Introduction
Spinal cord injury (SCI) is a severe and traumatic
disease; due to of the high rates of disability and fatality, it
results in a series of personal and social problems. The data of
the National SCI database show that the current prevalence of SCI
in the US has already reached 63,109 patients, with an estimated
annual incidence of ~54 cases per million in 2012 (1). In addition, functional deficits
always accompany a SCI, and only 0.4% of patients experience
complete neurological recovery. This is due to the fact that axons
fail to regenerate after SCI (2–4).
This failure to regenerate can be attributed to two causes: Mature
neurons exhibiting a low intrinsic ability to regrow axons and the
extrinsic environment determines the projection of the axon
regeneration after SCI (5–11). In addition, mature neurons have a
low intrinsic ability to regrow axons (12,13).
In past decades, a number of studies (2,14,15)
have focused on characterizing environmental inhibitory molecules
in the adult central nervous system. However, blocking the
inhibitory activities of chondroitin sulfate proteoglycans (CSPGs)
and myelin-associated molecules in the glial scar via either
genetic or pharmacological means allows for the regrowth of some
injured axons, even though their regeneration is limited (14,15).
This evidence suggests that removing inhibitory activities alone is
not sufficient to allow the majority of injured axons to regenerate
and gain adequate functional recovery. It is important to
understand the underlying mechanisms of both the diminished
intrinsic growth and axon regenerative abilities of neurons.
Numerous studies (13,16–18)
have indicated that manipulating certain signaling pathways,
including the PTEN/mTOR, Janus kinase/STAT, dual leucine zipper
kinase/JNK and suppressor of cytokine signaling 3/STAT3 pathways,
may allow certain populations of mature central nervous system
(CNS) neurons to mount regenerative growth after injury. However,
the primary cellular substrate of PTEN is the second messenger
phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which transmits
growth and survival signals (16).
PTEN removes the D3 phosphate from PIP3, inactivating the
PI3K/Akt/mTOR pathway and generating phosphatidylinositol (4,5)-bisphosphate (PIP2), which does not
function in the same manner as PIP3 (19–21).
In a previous study, Park et al (22) found that the regeneration and
survival abilities of a retinal ganglion cell with PTEN gene
knockdown were superior to those of normal retinal ganglion cells,
and were correlated with the content of mTOR. Our previous study
(23) also demonstrated that PTEN
silencing using short hairpin RNA (shRNA) promoted neurite
elongation and motor function improvement in a rat model of
SCI.
In the present study, an inhibitory microenvironment
of SCI was constructed in vitro. An inhibitor with a high
inhibition efficiency targeted against the PTEN/mTOR signaling
pathway was used to explore the mechanism of axon
growth/regeneration promotion. As PTEN also affects apoptosis in a
number of cell types, the effects of PTEN on neuronal apoptosis
were also explored.
Materials and methods
Animal subjects and ethics
statement
A total of 24 new born Wistar rats (5–6 g) were
provided by the Radiation Study Institute-Animal Center at Tianjin
Medical University. All experimental procedures involving animals
were approved by the Ethics Committee of Tianjin Medical University
and strictly complied with the Ethical Principles for the
Maintenance and Use of Animals In Neuroscience Research (24).
Neuron isolation and culture
In brief, forebrain cortices from postnatal day 0
(P0) Wistar rats were dissected under a stereomicroscope (LEICA
M501; Leica Microsystems GmbH) and dissociated into a single-cell
suspension through enzymatic digestion (Papain and DNase I;
Worthington Biochemical Corporation) and mechanical pipetting.
After centrifugation for 5 min at 200 × g and 4°C, the cells were
resuspended at a density of 6×105 cells/ml in fresh
plating medium [DMEM-high containing 10% FBS (both Gibco; Thermo
Fisher Scientific, Inc.) and 1% (vol/vol) penicillin/streptomycin
(Sigma-Aldrich; Merck KGaA)]. The cells were cultured in culture
plates (BD Falcon; BD Biosciences) coated with 0.01% poly-L-lysine
(PLL; Sigma-Aldrich; Merck KGaA) at 37°C in a humidified incubator
with 5% CO2. The plating medium was replaced by
serum-free medium [Neurobasal containing 10 ng/ml neuronal growth
factor, 2% (vol/vol) B27 supplement, 0.5 mM L-glutamine (all Gibco;
Thermo Fisher Scientific, Inc.), 0.5% (vol/vol) D-glucose and 0.5%
(vol/vol) penicillin/streptomycin (Sigma-Aldrich; Merck KGaA)] 4 h
later. Half of the serum-free medium was replaced every 3 days. A
primary antibody against β-tubulin III (1:500; Abcam, ab18207) was
applied as a specific axonal marker to identify the neurons. In
addition, Hoechst 33342 (1 µg/ml; Invitrogen; Thermo Fisher
Scientific, Inc.) was used to visualize the nuclei of all cells in
TUNEL staining.
Preliminary specific inhibitors
efficiency assay
The inhibitor of PTEN dipotassium bisperoxo
(picolinato) oxovanadate [bpV(pic); Sigma-Aldrich; Merck KGaA] was
reconstituted in ddH2O for a 500-µM stock; different
concentrations (100, 300, 500, and 700 nM) were tested (data not
shown) and the final concentration used was 500 nM. Inhibitive
efficiency of bpV(pic) was still lower than that of control group
at day 14 (data not shown). The highly selective inhibitor of PI3K
LY294002 (Cell Signaling Technology, Inc.) was reconstituted in
DMSO for a 10-mM stock; the final concentration used was 50 µM. The
inhibitor of mTOR ridaforolimus (Santa Cruz Biotechnology, Inc.)
was reconstituted in DMSO for a 100-µM stock; the final
concentration used was 100 nM. To evaluate the efficiency of the
inhibitors of the PTEN/Akt/mTOR signaling pathway, the neurons were
separated into four treatment groups [control, LY294002 + bpV(pic),
ridaforolimus + bpV(pic) and bpV(pic)]. Half of the culture medium
was replaced every 3 days. These samples were collected for western
blot analysis at day 7, based on a phosphorylation pattern study.
In addition, primary antibodies for Akt (cat. no. 4691, 1:1,000),
phosphorylated (p-)Akt (cat. no. 4060, 1:1,000), mTOR (cat. no.
