Introduction
Hepatocellular carcinoma (HCC) is derived from
hepatocytes and has become one of the most prevalent and malignant
cancer types worldwide (1). Every
year, a large number of cancer-associated mortalities are induced
by HCC (2). Despite certain
advances in HCC therapy being achieved, the outcomes of patients
with HCC remain unsatisfactory (3). A number of patients with HCC are
diagnosed at the advanced stage, which makes curative treatment is
no longer possible and increases the probability of cancer
recurrence and metastasis (4).
Therefore, in order to better treat HCC, it is vital to identify
novel biomarkers for HCC diagnosis and prognosis, and therapeutic
targets.
Long non-coding RNAs (lncRNAs) are a class of
noncoding RNAs with a length >200 nucleotides, which possess no
protein-coding potential (5). In
recent years, lncRNA has attracted much attention in the field of
biology. Increasing evidence demonstrated that lncRNA exerts pivot
functions in the majority of biological processes, including cell
survival, proliferation, migration and invasion (6–8).
Dysregulation of lncRNAs is closely associated with cancer
development and progression (9).
For instance, the lncRNA Sox2ot is overexpressed in
cholangiocarcinoma and promotes tumor cell proliferation and
invasion (10). Zhang et al
(11) reported that lncRNA
HOXD-AS1 promotes epithelial ovarian cancer cell proliferation and
invasion by targeting microRNA (miRNA/miR)-133a-3p and activating
the Wnt/β-catenin signaling pathway. Therefore, it is crucial to
determine the mechanism of lncRNAs in tumor progression.
FEZF1-AS1 has been reported to regulate tumor
progression in a number of cancer types, including colorectal
carcinoma (12), gastric cancer
(13) and non-small cell lung
cancer (14). However, whether
FEZF1-AS1 serves a role in HCC requires investigation. In the
present study, it was identified that FEZF1-AS1 was significantly
upregulated in HCC tissues and predicted a poor prognosis for
patients with HCC. It was demonstrated that knockdown of FEZF1-AS1
inhibited the proliferation, migration and invasion of HCC cells.
Additionally, it was identified that FEZF1-AS1 acted as a sponge to
miR-4443, which was significantly downregulated in HCC tissues.
Furthermore, it was identified that inhibition of miR-4443
abolished the effects of FEZF1-AS1 on HCC cell proliferation,
migration and invasion. Collectively, the present results
demonstrated that FEZF1-AS1 serves as an oncogene in HCC via
inhibition of miR-4443.
Materials and methods
Patient samples
A total of 116 specimens, including 58
tumor-adjacent tissues and 58 tumour tissues (female, 11 and male,
47; mean age, 49.16±13.42 years), were obtained from patients with
HCC, who underwent surgical resection from August 2010 to October
2016 at The First College of Clinical Medical Science, China Three
Gorges University (Yichang, China). Patients who received
chemotherapy or radiotherapy prior to surgery were excluded. The
final diagnosis was confirmed by pathological analysis. All the
specimens were collected immediately following liver resection and
stored in liquid nitrogen at −80°C until analysis. Written consent
was obtained from every patient and the research protocol was
approved by the Ethics Committee of The First College of Clinical
Medical Science, China Three Gorges University.
Cell culture and transfection
HCC cell lines, Hep3B and Huh7, and normal
hepatocyte LO2 were all purchased from the Cell Bank of the Chinese
Academy of Sciences (Shanghai, China). In addition, all the cell
lines were cultured with their specified basic culture medium
[Dulbecco's modified Eagle's medium (DMEM); Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA] supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin sulfate and maintained at
37°C in a humidified atmosphere containing 5% CO2.
FEZF1-AS1 small interfering (si)RNA
(5′-GAAAGUGUUGUGUCAAUAACG-3′) and non-targeting siRNA [si negative
control (siNC, 5′-AATTCTCCGAACGTGTCACGT-3′)], miR-4443 mimics
(5′-UUGGAGGCGUGGGUUUU-3′), inhibitors (5′-AAAACCCACGCCUCCAA-3′) and
controls (5′-ACAUCUGCGUAAGAUUCGAGUCUA-3′) were purchased from
Shanghai Integrated Biotech Solutions Co., Ltd. (Shanghai, China).
Transfection was performed with Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
according to the manufacturer's protocol, with plasmids or siRNAs
transfected at a concentration of 50 nM. A total of 48 h
post-transfection, efficiency was validated by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues or cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturers' protocol, and subsequently
converted into complementary DNA (cDNA) using a PrimeScript RT
reagent kit (Takara Biotechnology Co., Ltd., Dalian, China),
according to the manufacturer's protocol. The RNA expression levels
were examined by real-time PCR using a SYBR Premix Dimmer Eraser
kit (Takara Biotechnology Co., Ltd.). The thermocycling conditions
of qPCR were as follows: 94°C for 15 min, followed by 45 cycles at
94°C for 10 sec, 60°C for 30 sec and 72°C for 30 sec. Gene
expression in each sample was normalized to U6. The expression of
miR-4443 was quantified using TaqMan MicroRNA Assay kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.), and human U6 RNA,
which was amplified as a control. Data are presented as the mean ±
standard deviation from three independent experiments. The relative
expression fold-change of mRNA was calculated using the
2−ΔΔCq method (15).
The primer sequences were as follows: FEZF1-AS1, forward
5′-TTAGGAGGCTTGTTCTGTGT-3′, reverse 5′-GCGCAGGTACTTAAGAAAGA-3′;
miR-4443, forward 5′-GTTGGAGGCGTGGGT-3′, reverse
5′-GGTCCAGTTTTTTTTTTTTTTTAAAACC-3′; and U6, forward
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
RNA-FISH assay
Fluorescence-conjugated FEZF1-AS1 probes for
RNA-FISH were generated, according to protocols of LGC Biosearch
Technologies (Petaluma, CA, USA). HCC cells were treated in a
non-denaturing condition, followed by hybridization with DNA probe
sets, as previously described (9).
