Introduction
Neonatal hypoxic-ischemic encephalopathy (HIE) is a
neurological condition in newborns characterized by hypoxia and
ischemia, and is a major cause of neonatal mortality, neurological
behavior deficient and long-term disability (1). Not only are patients in distress, an
enormous burden and pressure is placed upon parents, and the rest
of society (2). Therefore, there
is an urgent need to identify effective treatments for HIE in
neonates. Furthermore, elucidating the mechanisms underlying HIE is
required (3). Energy failure,
intracellular calcium overload, glutamate-mediated excitotoxicity,
oxidative stress and inflammation have all been reported to
contribute to HIE (4–6). In the current study, the association
between the expression of ubiquilin-1 (Ubqln) and the extent of
neonatal HI brain injury was investigated.
Abnormal protein aggregation, and intracellular or
extracellular accumulation of misfolded and aggregated proteins are
major events in the pathogenesis of different neurodegenerative
diseases. The ubiquitin-proteasome system has a key role in
protecting neuronal homeostasis by removing misfolded/aggregating
proteins (7). Ubqln1, also known
as proepithelin, is a ubiquitin-like (UbL) protein, including a
N-terminal UbL domain, which regulates the interaction with the
proteasome and a C-terminal Ub-associated domain and preferentially
binds poly-ubiquitinated proteins (8). Previous studies have demonstrated
that Ubqln overexpression promotes the degradation of misfolded
proteins, inhibits misfolded protein-induced cytotoxicity, and
protects neurons against ischemia and oxidative stress-induced
brain injury; whereas knockdown of Ubqln aggravates cerebral
ischemia-induced neuronal injury and delays nerve function recovery
(7,8). Disruption of Ubqln function is
involved in the pathologic process of a number of human
neurodegenerative disorders, such as Alzheimer's disease (9,10)
and Huntington's disease (11);
however, the precise location and distribution of Ubqln in neonatal
HIE remains largely unknown. In the current study, the distribution
and co-localization of Ubqln in brain tissue was analyzed using
immunohistochemical methods. The study determined the level of
Ubqln during the development of neonatal hypoxic-ischemic (HI)
brain injury.
Materials and methods
Animals
Timed-pregnant C57 mice (n=5; age, 2–3-months) were
acquired from Sun Yat-sen University (Guangdong, China). The day of
birth of pups was designated day 0 (P0); postnatal 7-day-old (P7)
pups of either sex were used in the subsequent experiments. A total
of 40 pups were used in this study. All animal-related experiments
were approved and organized in accordance with the guidelines of
the Experimental Laboratory Animal Committee of Guangdong
Pharmaceutical University (permit no: gdpulac2017175), and under
the principles of the National Institutes of Health Guide for the
Care and Use of Laboratory (12).
The animals were housed under controlled temperature (23±2°C),
humidity (55±0%) and lighting conditions (12-h light/dark cycle).
Water and food were provided ad libitum.
HI brain injury model
Pups were divided into two groups (sham control and
HI). A HI brain injury model was established in P7 pups using the
Rice-Vannucci method (13), with
some modifications. Briefly, P7 pups weighing 5–5.5 g were
anesthetized with a 3% isoflurane-oxygen mixture for induction and
2% for maintenance. In pups subjected to the HI model, the left
common carotid artery (CCA) was permanently cut off using a bipolar
electrocoagulation device (gutta cutter, Jiangsu Kanghua Medical
Equipment Co., Ltd., Jiangsu, China). Pups were then transferred to
a 37°C incubator for 10 min until the pups regained consciousness,
and were then returned to their dams for 90 min. Subsequently, the
pups were placed in a hypoxia chamber containing 8% O2
in mixture with 92% N2 for 120 min. Sham controls
underwent anesthesia and the left CCA was exposed as in the HI
group, but there was no ligation or exposure to hypoxia.
Triphenyl tetrazolium chloride (TTC)
staining
At 24 h after completion of occlusion and hypoxic
injury, pups were sacrificed, and the whole brains of pups in the
HI and sham groups were quickly collected and sectioned coronally
into 2-mm slices for TTC staining. Tissue slices were stained with
2% TTC solution (cat. no. 17779; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) in a dark incubator at 37°C for 20 min. The
tissue slices were then placed in 4% paraformaldehyde overnight to
fix the brains for imaging. Tissue slices were imaged on each side.
Each slice of the two hemispheres of infarction area were
quantified by ImageJ (version 1.8.0; National Institutes of Health,
Bethesda, MD, USA).
Preparation of tissue sections
On the third day after surgery, the pups underwent
deep anesthesia by intraperitoneal injection of 10% chloral hydrate
(400 mg/kg; cat. no. 302-17-0; Sigma-Aldrich; Merck KGaA) and fixed
by transcardiac perfusion of cold PBS followed by ice cold 4%
paraformaldehyde in 0.1 M PBS. The brains were removed and further
fixed in the same fixative solution at 4°C overnight, and afterward
the brains were dehydrated serially in 10, 20 and 30% sucrose in
PBS at 4°C overnight until sinking occurred. Then, the brains were
implanted in Optimal Cutting Temperature compound (cat. no. 4583;
Sakura Finetek USA, Inc., Torrance, CA, USA). Serial coronal
sections were cut using a freezing microtome at 10-µm intervals and
mounted onto poly-L-lysine-coated glass slides.
