Introduction
Cerebral ischemia describes the situation in which
blood flow to the brain is insufficient in meeting metabolic
demand. This results in cerebral hypoxia, the death of brain
tissue, cerebral infarction or ischemic stroke (1,2).
Cerebral ischemia is associated with cerebrovascular disease or
disorder (CVD), which is a common disease with high morbidity and
mortality. Ischemic cerebrovascular disease (ICVD) has a high
incidence and high recurrence rate accounting for 70% of total
cerebrovascular disease, which is clinically associated with
myocardial infarction, thrombolytic therapy, coronary angioplasty,
coronary revascularization and heart transplantation (3). Although it is widely accepted that
blood reflow following ischemia is important for increasing
cardiomyocyte survival to minimize the cardiac dysfunction and
myocardial damage, reperfusion itself additionally aggravates
myocardial injury (4). Therefore,
the prevention and treatment of cerebral ischemia-reperfusion (IR)
injury is becoming increasing important in ICVD therapy.
However, the exact pathogenesis of cerebral IR
injury remains unknown. Cumulative evidence suggests that numerous
factors are involved in ischemic brain injury, including apoptosis,
acid poisoning, ionic imbalance, excitotoxicity, oxidative stress,
peri-infarct depolarization, nutritive stress, inflammation and
reactive oxygen species (ROS) (5–12).
Furthermore, numerous signaling pathways have been implicated in
the pathogenesis of cerebral IR injury, including mitogen-activated
protein kinases (13,14). Additionally, during ischemic brain
damage, the cell metabolism abnormalities may be derived from the
interrupted blood, oxygen and glucose supply, which ultimately
result in neuronal death or apoptosis (2). Therefore, anti-inflammation and
anti-oxidant treatment strategies are being developed to treat
cerebral IR injury.
Due to the complex alterations associated with IR
injury, it has become difficult to identify a novel pharmacological
drug with a protective effect. Previous studies preferentially
developed therapeutics for IR injury based on the advantages of
traditional Chinese medicine (TCM) (15,16).
Seaweed is one of the most abundant sources of polysaccharides used
in TCM, with alginate, agar, fucoidan, agarose and carrageenan all
present (16). A previous study
demonstrated that M. charantia polysaccharide (MCP) could be
a promising neuroprotective ingredient of M. charantia and its
mechanisms could be at least in part attributed to its antioxidant
activities and its inhibition of JNK3 signaling cascades during
cerebral ischemia/reperfusion injury in a rat model (17). Zhou et al (18) reported that treatment with
Ganoderma lucidum polysaccharides, aloe polysaccharide,
ginkgo leaf tablet and nimodipine following cerebral ischemic
injury significantly reduced caspase-3 protein and mRNA expression
levels in the cerebral cortex compared with the cerebral
ischemia/reperfusion Sprague-Dawley (SD) rat model group, and were
significantly increased compared with the sham surgery group
(P<0.05). Lu et al (19)
revealed that the protective effect of aloe polysaccharide on
cerebral ischemia may be due to the inhibition of neuronal cell
apoptosis via downregulation of caspase-3 protein expression.
Agaricus brasiliensis polysaccharides may affect
malondialdehyde and superoxide dismutase activity, and caspase-3
level in IR rats, resulting in cardiovascular protection (20). The effect of polysaccharide peptide
(PSP) on cerebral injury caused by IR remains unknown, even though
substantial research and numerous studies of the neuroprotective
effect of the polysaccharides have been performed. Additionally,
the mechanism requires further clarification.
At present, the study of cerebral IR injury has
attracted considerable research interest. Cerebral ischemia leads
to serious damage of local tissue and brain function. The degree of
damage is associated with the duration of ischemia and the
remaining blood flow. Severe ischemia with a longer duration may
lead to infarction. The primary therapeutic method is IR, allowing
ischemia brain tissue access to a supply of oxygen and the
necessary nutrients again. However, the therapeutic efficacy of
reperfusion is undesirable. Cerebral IR may result in neuron death
due to an increase in the generation of free radicals caused by IR
injury (21). Furthermore,
reperfusion may lead to a reduction in the content of excitatory
amino acid and alteration of the ultramicrostructure of brain
tissue, including mitochondrial swelling, calcium deposition and
endothelial swelling (1).
Therefore, in the present study, the neuroprotective action and
underlying mechanisms of PSP against cerebral IR-induced injury
were investigated in vitro and in vivo.
Materials and methods
Cell culture
Mouse neuroblastoma cell line N2a was purchased from
BeNa Culture Collection (Jiangsu, China) and incubated in culture
medium (Dulbecco's modified Eagle's medium and opti-Minimum
Essential Medium 1:1, both from Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing 10% fetal bovine serum (Hyclone;
GE Healthcare Life Sciences, Logan, UT, USA) in a 5% CO2
incubator at 37°C. The medium was replaced with fresh medium every
two days. The cells were used for propagation or experimental use
when 70–80% confluent.
Establishment of the model of IR in
vitro
The cells were divided into five groups; Negative
control (NC) group, IR injury group, PSP low dose treatment group
(PSP1; l 10 mg/ml), PSP middle dose treatment group (PSP2; 30
mg/ml) and PSP high dose treatment group (PSP3; 50 mg/ml).
When the cells were cultured to 80% confluence, the
cells in the NC group were continually cultured in complete culture
solution, while the cells in the IR and PSP groups were treated
with an equal volume of equilibrated medium, containing 116 mM
NaCl, 5.4 mM KCl, 0.8 mM MgSO4, 1 mM
NaH2PO4, 0.9 mM CaCl2 and 10 mg/l
phenol red. The cells were incubated in 5% CO2 and 95%
N2 for 30 min at 37°C, and subsequently the equilibrated
medium was replaced with complete culture solution. Different
concentrations of PSP (Xi'an Tianrui Biological Technology Co.,
Ltd. Xi'an, China) were added to the PSP groups.
MTT assay
The cells were seeded on a 96-well plate at a
density of 105 cells/well and incubated in 5%
CO2 at 37°C. Following cell growth to 80%, different
concentrations of PSP were added to the wells. After 24 h, 10 µl (1
g/l) of MTT solution (Gibco; Thermo Fisher Scientific, Inc.) was
added into each well and cultured for 4 h at 37°C. The supernatant
was removed and 150 µl dimethyl sulfoxide was added. The optical
(OD) was measured with the multifunctional microplate reader
SpectraMax M5 (Molecular Devices, LLC, Sunnyvale, CA, USA) at 490
nm. Each group was conducted in six parallel experiments.
