Introduction
Chronic obstructive pulmonary disease (COPD) is a
common respiratory disease characterized by incomplete
reversibility of airflow obstruction. The pathological changes
occurring in COPD include airway reconstruction and pulmonary
artery reconstruction, particularly chronic bronchitis and
emphysema (1). Airway remodeling
is a key step in regulating disease progression, whereas
uncontrollable activation of fibroblasts is central to airway
reconstruction (1,2). As previously documented, factors
including matrix metalloproteinase-9 (MMP-9), platelet-derived
growth factor (PDGF) and transforming growth factor-β1 (TGF-β1) are
notably involved in airway remodeling (3–5).
The hyperplasia suppressor gene (HSG) can inhibit
cell proliferation and the cell cycle (6,7). HSG
is reportedly highly expressed in normal cells, with reduced
expression in proliferating cells (8–10).
Overexpression of HSG can inhibit the malignant phenotype of breast
cancer (11) and induce apoptosis
in tumor cells (10). Of note,
recombinant HSG can suppress the proliferation, and induce the
apoptosis of airway fibroblasts (12); however, whether HSG is a suppressor
of airway fibroblasts, and the underlying mechanisms involved in
this process are yet to be verified.
The Wnt signaling pathway is involved not only in
lung development, but also in the pathogenesis of pulmonary
fibrosis (13), idiopathic
pulmonary hypertension (14) and
pulmonary interstitial disease (15). The Wnt/β-catenin signaling pathway
is required for cell differentiation, apoptosis, carcinogenesis,
immunity, and other physiological and pathological processes
(16,17) and provides targets for the
development of novel therapeutics to intervene with the development
of COPD. The molecular mechanisms underpinning the pathogenesis of
Wnt signaling components, including the destructive complex, in
COPD are unclear (18); however,
its role in cardiac and liver fibrosis has previously been explored
(19,20). As both the HSG and Wnt signaling
pathways contribute to fibrosis, it remains unclear whether HSG may
regulate the Wnt/β-catenin signaling pathway in airway
reconstruction. In the present study, an HSG overexpression vector
was constructed to transfect airway fibroblasts in vitro to
verify the effect of HSG overexpression on activation of airway
fibroblasts, and assess the underlying mechanisms.
Materials and methods
Animals
A total of 20 male Sprague-Dawley rats (age, 3
months; body weight, 250 g) were purchased from Hunan Slake Jingda
Laboratory Animal Co., Ltd. [license no. SCXK (Hunan) 2016-0002].
All animals were raised in non-toxic plastic boxes, stainless steel
wire cages and metal cages, and housed in a specific pathogen-free
environment that was automatically maintained at a temperature of
22±2°C and a relative humidity of 45–65%, under a controlled 12-h
light/dark cycle with access to food and water ad libitum.
All animal experiments were approved by the Ethics Committee of
Guizhou College of Traditional Chinese Medicine (Guiyang,
China).
Establishment of a rat model of
COPD
Following anesthesia with sodium pentobarbital (40
mg/kg, intraperitoneal), rat skins were disinfected with iodophor.
The epidermis and muscles along the middle of the neck were cut
using surgical scissors to expose the trachea. Papain solution (2
mg/ml) was injected into the trachea at a dose of 1 ml/kg from top
to bottom to reach the lungs. Subsequently, the animals were picked
up and rotated to ensure that the drug had fully entered the
trachea. Thereafter, the wound was sutured, and the animals were
placed on an electric blanket. The animals were anesthetized with
isoflurane (1% in oxygen) and decapitated 8 days after remodeling.
Airway tissues from 3 animals were collected on ice and fixed in 4%
paraformaldehyde overnight at 4°C. The tissues were sectioned into
5-µm thick sections and mounted on slides, followed by hematoxylin
and eosin staining for 5 min at room temperature. The stained
sections were observed under a light microscope (magnification:
×200; BX51; Olympus Corporation).
Preparation of airway fibroblasts
A total of 12 rats were sacrificed and
thoracotomized under aseptic conditions to collect airway
fibroblasts as previously described (21). A sterile scalpel was used to
collect the tissue surrounding the airway; freshly obtained airway
tissue from COPD rats was washed five times with phosphate-buffered
saline (PBS) on an aseptic worktable. The tissue was then cut into
2×2-mm pieces using sterilized ophthalmic scissors. The tissue was
pasted into the culture plate, and placed in a 5% CO2
incubator at 37°C for 4 h. After cell adherence, freshly prepared
medium containing DMEM (Gibco; Thermo Fisher Scientific, Inc.) and
10% fetal bovine serum (FBS; cat. no. 04-007-1A; Biological
Industries) was added to the culture plate, and the cells were
further cultured in a 5% CO2 incubator at 37°C. Cell
growth was monitored, the cell supernatant was periodically
discarded, and newly prepared DMEM +10% FBS culture medium was
added into the culture dish.
Immunohistochemistry
The cultured dishes (3×103/ml) were
rinsed three times in PBS (3 min/wash), fixed in 4%
paraformaldehyde for 15 min at room temperature, and permeated with
0.5% Triton X-100 PBS for 20 min at room temperature. Bovine serum
albumin (5%; Gibco; Thermo Fisher Scientific, Inc.) was added to
the culture dish, which was subsequently sealed at 37°C for 30 min.
Anti-vimentin antibody (1:250; cat. no. ab92547; Abcam) was added
to the dish and incubated at 37°C for 3 h. The diluted
Cy3-conjugated secondary antibody (1:200, cat. no. BA1032, Wuhan
Boster Biological Technology, Ltd.) was added and incubated at 37°C
for 30 min. Subsequently, DAPI was used to stain the nuclei for 3
min at room temperature, and the images were observed under a
fluorescence microscope (magnification: ×200) as previously
described (22).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Airway fibroblasts (3×103 cells/well)
were seeded in 6-well plates. Following transfection and/or
treatment, total mRNA was extracted using a TRIzol®
assay kit (Baosheng Science & Technology Innovation Co. Ltd.).
RNA quality was evaluated based on the optical density (260/280
nm). mRNA was transcribed into cDNA using a SMART® MMLV
Reverse Transcriptase kit (cat. no. 639522; Takara Biotechnology
Co., Ltd.) at 37°C using the following thermocycling conditions:
25°C for 10 min, 37°C for 120 min and 85°C for 5 min, which was
subsequently used as the template for fluorescent qPCR. The
reaction mixture included the following components: Deionized water
(9.5 µl), cDNA (1 µl), primers (2 µl) and 2X UltraSYBR Mixture
(12.5 µl; HY-K0501; MedChemExpress LLC) under the following cycling
conditions: 35 cycles of 94°C denaturation for 45 sec, 59°C
annealing for 45 sec and 72°C extension for 60 sec. The relative
mRNA expression of HSG, β-catenin and Ras homology family member A
(RhoA) was calculated using GAPDH as the reference using the
2−ΔΔCq method (23).