Immunofluorescence staining
To detect the expression and distribution of Ubqln
in the brains of neonatal HI pups, sections were washed with PBS.
Following blocking, using blocking buffer (QuickBlock™ Blocking
Buffer for Immunol Staining, cat. no. P0260, Beyotime Institute of
Biotechnology, Shanghai, China) for 1 h at room temperature to
reduce non-specific staining. Then, the sections were incubated
with primary antibody against Ubqln (dilution, 1:500; cat. no.
16400-I-AP; Proteintech Group, Inc., Rosemont, USA.) in PBS
containing 0.3% Triton X-100 at 4°C overnight. After sufficient
washing with PBS, appropriate secondary antibodies in Alexa
Fluor®594 (1:1,000 dilution; cat. no. A-11012;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) were
added and incubated for 2 h at 37°C in the dark, and the sections
were washed three times with PBS. Nuclei were stained with the
nuclear dye, DAPI (0.1 µg/ml), for 5 min at room temperature and
fully rinsed with PBS, then the coverslips were mounted on slides
with FluorSave reagent (cat. no. P0126; Beyotime Institute of
Biotechnology) and the morphologies observed under a fluorescence
microscope (BX51; Olympus Corporation, Tokyo, Japan). Ischemic
ipsilateral cortical and hippocampal regions were selected for
imaging, and five visual fields were selected for each section
(magnification, ×100 and ×200).
Double immunofluorescent staining
The cellular location of Ubqln was also determined
in the brains of HI pups. Frozen sections were heated and washed
with PBS then incubated with blocking solution with 10% normal goat
serum for 1 h at room temperature. The sections were then incubated
with primary antibody against RNA binding protein fox-1 homolog 3
(NeuN) for neurons (dilution, 1:1,000; cat. no. SAB4300883;
Sigma-Aldrich; Merck KGaA), glial fibrillary acidic protein (GFAP)
for astrocytes (dilution, 1:1,000; cat. no. ab10062, Abcam,
Cambridge, USA), allograft inflammatory factor 1 (Iba-1) for
microglial cells (dilution, 1:1,000; cat. no. ab15690, Abcam) and
Ubqln (dilution, 1:500; cat. no. SAB1305680, Sigma-Aldrich, Merck
KGaA) in PBS containing 0.3% Triton X-100 at 4°C overnight. The
sections were then washed with PBS and incubated with the
corresponding fluorescence-conjugated secondary antibodies (Alexa
Fluor®488 goat anti-mouse IgG (H+L), cat. no. A-11029;
Alexa Fluor®594 goat anti-rabbit IgG (H+L), cat. no.
A-11012; Invitrogen; Thermo Fisher Scientific, Inc.) for 1 h at
room temperature. Nuclei were stained for DAPI (0.1 µg/ml), for 5
min at room temperature. Images were obtained using a fluorescence
microscope (BX51; Olympus Corporation). Ischemic ipsilateral
cortical and hippocampal regions were selected for imaging, and
five visual fields were selected for each section (magnification,
×100 and ×200).
Western blot analysis
Western blotting was used to determine the level of
Ubqln semi-quantitatively following HI treatment. The expression of
β-actin was designated as the internal control. For western blot
analysis, the total protein of the ipsilateral hemisphere was
removed and extracted at 1 and 3 days after HI treatment with a
Micro BCA Protein Assay kit according to the manufacturer's
instructions (cat. no. P0012S; Beyotime Institute of
Biotechnology). The bicinchoninic acid assay (Beyotime Institute of
Biotechnology) was used to measure the protein concentration, with
BSA (cat. no. P0012S, Beyotime Institute of Biotechnology) as the
standard. For each run, 20 mg protein/well lysate was separated by
SDS-PAGE on 10% gels and transferred to a polyvinylidene fluoride
membrane (EMD Millipore) The membranes were blocked in 5% non-fat
milk for 1 h at room temperature and incubated overnight in the
presence of the primary antibodies against Ubqln (1:5,000, cat. no.
16400-I-AP; Proteintech Group, Inc) and β-actin (1:10,000; cat. no.
T0022; Affinity Biosciences, Cincinnati, OH, USA) at 4°C. The
membrane was fully washed three times with TBS containing 0.05%
Tween 20 (TBST) and subsequently reacted with the corresponding
secondary antibody (1:10,000, Goat Anti-Rabbit IgG (H+L) HRP; cat.
no. S0001; Goat Anti-Mouse IgG (H+L) HRP; cat. no. S0002; both
Affinity Biosciences) for 1 h at room temperature. After thorough
washing with TBST, the protein bands were developed using enhanced
chemiluminescence detection reagents (cat. no. WBKLS0500; Merck
KGaA). The optical density of the bands on the films was analyzed
using ImageJ version 1.8.0 (National Institutes of Health).