Lactate dehydrogenase (LDH) activity
determination
The medium was collected and centrifuged at 4°C and
380 × g for 5 min to obtain the supernatant. The LDH activity was
determined by ELISA. The experimental process was completely
performed according to the protocols of an LDH kit (AB102526;
Abcam, Cambridge, UK). The OD value of LDH protein was measured
using a microplate reader at 450 nm. The standard curve was
established, according to the concentration and OD value. The
experiment was repeated three times.
Detection of caspase-3 activity in
cells
The cells were harvested and lysed in RIPA buffer
(Beyotime Institute of Biotechnology, Haimen, China) on ice for 20
min. Following centrifugation at 44°C and 380 × g for 5 min, the
supernatant was collected and the total proteins were quantified,
according to a caspase-3 activity assay kit (Jiancheng
Bioengineering Institute, Nanjing, China). The measurement of the
caspase-3 activity was processed in protein analytic buffer (20
nmol/l HEPES, 2 mmol/l dithiothreitol, 10% glycerol and 20 µmol/l
DEVD-pNA) at 37°C for 2 h. The OD value of the sample was measured
at 405 nm using a microplate reader. The content of caspase-3 was
calculated using the standard curve. The experiment was repeated
three times.
Animals
A total of 75 healthy male SD rats (10–12 weeks;
200–250 g) were purchased from The Experimental Animal Center of
Guangzhou University of Traditional Chinese Medicine (Guangzhou,
China). All rats were kept in controlled conditions at 25±3°C and
35–60% relative humidity. A 12-h light/dark cycle was maintained.
All the animals were allowed to acclimate to laboratory conditions
for ≥1 week prior to the experiment and had free access to water
and food. All the animal works were performed stringently,
according to the regulation of the Guide for the Care and Use of
Laboratory Animals in China (22).
The protocol was approved by the Committee of the Ethics of Animal
Experiments of Shanghai Jiaotong University Affiliated Sixth
Hospital Provincial Centre for Disease Control and Prevention
(proposal authorization no. SYXK/2016/050122). All surgeries were
conducted using appropriate anesthesia and all efforts were made to
reduce the pain and suffering of the animals.
In vivo IR model
The rats were randomly divided into five groups;
sham operation group (sham; n=15), IR injury group (n=15), PSP1
(150 mg/kg; n=15), PSP2 (200 mg/kg; n=15) and PSP3 (250 mg/kg;
n=15). First, 10% chloral hydrate (Sinopharm Chemical Reagent Co.,
Ltd., Shanghai, China) was injected intraperitoneally into rats at
a dose of 300–350 mg/kg. There were no significant signs of
peritonitis. Following anesthesia, the model of the right middle
cerebral artery occlusion was selected for further experimental
use. In the sham group, the right common carotid artery, external
carotid artery and internal carotid artery were separated without a
plug bolt. The plug was pulled out in the other groups after 2 h of
ischemia, followed by reperfusion for 24 h. The low, middle and
high dose group rats received 150, 250, 250 mg/kg, respectively,
via intraperitoneal injection at 2 h after ischemia; the sham group
and IR group were intraperitoneally injected with an equal volume
of saline solution at the same time point.
Neurological assessment
The neurological assessment was processed, according
to the Zea Longa method (23). The
neurological deficiency was assessed consistently according to a
5-point scale system: 0, no neurological deficit; 1, the
contralateral torso and forelimb may not be thoroughly stretched;
2, when the tail was held, the animal may be turned to the
ipsilateral side; 3, no spontaneous motor activity or falling to
the left side; 4, unable to walk or loss of consciousness. Each
experiment was repeated three times.
Calculation of cerebral infarction
volume
The area of cerebral infarction was identified using
triphenyl tetrazolium chloride (TTC; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) staining. Following the neurological assessment
grading, five rats in each group were intraperitoneally injected
with 10% chloral hydrate. The brains were collected following
anesthesia and decapitation, and were subsequently placed in −20°C
for 15 min. The tissue slices were processed to ~1 mm thickness
using a cryostat microtome. The slices were placed in 2% TTC
solution, incubated at 37°C for 30 min, and were subsequently fixed
in 4% paraformaldehyde at 4°C for 24 h. Images of the slices were
captured, the infarct area in each section was calculated using an
image analyzer (Image-Pro Plus version 6.0; Media Cybernetics,
Inc., Rockville, MD, USA). and the cerebral infarction volume
percentage was calculated using the formula:
Infarction volume(%)=right cerebral
infarction volume xslice thicknessright brain tissue
volumex100
The infarcted volume was calculated as a percentage
of the total brain area.
Histopathological staining
analysis
Male Sprague-Dawley rats were injected
intraperitoneally with 10% chloral hydrate (300–350 mg/kg body
weight; Sinopharm, Chemical Reagent Co., Ltd.), and the hearts were
exposed through thoracotomy following anesthesia. The needle was
inserted from the apical position, the right atrial appendage vein
was cut open and 0.9% saline solution was rapidly perfused in the
heart until the clear liquid outflowed from right atrial appendage,
subsequently 4% paraformaldehyde (Sinopharm, Chemical Reagent Co.,
Ltd.) was perfused to the heart followed by quick decapitation and
collection of the brain, which was fixed at 4°C for 24 h with 4%
paraformaldehyde. The coronal brain tissue was cut into slices of 3
mm thickness followed by routine embedding in paraffin. The brains
were subsequently sliced to 5 µm thickness, dewaxed with xylene and
hydrated using a series of graded concentrations of ethanol (100%
ethanol, 5 min; 95% ethanol, 1 min; 80% ethanol, 5 min; 75%
ethanol, 5 min; distilled water, 2 min). Hematoxylin and eosin
(H&E) staining was performed using the routine method at room
temperature for 12 min. Following dehydration, sections were
treated with xylene at room temperature for 10 min twice. The
tissue sections was examined using a light microscope (OriGene
Technologies, Inc.) to examine the histopathological morphology at
a magnification of ×100.