The primers used in the study are presented in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene name | Primer sequences
(5′-3′) | Primer length
(bp) | Product length
(bp) | Annealing
temperature (°C) |
|---|
| HSG F |
AACTCCATCGTCACCGTCAA | 20 | 385 | 59 |
| HSG R |
CAACCCGCAGGAAGCAA | 17 |
|
|
| RhoA F |
AGAGTTGGCTTTATGGGACAC | 21 | 181 | 59 |
| RhoA R |
GATGATGGGCACATTTGGA | 19 |
|
|
| β-catenin F |
TTATGAGTGGGAGCAAGGC | 19 | 451 | 59 |
| β-catenin R |
ACAACGGGCTGTTTCTACG | 19 |
|
|
| GAPDH F |
GCAAGTTCAACGGCACAG | 18 | 141 | 59 |
| GAPDH R |
CGCCAGTAGACTCCACGAC | 19 |
|
|
Experimental groups
Airway fibroblasts were divided into nine groups:
Control, vector, HSG overexpression (HSG), lithium chloride (LiCl),
vector + LiCl, HSG + LiCl,
4-ethyl-5,6-dihydro-5-methyl-(1,3)dioxolo(4,5-j)phenanthridine
(HLY78), vector + HLY78, and HSG + HLY78 (Fig. 1A). Wnt signaling pathway agonists,
LiCl (cat. no. B6083) and HLY78 (cat. no. C5433) were purchased
from Apexbio Technology, LLC. The cells were treated with either 10
mM LiCl or 10 µM HLY78 for 24 h, at 48 h at 37°C following
transfection with the HSG vector.
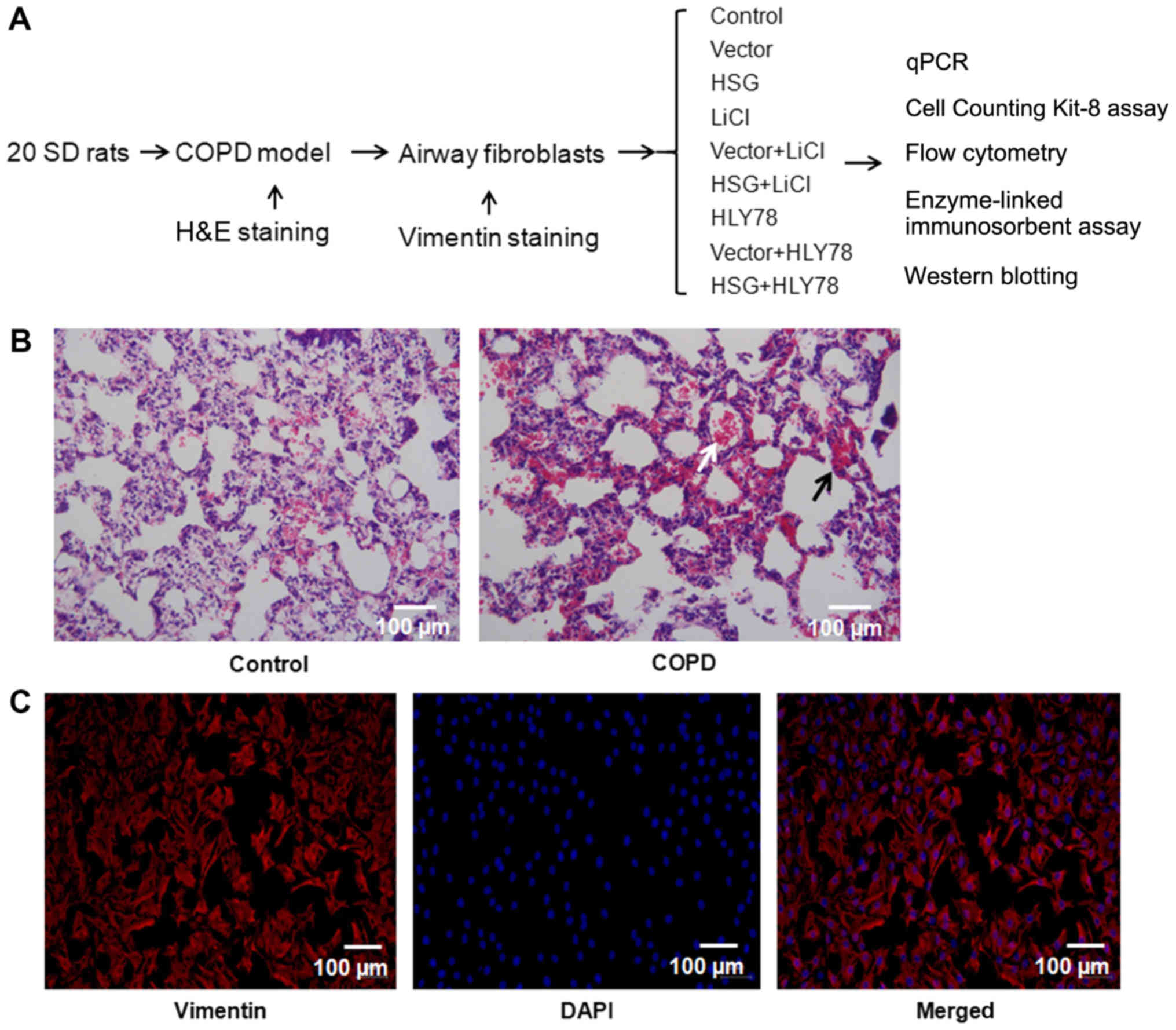 | Figure 1.Airway fibroblasts collected from
COPD rats. (A) Schematic illustration of the study design. (B)
H&E staining of the airway tissue. Black and white arrows
indicate infiltration of inflammatory cells and exfoliation,
respectively. (C) Identification of airway fibroblasts
(magnification, ×200). Red represents vimentin expression; blue
represents nuclear staining (DAPI). COPD, chronic obstructive
pulmonary disease; HSG, hyperplasia suppressor gene; LiCl, lithium
chloride; HLY78,
4-ethyl-5,6-dihydro-5-methyl-(1,3)dioxolo(4,5-j)phenanthridine;
qPCR, quantitative polymerase chain reaction; SD, Sprague-Dawley;
H&E, hematoxylin and eosin. |
Cell transfection
The HSG gene sequence (NM_130894) was
obtained from the National Centre for Biotechnology Information
database, and a biosynthetic gene fragment was cloned into the
pCDNA3.1 (+) vector (Shanghai GenePharma Co. Ltd.) at the digestion
site (Hind III/Xho I). Cells at 60% confluence were
transfected with empty vector or HSG (1 µg/ml) using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 6 h, the medium was replaced with fresh
DMEM containing 10% FBS and cultured in a 5% CO2
incubator at 37°C for 48 h. The cells were then used in the
following experiments.
