Introduction
Colorectal cancer (CRC) is among the most common
malignancies worldwide, with a high incidence in the United States
and Western Europe (1). In 2017,
>130,000 newly cases were diagnosed and >50,000 individuals
succumbed to mortality in the United States (1). The incidence rates and mortality
rates have declined in patients with CRC aged >50 years,
however, the incidence rates have increased by 22% in patients aged
<50 years and the mortality rates have increased by 13% in the
last decade (1). In China, the
incidence and mortality rates of CRC have markedly increased
(2). Despite substantial efforts
in the establishment of early detection systems and chemotherapy
reagents, the prognosis of CRC remains far from satisfactory for
the majority of patients (3).
The therapeutic responses and survival outcomes of
CRC are constrained by the clinical heterogeneity (4,5).
Therefore, molecular markers emerge as efficient classifiers for
CRC (6,7). This is exemplified by the mutations
identified in genes such as
KRAS/BRAF/phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic
subunit α (PIK3CA), which have been widely accepted as clinical
indicators for therapeutic decisions (6). However, the clinical outcomes of CRC
remain largely diverse.
Microsatellite instability (MSI) is one of the most
intensively investigated molecular markers in CRC (3). MSI indicates inactivation of the
mismatch repair (MMR) genes and is frequently associated with the
CpG island methylator phenotype, whereas microsatellite stable
(MSS) is associated with chromosomal instability (CIN) (3). MSI CRCs are divided into MSI-High
(MSI-H) or MSI-Low (MSI-L) subsets, based on the extent of the
instability (8). The essence of
MSI has been intensively evaluated in the National Comprehensive
Cancer Network guideline (8).
Noteworthy, 15% of patients with CRC show MSI whereas the remainder
are characterized by MSS (9,10).
CRCs with MSI-H often exhibit numerous distinct features, including
a more proximal tumor position (11). Furthermore, >80% of patients
with CRC with Lynch syndrome, a top-ranked inherited CRC-associated
disease, exhibit MSI (12,13). Of note, ~10-20% of patients with
CRC with Lynch syndrome manifest MSS, with diverse
immunohistochemistry results (14).
MSI is one of the most promising markers
investigated to date with prognostic and therapeutic values.
Previously, patients with MSI were associated with a favorable
prognosis compared with those with MSS (3,12,15).
However, the prognostic role of MSS and MSI in CRC remains
controversial. When patients with MSI-H and MSS/MSI-L received
fluorouracil (FU), the significantly different prognostic values
between the two became indistinguishable (16). In a MSI subset, those treated with
FU had a poorer prognosis than those without FU (16). A recent study highlighted the
predictive role of MMR status in immune checkpoint inhibition with
pembrolizumab (7).
However, the mechanism underlying the association
between MSI status and the clinicopathological characteristics of
CRC remains to be fully elucidated. To gain better insight into the
key genes and pathways involved in MSI of CRC, bioinformatics
analysis of the GSE25071 gene expression profile, including 38 MSS,
five MSI-H and three MSI-L samples, was conducted to identify
potential key genes and pathways associated with MSI.
Materials and methods
Gene expression profile from the Gene
Expression Omnibus (GEO) database
The gene expression profile, GSE25071, which
contained 38 MSS colorectal cancer cases, five MSI-H cases and
three MSI-L cases (17), was
downloaded from the GEO database (http://www.ncbi.nlm.nih.gov/geo/) (18). The GSE25071 profile was further
annotated by the profile GPL2986, ABI Human Genome Survey
Microarray Version 2 (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) (17).