Western blotting
Western blotting was performed to determine the
protein expression of silent information regulator protein (SIRT1),
peroxisome proliferator-activated receptor γ coactivator-1α
(PGC-1α), caspase-3, B-cell lymphoma 2 (Bcl-2) and Bcl-2-like
protein 4 (Bax), according to a standard protocol. The brain
tissues were lysed on ice and centrifuged at 4°C and 13,680 × g for
20 min. The supernatant was extracted using RIPA buffer and the
protein concentration was measured using a bicinchoninic acid
protein quantification kit. Total proteins (20 µg) were loaded and
separated by 12% SDS-PAGE, and transferred to polyvinylidine
difluoride membranes (GE Healthcare). The membranes were blocked at
25°C with 5% skimmed milk in Tris-buffered saline (TBS) containing
0.1% Tween-20 for 1 h and subsequently incubated overnight at 4°C
with the individual primary antibodies, rabbit-anti-SIRT1 (9475;
1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA), PGC-1α
(#2178, 1:1,000; Cell Signaling Technology, Inc.), Bax (2772;
1:1,000; Cell Signaling Technology, Inc.), Bcl-2 (15071; 1:1,000;
Cell Signaling Technology, Inc.), caspase-3 (9662; 1:1,000; IDUN
Pharmaceuticals, Inc., San Diego. CA, USA), β-actin (4970; 1:1,000;
Santa Cruz Biotechnology, Dallas, TX, USA). Following washing for
three times with TBS-Tween for 5 min every time, the membrane was
incubated with secondary antibodies of horseradish
peroxidase-conjugated immunoglobulin G (Sigma-Aldrich; Merck KGaA;
1:5,000) for 1 h at room temperature. Following washing three times
for 5 min, the proteins were visualized with enhanced
chemiluminescent substrates (GE Healthcare), and the image of each
band was further quantified using Multigauge computer software
version 3.0 (Berthold Australia Pty Ltd, Bundoora, Australia).
Statistical analysis
All statistical analyses were performed with SPSS
software version 13.0 for windows (SPSS, Inc., Chicago, IL, USA).
Data are presented as the mean ± standard deviation. One-way
analysis of variance with Bonferroni's correction was used to
compare the statistical difference between multiple groups. The
Least Significant Difference test was used for comparison between
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
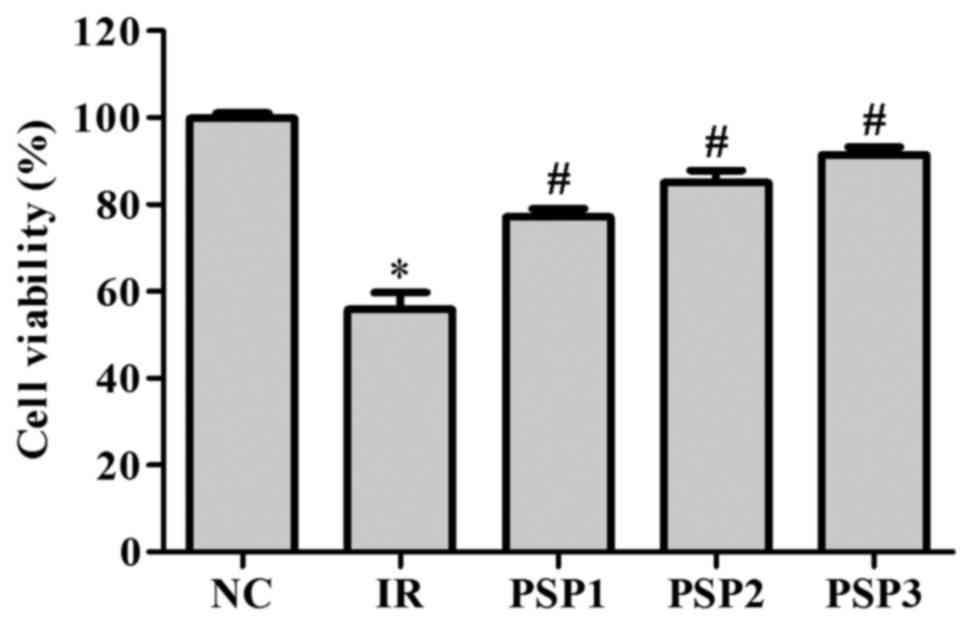
Effect of PSP on cell viability
Following simulation of IR conditions, the N2a cell
viability was analyzed in an MTT assay (Fig. 1). Compared with the NC, the cell
viability significantly decreased following IR with a survival rate
of ~60% (Fig. 1; P<0.05).
Following treatment with PSP, the cell activity was significantly
improved compared with the IR group and exhibited a dose-dependent
manner (P<0.05). The cell viabilities of the PSP1, PSP2 and PSP3
groups were ~84, 90 and 95%, respectively. The data indicated that
treatment with higher doses of PSP resulted in significantly higher
cell viability (P<0.05).
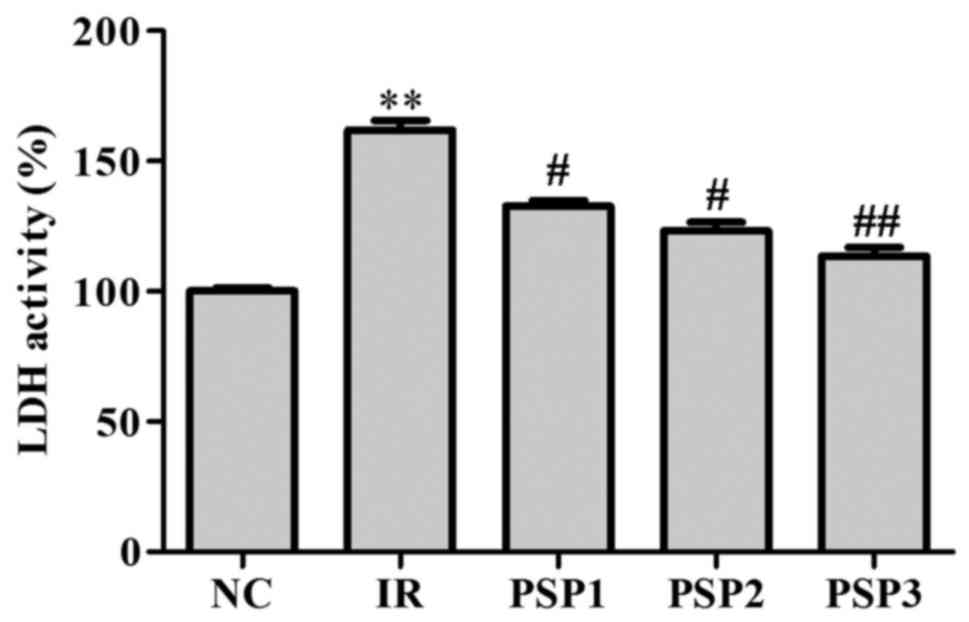
Effect of PSP on cell membrane
permeability
Neuronal injury or death was assessed according to
levels of LDH released into the extracellular medium as previously
described (24). As demonstrated
in Fig. 2, the IR group exhibited
a significantly higher LDH activity (170%) compared with the NC
(P<0.01). Following administration of PSP, the activity of LDH
decreased with the concentration of PSP increasing between 10 and
50 mg/ml. A high dose of PSP was able to significantly decrease the
activity of LDH with the expression percentage of 105% in culture
medium compared with the NC (P<0.01; Fig. 2), which suggests that PSP is able
to reduce the cell membrane permeability and consequently reduce
injury to protect cells.