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM Corp.). The results are expressed as the mean ±
standard error from at least three independent experiments. A
statistical evaluation was performed with a one-way analysis of
variance followed by Duncan's multiple range test, which was used
to compare the sham control and HI groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
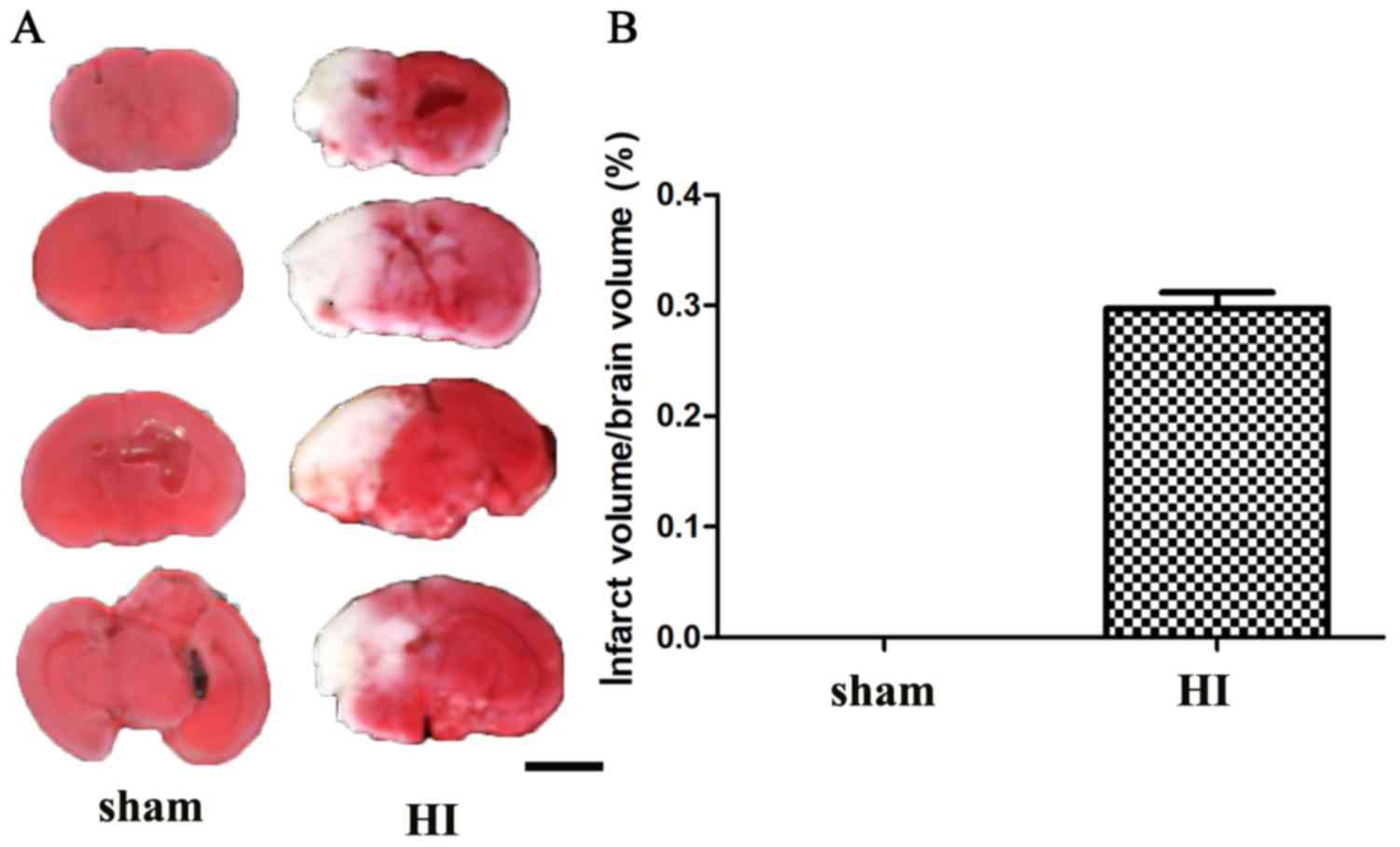
HI brain injury model
To determine the expression of Ubqln in the brains
of the HIE mouse model, a mouse pup model was established using the
Rice-Vannucci method with some modifications. At 24 h after HI
injury, the ischemic infarctions area appeared white and regularly
included the neocortex and basal ganglia, as confirmed by TTC
staining (Fig. 1). These results
suggested that the HIE model had been successfully established.