Cell Counting Kit-8 assay
After discarding the culture medium, the cells
(3×105/ml) were washed with PBS and digested with
trypsin for 2–3 min. Next, the cells were resuspended, inoculated
into 96-well plates and placed in a 5% CO2 incubator at
37°C for 24 h. The assay was performed as previously described
(24). The optical density was
determined using a microplate reader (Bio-Tek Instruments, Inc.) at
570 nm to determine cell viability.
Flow cytometry
The cells (3×105/ml) were incubated in
the dark with Annexin V-fluorescein isothiocyanate and propidium
iodide (cat. no. AP101-100; Multisciences Biotech Co., Ltd.) for 30
min at room temperature. Subsequently, apoptosis was detected using
a flow cytometer (BD Biosciences) within 1 h. Totally, four
quadrants were divided based upon the fluorescence intensity and
analyzed using FlowJo v10 (FlowJo, LLC). Quadrant Q2-2 was added to
box quadrant Q2-4 to obtain the percentage of apoptotic cells.
ELISA
The cells were collected and total protein was
isolated using a triple prep kit (cat. no. 28-9425-44, ReadyPrep;
GE Healthcare Life Sciences). Protein concentration was measured
using a bicinchoninic acid assay kit (cat. no. P0009: Beyotime
Institute of Biotechnology). Volumes were adjusted to normalize the
protein content, and then aliquots were processed for the ELISA
using rat TGF-β1 (cat. no. MM-0181R1), rat MMP-9 (cat. no.
MM-20918R1) and rat PDGF assay kits (cat. no. MM-0076R1; all MlBio;
Shanghai Enzyme Biotechnology Co., Ltd.).
Western blot analysis
Following transfection and/or treatment as
aforementioned, protein was extracted from each group using a
protein isolation kit (cat. no. 28-9425-44; GE Healthcare Life
Sciences). Protein levels were quantified with a bicinchoninic acid
protein assay kit (cat. no. P0009; Beyotime Institute of
Biotechnology). Proteins (25 µg/lane; 0.5 µg/µl) were separated via
12% SDS-PAGE and transferred onto polyvinylidene fluoride
membranes. The membranes were blocked in 5% skimmed milk for 2 h at
room temperature and incubated with the following primary
antibodies overnight at 4°C: Mouse monoclonal anti-GAPDH (1:2,000;
cat. no. TA-08; OriGene Technologies, Inc.); rabbit polyclonal
anti-HSG (1:5,000; cat. no. ab124773; Abcam); rabbit polyclonal
anti-β-catenin (1:300; cat. no. ab32572; Abcam) and rabbit
polyclonal anti-RhoA (1:2,000; cat. no. ab187026; Abcam). The IgG
horseradish peroxidase-conjugated secondary antibody (mouse: 1:100;
cat. no. ab131368; Abcam; rabbit: 1:100; ZB-2301; OriGene
Technologies, Inc.) was added and co-incubated for 2 h at room
temperature. An enhanced chemiluminescence exposure liquid droplet