DAPI (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used to
stain the nuclei at room temperature for 10 min. Treated samples
were visualized by confocal microscopy at ×600 magnification
(FV1000; Olympus Corporation, Tokyo, Japan).
Cellular proliferation assays
Cells were seeded into 96-well plates at a density
of 1×103 cells/well. Following 24, 48, 72 and 96 h of
incubation at 37°C in DMEM (Gibco; Thermo Fisher Scientific, Inc.),
cellular viability was evaluated using a Cell Counting Kit-8
(CCK-8) assay (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan), according to the manufacturer's protocol. The absorbance
was measured at a wavelength of 450 nm using a
Multiskan™ GO Microplate Spectrophotometer (Thermo
Fisher Scientific, Inc.).
A colony formation assay was also performed. A total
of 2×103 cells/well were seeded into 6-well plates with
DMEM supplemented with 10% FBS and cultured at 37°C with 5%
CO2 for 14 days. Colonies were fixed with 100% methanol
at room temperature for 20 min and stained with 0.1% crystal violet
(Sigma-Aldrich; Merck KGaA) at 25°C for 30 min. The total number of
visible colonies was determined under an optical light microscope
at ×40 magnification (Olympus Corporation). All experiments were
repeated three times.
Cellular migration and invasion
assays
Transwell migration and matrigel invasion assays
were assessed using transwell membranes (8 µm pore size, 6.5 mm
diameter; Corning Incorporated, Corning, NY, USA) in 24-well plates
of 8 µm (BD Biosciences, Franklin Lakes, NY, USA) according to the
manufacturer's protocol. For the transwell migration assay, cells
were resuspended in serum-free 200 µl DMEM medium at a density of
5×104 cells/well. The bottom chamber of transwell plates
was supplemented with 600 µl DMEM medium, containing 20% FBS as a
chemoattractant. Cells were subsequently seeded into the upper
chamber and incubated for another 20 h at 37°C with 5%
CO2. For the invasion assay, matrigel matrix (1:8; 50
µl/well, BD Biosciences) was polymerized in the transwell membrane,
according to the manufacturer's protocol. Cells (1×105
cells/well) were used and incubated for another 48 h at 37°C with
5% CO2. Following incubation, cells on the upper surface
of the membrane were scraped off with cotton swabs. Cells that
migrated to the lower surface were fixed with polyoxymethylene at
25°C for 30 min, stained with 0.1% Crystal Violet staining solution
at 25°C for 30 min. The cells on the bottom of the membrane were
calculated from five random light microscopic fields at ×40
magnification.
Luciferase assays
The potential targets of FEZF1-AS1 were predicted
using the miRDB tool (http://mirdb.org/miRDB/index.html). Subsequently,
FEZF1-AS1 sequences containing the wild-type (WT) binding site or
mutated-type (Mut) binding site for miR-4443 were synthesized by
Sangon Biotech Co., Ltd. (Shanghai, China) and cloned into the
pmiR-GLO vector (Promega Corporation, Madison, WI, USA). Luciferase
reporter gene assays were performed using the Dual-Luciferase
Reporter Assay System (Promega Corporation) according to the
manufacturer's protocol. Prior to transfection, cells were seeded
in 24-well plates (5×103 cells/well) and cultured for 24
h. Subsequently, the WT or Mut of FEZF1-AS1 was transiently
co-transfected with miR-4443 mimics or miR-NC (Guangzhou RiboBio
Co., Ltd., Guangzhou, China) into logarithmic phase cells using
Lipofectamine® 2000 transfection reagent. Luciferase
assays were performed using the Dual-Luciferase Reporter Assay
system (Promega Corporation), according to the manufacturer's
protocol, after a further 48 h culture. The value of relative
luciferase activity indicates the firefly luciferase activity
normalized to that of Renilla for each assay.
Statistical analysis
The statistical significance of the differences
between groups was assessed using Student's t-test for pair-wise
comparisons or one-way analysis of variance followed by Fisher's
least significant difference post hoc test for multiple
comparisons. Pearson's correlation coefficient analysis was used to
determine the correlations. Kaplan-Meier survival curve analysis
and log-rank test were used for survival analysis. P<0.05 was
considered to indicate a statistically significant difference. Data
are presented as the mean ± standard deviation of three independent
experiments. Statistical analysis was performedv using the SPSS
software version 20.0 (IBM Corp., Armonk, NY, USA).