Expression of Ubqln in the brains of
the HIE mouse model
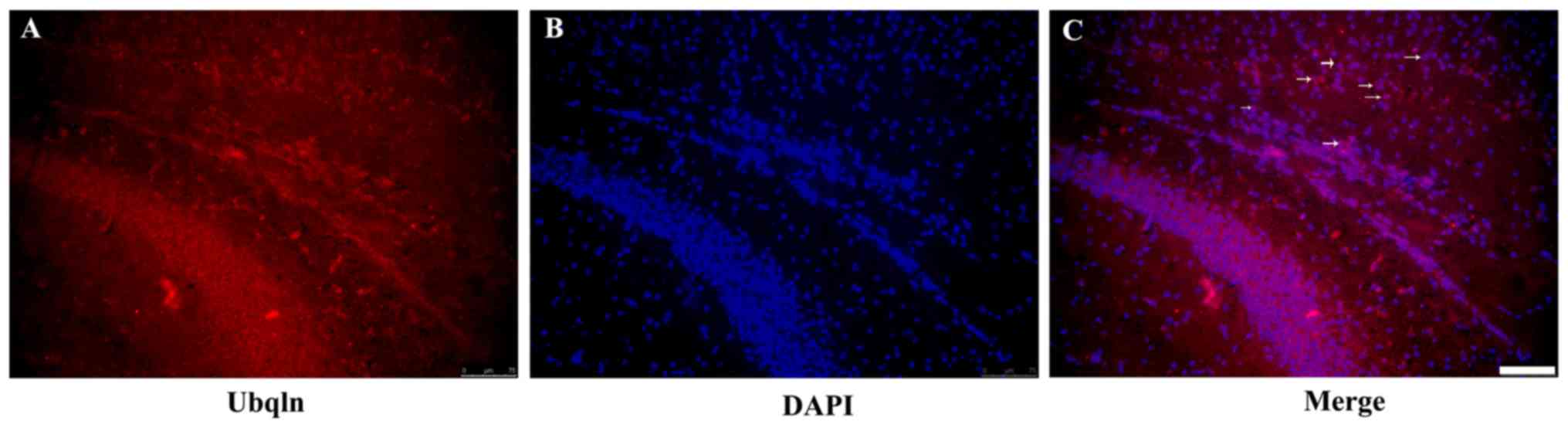
Immunofluorescence staining was performed of the
mouse brain tissues to determine the expression of Ubqln in the
brains of the HIE model mice. The results demonstrated that Ubqln
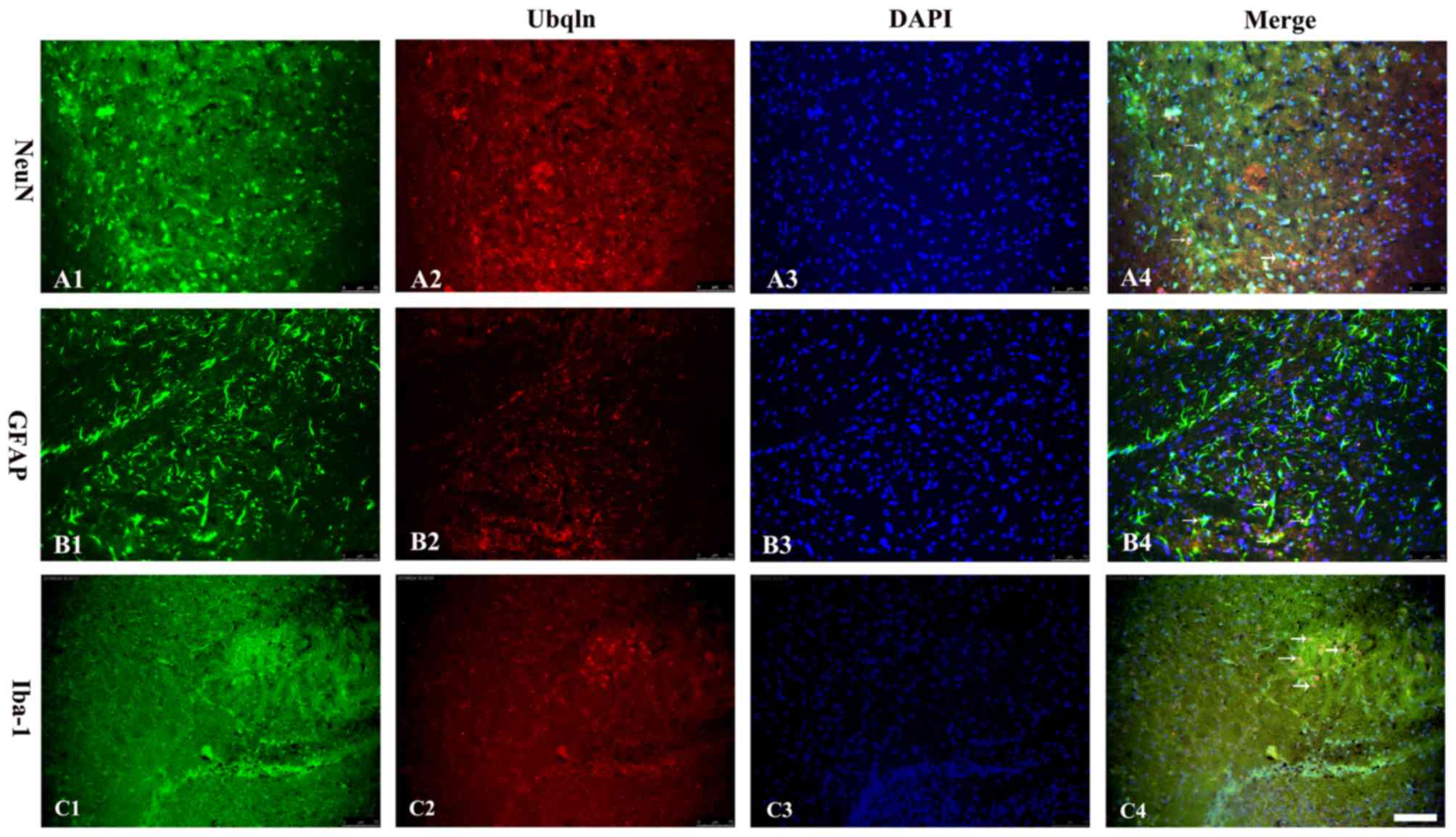
was expressed predominantly in the cortex and hippocampus (Fig. 2). To analyze the cell type-specific
expression of Ubqln in the brains of the HIE mouse model,
immunofluorescent double labeling was performed with specific