(cat. no. RJ239676; Thermo Fisher Scientific, Inc.) was added to
the membranes. The membranes were visualized using a gel imaging
system (Bio-Rad Laboratories, Inc.). Densitometry was performed
using Quantity One version 1.4.6 (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data were presented as the mean ± standard error of
the mean with 6 independent repeats and analyzed using SPSS version
17.0 (SPSS, Inc.). Significant differences were determined using
one-way analysis of variance, followed by the Student-Newman-Keuls
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
HSG overexpression reduces the
viability of airway fibroblasts
The airway tissue of the control group exhibited
regular alveolar structure (Fig.
1B). Pathological expansion and fusion of alveolar cavities
were not observed. The wall of the bronchial tube was normal, the
epithelium of the airway mucosa was smooth, cilia were neatly
arranged, and obvious inflammatory exudation was not found in the
tracheal cavity. In contrast, the airway tissue of the COPD group
showed evidence of emphysema, alveolar dilatation, an alveolar wall
that disintegrated and fused to form lung ulcers, and markedly
decreased numbers of alveoli (Fig.
1B). The goblet cells of the bronchial epithelium showed
proliferation, and a number of inflammatory cells such as
neutrophils and giant cells infiltrated the lumen, accompanied by
the proliferation of fibrous connective tissue.
The identification of airway fibroblasts is shown in
Fig. 1C, wherein red indicates
vimentin expression, and nuclear staining by DAPI is observed in
blue. All cells showed vimentin expression, indicating successful
preparation of the airway fibroblasts. Compared with the vector
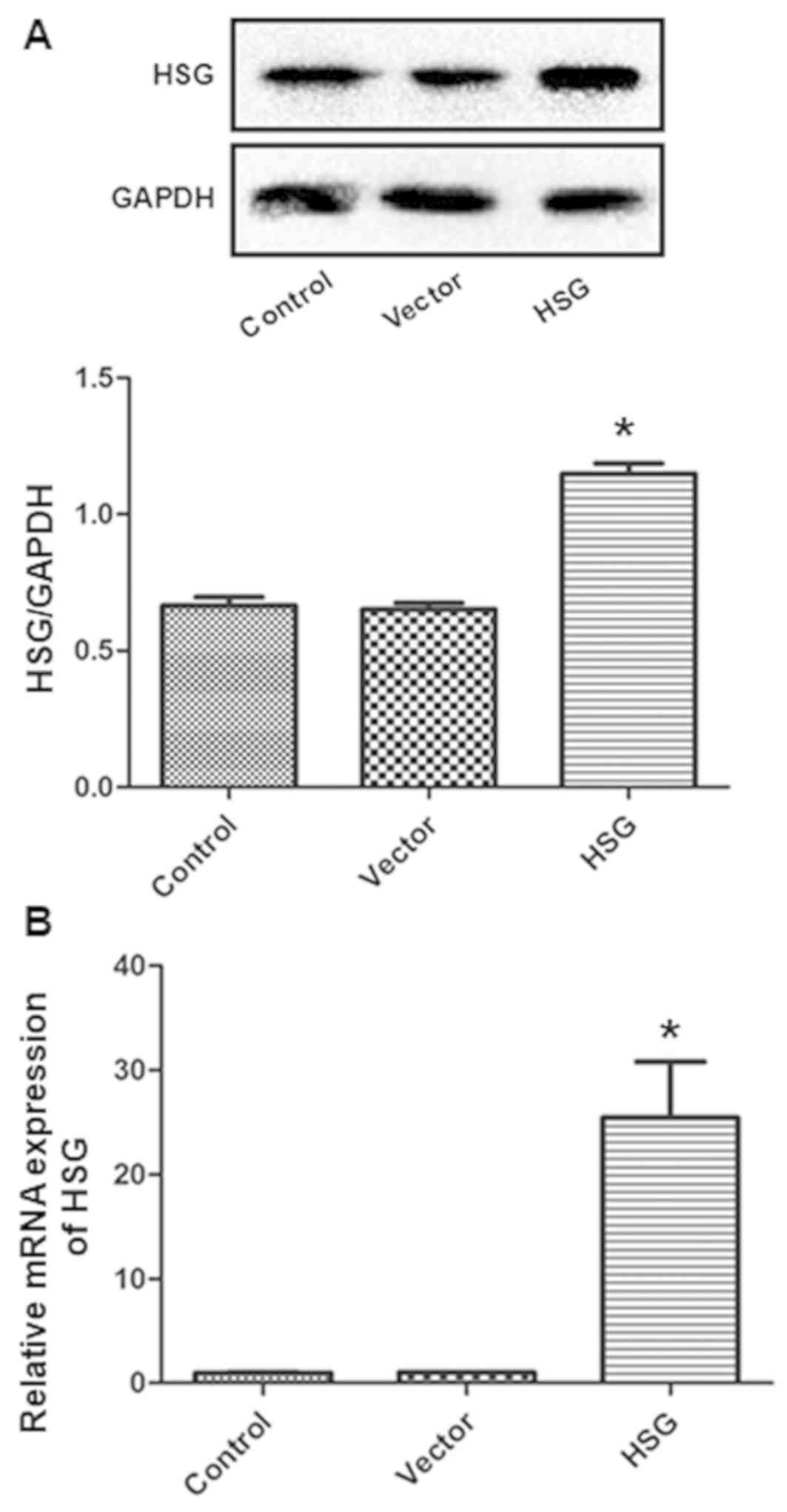
group, the HSG vector upregulated HSG expression at both the mRNA
and protein levels (both P<0.05; Fig. 2A and B), whereas the control vector
did not influence HSG expression. Thereafter, the effects of HSG
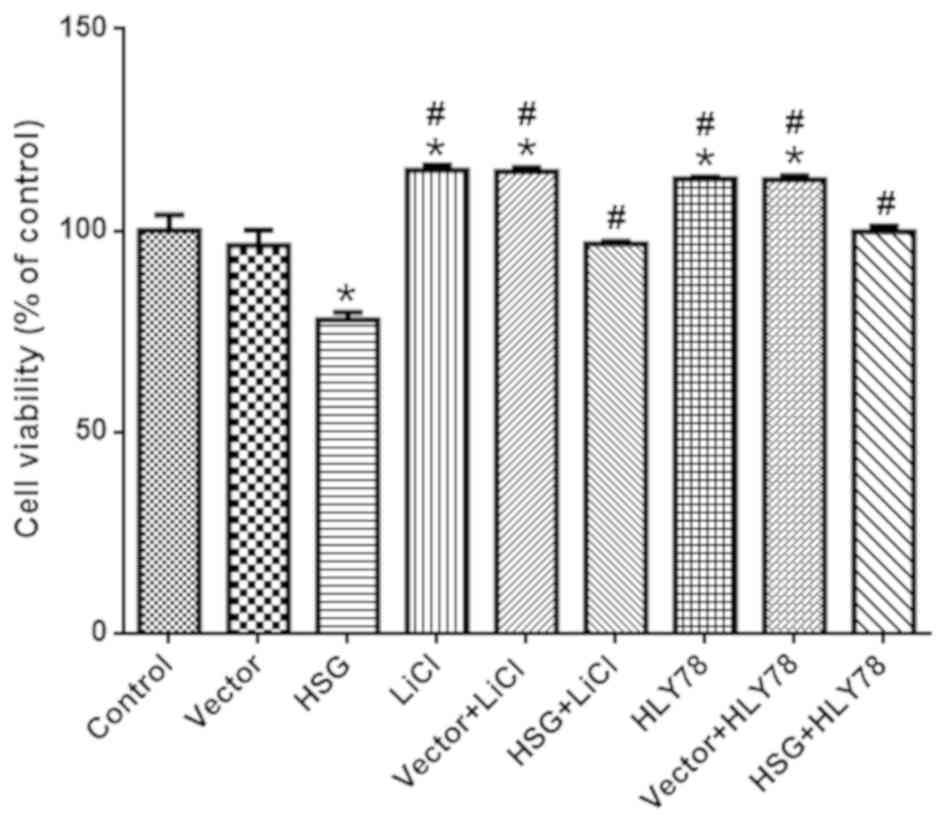
overexpression on cell viability were measured. The cell viability
of the HSG group was decreased (vs. vector, P<0.05), which was
inhibited by HLY78 and LiCl (vs. HSG, P<0.05; Fig. 3). Of note, LiCl and HLY78 alone
increased cell viability (vs. control, P<0.05).
HSG overexpression facilitates
apoptosis of airway fibroblasts
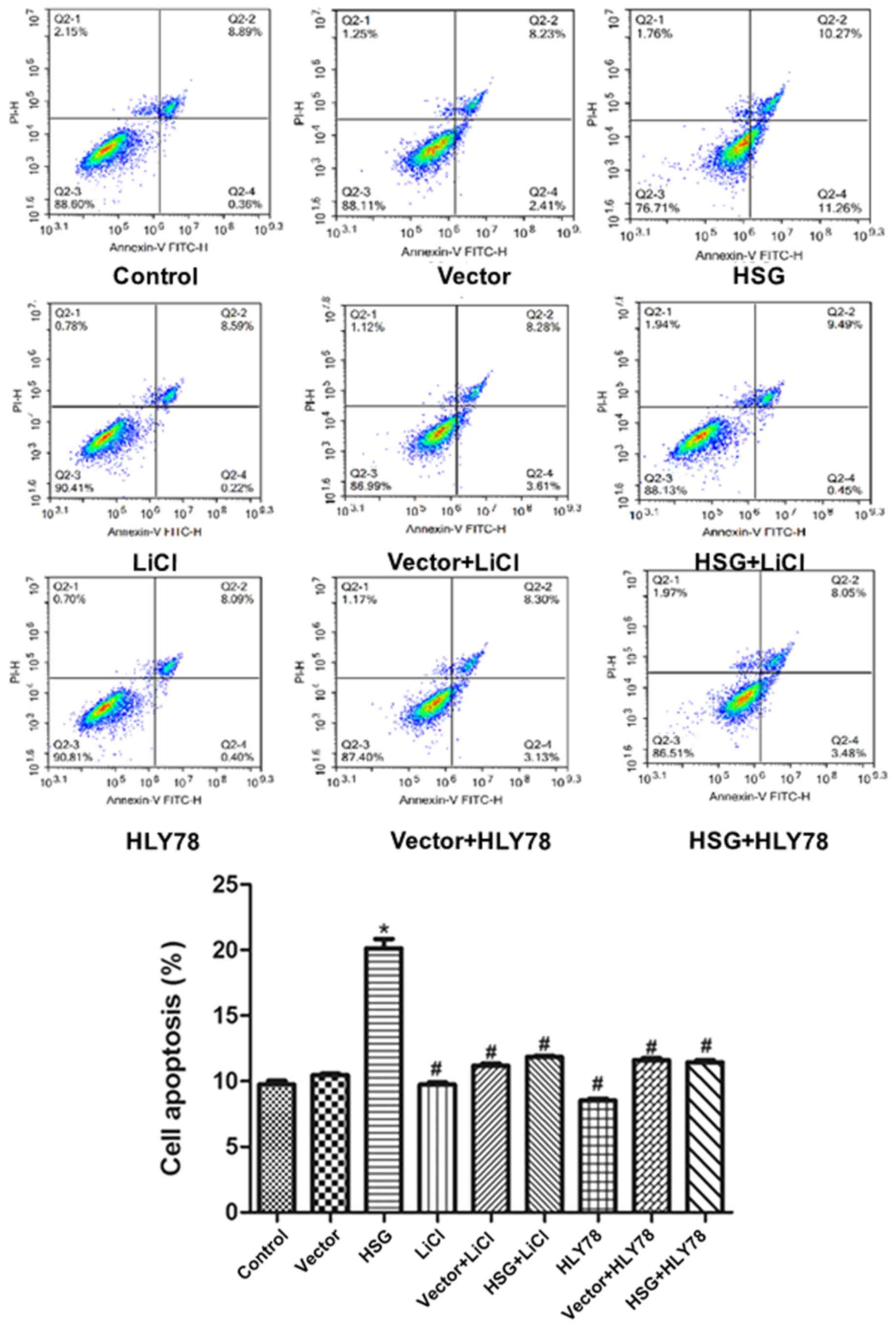
To further examine the effects of HSG overexpression
on apoptosis, flow cytometry was used to detect the cell
distribution following double staining. The apoptosis rate in the
HSG group was significantly higher compared with that observed in
the vector group (P<0.05; Fig.