Results
FEZF1-AS1 is upregulated in HCC
tissues
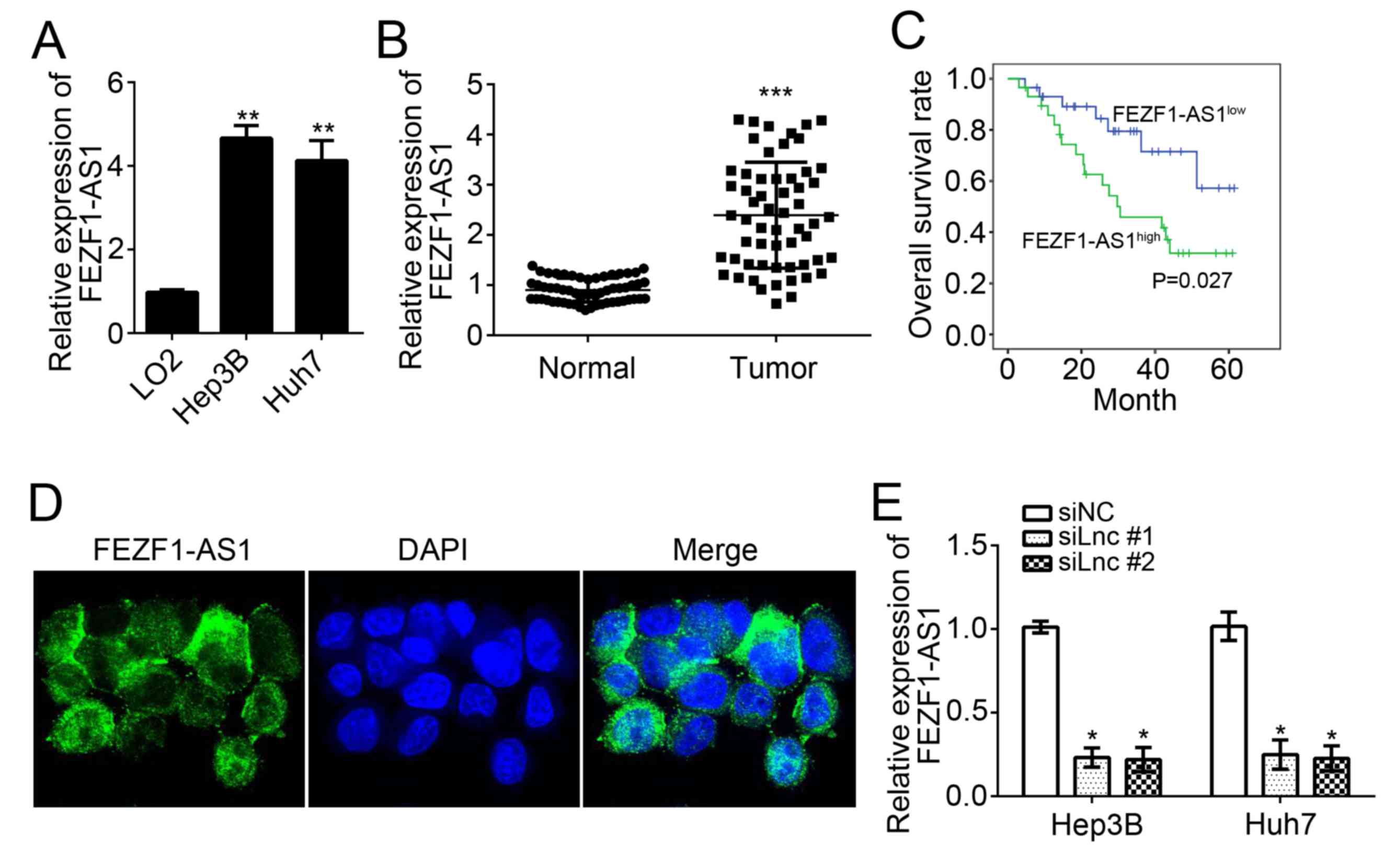
To examine the expression patterns of FEZF1-AS1,
RT-qPCR analysis was conducted using HCC cell lines and LO2 cells.
It was identified that FEZF1-AS1 expression was significantly
upregulated in HCC cell lines (Hep3B and Huh7) compared with normal
LO2 hepatocytes (P<0.01; Fig.
1A). To further verify its overexpression in HCC cells, 58 HCC
tissues and 58 adjacent normal tissues were obtained, and FEZF1-AS1
expression was analyzed using RT-qPCR. The results suggested that
FEZF1-AS1 expression levels were significantly upregulated in HCC
tissues compared with adjacent normal tissues (P<0.001; Fig. 1B). To determine whether FEZF1-AS1
is able to serve as a prognostic biomarker of patients with HCC, a
Kaplan-Meier survival curve analysis was conducted. It was
identified that patients with HCC with higher expression of
FEZF1-AS1 exhibited a significantly lower survival rate (P<0.05;
Fig. 1C). The location of
FEZF1-AS1 in HCC cells was analyzed and it was identified that
FEZF1-AS1 was located in the cytoplasm (Fig. 1D). FEZF1-AS1 expression was
subsequently knocked down in Hep3B and Huh7 cells by transfection
with specific RNAs. As presented, the expression levels of
FEZF1-AS1 were significantly downregulated in Hep3B and Huh7 cells
following transfection with the two siRNAs (P<0.05; Fig. 1E).