2983, 1:1,000), p-mTOR (cat. no. 5536, 1:1,000), p70-S6 kinase 1
(p70S6K; cat. no. 97596, 1:1,000) and p-p70S6K (cat. no. 97596,
1:1,000; all Cell Signaling Technology, Inc.) were used in this
procedure at 4°C overnight.
Plating preparation
To explore the effect of specific inhibitors on
axonal growth, 6-well plates were coated with 0.01% PLL overnight.
The next day, they were washed three times with PBS and dried at
37°C. Then, 3-µl droplets of CSPGs (50 µg/ml; EMD Millipore) were
spotted onto the 6-well plates for 4 h at 37°C. After the droplets
dried, the six-well plates were covered with laminin(LN) (10 µg/ml;
Invitrogen) for 2 h at 37°C. They were then washed three times with
PBS and stored at 37°C before neuron plating.
Immunocytochemistry
Immunocytochemistry was performed as described
previously (25). Neurons were
treated with the various inhibitors for 3 days, then the medium was
changed. Cytochemistry was performed on day 7, and all
cytochemistry was performed on cells cultured on CSPGs-coated
plates. After 10 days of culture, the four groups of neurons were
fixed with 4% PFA in PBS for 5 min and incubated in 0.3% Triton
X-100 (Sigma-Aldrich; Merck KGaA) for 5 min and blocking solution
containing 10% goat serum (OriGene Technologies, Inc.) in PBS for
60 min all these steps are at room temperature. The primary
antibodies, mouse (IgM) anti-CSPG (clone CS-56; 1:500;
Sigma-Aldrich; Merck KGaA, Cat no. SAB2100493) and rabbit
anti-β-tubulin III (1:100; Abcam, Cat no. ab18207), were used to
identify CSPGs and neurons, respectively. All cultures were stained
with primary antibodies overnight at 4°C. The cultures were then
washed three times and stained with the secondary antibodies
AMCA-conjugated goat anti-mouse IgM (Jackson ImmunoResearch cat.
no. 115-155-020) and TRITC-conjugated goat anti-rabbit IgG (OriGene
Technologies, Inc. cat. no. S0015) for 1 h at room temperature.
Hoechst 33342 was used to visualize the nuclei for 10 min at room
temperature.
Quantification of neurites
Neurite length and the extent of branching were
calculated from images of at least 120 neurons for each culture
condition that were acquired under a fluorescence microscope
(magnification, ×20; IX71; Olympus Corporation). Neurites extending
over the CSPG substrate were traced and measured using the
Image-Pro Plus 7.0 software (Media Cybernetics, Inc.) for each
field. Maximum neurite length was defined as the length of the
longest continuous neurite. The number of primary neurites was
defined as the number of neurites that were extending from the cell
body. In light of the enhanced crossing ability onto CSPGs, only
individually identified axons were analyzed, and the percentage of
neurites that crossed the CSPGs border was calculated.
Western blot analysis
After 7 days, the cells were placed for 30 min in
culture medium containing a final concentration of 100 µM
H2O2 at 37°C to induce apoptosis of neurons.
Western blotting analysis was performed as described previously
(26), with minor modifications.
The total protein of each treatment group was extracted from
neurons on the 10-cm dishes using RIPA buffer (Santa Cruz
Biotechnology, Inc.) and protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA) on ice. A bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.) was used to
measure the protein concentration. Equal amounts of the samples (20
µg) were separated via 10% SDS-PAGE. Proteins were transferred to
PVDF membranes for 1 h at 100 V and blocked with 5% milk for 1 h at
room temperature. Membranes were then incubated with the primary
antibodies [Bcl-2, 1:1,000, cat. no. 3498 and native caspase-3,
1:1,000, cat. no. 9662 (Cell Signaling Technology, Inc.); PTEN,
1:1,000; cat. no. 9188 (Cell Signaling Technology, Inc.] in
blocking solutions at 4°C overnight before detection with
HRP-conjugated secondary antibodies. Immunoreactive bands were
visualized on film by enhanced chemiluminescent substrate (Pierce,
Thermo Fisher Scientific, Inc.) ImagePro Plus version 7.0 (Media
Cybernetics) software was used to calculate the density.
TUNEL staining for
H2O2-mediated apoptosis
After 7 days, cells were placed for 30 min in
culture medium containing a final concentration of 100 µM
H2O2. Then, in 4% paraformaldehyde in PBS for
5 min at room temperature for fixation. Fragmented DNA in apoptotic
cells was quantified using a TUNEL System (Roche Diagnostics) on
96-well plates. TUNEL reagent was added to fixed neurons for 60 min
at 37°C in the dark. Fluorescein-12-dUTP-labelled DNA in the cortex
was visualized under a fluorescence microscope (magnification, ×10,
IX71; Olympus Corporation). The nuclei were counterstained with
Hoechst 33342 (1 µg/ml; Invitrogen; Thermo Fisher Scientific,
Inc.). Hoechst 33342 was added to the neurons for 10 min at room
temperature. For apoptotic cells, positive green fluorescent
apoptotic nuclei in three view fields that were selected randomly
in each plate were examined. ImagePro Plus version 7.0 image
analysis software (Media Cybernetics, Inc.) was used to calculate
the percentage of neuronal apoptosis. The average value was taken
as the final result.
Statistical analysis
All statistical analyses were performed using SPSS
18.0 software (SPSS, Inc.). All data are presented as the mean ±
standard error of mean, and χ2 test or one-way analysis
of variance (ANOVA) was used for comparisons among the four groups.
For all ANOVA measures, Tukey's test was used following ANOVA;
groups were treated as the independent variable and the levels of
the outcome variables as the dependent variables. P<0.05 was
considered to indicate a statistically significant difference.