4). LiCl and HLY78 reduced the effects of HSG overexpression on
apoptosis (vs. HSG, P<0.05). The rates of apoptosis in the HSG +
LiCl and HSG + HLY78 groups were significantly lower compared with
that detected in the HSG group (P<0.05). However, LiCl and HLY78
alone did not affect apoptosis (vs. control, P>0.05; Fig. 4).
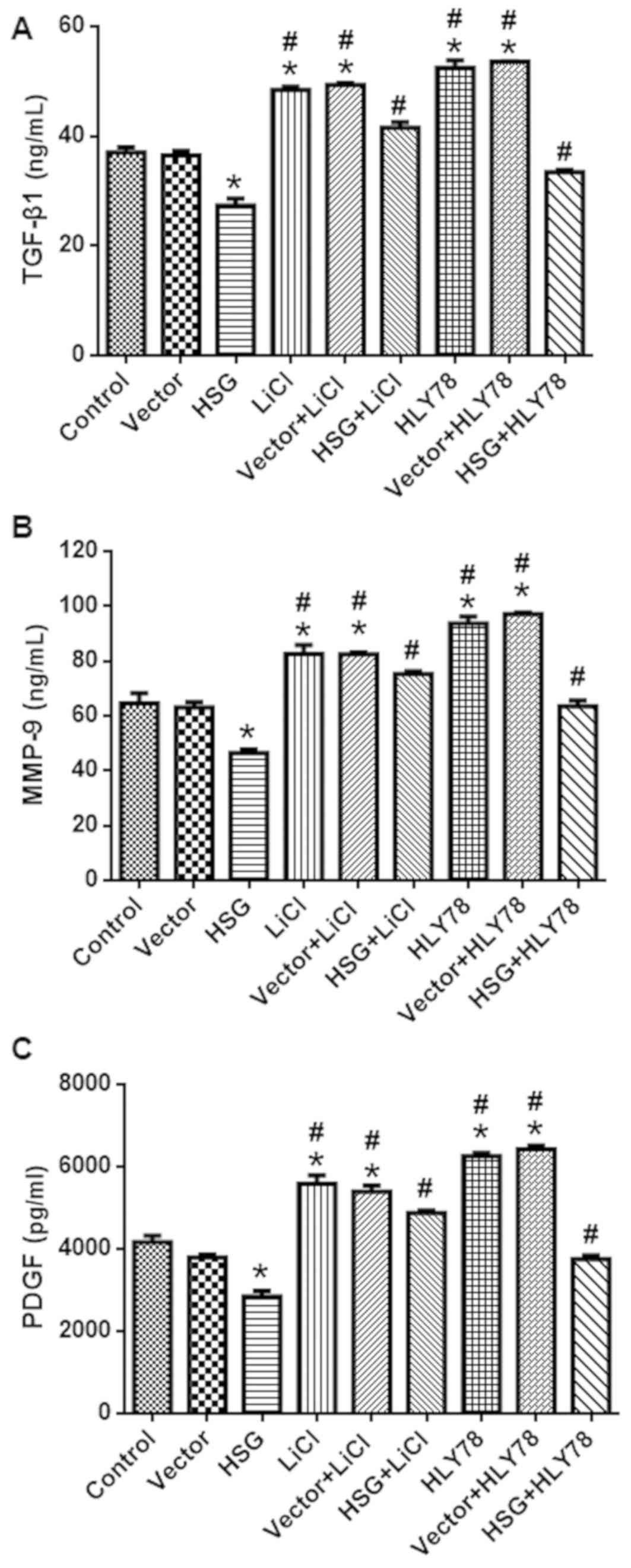
HSG overexpression reduces the levels
of TGF-β1, MMP-9, and PDGF
The protein levels of TGF-β1, MMP-9, and PDGF in
each group are shown in Fig. 5.
The levels of TGF-β1 (Fig. 5A),
MMP-9 (Fig. 5B), and PDGF
(Fig. 5C) in the HSG group were
decreased compared with in the control group. In contrast, these
levels were higher in the HSG + LiCl and HSG + HLY78 groups
compared with the HSG group (P<0.05). LiCl and HLY78 alone also
increased the levels of TGF-β1, MMP-9, and PDGF (vs. control;
P<0.05).