markers for neurons (NeuN), astrocytes (GFAP) and microglia
(Iba-1). The results indicated that the mature neuronal marker,
astrocyte marker and microglia marker were double-labeled with
Ubqln (Fig. 3), which indicated
that was predominantly expressed in neurons and microglia during
the early stages of mouse brain development.
Western blot analysis of Ubqln in the
HIE mouse model
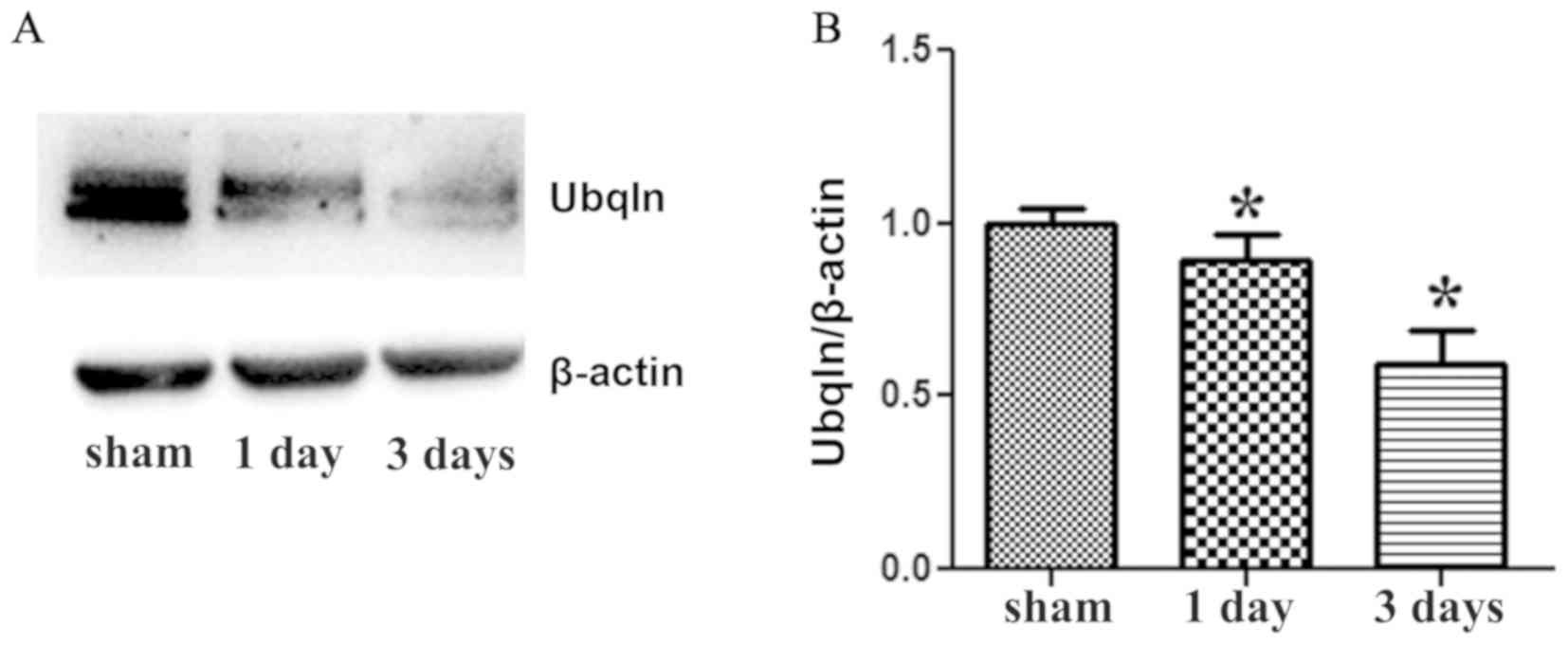
Western blot was then performed to further determine
the protein expression of Ubqln in the brains of the two groups.