Results
Culture and identification of
forebrain cortical neurons
The procedures of isolation and neuron seeding were
completed within 4 h of animal sacrifice. The plating medium was
replaced with serum-free medium 4 h later. After 7 days of culture,
immunocytochemistry was performed to identify the cerebral cortical
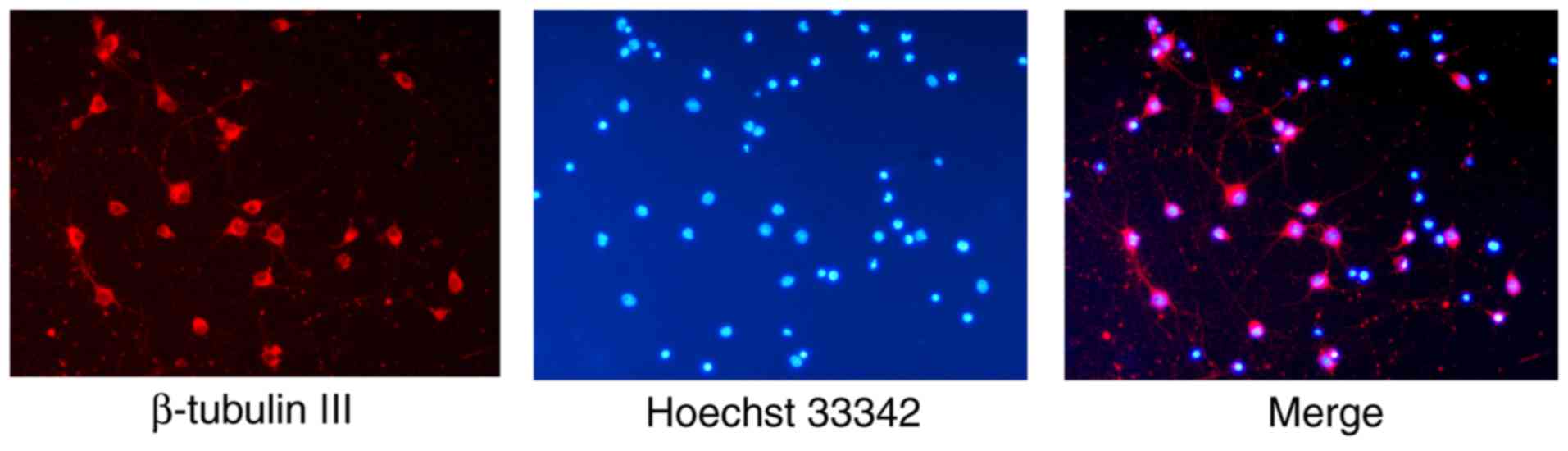
neurons (Fig. 1). The neurons were
immunostained red by β-tubulin III, a widely used neuron marker
(27–29), and their nuclei were stained blue
by Hoechst 33342. The purity of the neurons in the cultures
(proportion of β-tubulin III positive neurons) was 94.78±1.56%.
Efficiency of specific inhibitors of
the PTEN/Akt/mTOR signaling pathway
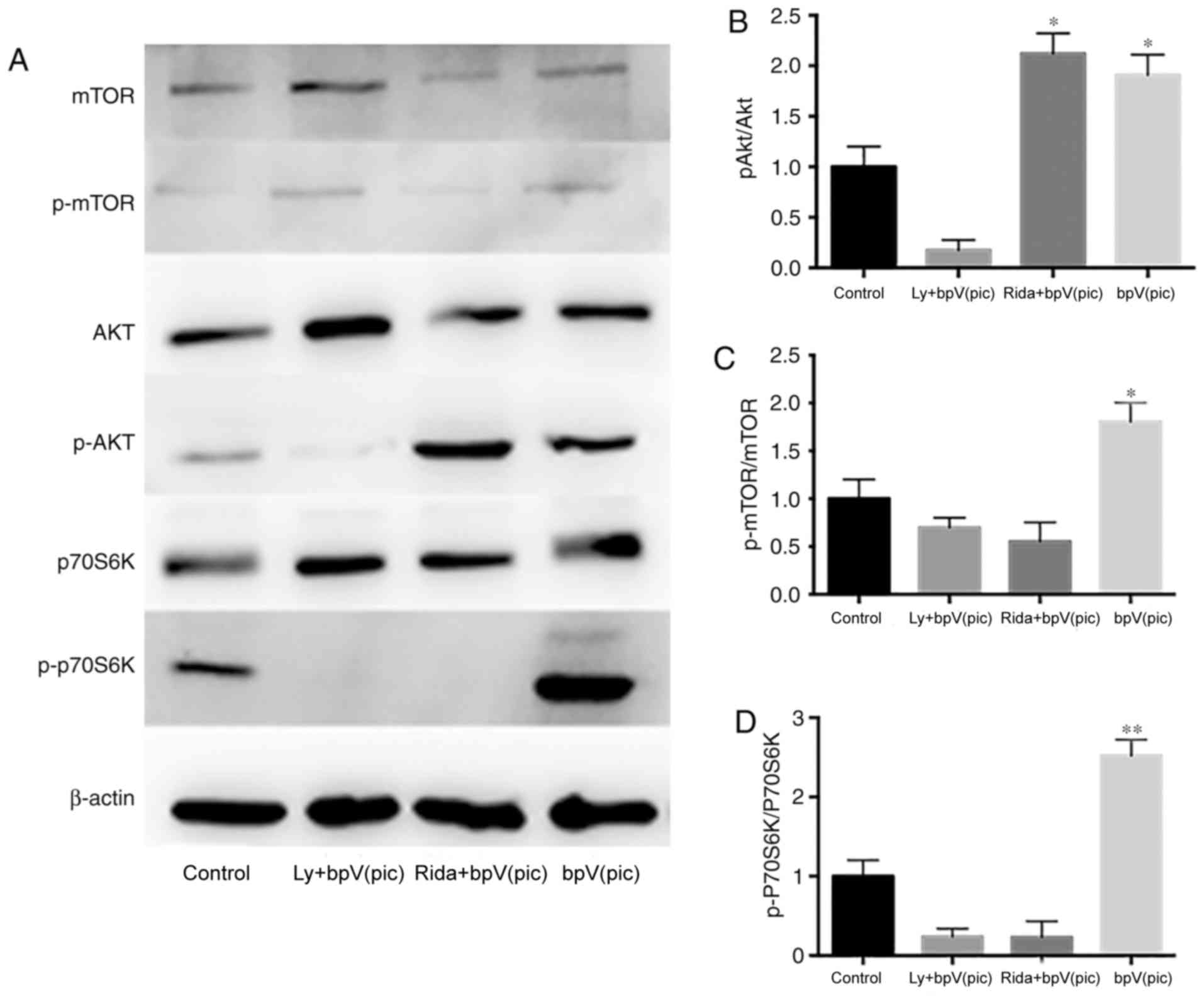
After 7 days of culture, samples from the four
treatment groups were collected for western blot analysis (Fig. 2). The inhibition efficiency of
bpV(pic) was detected, and it was demonstrated that it could
effectively inhibit the expression of PTEN (Fig. S1). After adding PTEN inhibitor
bpV(pic), the phosphorylation levels of Akt and p70S6K
(Thr421/Ser424) of the bpV(pic) group were increased significantly
compared with the control group (Fig.