Briefly, familial adenomatous polyposis (FAP) syndromes, hereditary
nonpolyposis colorectal cancer (HNPCC) and other types of cancer
were excluded from the included cases. Clinical information
including gender, age, tumor localization, tumor stage according to
The International Union Against Cancer (UICC)/American Joint
Committee on Cancer and MSI status were recorded (17). The total RNA of the samples was
processed using the AllPrep DNA/RNA Mini kit (Qiagen, Inc.,
Valencia, CA, USA) for tumors and the Ambion RiboPure kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) for normal colonic
mucosa. The Chemilunimescent Detection kit from Applied Biosystems;
Thermo Fisher Scientific, Inc. was used following the labeling
process (NanoAmp RT-IVT Labeling kit; Applied Biosystems; Thermo
Fisher Scientific, Inc., Waltham, MA, USA; DIG-UTP, Roche
Diagnostics, Basel, Switzerland). Subsequently, the microarrays
were scanned using the AB1700 Chemilunimescent microarray analyzer
and further processed by the accompanying software (Applied
Biosystems; Thermo Fisher Scientific, Inc.; version 1.1.1)
(17). GSE18088, GSE13067 and
GSE78220 were included for external validation of the
differentially expressed genes (DEGs) determined in GSE25071.
GSE18088 contained 34 cases of MSS and 19 cases of MSI with primary
stage II colon cancer based on the UICC. The profile was based on
the Affymetrix Human Genome U133 Plus 2.0 Array (Affymetrix; Thermo
Fisher Scientific, Inc.) (19).
GSE13067 contained 63 MSS and 11 MSI-H fresh-frozen primary CRC
samples for the Affymetrix Human Genome U133 Plus 2.0 Array
(Affymetrix; Thermo Fisher Scientific, Inc.) (20). GSE78220 contained 28 melanoma
samples for transcriptomic analysis of anti-PD-1 therapy
(pembrolizumab), with the Illumina HiSeq 2000 platform (GPL11154;
Illumina Inc., San Diego, CA, USA) (21).
Identification of the DEGs
The DEGs listed in three groups of MSS vs. MSI, MSS
vs. MSI-L and MSS vs. MSI-H were identified based on the GEO2R
web-based tool (www.ncbi.nlm.nih.gov/geo/) (22). The Benjamini-Hochberg procedure
(false discovery rate) was applied. The predefined cut-off values
included adj. P<0.05 and |log fold change (logFC)|>2. Given
the limited DEGs in MSS vs. MSI-L and MSS vs. MSI (H+L), only the
expression data of DEGs identified in MSS vs. MSI-H group were
processed in FunRich software (version 2.1.2; www.funrich.org) for a bidirectional hierarchical
clustering plot (23).
Gene ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway analyses of DEGs
in the MSS vs
MSI-H group
For further functional enrichment analysis of the
DEGs in MSS vs. MSI-H, GO enrichment analysis, including biological
process (BP), molecular function (MF) and cellular component (CC),
and KEGG analysis were performed using the Database for Annotation,
Visualization, and Integrated Discovery (http://david.abcc.ncifcrf.gov/) web-based platform
(24–26).
Gene co-expression network
analysis
Only the DEGs of MSS vs. MSI-H were included for the
subsequent analytic processes. All DEGs were imported into
GeneMANIA, a web-based gene-gene interactions identification
database (www.genemania.org) (27). The interaction list of the DEGs and
additional genes (node degree ≥2) was output for visualization in
the Cytoscape program (version 3.6.0; www.cytoscape.org/) (28). The hub genes with highest connected
edges within the co-expression networks were determined. The
highest ranked three modules were identified using the Molecular
Complex Detection (MCODE) program (29). The mRNA expression of the hub genes
were further externally validated in GSE18088, GSE13067 and
GSE78220.
Protein-protein interaction (PPI)
networks
All DEGs were further input to the Search Tool for
the Retrieval of Interacting Genes/Proteins database (STRING
version 10.0; www.string-db.org/) for interaction network at the
protein level (30). The results
were further visualized using Cytoscape with predefined cut-off
values: Node degree, ≥2; k-score(value=2); node score, 0.2; max.
depth, 100 (28).
Gene set enrichment analysis
(GSEA)
GSEA, released by the Broad Institute (software.broadinstitute.org/gsea/index.jsp), was
used to cluster significant gene sets associated with given
annotation terms (31). In the
present study, the MSS and MSI-H samples were analyzed by GSEA with
the annotation of ‘hallmark gene sets’. The significant cut-off
value was defined as P<0.05.