Ubqln expression was significantly decreased in the HI group at 1
and 3 days after HI injury compared with the sham group (Fig. 4). β-actin (43 kDa) was used as an
internal control. Therefore, the western blot results indicated
that Ubqln expression was markedly decreased following exposure to
HI brain injury.
Discussion
This study if the first to report, to the best of
our knowledge, Ubqln expression in the brains of a HIE mouse model.
The present work demonstrated the following: i) Ubqln was widely
expressed in the cortex and hippocampus in brains of the HIE mouse
model; ii) Ubqln was expressed in neurons, astrocytes, and
microglia in the brains of the HIE mouse model; and iii) Ubqln
expression was downregulated after neonatal HI brain injury in
mice.
The pathogenesis of HIE is not fully understood,
thus studying the underlying molecular mechanism is important for
the development of novel prophylactics and therapeutics against
neuronal death in neurodegenerative diseases. In particular, the
isolation and identification of novel molecules associated neuronal
survival/death is important. Currently, increasing evidence links
Ubqln to the pathogenic mechanism underlying Alzheimer's disease
(AD) and other neurodegenerative diseases (14). Ubqln has been reported to have a
critical role in the regulation of the levels, subcellular
targeting, aggregation, and degradation of various
neurodegenerative disease-associated proteins (15). Despite a number of studies
regarding the role of Ubqln in anti-oxidation (16), regulation of autophagy (17), cell protection and involvement in
tumorigenesis (18), no precise
location and distribution of Ubqln in neonatal HIE has been
reported previously. In the present study, Ubqln immunofluorescence
staining revealed that Ubqln was expressed abundantly in the brains
of neonatal sham and HI pups. The expression of Ubqln and prognosis
in the brains of neonatal HI pups was clarified for the first time,
to the best of our knowledge, and Ubqln may be a novel molecular
marker to predict prognosis in HIE.
Numerous studies have demonstrated that the secreted
protein, Ubqln, has an essential role in the regulation of protein
degradation, which is involved in the pathophysiology of cancer and
neurodegenerative diseases. Ubqln is frequently overexpressed in
breast (19), gastric (20) and lung cancers (21). It has been suggested that high
Ubqln expression is associated with tumor size, lymph node
metastasis, TNM stage and vascular invasion, and is significantly
associated with a worse prognosis in patients with gastric and
breast cancer (19,20); however, Shah et al (21) reported that expression of Ubqln
serves as a potential predictive biomarker for therapeutic efficacy
in patients of non-small cell lung cancer. In the central nervous
system, Ubqln is an AD-associated protein, which is known to
modulate amyloid precursor protein processing, amyloid-β secretion,
and presenilin-1 accumulation (22). A study by Satoh et al
(23) showed Ubqln expression in
the frontal cortex and hippocampus in brains from patients with AD.
Furthermore, Ubqln immunoreactivity is concentrated in Hirano
bodies and dystrophic neurites in brains from patients with AD,
which suggests that aberrant expression of Ubqln may be a
pathologic hallmark of AD. Based on in vitro studies, Ubqln
expression has been reported in human neuroblastoma cells and rat
cortical neurons (24). In the
present study, cell localization of Ubqln in the brains of neonatal
HI pups was identified. Ubqln was expressed in neurons, astrocytes
and microglia in vivo. The results of the current study
agree with a previous study (24).
Ubqln has an important role in clearing mislocalized
mitochondrial proteins upon cell stimulation, and the absence leads
to suppression of protein synthesis and cell cycle arrest (25). In the present study, the expression
of Ubqln protein was significantly decreased in HI model mice
compared with sham controls, as determined by western blot
analysis, suggesting that decreased Ubqln may have a role in the
development of HIE. Liu et al (16,26)
demonstrated that Ubqln protects cells from oxidative stress and
ischemic stroke causing tissue injury in mice by developing Ubqln
transgenic and conditional knockout mouse models to perform gain-
and loss-of-function analysis of Ubqln. The yeast two-hybrid system
has shown that Ubqln interacts with protein disulfide isomerase
(PDI), and observed that Ubqln, together with PDI, is localized in
the endoplasmic reticulum (ER) and upregulated in response to
hypoxia (27). It has also been
demonstrated that Ubqln association with PDI in the ER is involved
in tolerance to stress-induced apoptotic cell death (28). In neonatal brains, HI brain injury
usually causes cell death via necrosis or apoptosis (29). Previous studies have reported that
apoptosis is more frequent in HI brain injury (30,31).
Kojima et al (32) reported
that many genes upregulated following HI injury are associated with
cell death signaling, such as the arachidonic acid cascade. By
contrast, many downregulated genes affect the expression of target
genes, reflecting progressive damage by the HI insult. The lower
expression of Ubqln in the brains of neonatal HI pups indicates
that Ubqln may contribute to the pathogenesis of HIE by regulating
apoptosis.