2). In addition, the phosphorylation levels of mTOR were
slightly increased. No clear dose-dependent effects of bpV(pic)
were observed. Neurons were cultured to analyze the inhibitive
efficiency of bpV(pic) at the day 14 time point, and it was
demonstrated that the inhibition was still effective. In the
LY294002 + bpV(pic) group, the high expression levels of p-Akt and
p-p70S6K (Thr421/Ser424) induced by bpV(pic) were eliminated by
LY294002. The p-mTOR/mTOR ratio decreased compared with the control
group, as LY294002 inhibited the kinase activity of PI3K on mTOR.
The levels of total mTOR and Akt were markedly increased in the
LY+bpv group compared with control. In the ridaforolimus + bpV(pic)
group, the relative expression of p-Akt was increased; however, the
phosphorylation levels of mTOR and p70S6K(Thr421/Ser424) were
significantly decreased.
Effects of specific inhibitors on axon
growth
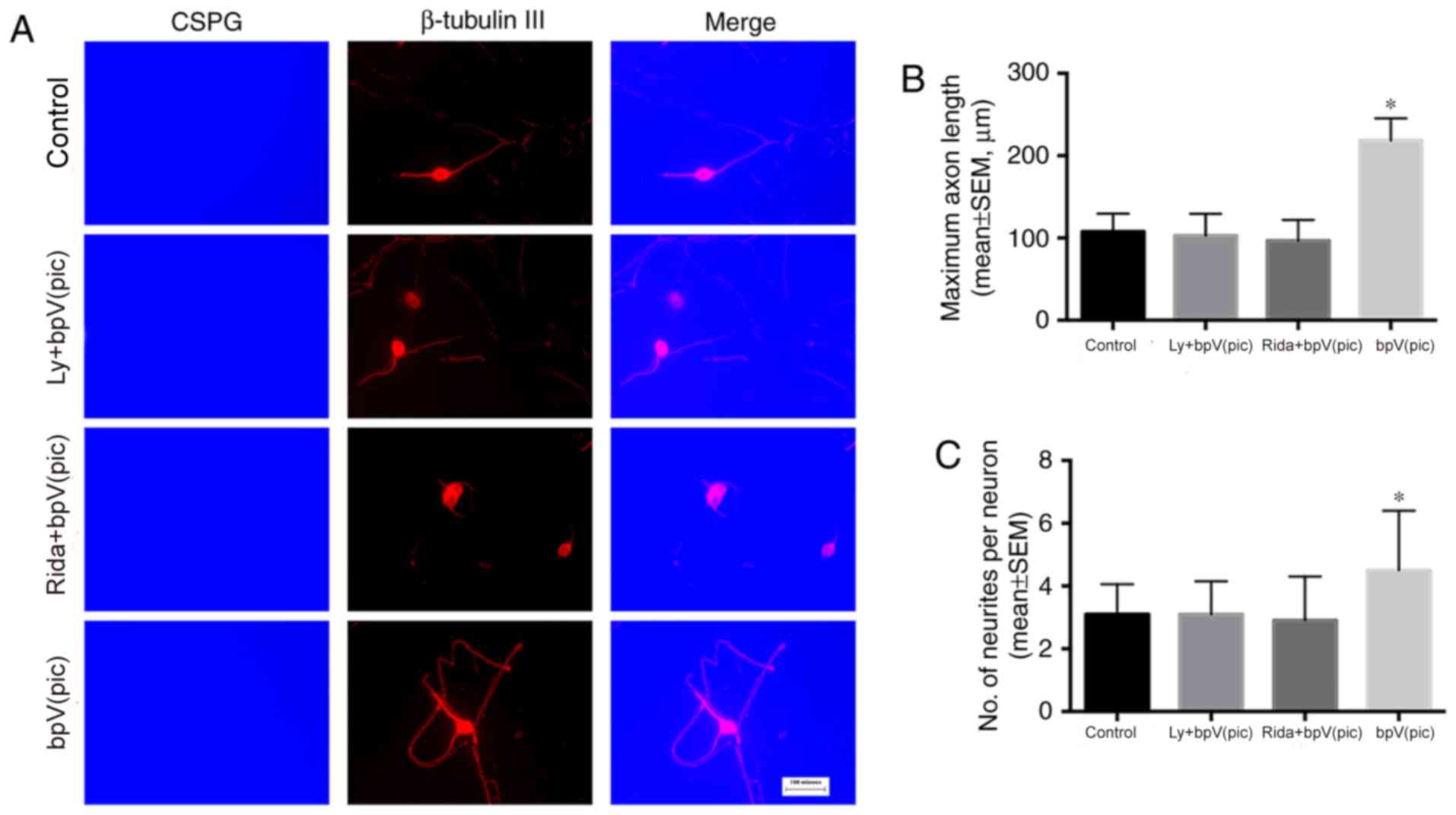
The neurons were seeded on a surface coated with PLL
+ CSPGs + LN. After plating, the medium was replaced with
serum-free medium, and the inhibitors were added to the medium.
After 10 days of culture, immunocytochemistry was performed to
evaluate the neurite initiation and elongation abilities in the
presence of CSPGs. Mouse anti-CSPG and rabbit anti-β-tubulin III
antibodies were used to identify CSPG and neurons, respectively.
The mean maximum axon lengths were 107.78±21.95, 102.92±26.51,
96.55±25.37 and 218.57±26.77 µm in the control, LY294002 +
bpV(pic), ridaforolimus + bpV(pic) and bpV(pic) groups,
respectively (Fig. 3A and B); a
significant increase was observed in the bpV(pic) group compared
with control (one-way ANOVA, P<0.001). The number of primary
neurites was 3.1±0.96, 3.1±1.05, 2.9±1.40 and 4.5±1.90 in the
control, LY294002 + bpV(pic), ridaforolimus + bpV(pic) and bpV(pic)
groups, respectively (Fig. 3A and
C). The number of neurites was significantly increased in the
bpV(pic) group compared with control (one-way ANOVA, P<0.001).