Weighted gene correlation network
analysis (WGCNA)
WGCNA was used for co-expression network
construction based on the correlations among genes and
identification of top-ranked gene modules and hub genes. The
‘WGCNA’ R package was implemented for all the genes in 46 samples
(38 MSS, five MSI-H and three MSI-L, normal samples were excluded).
Initially, Pearson's correlation of each pair genes was calculated
and an adjacency matrix was constructed based on the Pearson's
results and a predefined soft-thresholding parameter (β).
Subsequently, the topological overlap matrix of the included genes
and adjacency matrix was produced. Genes with similar expression
trends were classified as the same modules eigengenes for further
clinical traits correlation (32,33).
The genes of the most correlated module were extracted for GO and
KEGG analyses and PPI network construction. Hub genes were defined
with the highest degrees.
Prognostic values of hub genes in
WGCNA
The overall survival of hub genes determined by
WGCNA was further examined in the PrognoScan database, a
comprehensive platform for prognostic annotation
(dna00.bio.kyutech.ac.jp/PrognoScan/) (34).
Results
Identification of upregulated and
downregulated DEGs
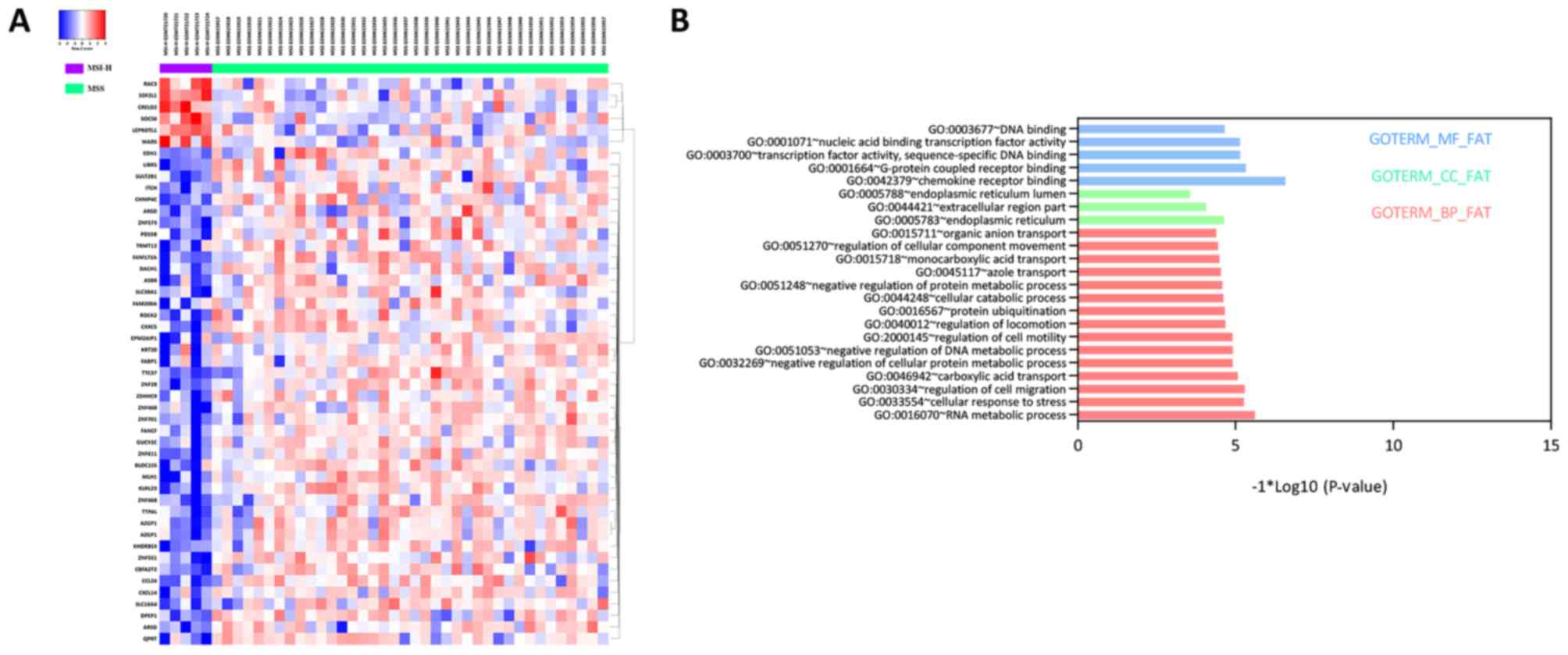
A total of 49 DEGs, including six upregulated and 43
downregulated DEGs, were identified between MSI-H and MSS groups,
illustrated by the bidirectional clustering heat map (Fig. 1A). However, no significant DEGs
were identified between MSI-L and MSS, and only one DEG was
identified between MSI-H and MSI-L. Therefore, subsequent
investigations focused on the DEGs between MSI-H and MSS.