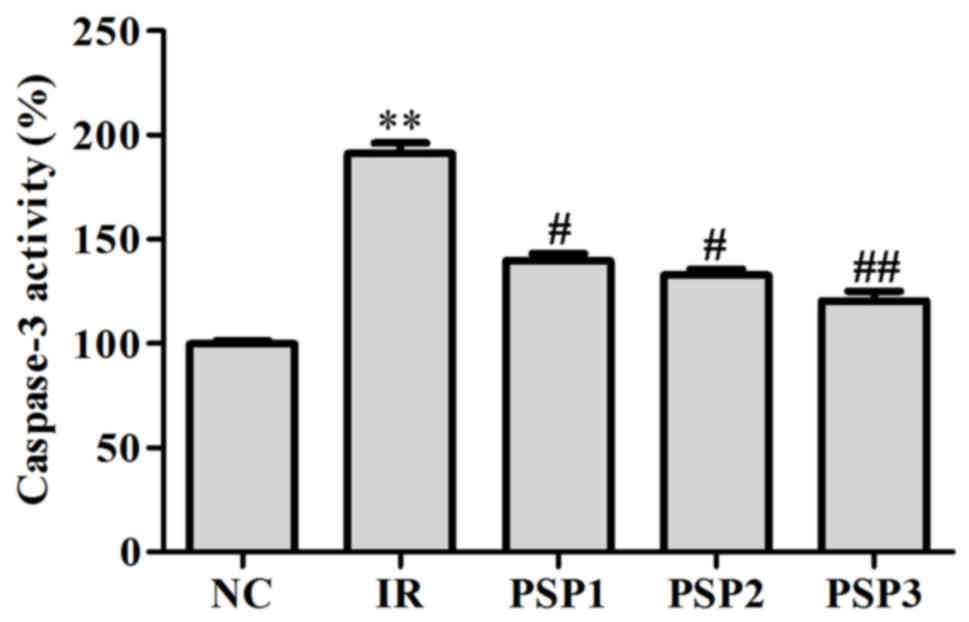
Effect of PSP on caspase-3 activity in
cells
The activity of caspase-3 was determined by
caspase-3 specific substrates. The results demonstrated that the
activity of caspase-3 was significantly increased by up to ~90%
following IR, and decreased markedly following treatment with PSP
(P<0.01; Fig. 3). The activity
of caspase-3 following treatment with PSP1, PSP2 and PSP3 was ~125,
120 and 110%, respectively. The results suggested that PSP may
reduce the activity of caspase-3 to reduce apoptosis.
Effect of PSP on neurological function
score
Neurological function assessment is a crucial
procedure for the medical care of neurological patients, which may
detect the presence of a neurological disease or injury and monitor
its progression. In the present study, each group of rats was
scored as 0–3 with no subarachnoid hemorrhage observed, suggestive
of successful establishment of the model. As demonstrated in
Table I, the mice in the sham
group had no neurological deficit symptoms. In the IR group, the
rats exhibited a neurological deficit with a higher score of 2.63
(P<0.01) compared with the sham group. Compared with the IR
group, the neurological deficits following treatment with PSP were
reduced (P<0.05), particularly in the PSP3 group (P<0.01).
The neurological function scores of the low and middle-dose PSP
groups were 2.17±0.25 and 1.53±0.33, respectively, which is higher
compared with high-dose PSP (1.16±0.19). The neurological deficit
score was the lowest in the PSP3 group.
 | Table I.Neural function defect scale in
different treatment groups. |
Table I.
Neural function defect scale in
different treatment groups.
| Groups | n | Score |
|---|
| Sham | 15 | 0 |
| IR | 15 |
2.63±0.48a |
| PSP1 | 15 |
2.17±0.25b |
| PSP2 | 15 |
1.53±0.33b |
| PSP3 | 15 |
1.16±0.19c |
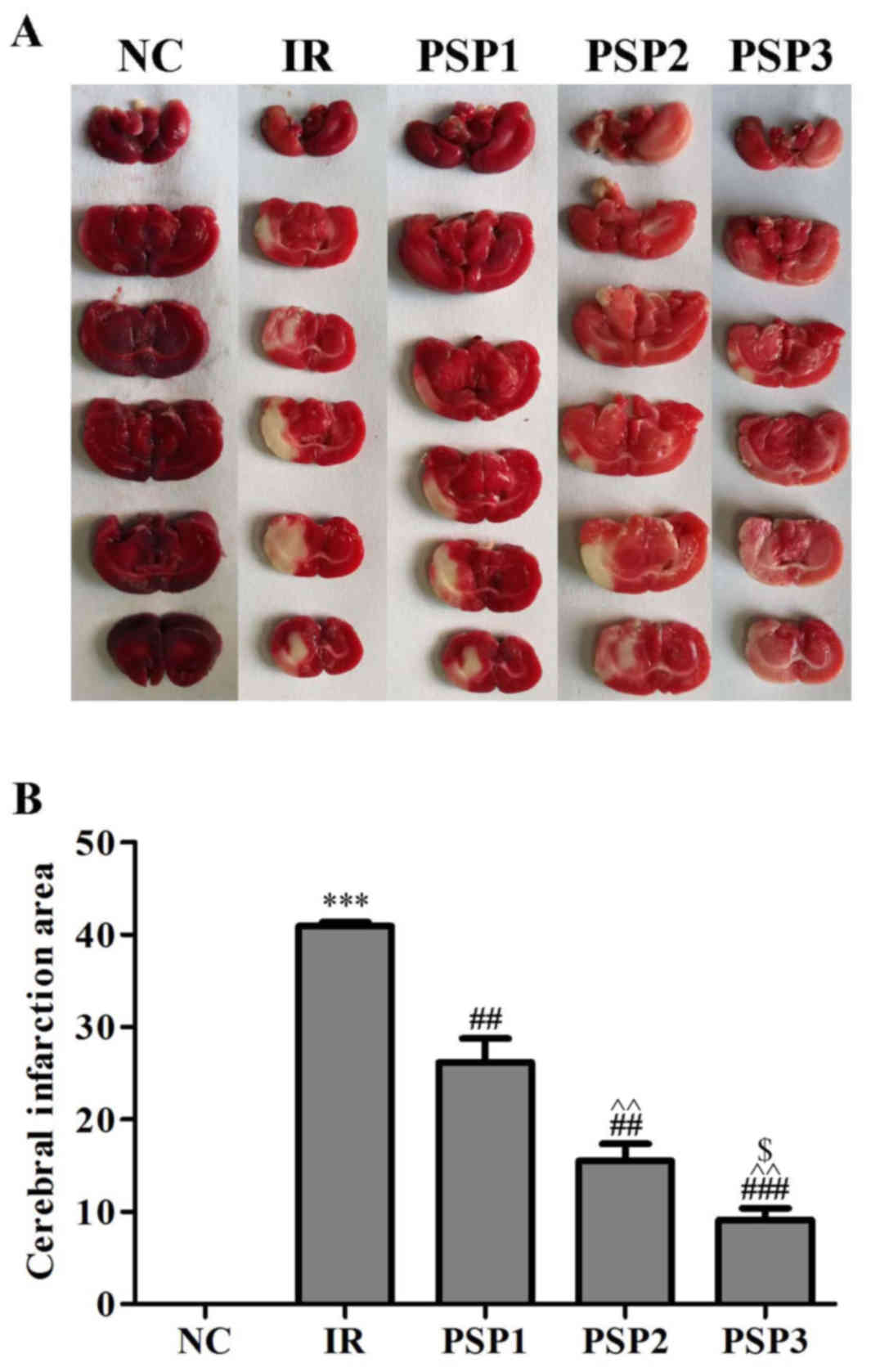
Effect of PSP on cerebral infarction
volume
To assess the protective effects of PSP against
brain ischemia injury in vivo, a model of ischemia was
established. Infarction size was assessed by the appearance of a
white region following TTC staining (Fig. 4). The results demonstrated that
there was no white region observed in brain tissue in the sham
group, and the infarction was observed in the IR group and PSP
groups. Compared with the IR group (42.93%), the infarction volume
in the PSP treatment groups decreased significantly (P<0.01).