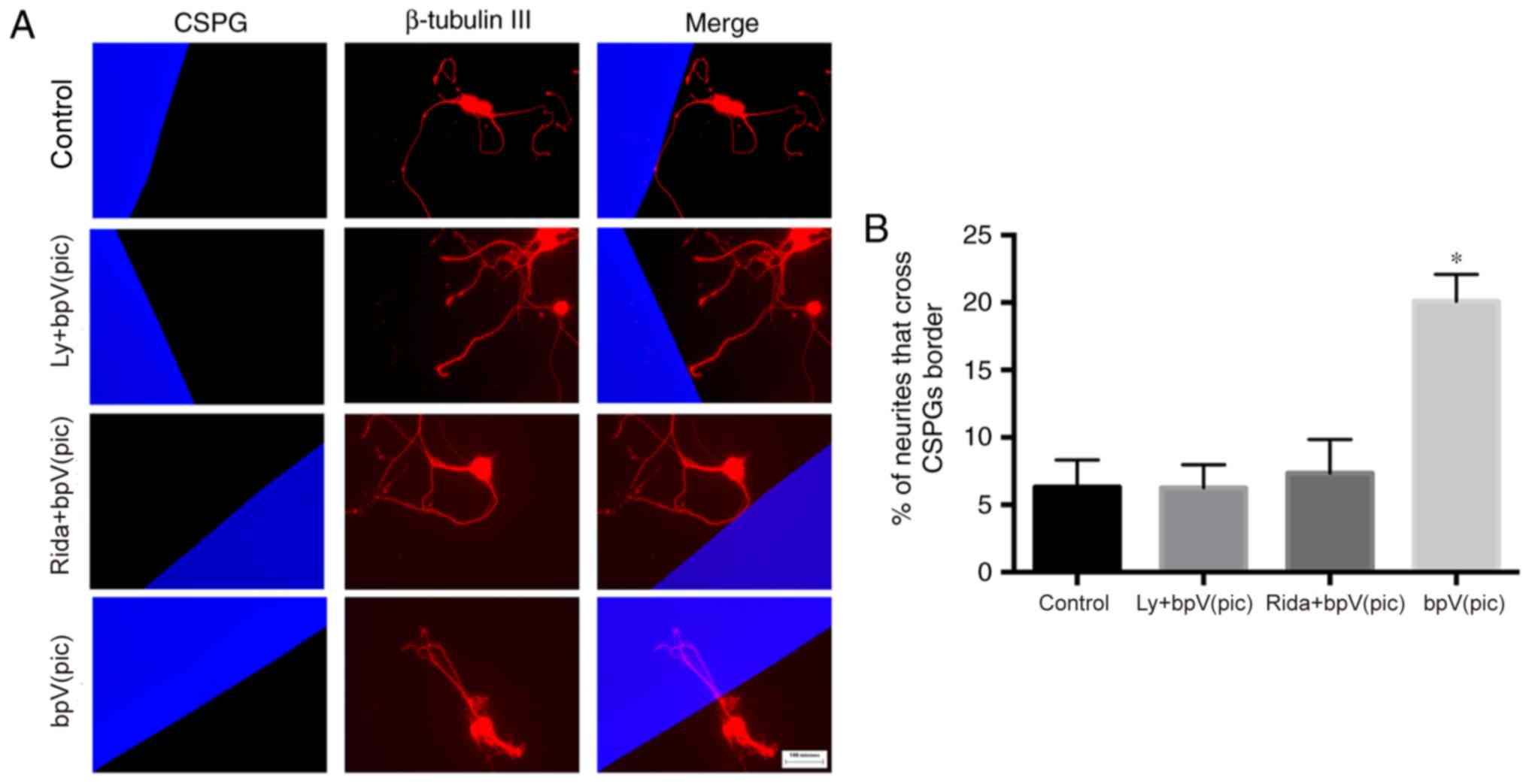
The percentage of neurites that crossed the CSPG border was
6.32±2.01%, 6.25±1.71, 7.34±2.51 and 20.1±1.99% in the four groups
(Fig. 4). The percentage of
neurites that crossed the CSPG border was significantly increased
in the bpV(pic) group compared with control (one-way ANOVA,
P<0.01).
Effects of specific inhibitors on
H2O2-mediated apoptosis
After the plating medium was replaced with
serum-free medium, the inhibitors were added to the medium. After 7
days of culture, cells were incubated in culture medium containing
100 µM H2O2 for 30 min. Western blotting and
TUNEL staining were performed to explore the effects of specific
inhibitors on neuronal apoptosis. Using western blotting analysis
(Fig. 5), it was determined that
Bcl-2 protein exhibited a higher level of expression in the
ridaforolimus + bpV(pic) and bpV(pic) groups, and reduced
expression in the LY294002 + bpV(pic) group compared with the
control group. The expression of caspase-3 was not significantly
different across the four groups.