GO enrichment and KEGG pathway
analyses of DEGs
A total of 15 BP, three CC and five MF terms were
significantly enriched. Specifically, RNA metabolic process,
endoplasmic reticulum and chemokine receptor binding were the
top-ranked in each term, respectively (Fig. 1B). No significant KEGG pathway was
enriched in the DEGs.
Co-expressed genes network
analysis
To delineate the biological functions of the DEGs, a
co-expression network of the DEGs with correlated additional genes
was established based on the GeneMANIA program. A total of 66 nodes
and 678 edges were determined (Fig.
2A). The top five hub genes with the highest degree were
identified, including zinc finger protein (ZNF) 813, ZNF426,
ZNF611, ZNF320 and ZNF573.
The top three modules with the highest scores were
identified using the MCODE plugin (Fig. 2B-D). Among them, no particular KEGG
pathway was significantly enriched. However, in the GO enrichment,
the regulation of transcription, DNA-templated term was the highest
ranked BP for module 1, and negative regulation of hydrolase
activity for module 2. Subsequently, the hub genes were externally
validated in GSE18088, GSE13067 and GSE78220. The expression levels
of ZNF426, ZNF320 and ZNF573 were significantly reduced in MSI-H
compared with MSS in GSE18088. The expression levels of ZNF813,