Among the PSP administration groups, the score of PSP3 group
(25.16%) was lower compared with PSP1 (36.17%; P<0.01) and PSP2
(33.53%; P<0.05). In addition, there was a significant
difference between low- and middle-dose group (P<0.01). These
results indicated that PSP may effectively reduce the infarction
size in a dose-dependent manner.
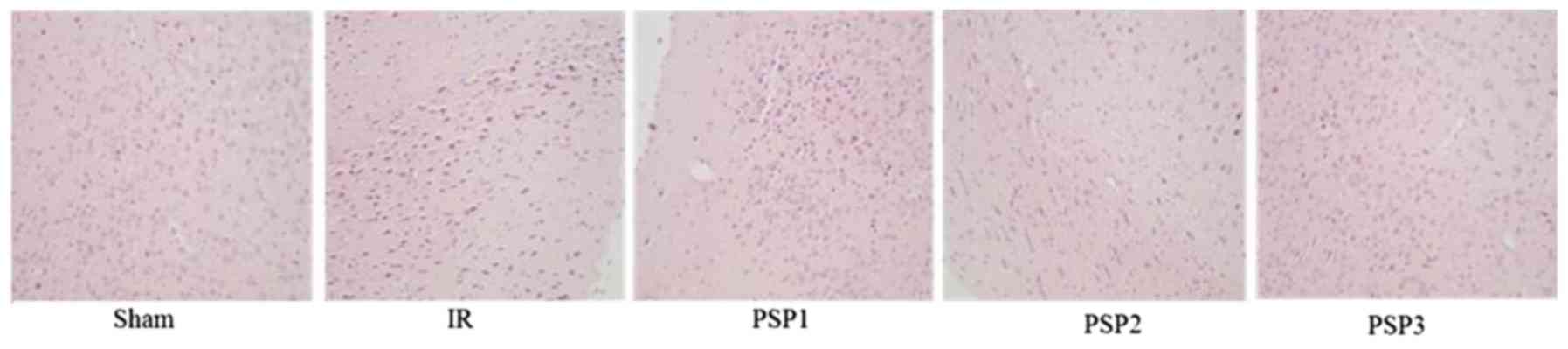
Histopathological evaluation
H&E staining was used to examine the
morphological alterations of the hippocampus area upon treatment
with PSP following IR injury. As demonstrated in Fig. 5, the nerve cells in the sham group
were structurally complete, arranged neatly, stained evenly and
clearly. In the IR group, the number of nerve cells decreased, were
arranged disorderly and the stain color deepened. Compared with the
IR group, the number of cells in the PSP-treated groups increased,
and the morphology of the nerve cells recovered well. These results
indicated that PSP may reduce brain cell death in the hippocampus
area caused by IR and reduce brain injury.
Effect of PSP on SIRT1, PGC-1α, Bax,
Bcl-2 and caspase-3 protein expression
To verify whether the SIRT1 signaling pathway
participates in the protection from IR injury mediated by PSP, the
expression of SIRT1 and PGC-1α was examined by western blotting
following IR injury. As demonstrated in Fig. 6, the expression of SIRT1 and PGC-1α
was decreased in the IR group, with ~0.3 fold and ~0.25 fold
change, respectively, compared with the sham group (P<0.01).
Following intraperitoneal injection of different concentrations of
PSP (150, 200 and 250 mg/kg), the expression of SIRT1 and PGC-1α
was upregulated (P<0.05), particularly in the PSP3 group (~0.9
fold change and ~1.1 fold change, respectively) compared with the
IR group. These results demonstrated that PSP may upregulate the
expression of neuroprotective factors SIRT1 and PGC-1α, which may
protect cerebral IR injury in rats.