FEZF1-AS1 knockdown inhibits the
proliferation, migration and invasion of Hep3B and Huh7 cells
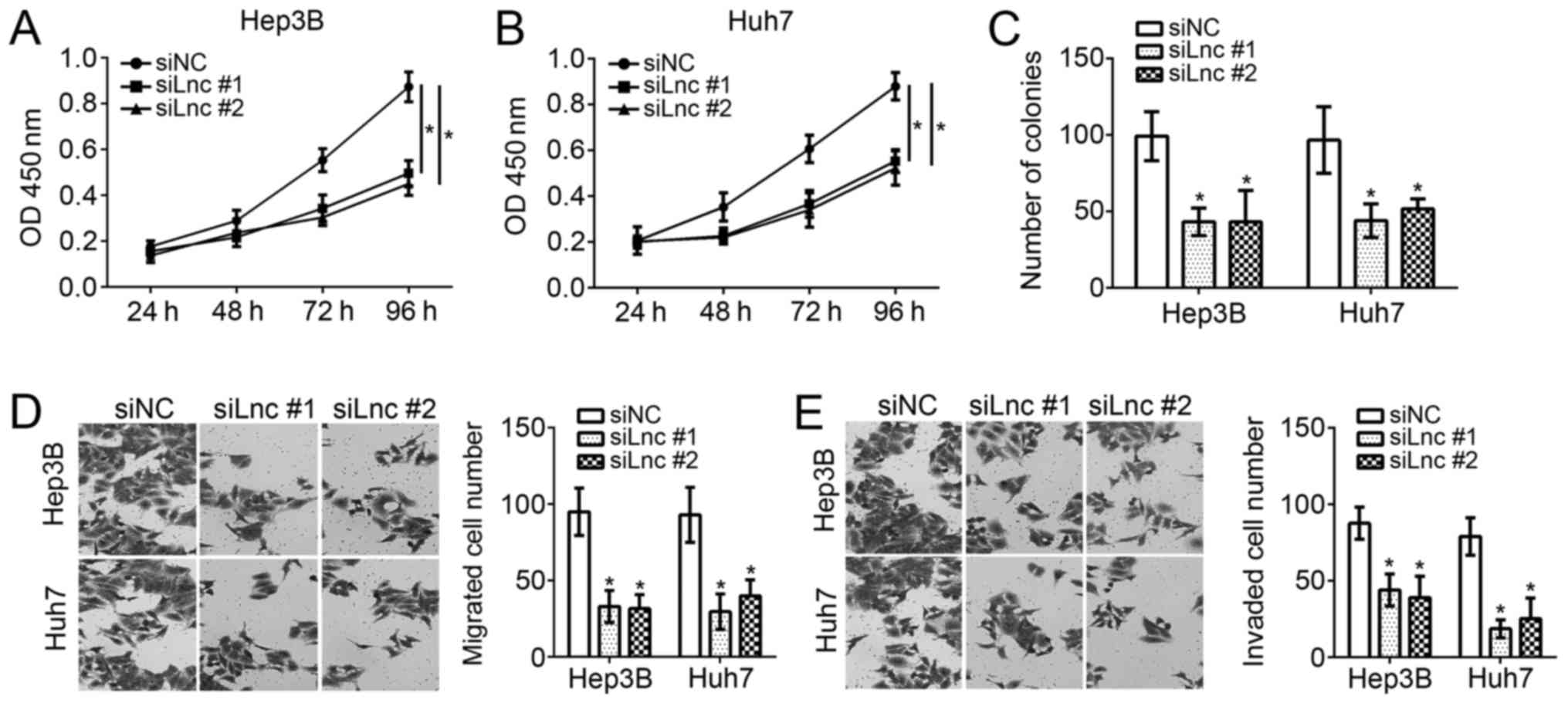
To examine the function of FEZF1-AS1 in HCC cells,
CCK-8, colony formation and transwell assays were performed.
Through CCK-8 assays, it was observed that knockdown of FEZF1-AS1
significantly inhibited the proliferation of Hep3B and Huh7 cells
(Fig. 2A and B) at 96 h after
culture. The colony formation assays suggested that the
downregulation of FEZF1-AS1 significantly decreased the number of
colonies formed by Hep3B and Huh7 cells (P<0.05; Fig. 2C). Furthermore, it was identified
that FEZF1-AS1 depletion significantly decreased the numbers of
migrated and invaded Hep3B and Huh7 cells (P<0.05; Fig. 2D and E). Collectively, these data
demonstrated that FEZF1-AS1 is important for the proliferation,
migration and invasion of HCC cells.
FEZF1-AS1 acts as a sponge for
miR-4443
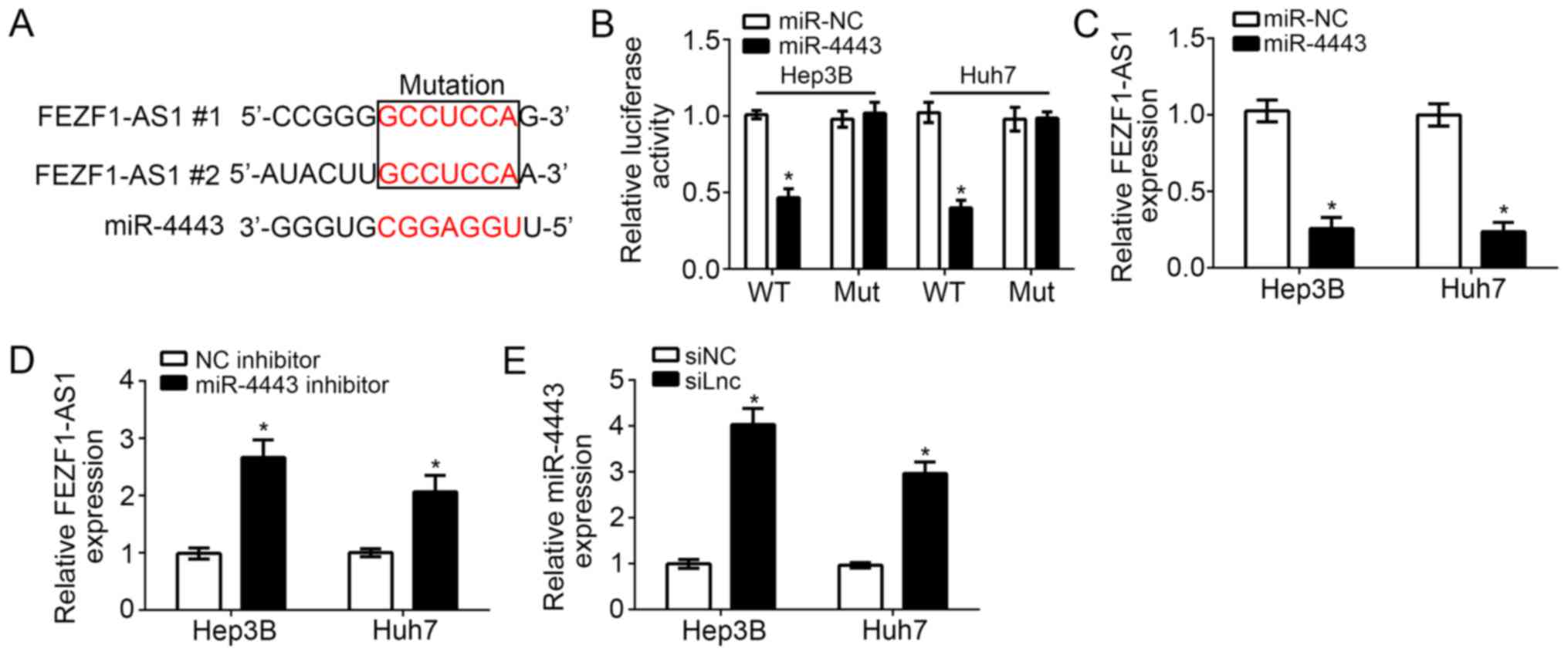
lncRNAs serve as competing endogenous RNAs (ceRNAs)
to bind with miRs and regulate gene expression (16). In order to investigate the
downstream mechanism of FEZF1-AS1, bioinformatics analysis was
performed, which identified that numerous miRs exhibited potential
binding sites for FEZF1-AS1. Among all targeting miRs, miR-4443
ranked top and has been reported to inhibit colon cancer
progression (17). Therefore,
miR-4443 was selected for further examination. It was identified
that there were two potential binding sites of miR-4443 in
FEZF1-AS1 (Fig. 3A). Through
luciferase reporter assays, it was demonstrated that miR-4443