ZNF426 and ZNF573 were significantly reduced in the MSI-H group
compared with the MSS group in GSE13067. The expression levels of
ZNF813 and ZNF573 were significantly reduced in the pembrolizumab
no-response group compared with the response group (Fig. 3A-C).
PPI network analysis
The minimum required interaction score of STRING was
medium confidence (0.4) and the cut-off degree for the included
nodes in Cytoscape was ≥1. The PPI networks included 14 nodes and
eight edges (Fig. 4), being
distinct from the co-expression networks (Fig. 2A).
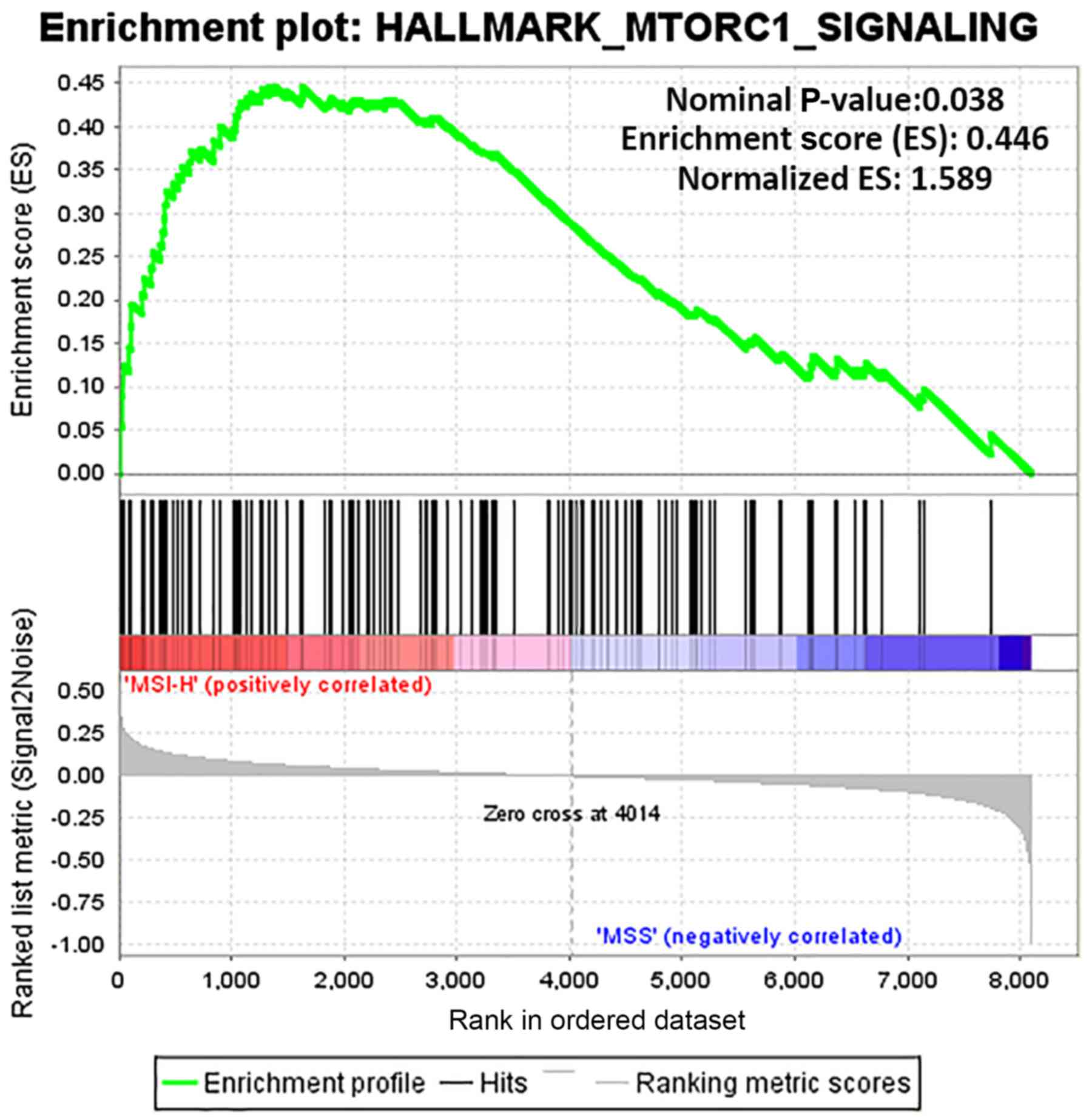
GSEA results
GSEA was used to determine the functions of gene
sets between the MSI-H and MSS groups. Only one gene set, the
mammalian target of rapamycin complex 1 (mTORC1) signaling (nominal
P-value, 0.038; normalized enrichment score, 0.446) was
significantly enriched in MSI-H, with none significantly enriched
in MSS (Fig. 5). Furthermore, the
50 top ranked genes correlated with each phenotype (MSI-H and MSS)
are illustrated with a heat map (Fig.
6).
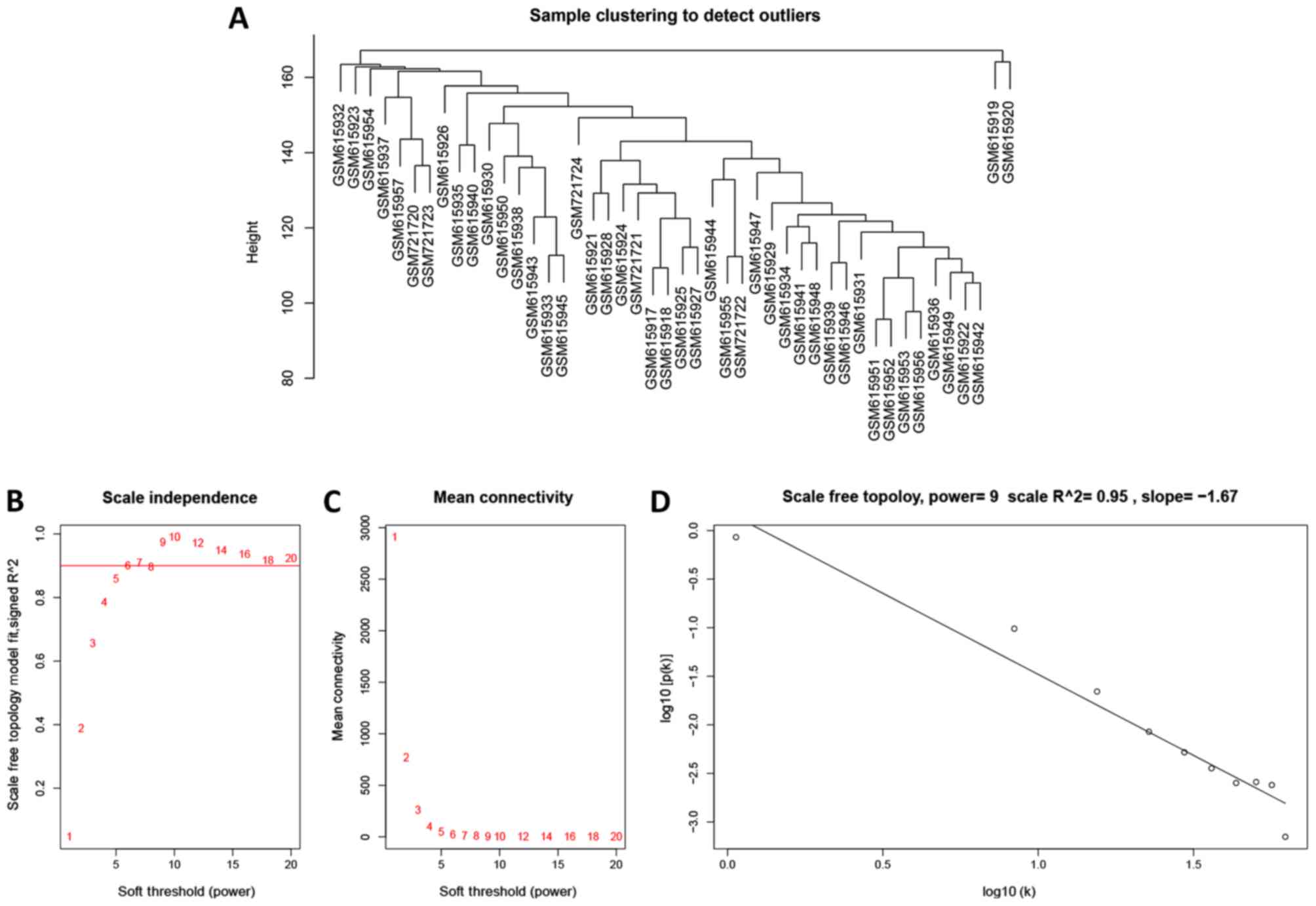
WGCNA of the gene expression profile
in all tumor samples
To further investigate the potential gene modules
associated with MSI status, the WGCNA was conducted with R package.