In summary, immunofluorescence staining and western
blot analysis demonstrated the expression and cell location of
Ubqln in the brains of neonatal HI pups. Decreased expression of
Ubqln was detected following HI brain injury, which suggests that
the decreased expression of Ubqln may contribute to the development
of HIE. Therefore, further studies should focus on the mechanism
underlying the regulation of the changes in Ubqln during HIE.
Acknowledgements
Not applicable.
Funding
This project was financially supported by the
Natural Science Foundation of Guangdong Province (grant no.
2018A030313579), the Natural Science Foundation of Guangdong
Province, the Fundamental Research Funds for the Central
Universities (grant no. 14ykpy33), the Science Foundation of
Guangdong No. 2 Provincial People's Hospital for Youth (grant no.
YQ2017-001), the Science and Technology Programs in Educational
Commission of Guangdong Province (2016), the Innovative and
Efficient Projects of Guangdong Pharmaceutical University (2016),
the Guangdong Province Innovation and Entrepreneurship Training
Program for University Students (grant no. 201710573046) and the
Medical Scientific Research Foundation of Guangdong Province, China
(grant no. A2015131).
Availability of data and materials
statements
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LiL contributed to the general administration,
statistical analysis, manuscript writing; YL, XT, XR and WZ
assisted in the completion of western blotting, behavioral tests
and other procedures; JL, LZ and WC assisted in the completion of
western blotting, immunofluorescence and other procedures. PZ and
WW established animal models and providing technical guidance; LaL
and MW provided technical guidance. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The animal-related experiments were approved and
organized in accordance with the guidelines of the Experimental
Laboratory Animal Committee of Guangdong Pharmaceutical University
(permit no: gdpulac2017175).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Ubqln1
|
ubiquilin-1
|
|
CCA
|
common carotid artery
|
|
HIE
|
hypoxic-ischemic encephalopathy
|
|
TTC
|
triphenyl tetrazolium chloride
|
References
|
1
|
Lv H, Wang Q, Wu S, Yang L, Ren P, Yang Y,
Gao J and Li L: Neonatal hypoxic ischemic encephalopathy-related
biomarkers in serum and cerebrospinal fluid. Clin Chim Acta.
450:282–297. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Silveira RC and Procianoy RS: Hypothermia
therapy for newborns with hypoxic ischemic encephalopathy. J
Pediatr (Rio J) 91(6 Suppl 1). S78–S83. 2015. View Article : Google Scholar
|
|
3
|
Douglas-Escobar M and Weiss MD:
Hypoxic-ischemic encephalopathy: A review for the clinician. JAMA
Pediatr. 169:397–403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vannucci RC, Connor JR, Mauger DT, Palmer
C, Smith MB, Towfighi J and Vannucci SJ: Rat model of perinatal
hypoxic-ischemic brain damage. J Neurosci Res. 55:158–163. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Perlman JM: Intervention strategies for
neonatal hypoxic-ischemic cerebral injury. Clin Ther. 28:1353–1365.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li L, Klebe D, Doycheva D, McBride DW,
Krafft PR, Flores J, Zhou C, Zhang JH and Tang J: G-CSF ameliorates
neuronal apoptosis through GSK-3β inhibition in neonatal
hypoxia-ischemia in rats. Exp Neurol. 263:141–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jansen AH, Reits EA and Hol EM: The
ubiquitin proteasome system in glia and its role in
neurodegenerative diseases. Front Mol Neurosci. 7:732014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Massey LK, Mah AL and Monteiro MJ:
Ubiquilin regulates presenilin endoproteolysis and modulates
gamma-secretase components, Pen-2 and nicastrin. Biochem J.
391:513–525. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Natunen T, Takalo M, Kemppainen S, Leskelä
S, Marttinen M, Kurkinen KMA, Pursiheimo JP, Sarajärvi T,
Viswanathan J, Gabbouj S, et al: Relationship between ubiquilin-1
and BACE1 in human Alzheimer's disease and APdE9 transgenic mouse
brain and cell-based models. Neurobiol Dis. 85:187–205. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang FF and Li J: Inhibitory effect of
chloroquine derivatives on presenilin 1 and ubiquilin 1 expression
in Alzheimer's disease. Int J Clin Exp Pathol. 8:7640–7643.
2015.PubMed/NCBI
|
|
11
|
Rutherford NJ, Lewis J, Clippinger AK,
Thomas MA, Adamson J, Cruz PE, Cannon A, Xu G, Golde TE, Shaw G, et
al: Unbiased screen reveals ubiquilin-1 and −2 highly associated
with huntingtin inclusions. Brain Res. 1524:62–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Institute of Laboratory Animal Resources
(US), . Committee on Care, Use of Laboratory Animals, National
Institutes of Health (US). Division of Research Resources. Guide
for the care and use of laboratory animals. National Academies.