overexpression inhibited the luciferase activity in Hep3B and Huh7
cells transfected with FEZF1-AS1-WT, while mutation of the binding
sites in FEZF1-AS1 abrogated this effect (Fig. 3B). Furthermore, it was identified
that overexpression of miR-4443 significantly inhibited the
expression levels of FEZF1-AS1 in Hep3B and Huh7 cells (P<0.01;
Fig. 3C); whereas, inhibition of
miR-4443 significantly increased the expression of FEZF1-AS1
(P<0.05; Fig. 3D).
Consistently, knockdown of FEZF1-AS1 additionally significantly
upregulated the expression levels of miR-4443 in Hep3B and Huh7
cells (P<0.05; Fig. 3E).
Collectively, these data suggested that FEZF1-AS1 directly targets
miR-4443 in HCC cells.
miR-4443 is downregulated in HCC
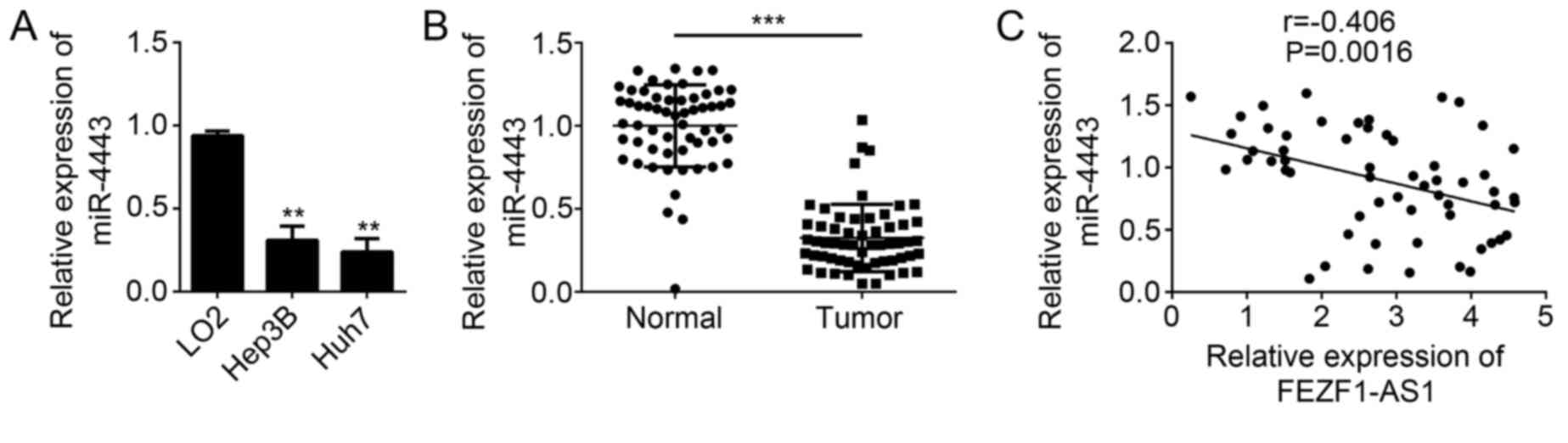
The function of miR-4443 remains unknown in HCC. To
further determine whether miR-4443 is involved in HCC progression,
the expression patterns of miR-4443 in HCC cell lines were first
analyzed by RT-qPCR. The results suggested that miR-4443 was
downregulated in HCC cell lines compared with LO2 cells (Fig. 4A). Similarly, the expression of
miR-4443 was significantly downregulated in HCC tissues compared
with adjacent normal tissues (P<0.001; Fig. 4B). Subsequently, the expression
correlation between FEZF1-AS1 and miR-4443 was determined in HCC
tissues by RT-qPCR. The results demonstrated that FEZF1-AS1
expression was negatively correlated with that of miR-4443 in HCC
tissues (Fig. 4C). In summary,
these data implied that miR-4443 may serve a function in HCC
progression.