A total of 46 cases, including 38 MSS, eight MSI (five MSI-H and
three MSI-L) were clustered (Fig.
7A). The power of β=9 was defined as the soft-thresholding
value (scale free R2=0.95; slope=−1.67; Fig. 7B-D). A total of 25 modules were
identified (Fig. 8A). Noteworthy,
the pink module was the most significantly correlated with MSI
status (R2=0.5, P=0.0004). Of note, the pink module also
featured a high correlation (R2=0.53, P=0.0002) with
tumor stage (Fig. 8B).
Identification of hub genes in the
pink module associated with MSI status
The genes in the pink module were significantly
enriched by protein targeting to endoplasmic reticulum in BP terms,
cytosolic part in CC terms, structural constituent of ribosome in
MF terms and ribosome in KEGG pathways. Subsequently, a PPI network
(degree ≥1), with 55 nodes and 96 edges, was constructed based on
the genes in the pink module. The top 10 hub genes with the highest
degrees were determined, including ribosomal protein L12 (RPL12),
ribosomal protein S3A (RPS3A), ribosomal protein S9 (RPS9),
ribosomal protein L27a (RPL27A), ribosomal protein L7 (RPL7),
ribosomal protein L28 (RPL28), ribosomal protein L14 (RPL14),
ribosomal protein S17 (RPS17), mitochondrial ribosomal protein L16
(MRPL16) and G elongation factor, mitochondrial 2 (GFM2), as shown
in Fig. 9.