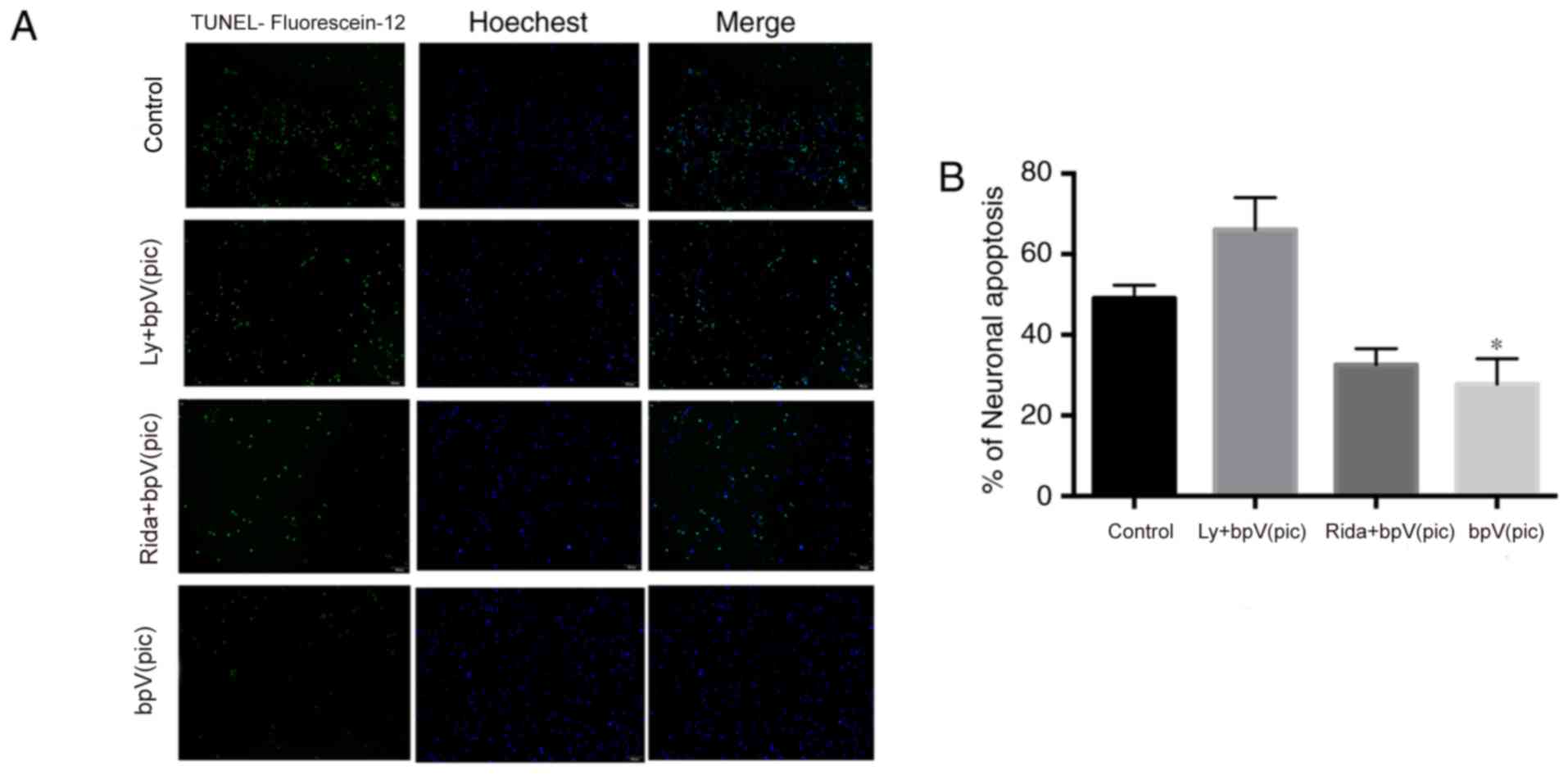
Apoptosis was further analyzed using TUNEL staining;
cells and apoptotic cells were stained blue and green,
respectively. The percentage of neuronal apoptosis was 49.1±3.2,
66.1±7.9, 32.6±4.0 and 27.8±6.3% in the control, LY294002 +
bpV(pic), ridaforolimus + bpV(pic) and bpV(pic) groups,
respectively (Fig. 6). The
percentage of neuronal apoptosis was significantly reduced in the
ridaforolimus + bpV(pic) and bpV(pic) groups compared with control
(one-way ANOVA, P<0.05).
Discussion
Studies (15) in
the last few decades have focused on characterizing environmental
inhibitory molecules in the adult central nervous system. Our
previous study (23) showed that
an shRNA against PTEN promoted neurite outgrowth of cortical
neurons and functional recovery in rats following spinal cord
contusion. The present study explored the role of the PTEN/Akt/mTOR
signaling pathway in axonal growth/regeneration and neuronal
apoptosis in the presence of CSPGs-mediated inhibition.
PTEN is an important potent tumor suppressor and has
been found to be mutated in the development of various cancers,
including hepatocellular carcinoma and lung cancer (30,31).
It not only serves an essential role in cell proliferation,
differentiation, growth and migration, but also affects apoptosis
(32–35). According to previous studies
(36,37), PTEN dramatically enhances the
intrinsic growth/regenerative ability of corticospinal neurons to
promote the extension of injured corticospinal tract (CST) axons
following SCI with PTEN inactivation. It negatively regulates
intracellular levels of PIP3; this dephosphorylation (p-mTOR
dephosphorylated into mTOR) is important as it results in
inhibition of the Akt signaling pathway (20). The PI3K/Akt signaling pathway has a
central role in cell growth and survival through the
phosphorylation and inhibition of a number of vital substrates. For
example, Akt can activate the transcription of the proapoptotic
genes encoding Fas receptor and Bcl-2-like protein 11, and
inactivate the proapoptotic Bcl-2 family member Bcl-2-associated
agonist of cell death (20,38–40).
mTOR is a serine/threonine protein kinase that consists of two
types of protein complexes (41).
The protein kinase mTOR phosphorylates multiple downstream proteins
of the PI3K/Akt pathway (41). In
this experiment, p70S6K was selected as a biomarker of mTOR
activity (41).
The present study used pharmacological methods. The
specific inhibitors of PTEN, Akt and mTOR, bpV(pic), LY294002 and
ridaforolimus, were used to treat cells (10,42,43).
The experiments comprised four groups based on the inhibitor used
in each. Western blot analysis was used to verify the efficiency of
the specific inhibitors. Compared with the control group, the
phosphorylation levels of Akt, p70S6K and mTOR of the bpV(pic)
group were significantly increased. This suggested that bpV(pic)
had an effect on the PTEN/Akt/mTOR signaling pathway. When LY294002
was added, the phosphorylation levels of Akt, mTOR and p70S6K were
downregulated. When the ridaforolimus was added, the
phosphorylation levels of mTOR and p70S6K were downregulated, but
the level of p-Akt was unchanged. These results demonstrated that
these specific inhibitors exhibited high inhibitory efficiency, and
that the PI3K/Akt/mTOR signaling pathway was activated in cells
following PTEN inhibition.
Axonal regeneration and sprouting are two important
strategies for SCI repair (8). In
addition, they may be central to promoting reinnervation and
functional recovery after SCI. It was first explored whether
cerebral cortical neurons would show significant improvement in
overcoming CSPGs-mediated axonal inhibition following PTEN
inhibition. The maximum axon length, number of neurites and
percentage of neurites that crossed the CSPGs border were all
significantly higher in the bpV(pic) group. The neurons gained an
increased ability to extend their axons over the CSPGs substrate
and to cross into the CSPGs-rich regions. This finding is
consistent with our and other previous studies (8,23,44).