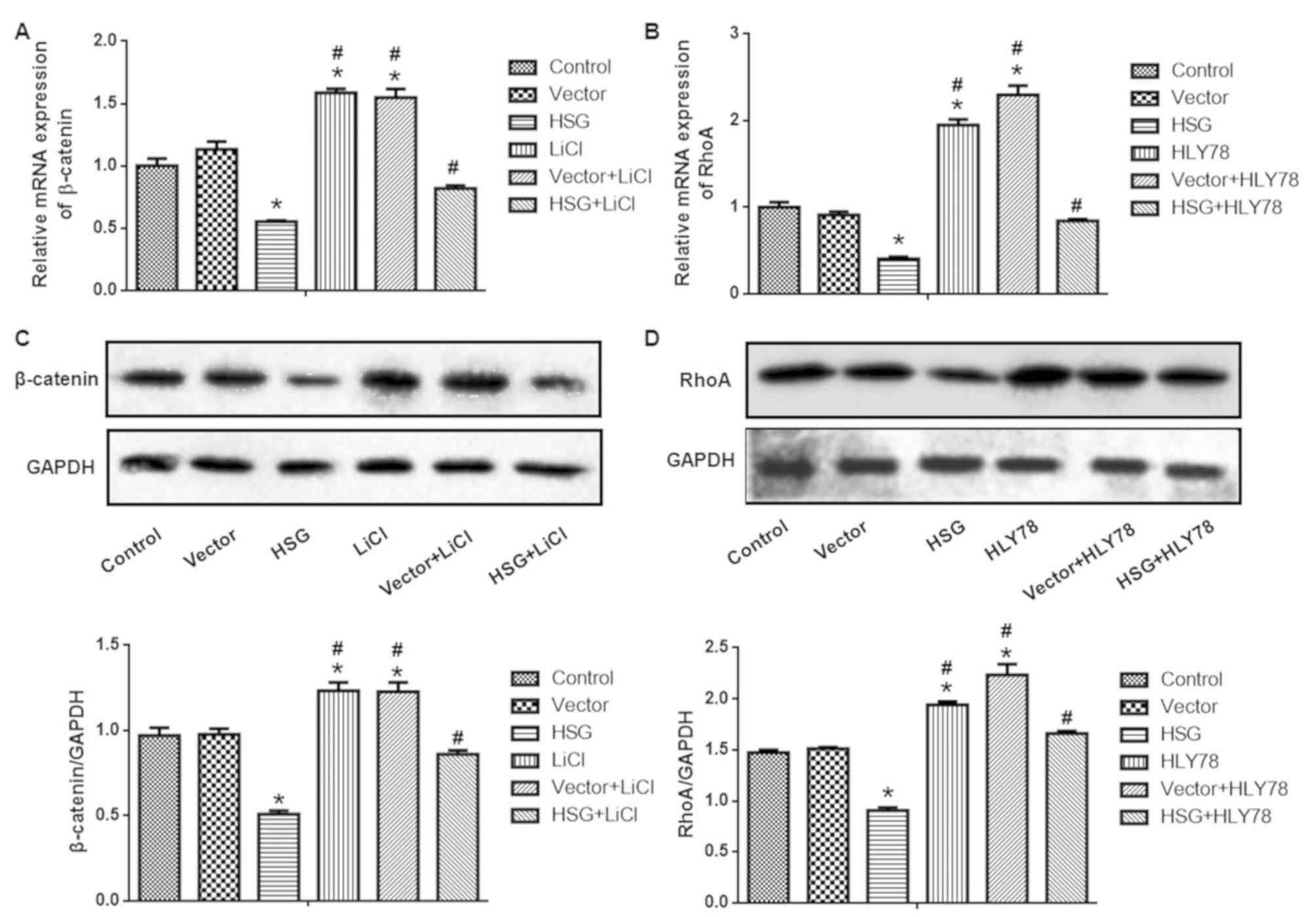
HSG overexpression reduces the
expression of β-catenin and RhoA
The expression of β-catenin and RhoA in each group
was normalized to GAPDH and is presented in Fig. 6. The mRNA levels of β-catenin
(Fig. 6A) and RhoA (Fig. 6B) were higher in the HSG group
compared with the vector group (both P<0.05). However, the
expression of β-catenin and RhoA was reduced by LiCl and HLY78,
respectively (vs. HSG, P<0.05). The protein expression of
β-catenin (Fig. 6C) and RhoA
(Fig. 6D) was also detected using
western blot analysis. Consistent with the mRNA expression, the
protein levels of β-catenin and RhoA (normalized to GAPDH) in the
HSG group were higher (vs. control, P<0.05), and the expression
of β-catenin and RhoA was reduced by LiCl and HLY78, respectively
(vs. HSG, P<0.05). The expression of β-catenin and RhoA was
promoted by LiCl and HLY78 alone, respectively (vs. control,
P<0.05).
Discussion
Although the effects of HSG on tumors are well
documented (10,11), the role of HSG in airway fibroblast
proliferation in COPD remains unknown. The present study
demonstrated that HSG overexpression reduced cell viability and
facilitated the apoptosis of airway fibroblasts in a rat model of
COPD. This mechanism involved inactivation of the Wnt signaling
pathway. These data may have important implications for the
treatment of airway remodeling in the future.
The pathogenesis of COPD is closely associated with
factors, including smoking, exposure to air pollution and
inhalation of dust (25,26). Pathological changes observed in
COPD include chronic bronchitis, emphysema, airway reconstruction
and pulmonary artery reconstruction (27). In the present study, the alveolar
wall became thinner, the number of alveolar septa was markedly
reduced, the alveolar cavity was enlarged and ruptured, and lung
ulcers formed following inhalation of papain. A previous study
reported that exogenous recombinant HSG may inhibit cell
proliferation and airway reconstruction (12). In the present study, it was further
reported that an exogenous HSG vector may reduce the viability, and
promote the apoptosis of airway fibroblasts. Collectively, these
studies indicated the importance of HSG as a potential target for
the treatment of airway remodeling in patients with COPD.
In the respiratory system, the Wnt signaling pathway
participates in the development of lung tissue and is involved in
pulmonary fibrosis (28). In the
present study, activators of the Wnt/β-catenin signaling pathways
were used to confirm the role of these pathways in the HSG-induced
decrease in cell viability and increase in apoptosis. These results
further indicated that Wnt signaling is involved in airway
remodeling of COPD (29–32).
TGF-β1 is a known target of the Wnt pathway and a
multifunctional protein (33). It
plays a role in cell growth, differentiation, apoptosis, and
regulation of the immune system (34–36).
TGF-β1 is widely distributed in the trachea and bronchus, and found
in infiltrating neutrophils, macrophages, and airway fibroblasts
during the progression of COPD (37). It is the most important fibrogenic
cytokine and can reverse-regulate and stimulate fibroblasts to
synthesize and secrete components of the extracellular matrix,
leading to airway fibrosis (38–40).
In the pathogenesis of COPD, MMP-9 mainly functions by degrading
extracellular components of the alveolar matrix and aggravating
alveolar cavity expansion, thereby resulting in decreased
elasticity and low retraction (41). On the other hand, MMP-9 is involved
in airway remodeling by destroying epithelial and endothelial cells
(42). In addition, MMP-9 is a
target of the Wnt pathway (30,43).
PDGF can stimulate the chemotaxis and proliferation of airway
fibroblasts and extracellular matrix synthesis, one of the
mechanisms of airway reconstruction (44). The results from the present study
showed that HSG regulates the expression of TGF-β1, MMP-9 and PDGF.
Additionally, the expression of these factors was regulated by the
Wnt/β-catenin and Wnt/PCP signaling pathways. These results
suggested that HSG may interfere with airway remodeling in
COPD.
