokines and angiogenesis in the pathophysiology of
endometriosis. Reproduction. 123:217–226. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Van Langendonckt A, Casanas-Roux F and
Donnez J: Oxidative stress and peritoneal endometriosis. Fertil
Steril. 77:861–870. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arablou T and Kolahdouz-Mohammadi R:
Curcumin and endometriosis: Review on potential roles and molecular
mechanisms. Biomed Pharmacother. 97:91–97. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mohammadi Kolahdouz R and Arablou T:
Resveratrol and endometriosis: In vitro and animal studies and
underlying mechanisms (Review). Biomed Pharmacother. 91:220–228.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Laganà AS, La Rosa VL, Rapisarda AMC,
Valenti G, Sapia F, Chiofalo B, Rossetti D, Frangež Ban H, Bokal
Vrtačnik E and Vitale SG: Anxiety and depression in patients with
endometriosis: Impact and management challenges. Int J Womens
Health. 9:323–330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Laganà AS, La Rosa V, Petrosino B and
Vitale SG: Comment on ‘Risk of developing major depression and
anxiety disorders among women with endometriosis: A longitudinal
follow-up study’. J Affect Disord. 208:672–673. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Laganà AS, Vitale SG, Granese R, Palmara
V, Frangež Ban H, Vrtačnik-Bokal E, Chiofalo B and Triolo O:
Clinical dynamics of Dienogest for the treatment of endometriosis:
From bench to bedside. Expert Opin Drug Metab Toxicol. 13:593–596.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Simoens S, Dunselman G, Dirksen C,
Hummelshoj L, Bokor A, Brandes I, Brodszky V, Canis M, Colombo GL,
DeLeire T, et al: The burden of endometriosis: Costs and quality of
life of women with endometriosis and treated in referral centres.
Hum Reprod. 27:1292–1299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo Y, Wu Y, Peng Y, Liu X, Bie J and Li
S: Systematic analysis to identify a key role of CDK1 in mediating
gene interaction networks in cervical cancer development. Ir J Med
Sci. 185:231–239. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang C, Huang Y, Pei Y, Zhang HH, Chen X,
Guo H, Li S, Ji X and Hu J: Genome-wide gene expression profiling
reveals that CD274 is up-regulated new-onset type 1 diabetes
mellitus. Acta Diabetol. 54:757–767. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Benagiano G, Brosens I and Lippi D: The
history of endometriosis. Gynecol Obstet Invest. 78:1–9. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Burney RO, Talbi S, Hamilton AE, Vo KC,
Nyegaard M, Nezhat CR, Lessey BA and Giudice LC: Gene expression
analysis of endometrium reveals progesterone resistance and
candidate susceptibility genes in women with endometriosis.
Endocrinology. 148:3814–3826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sean D and Meltzer PS: GEOquery: A bridge
between the gene expression omnibus (GEO) and BioConductor.
Bioinformatics. 23:1846–1847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deng W, Wang Y, Liu Z, Cheng H and Xue Y:
HemI: A toolkit for illustrating heatmaps. PLoS One. 9:e1119882014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mootha VK, Lindgren CM, Eriksson KF,
Subramanian A, Sihag S, Lehar J, Puigserver P, Carlsson E,
Ridderstråle M, Laurila E, et al: PGC-1alpha-responsive genes
involved in oxidative phosphorylation are coordinately
downregulated in human diabetes. Nat Genet. 34:267–273. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39:(Database Issue). D561–D568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cho S, Mutlu L, Zhou Y and Taylor HS:
Aromatase inhibitor regulates let-7 expression and let-7f-induced
cell migration in endometrial cells from women with endometriosis.
Fertil Steril. 106:673–680. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Di Stefano V, Wang B, Parobchak N, Roche N
and Rosen T: RelB/p52-mediated NF-κB signaling alters histone
acetylation to increase the abundance of corticotropin-releasing
hormone in human placenta. Sci Signal. 8:ra852015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Simoens S, Hummelshoj L and D'Hooghe T:
Endometriosis: Cost estimates and methodological perspective. Hum
Reprod Update. 13:395–404. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mu F, Harris HR, Rich-Edwards JW,
Hankinson SE, Rimm EB, Spiegelman D and Missmer SA: A prospective
study of inflammatory markers and risk of endometriosis. Am J
Epidemiol. 187:515–522. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bulun SE: Endometriosis. N Engl J Med.
360:268–279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Allen C, Hopewell S and Prentice A:
Non-steroidal anti-inflammatory drugs for pain in women with
endometriosis. Cochrane Database Syst Rev: CD004753. 2005.
View Article : Google Scholar
|
|
35
|
Burney RO and Giudice LC: Pathogenesis and
pathophysiology of endometriosis. Fertil Steril. 98:511–519. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ulukus M, Ulukus EC, Seval Y, Zheng W and
Arici A: Expression of interleukin-8 receptors in endometriosis.
Hum Reprod. 20:794–801. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nishida M, Nasu K and Narahara H: Role of
chemokines in the pathogenesis of endometriosis. Front Biosci
(Schol Ed). 3:1196–1204. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Arici A, Seli E, Zeyneloglu HB, Senturk
LM, Oral E and Olive DL: Interleukin-8 induces proliferation of
endometrial stromal cells: A potential autocrine growth factor. J
Clin Endocrinol Metab. 83:1201–1205. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garcia-Velasco JA and Arici A:
Interleukin-8 expression in endometrial stromal cells is regulated
by integrin-dependent cell adhesion. Mol Hum Reprod. 5:1135–1140.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Iwabe T, Harada T, Tsudo T, Tanikawa M,
Onohara Y and Terakawa N: Pathogenetic significance of increased
levels of interleukin-8 in the peritoneal fluid of patients with
endometriosis. Fertil Steril. 69:924–930. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Premack BA and Schall TJ: Chemokine
receptors: Gateways to inflammation and infection. Nat Med.
2:1174–1178. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ulukus M, Ulukus EC, Seval Y, Cinar O,
Zheng W and Arici A: Expression of interleukin-8 receptors in
patients with adenomyosis. Fertil Steril. 85:714–720. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Antiñolo G, Fernández RM, Noval JA, Molini
JL and Borrego S: Analysis of the involvement of CCR5-Delta32 and
CCR2-V64I variants in the development of endometriosis. Mol Hum
Reprod. 10:155–157. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wieser F, Dogan S, Klingel K, Diedrich K,
Taylor RN and Hornung D: Expression and regulation of CCR1 in
peritoneal macrophages from women with and without endometriosis.
Fertil Steril. 83:1878–1881. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chapron C, Borghese B, Streuli I and de
Ziegler D: Markers of adult endometriosis detectable in
adolescence. J Pediatr Adolesc Gynecol. 24 5 Suppl:S7–S12. 2011.
View Article : Google Scholar : PubMed/NCBI
|