In addition, when LY294002 and ridaforolimus were added, the
ability for neuronal regeneration was attenuated. This suggested
that the PTEN inhibition promoted axonal growth/regeneration
through the PI3K/Akt/mTOR signaling pathway. Then, the
H2O2-induced apoptosis of cerebral cortical
neurons following PTEN inhibition was explored. The results of
TUNEL staining showed that the percentage of neuronal apoptosis was
significantly lower in the ridaforolimus + bpV(pic) and bpV(pic)
groups. However, it was higher in the LY294002 + bpV(pic) group,
suggesting that PI3K signaling was critically involved in apoptotic
induction. In addition, mTOR appeared to not be a critical
downstream target; based on previous findings, glycogen synthase
kinase 3β may be an important downstream target for neuronal
apoptosis (1,45).
SCI is a complicated pathophysiological process that
involves a cascade of cellular and biochemical events (2,46–48).
The present study found that the PI3K/Akt/mTOR signaling pathway
served an important role in neuronal regeneration. Whether other
signaling pathways are involved in neuronal regeneration, and
whether there is crossover activity between these signaling
pathways, requires further investigation. In any case, a single
treatment strategy, in spite of decreases in the inhibitory factors
present in the microenvironment or the promotion of
neuron-intrinsic regenerative abilities, is not sufficient to
support neuronal regeneration; thus, combining treatments with
other successful strategies targeting different mechanisms for
repair may be more successful. Further studies should be performed
to address the issues associated with SCI.
bpV(pic) is an effective PTEN inhibitor. PTEN
inhibition mediated by bpV(pic) promoted the axonal elongation and
initiation abilities of cerebral cortical neurons, as well as the
ability of these axons to cross the CSPG border; these effects were
mediated via the PTEN/PI3K/Akt/mTOR signaling pathway. Furthermore,
PTEN inhibition mediated by bpV(pic) protected neurons from
apoptosis. The results of the present study may provide a potential
novel strategy for the treatment of SCI through PTEN
inhibition.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Jianming Yang
(Key Laboratory of Immuno Microenvironment and Disease of the
Educational Ministry of China, Department of Immunology, Tianjin
Medical University, Tianjin), for her help in writing.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81501899), the State Key
Program of the National Natural Science Foundation of China (grant
no. 81330042), the Special Program for Sino-Russian Joint Research
Sponsored by the Ministry of Science and Technology, China (grant
no. 2014DFR31210), the Key Program Sponsored by the Tianjin Science
and Technology Committee, China (grant nos. 13RCGFSY19000 and
14ZCZDSY00044), the Science Foundation of Tianjin Medical
University for Young Scholars (grant no. 2014KYQ01) and the Science
Foundation of Tianjin Medical University General Hospital for Young
Scholars (grant no. ZYYFY2014037).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XK and SF conceived and designed the study. SL, JJ,
HZ and CZ performed the experiments. These four authors contributed
equally. LLiu and JL analyzed the data and drafted the Abstract and
Introduction; LLu, XL and CZ interpreted the data and wrote the
remaining parts of this manuscript; YK and YL performed the
secondary data analyses and revised the manuscript for intellectual
and scientific content, and ZC and YR contributed to the conception
of the study. All authors read and approved the manuscript.
Ethics approval and consent to
participate
All experimental procedures involving animals were
approved by the Ethics Committee of Tianjin Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jain NB, Ayers GD, Peterson EN, Harris MB,
Morse L, O'Connor KC and Garshick E: Traumatic spinal cord injury
in the United States, 1993–2012. JAMA. 313:2236–2243. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blesch A and Tuszynski MH: Spinal cord
injury: Plasticity, regeneration and the challenge of translational
drug development. Trends Neurosci. 32:41–47. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sun F and He Z: Neuronal intrinsic
barriers for axon regeneration in the adult CNS. Curr Opin
Neurobiol. 20:510–518. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang P and Yang Z: Enhancing intrinsic
growth capacity promotes adult CNS regeneration. J Neurol Sci.
312:1–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cao L, Zhu YL, Su Z, Lv B, Huang Z, Mu L
and He C: Olfactory ensheathing cells promote migration of Schwann
cells by secreted nerve growth factor. Glia. 55:897–904. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fouad K, Schnell L, Bunge MB, Schwab ME,
Liebscher T and Pearse DD: Combining Schwann cell bridges and
olfactory-ensheathing glia grafts with chondroitinase promotes
locomotor recovery after complete transection of the spinal cord. J
Neurosci. 25:1169–1178. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Harel NY and Strittmatter SM: Can
regenerating axons recapitulate developmental guidance during
recovery from spinal cord injury? Nat Rev Neurosci. 7:603–616.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Klapka N and Müller HW: Collagen matrix in
spinal cord injury. J Neurotrauma. 23:422–435. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Paveliev M, Lume M, Velthut A, Phillips M,
Arumäe U and Saarma M: Neurotrophic factors switch between two
signaling pathways that trigger axonal growth. J Cell Sci.
120:2507–2516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schwab ME: Functions of Nogo proteins and
their receptors in the nervous system. Nat Rev Neurosci.
11:799–811. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Silver J and Miller JH: Regeneration
beyond the glial scar. Nat Rev Neurosci. 5:146–156. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu K, Tedeschi A, Park KK and He Z:
Neuronal intrinsic mechanisms of axon regeneration. Annu Rev
Neurosci. 34:131–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun F, Park KK, Belin S, Wang D, Lu T,
Chen G, Zhang K, Yeung C, Feng G, Yankner BA and He Z: Sustained
axon regeneration induced by co-deletion of PTEN and SOCS3. Nature.
480:372–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Busch SA and Silver J: The role of
extracellular matrix in CNS regeneration. Curr Opin Neurobiol.
17:120–127. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yiu G and He Z: Glial inhibition of CNS
axon regeneration. Nat Rev Neurosci. 7:617–627. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park KK, Liu K, Hu Y, Kanter JL and He Z:
PTEN/mTOR and axon regeneration. Exp Neurol. 223:45–50. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shin JE, Cho Y, Beirowski B, Milbrandt J,
Cavalli V and DiAntonio A: Dual leucine zipper kinase is required
for retrograde injury signaling and axonal regeneration. Neuron.
74:1015–1022. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Smith PD, Sun F, Park KK, Cai B, Wang C,
Kuwako K, Martinez-Carrasco I, Connolly L and He Z: SOCS3 deletion
promotes optic nerve regeneration in vivo. Neuron. 64:617–623.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maehama T and Dixon JE: The tumor
suppressor, PTEN/MMAC1, dephosphorylates the lipid second
messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem.