Additionally, HSG can inhibit the expression of
MMP-9, PDGF, TGF-β1 in fibroblasts, thus inhibiting airway
remodeling (45). These regulatory
effects are mediated by the Wnt signaling pathway. In the present
study HSG was negatively associated with the Wnt/β-catenin and
Wnt/PCP signaling pathways, which further confirmed the association
between HSG and the Wnt signaling pathway. It can be concluded that
HSG inhibits the proliferation of airway fibroblasts by inhibiting
the abnormal activation of the Wnt signaling pathway. Therefore,
the promotion of HSG expression, or inhibition of abnormal
activation of the Wnt/PCP signaling pathway to inhibit the
overexpression of MMP-9, PDGF and TGF-β1, may be an effective
therapeutic strategy for airway remodeling in COPD. Although HSG
overexpression inactivated the Wnt signaling pathway, the specific
mechanisms could not be verified. A previous study demonstrated
that PTEN, another tumor suppressor gene, interacts with Wnt1 to
regulate the Wnt signaling pathway (46). Whether HSG is directly or
indirectly linked to the Wnt signaling pathway requires further
investigation.
The present study has certain limitations. First,
the majority of the experiments were conducted in the cellular
model, although airway fibroblasts were collected from COPD rats.
In vivo experiments are required to confirm the present
results. Second, an association was established between HSG and the
Wnt signaling pathway in apoptosis; however, the exact mechanism of
regulation requires further investigation. In addition, the
contribution of autophagy and other types of cell death should also
be determined in future studies.
In conclusion, our data demonstrated that HSG
overexpression inactivates airway fibroblasts via regulation of the
Wnt signaling pathway. This may represent a potential therapeutic
target for COPD.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81473533) and the
Major Basic Research in Guizhou Province [grant no. (2015)
2002].
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZG and YY conceived and designed the experiments;
ZG, YY, XZ, JZ, BL, XW and XL performed the experiments and
analyzed the data; ZG and YY wrote the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of Guizhou College of Traditional Chinese Medicine
(Guiyang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yu ZW, Xu YQ, Zhang XJ, Pan JR, Xiang HX,
Gu XH, Ji SB and Qian J: Mutual regulation between miR-21 and the
TGFβ/Smad signaling pathway in human bronchial fibroblasts promotes
airway remodeling. J Asthma. 56:341–349. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lai T, Tian B, Cao C, Hu Y, Zhou J, Wang
Y, Wu Y, Li Z, Xu X, Zhang M, et al: HDAC2 suppresses
IL17A-mediated airway remodeling in human and experimental modeling
of COPD. Chest. 153:863–875. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ricciardolo FLM, Folkerts G, Folino A and
Mognetti B: Bradykinin in asthma: Modulation of airway inflammation
and remodelling. Eur J Pharmacol. 827:181–188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eapen MS, Myers S, Lu W, Tanghe C, Sharma
P and Sohal SS: sE-cadherin and sVE-cadherin indicate active
epithelial/endothelial to mesenchymal transition (EMT and EndoMT)
in smokers and COPD: Implications for new biomarkers and
therapeutics. Biomarkers. 23:709–711. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mahmood MQ, Reid D, Ward C, Muller HK,
Knight DA, Sohal SS and Walters EH: Transforming growth factor
(TGF) β1 and Smad signalling pathways: A likely key to
EMT-associated COPD pathogenesis. Respirology. 22:133–140. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun ZQ, Chen G, Guo Q, Li HF and Wang Z:
In vivo and in vitro effects of hyperplasia suppressor gene on the
proliferation and apoptosis of lung adenocarcinoma A549 cells.
Biosci Reports. 38:BSR201803912018. View Article : Google Scholar
|
|
7
|
Luo L, Gong YQ, Qi X, Lai W, Lan H and Luo
Y: Effect of tumor suppressor PTEN gene on apoptosis and cell cycle
of human airway smooth muscle cells. Mol Cell Biochem. 375:1–9.
2013.PubMed/NCBI
|
|
8
|
Jiang GJ, Han M, Zheng B and Wen JK:
Hyperplasia suppressor gene associates with smooth muscle
alpha-actin and is involved in the redifferentiation of vascular
smooth muscle cells. Heart Vessels. 21:315–320. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guo YH, Li Q, Yu HY and Gao W: Hyperplasia
suppressor gene induces vascular smooth muscle cell apoptosis.
Beijing Da Xue Xue Bao Yi Xue Ban. 39:394–398. 2007.(In Chinese).
PubMed/NCBI
|
|
10
|
Wu L, Li Z, Zhang Y, Zhang P, Zhu X, Huang
J, Ma T, Lu T, Song Q, Li Q, et al: Adenovirus-expressed human
hyperplasia suppressor gene induces apoptosis in cancer cells. Mol
Cancer Ther. 7:222–232. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang Y, Du Q, Qiu XY, Tian XX and Fang
WG: Over expression of hyperplasia suppressor gene inhibits the
malignant phenotype of breast cancer cell. Zhonghua Bing Li Xue Za
Zhi. 39:259–263. 2010.(In Chinese). PubMed/NCBI
|
|
12
|
Zheng-Xing GE, Bo LI, Zhou X and Chang LI:
rHSG gene regulates airway fibroblast proliferation and apoptosis
of COPD rats. Basic Clin Med. 33:1235–1241. 2013.
|
|
13
|
Villar J, Cabrera NE, Valladares F, Casula
M, Flores C, Blanch L, Quilez ME, Santana-Rodríguez N, Kacmarek RM
and Slutsky AS: Activation of the Wnt/β-catenin signaling pathway
by mechanical ventilation is associated with ventilator-induced
pulmonary fibrosis in healthy lungs. PLoS One. 6:e239142011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chilosi M, Poletti V, Zamò A, Lestani M,
Montagna L, Piccoli P, Pedron S, Bertaso M, Scarpa A, Murer B, et
al: Aberrant Wnt/beta-catenin pathway activation in idiopathic
pulmonary fibrosis. Am J Pathol. 162:1495–1502. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Königshoff M, Balsara N, Pfaff EM, Kramer
M, Chrobak I, Seeger W and Eickelberg O: Functional Wnt signaling
is increased in idiopathic pulmonary fibrosis. PLoS One.
3:e21422008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Peng Y, Zhang X, Ma Q, Yan R, Qin Y, Zhao
Y, Cheng Y, Yang M, Wang Q, Feng X, et al: MiRNA-194 activates the
Wnt/β-catenin signaling pathway in gastric cancer by targeting the
negative Wnt regulator, SUFU. Cancer Lett. 385:117–127. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qu J, Yue L, Gao J and Yao H: Perspectives
on Wnt signal pathway in the pathogenesis and therapeutics in
chronic obstructive pulmonary disease. J Pharmacol Exp Ther.
369:473–480. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tao H, Yang JJ, Shi KH and Li J: Wnt
signaling pathway in cardiac fibrosis: New insights and directions.
Metabolism. 65:30–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nishikawa K, Osawa Y and Kimura K:
Wnt/β-catenin signaling as a potential target for the treatment of
liver cirrhosis using antifibrotic drugs. Int J Mol Sci.