1985.
|
|
13
|
Rice JE III, Vannucci RC and Brierley JB:
The influence of immaturity on hypoxic ischemic brain damage in the
rat. Ann Neurol. 9:131–141. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takalo M, Haapasalo A, Natunen T,
Viswanathan J, Kurkinen KM, Tanzi RE, Soininen H and Hiltunen M:
Targeting ubiquilin-1 in Alzheimer's disease. Expert Opin Ther
Targets. 17:795–810. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gadhave K, Bolshette N, Ahire A, Pardeshi
R, Thakur K, Trandafir C, Istrate A, Ahmed S, Lahkar M, Muresanu DF
and Balea M: The ubiquitin proteasomal system: A potential target
for the management of Alzheimer's disease. J Cell Mol Med.
20:1392–1407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Lü L, Hettinger CL, Dong G, Zhang
D, Rezvani K, Wang X and Wang H: Ubiquilin-1 protects cells from
oxidative stress and ischemic stroke caused tissue injury in mice.
J Neurosci. 34:2813–2821. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
N'Diaye EN, Kajihara KK, Hsieh I, Morisaki
H, Debnath J and Brown EJ: PLIC proteins or ubiquilins regulate
autophagy-dependent cell survival during nutrient starvation. EMBO
Rep. 10:173–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yadav S, Singh N, Shah PP, Rowbotham DA,
Malik D, Srivastav A, Shankar J, Lam WL, Lockwood WW and Beverly
LJ: MIR155 regulation of ubiquilin1 and ubiquilin2: Implications in
cellular protection and tumorigenesis. Neoplasia. 19:321–332. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Lu J, Zhao X, Feng Y, Lv S, Mu Y,
Wang D, Fu H, Chen Y and Li Y: Prognostic significance of
Ubiquilin1 expression in invasive breast cancer. Cancer Biomark.
15:635–643. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bao J, Jiang X, Zhu X, Dai G, Dou R, Liu
X, Sheng H, Liang Z and Yu H: Clinical significance of ubiquilin 1
in gastric cancer. Medicine (Baltimore). 97:e97012018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shah PP, Lockwood WW, Saurabh K, Kurlawala
Z, Shannon SP, Waigel S, Zacharias W and Beverly LJ: Ubiquilin1
represses migration and epithelial-to-mesenchymal transition of
human non-small cell lung cancer cells. Oncogene. 34:1709–1717.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Viswanathan J, Haapasalo A, Kurkinen KM,
Natunen T, Mäkinen P, Bertram L, Soininen H, Tanzi RE and Hiltunen
M: Ubiquilin-1 modulates γ secretase mediated ε-site cleavage in
neuronal cells. Biochemistry. 52:3899–3912. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Satoh J, Tabunoki H, Ishida T, Saito Y and
Arima K: Ubiquilin-1 immunoreactivity is concentrated on Hirano
bodies and dystrophic neurites in Alzheimer's disease brains.
Neuropathol Appl Neurobiol. 39:817–830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Z, Ruan Y, Yue W, Zhu Z, Hartmann T,
Beyreuther K and Zhang D: GM1 up-regulates Ubiquilin 1 expression
in human neuroblastoma cells and rat cortical neurons. Neurosci
Lett. 407:59–63. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Whiteley AM, Prado MA, Peng I, Abbas AR,
Haley B, Paulo JA, Reichelt M, Katakam A, Sagolla M, Modrusan Z, et
al: Ubiquilin1 promotes antigen-receptor mediated proliferation by
eliminating mislocalized mitochondrial proteins. Elife.
6:e264352017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Qiao F and Wang H: Enhanced
proteostasis in post-ischemic stroke mouse brains by ubiquilin-1
promotes functional recovery. Cell Mol Neurobiol. 37:1325–1329.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nomura Y: Neuronal apoptosis and
protection: Effects of nitric oxide and endoplasmic
reticulum-related proteins. Biol Pharm Bull. 27:961–963. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ko HS, Uehara T and Nomura Y: Role of
ubiquilin associated with protein-disulfide isomerase in the
endoplasmic reticulum in stress-induced apoptotic cell death. J
Biol Chem. 277:35386–35392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gill MB and Perez-Polo JR: Hypoxia
ischemia-mediated cell death in neonatal rat brain. Neurochem Res.
33:2379–2389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ekert P, MacLusky N, Luo XP, Lehotay DC,
Smith B, Post M and Tanswell AK: Dexamethasone prevents apoptosis
in a neonatal rat model of hypoxic-ischemic encephalopathy (HIE) by
a reactive oxygen species-independent mechanism. Brain Res.
747:9–17. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hernández-Jiménez M, Sacristán S, Morales
C, García-Villanueva M, García-Fernández E, Alcázar A, González VM
and Martín ME: Apoptosis-related proteins are potential markers of
neonatal hypoxic-ischemic encephalopathy (HIE) injury. Neurosci
lett. 558:143–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kojima T, Ueda Y, Sato A, Sameshima H and
Ikenoue T: Comprehensive gene expression analysis of cerebral
cortices from mature rats after neonatal hypoxic-ischemic brain
injury. J Mol Neurosci. 49:320–327. 2013. View Article : Google Scholar : PubMed/NCBI
|