273:13375–13378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sansal I and Sellers WR: The biology and
clinical relevance of the PTEN tumor suppressor pathway. J Clin
Oncol. 22:2954–2963. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stambolic V, Suzuki A, de la Pompa JL,
Brothers GM, Mirtsos C, Sasaki T, Ruland J, Penninger JM,
Siderovski DP and Mak TW: Negative regulation of PKB/Akt-dependent
cell survival by the tumor suppressor PTEN. Cell. 95:29–39. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park KK, Liu K, Hu Y, Smith PD, Wang C,
Cai B, Xu B, Connolly L, Kramvis I, Sahin M and He Z: Promoting
axon regeneration in the adult CNS by modulation of the PTEN/mTOR
pathway. Science. 322:963–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou H, Li X, Wu Q, Li F, Fu Z, Liu C,
Liang Z, Chu T, Wang T, Lu L, et al: shRNA against PTEN promotes
neurite outgrowth of cortical neurons and functional recovery in
spinal cord contusion rats. Regen Med. 10:411–429. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zimmermann M: Ethical principles for the
maintenance and use of animals in neuroscience research. Neurosci
Lett. 73:11987. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tom VJ, Steinmetz MP, Miller JH, Doller CM
and Silver J: Studies on the development and behavior of the
dystrophic growth cone, the hallmark of regeneration failure, in an
in vitro model of the glial scar and after spinal cord injury. J
Neurosci. 24:6531–6539. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jordan PM, Ojeda LD, Thonhoff JR, Gao J,
Boehning D, Yu Y and Wu P: Generation of spinal motor neurons from
human fetal brain-derived neural stem cells: Role of basic
fibroblast growth factor. J Neurosci Res. 87:318–332. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Geisert EE Jr and Frankfurter A: The
neuronal response to injury as visualized by immunostaining of
class III beta-tubulin in the rat. Neurosci Lett. 102:137–141.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ignatova TN, Kukekov VG, Laywell ED,
Suslov ON, Vrionis FD and Steindler DA: Human cortical glial tumors
contain neural stem-like cells expressing astroglial and neuronal
markers in vitro. Glia. 39:193–206. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kempermann G, Gast D, Kronenberg G,
Yamaguchi M and Gage FH: Early determination and long-term
persistence of adult-generated new neurons in the hippocampus of
mice. Development. 130:391–399. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li DM and Sun H: TEP1, encoded by a
candidate tumor suppressor locus, is a novel protein tyrosine
phosphatase regulated by transforming growth factor beta. Cancer
Res. 57:2124–2129. 1997.PubMed/NCBI
|
|
31
|
Steck PA, Pershouse MA, Jasser SA, Yung
WK, Lin H, Ligon AH, Langford LA, Baumgard ML, Hattier T, Davis T,
et al: Identification of a candidate tumour suppressor gene, MMAC1,
at chromosome 10q23.3 that is mutated in multiple advanced cancers.
Nat Genet. 15:356–362. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alexiou GA and Voulgaris S: The role of
the PTEN gene in malignant gliomas. Neurol Neurochir Pol. 44:80–86.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Goberdhan DC and Wilson C: PTEN: Tumour
suppressor, multifunctional growth regulator and more. Hum Mol
Genet. 12(Spec No 2): R239–R248. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Leslie NR, Maccario H, Spinelli L and
Davidson L: The significance of PTEN's protein phosphatase
activity. Adv Enzyme Regul. 49:190–196. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi Y, Paluch BE, Wang X and Jiang X: PTEN
at a glance. J Cell Sci. 125:4687–4692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu K, Lu Y, Lee JK, Samara R, Willenberg
R, Sears-Kraxberger I, Tedeschi A, Park KK, Jin D, Cai B, et al:
PTEN deletion enhances the regenerative ability of adult
corticospinal neurons. Nat Neurosci. 13:1075–1081. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zukor K, Belin S, Wang C, Keelan N, Wang X
and He Z: Short hairpin RNA against PTEN enhances regenerative
growth of corticospinal tract axons after spinal cord injury. J
Neurosci. 33:15350–15361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo
P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Datta SR, Dudek H, Tao X, Masters S, Fu H,
Gotoh Y and Greenberg ME: Akt phosphorylation of BAD couples
survival signals to the cell-intrinsic death machinery. Cell.
91:231–241. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dijkers PF, Medema RH, Pals C, Banerji L,
Thomas NS, Lam EW, Burgering BM, Raaijmakers JA, Lammers JW,
Koenderman L and Coffer PJ: Forkhead transcription factor FKHR-L1
modulates cytokine-dependent transcriptional regulation of
p27(KIP1). Mol Cell Biol. 20:9138–9148. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Park KK, Hu Y, Muhling J, Pollett MA,
Dallimore EJ, Turnley AM, Cui Q and Harvey AR: Cytokine-induced
SOCS expression is inhibited by cAMP analogue: Impact on
regeneration in injured retina. Mol Cell Neurosci. 41:313–324.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schmid AC, Byrne RD, Vilar R and
Woscholski R: Bisperoxovanadium compounds are potent PTEN
inhibitors. FEBS Lett. 566:35–38. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vlahos CJ, Matter WF, Hui KY and Brown RF:
A specific inhibitor of phosphatidylinositol 3-kinase,
2-(4-morpholinyl)- 8-phenyl-4H-1-benzopyran-4-one (LY294002). J
Biol Chem. 269:5241–5248. 1994.PubMed/NCBI
|
|
44
|
Lu Y, Belin S and He Z: Signaling
regulations of neuronal regenerative ability. Curr Opin Neurobiol.
27:135–142. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dill J, Wang H, Zhou F and Li S:
Inactivation of glycogen synthase kinase 3 promotes axonal growth
and recovery in the CNS. J Neurosci. 28:8914–8928. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bartus K, James ND, Bosch KD and Bradbury
EJ: Chondroitin sulphate proteoglycans: Key modulators of spinal
cord and brain plasticity. Exp Neurol. 235:5–17. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gervasi NM, Kwok JC and Fawcett JW: Role
of extracellular factors in axon regeneration in the CNS:
Implications for therapy. Regen Med. 3:907–923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jiang H, Guo W, Liang X and Rao Y: Both
the establishment and the maintenance of neuronal polarity require
active mechanisms: Critical roles of GSK-3beta and its upstream
regulators. Cell. 120:123–135. 2005. View Article : Google Scholar : PubMed/NCBI
|