19:E31032018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lewis CC, Chu HW, Westcott JY, Tucker A,
Langmack EL, Sutherland ER and Kraft M: Airway fibroblasts exhibit
a synthetic phenotype in severe asthma. J Allergy Clin Immunol.
115:534–540. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song Z, Chen H, Xu W, Wu S and Zhu G:
Basolateral amygdala calpain is required for extinction of
contextual fear-memory. Neurobiol Learn Mem. 155:180–188. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu G, Wang X, Wu S and Li Q: Involvement
of activation of PI3K/Akt pathway in the protective effects of
puerarin against MPP+-induced human neuroblastoma SH-SY5Y cell
death. Neurochem Int. 60:400–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aigon A and Billecocq S: Prevalence and
impact on quality of life of urinary incontinence in an adult
population with chronic obstructive pulmonary diseases, literature
review. Prog Urol. 28:962–972. 2018.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oshagbemi OA, Keene SJ, Driessen JHM,
Jordan R, Wouters EFM, de Boer A, de Vries F and Franssen FME:
Trends in moderate and severe exacerbations among COPD patients in
the UK from 2005 to 2013. Respir Med. 144:1–6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moon JY, Leitao Filho FS, Shahangian K,
Takiguchi H and Sin DD: Blood and sputum protein biomarkers for
chronic obstructive pulmonary disease (COPD). Expert Rev
Proteomics. 15:923–935. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Reuter S, Beckert H and Taube C: Take the
Wnt out of the inflammatory sails: Modulatory effects of Wnt in
airway diseases. Lab Invest. 96:177–185. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bartel S, Carraro G, Alessandrini F,
Krauss-Etschmann S, Ricciardolo FLM and Bellusci S: miR-142-3p is
associated with aberrant WNT signaling during airway remodeling in
asthma. Am J Physiol Lung Cell Mol Physiol. 315:L328–L333. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Royer PJ, Henrio K, Pain M, Loy J, Roux A,
Tissot A, Lacoste P, Pison C, Brouard S and Magnan A; COLT
consortium, : TLR3 promotes MMP-9 production in primary human
airway epithelial cells through Wnt/β-catenin signaling. Respir
Res. 18:2082017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Koopmans T and Gosens R: Revisiting asthma
therapeutics: Focus on WNT signal transduction. Drug Discov Today.
23:49–62. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hussain M, Xu C, Lu M and Wu X, Tang L and
Wu X: Wnt/β-catenin signaling links embryonic lung development and
asthmatic airway remodeling. Biochim Biophys Acta Mol Basis Dis.
1863:3226–3242. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vallée A, Lecarpentier Y, Guillevin R and
Vallée JN: Interactions between TGF-β1, canonical WNT/β-catenin
pathway and PPAR γ in radiation-induced fibrosis. Oncotarget.
8:90579–90604. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Si Y, Bai J, Wu J, Li Q, Mo Y, Fang R and
Lai W: LncRNA PlncRNA1 regulates proliferation and differentiation
of hair follicle stem cells through TGFbeta1mediated
Wnt/betacatenin signal pathway. Mol Med Rep. 17:1191–1197.
2018.PubMed/NCBI
|
|
35
|
Ma F, Li W, Liu C, Li W, Yu H, Lei B, Ren
Y, Li Z, Pang D and Qian C: MiR-23a promotes TGF-β1-induced EMT and
tumor metastasis in breast cancer cells by directly targeting CDH1
and activating Wnt/β-catenin signaling. Oncotarget. 8:69538–69550.
2017.PubMed/NCBI
|
|
36
|
Li M, Yuan Y, Chen Q, Me R, Gu Q, Yu Y,
Sheng M and Ke B: Expression of Wnt/β-catenin signaling pathway and
its regulatory role in type I collagen with TGF-β1 in scleral
fibroblasts from an experimentally induced myopia guinea pig model.
J Ophthalmol. 2016:51265602016.PubMed/NCBI
|
|
37
|
Godinas L, Corhay JL, Henket M, Guiot J,
Louis R and Moermans C: Increased production of TGF-β1 from sputum
cells of COPD: Relationship with airway obstruction. Cytokine.
99:1–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Westergren-Thorsson G, Bagher M,
Andersson-Sjoland A, Andersson-Sjöland A, Thiman L, Löfdahl CG,
Hallgren O, Bjermer L and Larsson-Callerfelt AK: VEGF synthesis is
induced by prostacyclin and TGF-beta in distal lung fibroblasts
from COPD patients and control subjects: Implications for pulmonary
vascular remodelling. Respirology. 23:68–75. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen H, Zhang R, Zheng Q, Yan X, Wu S and
Chen Y: Impact of body mass index on long-term blood pressure
variability: A cross-sectional study in a cohort of Chinese adults.
BMC Public Health. 18:11932018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Di Stefano A, Sangiorgi C, Gnemmi I,
Casolari P, Brun P, Ricciardolo FLM, Contoli M, Papi A, Maniscalco
P, Ruggeri P, et al: TGF-β signaling pathways in different
compartments of the lower airways of patients with stable COPD.
Chest. 153:851–862. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhdan VN, Potyazhenko MМ, Khaymenova GS,
Lyulka NN, Dubrovinskaya TV and Ivanitsky IV: Intensifying approach
to the therapy of patients with constellation of the diseases:
Chronic obstructive pulmonary disease and osteoarthritis. Wiad Lek.
70:578–580. 2017.PubMed/NCBI
|
|
42
|
Zhao Y, Qiao X, Wang L, Tan TK, Zhao H,
Zhang Y, Zhang J, Rao P, Cao Q, Wang Y, et al: Matrix
metalloproteinase 9 induces endothelial-mesenchymal transition via
Notch activation in human kidney glomerular endothelial cells. BMC
Cell Biol. 17:212016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang Z, Li K, Liang Q, Zheng G, Zhang S,
Lao X, Liang Y and Liao G: Elevated hydrostatic pressure promotes
ameloblastoma cell invasion through upregulation of MMP-2 and MMP-9
expression via Wnt/β-catenin signalling. J Oral Pathol Med.
47:836–846. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu J, Zhu Y, Feng W, Pan Y, Li S, Han D,
Liu L, Xie X, Wang G and Li M: Platelet-derived growth factor
mediates interleukin-13-induced collagen I production in mouse
airway fibroblasts. J Biosci. 39:693–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Song S, Zhang M, Yi Z, Zhang H, Shen T, Yu
X, Zhang C, Zheng X, Yu L, Ma C, et al: The role of
PDGF-B/TGF-β1/neprilysin network in regulating
endothelial-to-mesenchymal transition in pulmonary artery
remodeling. Cell Signal. 28:1489–1501. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao H, Cui Y, Dupont J, Sun H,
Hennighausen L and Yakar S: Overexpression of the tumor suppressor
gene phosphatase and tensin homologue partially inhibits
wnt-1-induced mammary tumorigenesis. Cancer Res. 65:6864–6873.
2005. View Article : Google Scholar : PubMed/NCBI
|