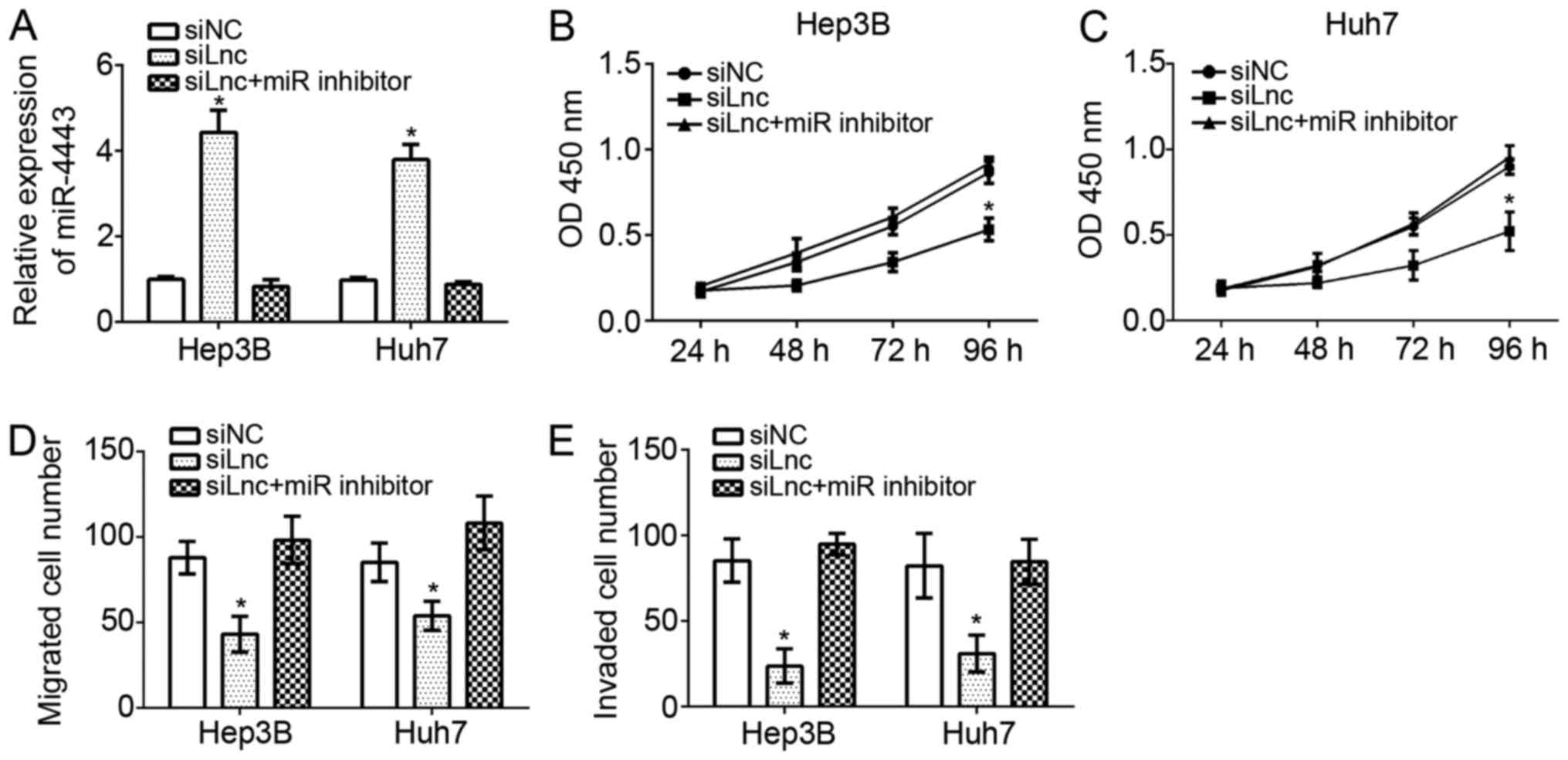
FEZF1-AS1 promotes HCC cell
proliferation, migration and invasion by regulating miR-4443
To verify whether miR-4443 was involved in
FEZF1-AS1-mediated effects on HCC progression, the expression
levels of miR-4443 were demonstrated to be downregulated by RT-qPCR
in FEZF1-AS1-depleted Hep3B and Huh7 cells. The results suggested
that the miR-4443 expression level was markedly downregulated in
Hep3B and Huh7 cells transfected siFEZF1-AS1 plus miR-4443
inhibitor compared with the siFEZF1-AS1 group (Fig. 5A). CCK-8 assays were performed to
evaluate the proliferation of Hep3B and Huh7 cells. It was
identified that FEZF1-AS1 knockdown inhibited cellular
proliferation while inhibition of miR-4443 significantly reversed
this trend in Hep3B and Huh7 cells (P<0.05; Fig. 5B and C). Furthermore, transwell
assays additionally demonstrated that FEZF1-AS1 depletion decreased
the migration and invasion of Hep3B and Huh7 cells; whereas,
knockdown of miR-4443 at the simultaneously promoted cell migration
and invasion (Fig. 5D and E).
FEZF1-AS1 promoted HCC cell proliferation, migration and invasion
by acting as a sponge to miR-4443.
Discussion
HCC contributes to a large number of
cancer-associated mortalities around the world each year (2). Although HCC becomes a primary public
health problem, there are no effective therapeutic methods for
curative treatment of HCC. Therefore the examination of the
underlying mechanism of hepatocarcinogenesis and the search for
novel therapeutic targets for HCC treatment is urgently required.
In the present study, it was identified that FEZF1-AS1 was highly
expressed in HCC tissues and cell lines compared with adjacent
normal tissues or hepatocytes. Additionally, it was demonstrated
that FEZF1-AS1 expression serves as a prognostic marker for
patients with HCC. Furthermore, the present results suggested that
knockdown of FEZF1-AS1 suppressed the proliferation, migration and
invasion of Hep3B and Huh7 cells in vitro. In terms of
mechanism, it was observed that FEZF1-AS1 served as a ceRNA sponge
to miR-4443 in HCC cells. Overexpression of miR-4443 repressed the
expression of FEZF1-AS1 in HCC cells, and vice versa. Furthermore,
it was demonstrated that miR-4443 was downregulated in HCC tissues
and negatively correlated with the expression of FEZF1-AS1.