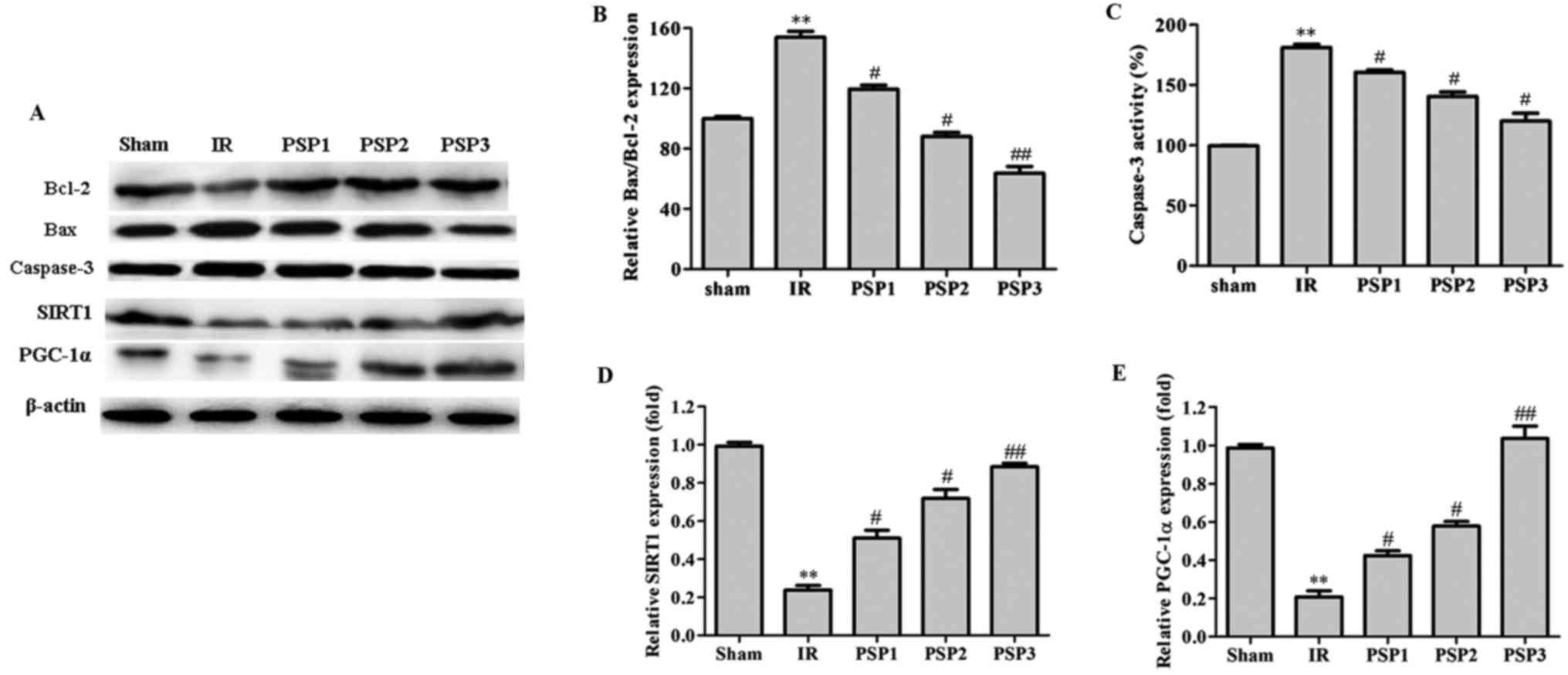 | Figure 6.Effect of PSP on the expression of
related proteins (Bcl-2, Bax and caspase-3, SIRT1 and PGC-1α) in
the ischemic hippocampus of rats in each group. (A) The protein
analyzed by western blotting. (B) The ratio of Bax to Bcl protein
level. The quantitative graph of (C) caspase-3, (D) SIRT1 and (E)
PGC-1 α protein levels. PSP1 is the low dose group (150 mg/kg),
PSP2 is the middle dose group (200 mg/kg) and PSP3 is the high dose
group (250 mg/kg). The data are expressed as the mean ± standard
deviation. **P<0.01 vs. respective sham; #P<0.05,
##P<0.01 vs. respective IR. PGC-1α, peroxisome
proliferator-activated receptor γ coactivator-1α; Bcl-2, B-cell
lymphoma 2; Bax, Bcl-2-like protein 4; SIRT1, silent information
regulator protein 1; PGC-1α, peroxisome proliferator-activated
receptor γ coactivator-1α; PSP, polysaccharide peptide; IR,
ischemia reperfusion. |
The effect of PSP on Bax, Bcl-2 and caspase-3 was
additionally examined by western blotting (Fig. 6). Compared with the sham group, the
Bax/Bcl-2 protein expression level in the IR group was
significantly increased at ~160% (P<0.01). Compared with the IR
group, the Bax/Bcl-2 protein expression in the PSP group was
downregulated to ~120, 90 and 68%, corresponding to PSP1, PSP2 and
PSP3 (P<0.05). It is demonstrated in Fig. 6 that the caspase-3 protein
expression level in the IR group was significantly increased
(P<0.01) compared with the sham group. However, following
treatment with PSP, the expression of caspase-3 protein was
significantly decreased (P<0.05). A dose of 250 mg/kg PSP was
able to downregulate the expression of caspase-3 more effectively.
These data suggested that the protection of PSP against cerebral IR
in rats may be due to an increase in Bcl-2 protein expression and a
decrease in Bax and caspase-3 protein expression, thus reducing the
apoptosis rate and reducing injury.
Discussion
In the present study, the cell viability, and the
activity of LDH and caspase-3 were examined by an MTT assay and
ELISA in vitro. A similar environment of cerebral IR injury
was stimulated for the N2a cells. It was identified that higher
concentrations of PSP markedly improved the cell viability of the
N2a cells. PSP reduced the activity of LDH and caspase-3.
Therefore, the in vitro study suggested that PSP may be able
to improve cell viability, increase the expression of caspase-3
protein and protect N2a cells against cerebral IR injury in
vitro.
Following establishment of an IR model in SD rats,
the PSP effect on neuroprotection was examined in vivo. It
is reported that focal cerebral ischemia may cause cerebral cell
death, and subsequently result in neuronal deficit symptoms and
local infarction (25). The
neuronal deficit severity and infarction area indicate the grade of
brain injury (26). In the animal
experiments, treatment with PSP exhibited neuroprotective effects,
demonstrated by alleviation of histological injury in the ischemic
cortex and neurological deficits derived from IR injury in rats.
All treatment groups with different doses of PSP has reduced the
cerebral infraction and improved neurological scores at 24 h post
reperfusion, and the most marked effect of PSP was identified in
the group treated with 250 mg/kg PSP. Therefore, a higher
concentration of PSP was used to evaluate the protection and its
side effect on the other functions. The morphology of infarct nerve
cells determined by H&E staining the neuroprotection mediated
by PSP. The results of the H&E staining and the caspase-3
activity assay revealed the neuroprotective effect of PSP was
associated with a reduction of neuronal loss and apoptosis induced
by IR injury.
During the pathological process of cerebral IR
injury, apoptosis in neurons serves a key role, according to a
previous study (4). Apoptosis is
controlled cell death regulated by the cell's natural
self-destruction. In the process of apoptosis, specific caspase
enzymes affect proteins that have crucial functions (27). At present, two pathways of
apoptosis have been well documented: The extrinsic and intrinsic
pathways. The extrinsic pathway is initiated by extracellular
insult and predominately relies on caspase-8 activation. A cascade
of proteases are activated by caspase-8, including caspase-9 and
caspase-7, to trigger caspase-3, which executes apoptosis (28). Mechanically, caspase-3 is able to
cleave itself and affect the other key proteins, including the DNA
breaks of canonical apoptosis (29). Therefore, activation and expression
of caspase-3 promotes cell apoptosis. In the normal mouse brain,
caspase-3 expression is at lower level (30). In the present study, caspase-3
activity was increased significantly following IR, and decreased
markedly following treatment with PSP for 24 h.
The intrinsic pathway, which is also termed the
mitochondrial pathway due to the essential role of mitochondria, is
initiated by Bax. Bax translocates to the mitochondrial membrane
from the cytosol and competes with other members of the Bcl-2
family. Therefore, the activation of the intrinsic pathway is
associated with a ratio of Bcl-2 and Bax. An imbalance between
Bcl-2 and Bax results in the activation of caspase-9 (31). To examine the mechanisms that are
altered by PSP in the IR injury model, the Bcl-2/Bax ratio was
assessed. The present results suggested a significant difference in
Bax/Bcl-2 ratio between the sham group and IR group. The Bax/Bcl-2
ratio in the IR group reached its highest value after 22 h of
reperfusion. However, the Bax/Bcl-2 ratio was reverted and
decreased following treatment with PSP, as the decreased Bax/Bcl-2
ratio may prevent neuronal cell apoptosis by impeding the Bax/Bax
homodimer formation and promoting Bcl-2/Bax heterodimer formation.
Thus, the present results again indicated that treatment with PSP
following IR injury may protect neuronal cells from apoptosis.
SIRT1, a homologue of silent information regulator
(Sir2) protein, is the closest mammalian homologue of yeast Sir2.
It is classified as a NAD-dependent deacetylase and a nuclear
sirtuin, although it is not restricted to the nucleus and has
important non-nuclear functions (32). SIRT1 is a key factor in apoptotic
cell death, cellular senescence and vascular growth associated with
IR (33). SIRT1 physically
interacts with and deacetylates PGC-1α at multiple lysine sites,
resulting in increased activity of PGC-1α. Xia et al
(34) reported that SIRT1 and
AMP-activated protein kinase (AMPK) were involved in the
stimulation of nitric oxide (NO) production by endothelial NO
synthase and enhancement of NO bioavailability, which may be an
important mechanism associated with the protective effect of SIRT1
in IR injury. Feng et al (35) demonstrated that treatment with
bakuchiol attenuates IR injury by reducing IR-induced mitochondrial
oxidative damage via the activation of SIRT1/PGC-1α signaling. The
SIRT1/AMPK pathway is involved in the protection induced by a
number of agents, including aspirin (36), resveratrol (37). Notably, SIRT1 and AMPK expression
were reported to be increased by heat shock protein 70 (HSP70)
overexpression (38). The present
study additionally demonstrated that the levels of SIRT1 and PGC-1α
protein were elevated in all three PSP groups. The next step will
be to examine whether HSP70 also has a role in cerebral IR injury,
and the association of HSP70, SIRT1 and PGC-1α and Bcl-2/Bax, and,
subsequently, the effect on cerebral IR injury by the intervention
of these factors. These results indicated that the mechanism of
seaweed-derived PSP protection of the brain from IR injury may be
attributed to these alterations and their associated signaling
pathway.
In the present study, the results revealed that the
protective effect of seaweed polysaccharide on cerebral IR injury
in vitro and in vivo and the mechanism may be
associated with its effect on the regulation of the SIRT1/PGC-1α
signaling pathway. Thus, application of seaweed polysaccharide may
provide a novel promising approach for cerebral IR injury
therapy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
PX and DW participated in the conception and design
of the study. PX and KM conducted the experiments and data
acquisition. JW and WL conducted the histological examination of
the brain tissue and analyzed and interpreted the data. PX was a
major contributor in writing the manuscript. DW revised the
manuscript and provide the final approval of the submitted
version.
Ethics approval and consent to
participate
All the animal works were performed stringently
according to the regulation of the Guide for the Care and Use of
Laboratory Animals in China. The protocol was approved by the
Committee of the Ethics of Animal Experiments of Shanghai Jiaotong
University Affiliated Sixth Hospital Provincial Centre for Disease
Control and Prevention (proposal authorization no.
SYXK/2016/050122).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IR
|
ischemia-reperfusion
|
|
PSP
|
polysaccharide peptide
|
|
LDH
|
lactate dehydrogenase
|
|
CVD
|
cerebrovascular disease
|
|
ICVD
|
ischemic cerebrovascular disease
|
|
ROS
|
reactive oxygen species
|
|
TCM
|
traditional Chinese medicine
|
|
NC
|
negative control
|
|
TTC
|
triphenyl tetrazolium chloride
|
|
TBS
|
Tris-buffered saline
|
|
SIRT1
|
silent information regulator protein
1
|
|
PGC-1α
|
peroxisome proliferator-activated
receptor γ coactivator-1α
|
References
|
1
|
Yamashita T and Abe K: Recent progress in
therapeutic strategies for ischemic stroke. Cell Transplant.
25:893–898. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dong Q, Lin X, Shen L and Feng Y: The
protective effect of herbal polysaccharides on ischemia-reperfusion
injury. Int J Biol Macromol. 92:431–440. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rao PR, Kumar VK, Viswanath RK and
Subbaraju GV: Cardioprotective activity of alcoholic extract of
Tinospora cordifolia in ischemia reperfusion induced myocardial
infarction in rats. Biol Pharm Bull. 28:2319–2322. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalogeris T, Baines CP, Krenz M and
Korthuis RJ: Cell biology of ischemia/reperfusion injury. Int Rev
Cell Mol Biol. 298:229–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zerna C, Assis Z, d'Esterre CD, Menon BK
and Goyal M: Imaging, intervention, and workflow in acute ischemic
stroke: The calgaryapproach. AJNR Am J Neuroradiol. 37:978–984.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sugawara T and Chan PH: Reactive oxygen
radicals and pathogenesis of neuronal death after cerebral
ischemia. Antioxid Redox Signal. 5:597–607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adibhatla Muralikrishna R and Hatcher JF:
Phospholipase A2, reactive oxygen species, and lipid peroxidation
in cerebral ischemia. Free Radic Biol Med. 40:376–387. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu M, Wang HF, Zhang YY and Zhuang HW:
Protection of rats spinal cord ischemia-reperfusion injury by
inhibition of MiR497 on inflammation and apoptosis: Possible role
in pediatrics. Biomed Pharmacother. 81:337–344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu T, Yao Q, Hu X, Chen C, Yao H and Chao
J: The role of MCPIP1 in ischemia/reperfusion injury-induced HUVEC
migrationand apoptosis. Cell Physiol Biochem. 37:577–591. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arda-Pirincci P and Bolkent S: The role of
epidermal growth factor in prevention of oxidative injury and
apoptosis induced by intestinal ischemia/reperfusion in rats. Acta
Histochem. 116:167–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shokeir AA, Barakat N, Hussein AM,
Awadalla A, Harraz AM, Khater S, Hemmaid K and Kamal AI: Activation
of Nrf2 by ischemic preconditioning and sulforaphane in
renalischemia/reperfusion injury: A comparative experimental study.