Finally, through functional experiments, it was demonstrated that
inhibition of miR-4443 abrogated the effects of FEZF1-AS1 on HCC
cell proliferation, migration and invasion.
In recent years, an increasing number of studies
document that lncRNAs are essential regulator in various biological
processes (18–20). In human cancer, lncRNAs have been
demonstrated to serve pivotal roles in regulating cellular
proliferation, apoptosis, migration and invasion (19). Increasing evidence suggests that
lncRNAs are involved in the development and progression of a
diverse range of cancer types, including osteosarcoma (20), gastric cancer (21), non-small cell lung cancer (22), colorectal cancer (23), breast cancer (24), clear cell renal cell carcinoma
(25) and hepatocellular carcinoma
(26). However, the functions of
the majority of lncRNAs in cancer remain unclear. As for FEZF1-AS1,
a previous study suggested that FEZF1-AS1 facilitates cell
proliferation and migration in colorectal carcinoma (12). Another previous study reported that
FEZF1-AS1 represses p21 expression to promote gastric cancer
proliferation through LSD1-Mediated H3K4me2 demethylation (13). He et al (14) demonstrated that FEZF1-AS1 enhances
epithelial-mesenchymal transition by suppressing
epithelial-cadherin and regulating WNT pathway in non-small cell
lung cancer. Whether FEZF1-AS1 regulates HCC progression remains
largely unknown. In the present study, it was observed that
FEZF1-AS1 was upregulated in HCC cells and promoted cancer cell
proliferation, migration and invasion. The present results
suggested that FEZF1-AS1 additionally serves as an oncogene in
HCC.
In the past decades, the functions of miRNAs have
been widely investigated. A large number of studies suggest that
miRNAs are closely associated with tumorigenesis. Additionally,
miRNAs are demonstrated to regulate tumor cell proliferation,
migration and invasion by suppressing target gene expression
post-transcriptionally (27).
Abnormal expression of miRNAs is frequently observed in types of
cancer, including breast cancer (28), colorectal cancer (29), non-small cell lung cancer (30) and hepatocellular carcinoma
(31). Recently, a previous study
suggested that miRNA expression levels may be regulated by lncRNAs
(18). In the present study,
FEZF1-AS1 was identified to act as a sponge of miR-4443 in HCC
cells. miR-4443 has been demonstrated to decrease the invasiveness
of human colon cancer cells (17).
In contrast, another previous study suggested that miR-4443 induced
malignancy of breast cancer (32).
Whether miR-4443 is involved in HCC requires further investigation.
Through functional experiments, it was observed that miR-4443 was
downregulated in HCC tissues. Inhibition of miR-4443 abolished the
effects of FEZF1-AS1 on HCC cell proliferation, migration and
invasion, which suggested that miR-4443 suppressed HCC
progression.
In conclusion, the results of the present study
demonstrated that FEZF1-AS1 promoted the proliferation, migration
and invasion of HCC cells by regulating miR-4443 expression levels.
The present results highlight the importance of the
FEZF1-AS1/miR-4443 axis during HCC development and progression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JG and JW conceived and designed the study, and
analyzed and interpreted the results. JZ performed experiments and
wrote this manuscript. TL and JH conducted the experiments. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
For the use of human samples, the protocol for the
present study was approved by the Institutional Ethics Committee of
China Three Gorges University (Yichang, China) and all enrolled
patients signed a written informed consent document.
Patient consent for publication
All patients within the present study provide
consent for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Pisani P and Ferlay J: Global
cancer statistics. CA Cancer J Clin. 49:33–64. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Colecchia A, Schiumerini R, Cucchetti A,
Cescon M, Taddia M, Marasco G and Festi D: Prognostic factors for
hepatocellular carcinoma recurrence. World J Gastroenterol.
20:5935–5950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pan Y, Qin T, Yin S, Zhang X, Gao X and Mu
L: Long non-coding RNA UC001kfo promotes hepatocellular carcinoma
proliferation and metastasis by targeting α-SMA. Biomed
Pharmacother. 87:669–677. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu L, Dai Z, Luo Q and Lv G: The long
noncoding RNA cancer susceptibility candidate 2 inhibits tumor
progression in osteosarcoma. Mol Med Rep. 17:1947–1953.
2018.PubMed/NCBI
|
|
6
|
Chen Y, Wei G, Xia H, Tang Q and Bi F:
Long noncoding RNA-ATB promotes cell proliferation, migration and
invasion in gastric cancer. Mol Med Rep. 17:1940–1946.
2018.PubMed/NCBI
|
|
7
|
Tang WG, Hu B, Sun HX, Sun QM, Sun C, Fu
PY, Yang ZF, Zhang X, Zhou CH, Fan J, et al: Long non-coding
RNA00364 represses hepatocellular carcinoma cell proliferation via
modulating p-STAT3-IFIT2 signaling axis. Oncotarget.
8:102006–102019. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shi J, Wang YJ, Sun CR, Qin B, Zhang Y and
Chen G: Long noncoding RNA lncHERG promotes cell proliferation,
migration and invasion in glioblastoma. Oncotarget.