Physiol Res. 64:313–323. 2015.PubMed/NCBI
|
|
12
|
Ju J, Wu J and Hou R: Role of the p38
mitogen-activated protein kinase signaling pathway
inestrogen-mediated protection following flap ischemia-reperfusion
injury. Cell Biochem Funct. 34:522–530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kovalska M, Kovalska L, Pavlikova M,
Janickova M, Mikuskova K, Adamkov M, Kaplan P, Tatarkova Z and
Lehotsky J: Intracellular signaling MAPK pathway after cerebral
ischemia-reperfusion injury. Neurochem Res. 37:1568–1577. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Imahashi K, Schneider MD, Steenbergen C
and Murphy E: Transgenic expression of Bcl-2 modulates energy
metabolism, preventscytosolic acidification during ischemia, and
reduces ischemia/reperfusion injury. Circ Res. 95:734–741. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei DF, Chen T, Yan M, Zhao W, Li F, Cheng
W and Yuan L: Synthesis, characterization, antioxditant activity
and neuroprotective effects of selenium polysaccharide from Radix
hedysari. Carbohydr Polym. 125:161–168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sanagi MM, Loh SH, Ibrahim Wan WN,
Pourmand N, Salisu A, Ibrahim Wan WA and Ali I: Agarose- and
alginate based biopolymers for sample preparation: Excellentgreen
extraction tools for this century. J Sep Sci. 39:1152–1159. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gong J, Sun F, Li Y, Zhou X, Duan Z, Duan
F, Zhao L, Chen H, Qi S and Shen J: Momordica charantia
polysaccharides could protect against cerebral ischemia/reperfusion
injury through inhibiting oxidative stress mediated c-Jun
N-terminal kinase 3 signaling pathway. Neuropharmacology.
91:123–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou ZY, Tang YP, Xiang J, Wua P, Jin HM,
Wang Z, Mori M and Cai DF: Neuroprotective effects of water-soluble
ganoderma lucidum polysaccharideson cerebral ischemic injury in
rats. J Ethno pharmacol. 131:154–164. 2010. View Article : Google Scholar
|
|
19
|
Lu ZQ, Deng YJ and Lu JX: Effect of aloe
polysaccharide on caspase-3 expression following cerebral ischemia
and reperfusion injury in rats. Mol Med Rep. 6:371–374. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang S, He B, Ge J, Zhai C, Liu X and Liu
P: Characterization of chemical composition of agaricus
brasiliensis polysaccharides and its effect on myocardial SOD
activity, MDA and caspase-3 level in ischemia-reperfusion rats. Int
J Biol Macromol. 46:363–366. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reis C, Wang Y, Akyol O, Ho WM, Ii RA,
Stier G, Martin R and Zhang JH: What's new in traumatic brain
injury: Update on tracking, monitoring and treatment. Int J Mol
Sci. 16:11903–11965. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi P, Zhang L, Wang J, Lu D, Li Y, Ren J,
Shen M, Zhang L and Huang J: Porcine FcεRI mediates porcine
reproductive and respiratory syndrome virus multiplication and
regulates the inflammatory reaction. Virol Sin. 33:249–260. 2018.
View Article : Google Scholar
|
|
23
|
Bösel J, Ruscher K, Ploner CJ and Valdueza
JM: Delayed neurological deterioration in a stroke patient with
postoperative acute anemia. Eur Neurol. 53:36–38. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haigis MC and Sinclair DA: Mammalian
sirtuins: Biological insights and disease relevance. Annu Rev
Pathol. 5:253–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zille M, Farr TD, Przesdzing I, Müller J,
Sommer C, Dirnagl U and Wunder A: Visualizing cell death in
experimental focal cerebral ischemia: Promises, problems, and
perspectives. J Cereb Blood Flow Metab. 32:213–231. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hyman BT and Yuan J: Apoptotic and non
apoptotic roles of caspases in neuronal physiology and
pathophysiology. Nat Rev Neurosci. 13:395–406. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Walsh JG, Cullen SP, Sheridan C, Lüthi AU,
Gerner C and Martin SJ: Executioner caspase-3 and caspase-7 are
functionally distinct proteases. Proc Natl Acad Sci USA.
105:12815–12819. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Manabat C, Han BH, Wendland M, Derugin N,
Fox CK, Choi J, Holtzman DM, Ferriero DM and Vexler ZS: Reperfusion
differentially induces caspase-3 activation in ischemic core and
penumbra after stroke in immature brain. Stroke. 34:207–213. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ferrer I and Planas AM: Signaling of cell
death and cell survival following focal cerebral ischemi: Lifeand
death struggle in the penumbra. J Neuropathol Exp Neurol.
62:329–339. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Potente M and Dimmeler S: Emerging roles
of SIRT1 in vascular endothelial homeostasis. Cell Cycle.
7:2117–2122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maiese K, Chong ZZ, Shang YC and Wang S:
Translating cell survival and cell longevity into treatment
strategies with SIRT1. Rom J Morphol Embryol. 52:1173–1185.
2011.PubMed/NCBI
|
|
33
|
Rodgers JT, Lerin C, Haas W, Gygi SP,
Spiegelman BM and Puigserver P: Nutrient control of glucose
homeostasis through a complex of PGC-1alpha and SIRT1. Nature.
434:113–118. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xia N, Förstermann U and Li HG:
Resveratrol and endothelial nitric oxide. Molecules.
19:16102–16121. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng J, Yang Y, Zhou Y, Wang B, Xiong H,
Fan C, Jiang S, Liu J, Ma Z, Hu W, et al: Bakuchiol attenuates
myocardial ischemia reperfusion injury by maintaining mitochondrial
function: The role of silent information regulator 1. Apoptosis.
21:532–545. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsai KL, Huang PH, Kao CL, Leu HB, Cheng
YH, Liao YW, Yang YP, Chien Y, Wang CY, Hsiao CY, et al: Aspirin
attenuates vinorelbine-induced endothelial inflammation via
modulating SIRT1/AMPK axis. Biochem Pharmacol. 88:189–200. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tamaki N, Cristina Orihuela-Campos R,
Inagaki Y, Fukui M, Nagata T and Ito HO: Resveratrol improves
oxidative stress and prevents the progression of periodontitis via
the activation of the Sirt1/AMPK and the Nrf2/antioxidant defense
pathways in a rat periodontitis model. Free Radic Biol Med.
75:222–229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu S, Xu J, Fang C, Shi C, Zhang X, Yu B
and Yin Y: Over-expression of heat shock protein 70 protects mice
against lung ischemia/reperfusion injury through SIRT1/AMPK/eNOS
pathway. Am J Transl Res. 8:4394–4404. 2016.PubMed/NCBI
|