8:108031–108041. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu P, Wang Y, Wu J, Huang G, Liu B, Ye B,
Du Y, Gao G, Tian Y, He L and Fan Z: LncBRM initiates YAP1
signalling activation to drive self-renewal of liver cancer stem
cells. Nat Commun. 7:136082016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Z, Li J, Ji D, Leng K, Xu Y, Huang L,
Jiang X and Cui Y: Overexpressed long noncoding RNA Sox2ot predicts
poor prognosis for cholangiocarcinoma and promotes cell
proliferation and invasion. Gene. 645:131–136. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Dun Y, Zhou S and Huang XH:
LncRNA HOXD-AS1 promotes epithelial ovarian cancer cells
proliferation and invasion by targeting miR-133a-3p and activating
Wnt/β-catenin signaling pathway. Biomed Pharmacother. 96:1216–1221.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen N, Guo D, Xu Q, Yang M, Wang D, Peng
M, Ding Y, Wang S and Zhou J: Long non-coding RNA FEZF1-AS1
facilitates cell proliferation and migration in colorectal
carcinoma. Oncotarget. 7:11271–11283. 2016.PubMed/NCBI
|
|
13
|
Liu YW, Xia R, Lu K, Xie M, Yang F, Sun M,
De W, Wang C and Ji G: LincRNAFEZF1-AS1 represses p21 expression to
promote gastric cancer proliferation through LSD1-Mediated H3K4me2
demethylation. Mol Cancer. 16:392017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He R, Zhang FH and Shen N: LncRNA
FEZF1-AS1 enhances epithelial-mesenchymal transition (EMT) through
suppressing E-cadherin and regulating WNT pathway in non-small cell
lung cancer (NSCLC). Biomed Pharmacother. 95:331–338. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao C, Zhang T, Zhang D, Xie L, Zou X, Lei
L, Wu D and Liu L: The long non-coding RNA, SNHG6-003, functions as
a competing endogenous RNA to promote the progression of
hepatocellular carcinoma. Oncogene. 36:1112–1122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meerson A and Yehuda H: Leptin and insulin
up-regulate miR-4443 to suppress NCOA1 and TRAF4, and decrease the
invasiveness of human colon cancer cells. BMC Cancer. 16:8822016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng P, Yin Z, Wu Y, Xu Y, Luo Y and
Zhang TC: LncRNA HOTAIR promotes cell migration and invasion by
regulating MKL1 via inhibition miR206 expression in HeLa cells.
Cell Commun Signal. 16:52018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li W, Zhao W, Lu Z, Zhang W and Yang X:
Long noncoding RNA GAS5 promotes proliferation, migration, and
invasion by regulation of miR-301a in esophageal cancer. Oncol Res.
26:1285–1294. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang C, Wu K, Wang S and Wei G: Long
non-coding RNA XIST promotes osteosarcoma progression by targeting
YAP via miR-195-5p. J Cell Biochem. 119:5646–5656. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Wang Y, Sun L, Min J, Liu J, Chen
D, Zhang H, Zhang H, Zhang H, Zhou Y and Liu L: Long noncoding RNA
BC005927 upregulates EPHB4 and promotes gastric cancer metastasis
under hypoxia. Cancer Sci. 109:988–1000. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liang Y, Ma Y, Li L, Shen X, Xin T, Zhao Y
and Ma R: Effect of long non-coding RNA LINC01116 on biological
behaviors of non-small cell lung cancer cells via the hippo
signaling pathway. J Cell Biochem. 119:63102018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Tao Y, Li Y, Zhao J, Zhang L,
Zhang X, Dong C, Xie Y, Dai X, Zhang X and Liao Q: The regulatory
network analysis of long noncoding RNAs in human colorectal cancer.
Funct Integr Genomics. 18:261–275. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang S, Wang J, Ghoshal T, Wilkins D, Mo
YY, Chen Y and Zhou Y: lncRNA gene signatures for prediction of
breast cancer intrinsic subtypes and prognosis. Genes (Basel).
9:pii: E65. 2018. View Article : Google Scholar
|
|
25
|
Wu Y, Tan C, Weng WW, Deng Y, Zhang QY,
Yang XQ, Gan HL, Wang T, Zhang PP, Xu MD, et al: Long non-coding
RNA Linc00152 is a positive prognostic factor for and demonstrates
malignant biological behavior in clear cell renal cell carcinoma.
Am J Cancer Res. 6:285–299. 2016.PubMed/NCBI
|
|
26
|
Ma X, Liu H, Murphy JT, Foyil SR, Godar
RJ, Abuirqeba H, Weinheimer CJ, Barger PM and Diwan A: Regulation
of the transcription factor EB-PGC1α axis by beclin-1 controls
mitochondrial quality and cardiomyocyte death under stress. Mol
Cell Biol. 35:956–976. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Wang W, Huang K, Wang Y, Li J and
Yang X: MicroRNA-34a inhibits cells proliferation and invasion by
downregulating Notch1 in endometrial cancer. Oncotarget.
8:111258–111270. 2017.PubMed/NCBI
|
|
28
|
Xiong H, Yan T, Zhang W, Shi F, Jiang X,
Wang X, Li S, Chen Y, Chen C and Zhu Y: miR-613 inhibits cell
migration and invasion by downregulating Daam1 in triple-negative
breast cancer. Cell Signal. 44:33–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tong F, Ying Y, Pan H, Zhao W, Li H and
Zhan X: MicroRNA-466 (miR-466) functions as a tumor suppressor and
prognostic factor in colorectal cancer (CRC). Bosn J Basic Med Sci.
18:252–259. 2018.PubMed/NCBI
|
|
30
|
Jiang H, Zhang H, Hu X and Li W: Knockdown
of long non-coding RNA XIST inhibits cell viability and invasion by
regulating miR-137/PXN axis in non-small cell lung cancer. Int J
Biol Macromol. 111:623–631. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cui S, Qian Z, Chen Y, Li L, Li P and Ding
H: Screening of up- and downregulation of circRNAs in HBV-related
hepatocellular carcinoma by microarray. Oncol Lett. 15:423–432.
2018.PubMed/NCBI
|
|
32
|
Chen X, Zhong SL, Lu P, Wang DD, Zhou SY,
Yang SJ, Shen HY, Zhang L, Zhang XH, Zhao JH and Tang JH: miR-4443
participates in the malignancy of breast cancer. PLoS One.
11:e01607802016. View Article : Google Scholar : PubMed/NCBI
|