Prognostic values of the hub genes in
the pink module
The prognostic values of the hub genes were assessed
in various independent datasets with different probe IDs and array
types. In all, none of the genes exhibited significant prognostic
values (Table I).
 | Table I.Prognostic values of hub genes from
PrognoScan. |
Table I.
Prognostic values of hub genes from
PrognoScan.
| ID_NAME | Dataset | Array type | Probe ID | N | COX P-value | HR (95% CI lower-CI
upper) |
|---|
| GFM2 | GSE17536 | HG-U133_Plus_2 | 231917_at | 177 | 0.52 | 1.20
(0.69–2.10) |
| GFM2 | GSE17536 | HG-U133_Plus_2 | 231918_s_at | 177 | 0.43 | 1.19
(0.78–1.81) |
| GFM2 | GSE17536 | HG-U133_Plus_2 | 225392_at | 177 | 0.35 | 1.33
(0.73–2.42) |
| GFM2 | GSE17537 | HG-U133_Plus_2 | 231917_at | 55 | 0.64 | 0.75
(0.22–2.54) |
| GFM2 | GSE17537 | HG-U133_Plus_2 | 231918_s_at | 55 | 0.54 | 1.24
(0.63–2.44) |
| GFM2 | GSE17537 | HG-U133_Plus_2 | 225392_at | 55 | 0.47 | 1.37
(0.59–3.21) |
| MRPL16 | GSE12945 | HG-U133A | 217980_s_at | 62 | 0.82 | 1.12
(0.41–3.08) |
| MRPL16 | GSE17536 | HG-U133_Plus_2 | 217980_s_at | 177 | 0.86 | 1.06
(0.59–1.89) |
| MRPL16 | GSE17537 | HG-U133_Plus_2 | 217980_s_at | 55 | 0.623951 | 1.23
(0.53–2.86) |
| RPL12 | GSE12945 | HG-U133A | 214271_x_at | 62 | 0.76 | 2.21
(0.01–326.11) |
| RPL12 | GSE12945 | HG-U133A | 200809_x_at | 62 | 0.39 | 107.59
(0.00–4600568.31) |
| RPL12 | GSE17536 | HG-U133_Plus_2 | 214271_x_at | 177 | 0.29 | 2.03
(0.55–7.52) |
| RPL12 | GSE17536 | HG-U133_Plus_2 | 200809_x_at | 177 | 0.34 | 1.91
(0.50–7.28) |
| RPL12 | GSE17537 | HG-U133_Plus_2 | 214271_x_at | 55 | 0.47 | 1.76
(0.38–8.13) |
| RPL12 | GSE17537 | HG-U133_Plus_2 | 200809_x_at | 55 | 0.23 | 2.31
(0.59–8.99) |
| RPL14 | GSE12945 | HG-U133A | 213588_x_at | 62 | 0.45 | 7654467.33 |
|
|
|
|
|
|
|
(0.00–6592665664338867322355712.00) |
| RPL14 | GSE12945 | HG-U133A | 219138_at | 62 | 0.76 | 0.78
(0.17–3.69) |
| RPL14 | GSE12945 | HG-U133A | 200074_s_at | 62 | 0.90 | 1.07
(0.40–2.83) |
| RPL14 | GSE17536 | HG-U133_Plus_2 | 213588_x_at | 177 | 0.78 | 0.84
(0.24–2.93) |
| RPL14 | GSE17536 | HG-U133_Plus_2 | 200074_s_at | 177 | 0.57 | 0.75
(0.28–2.00) |
| RPL14 | GSE17536 | HG-U133_Plus_2 | 219138_at | 177 | 0.52 | 0.72
(0.26–1.97) |
| RPL14 | GSE17537 | HG-U133_Plus_2 | 200074_s_at | 55 | 0.95 | 0.96
(0.28–3.26) |
| RPL14 | GSE17537 | HG-U133_Plus_2 | 219138_at | 55 | 0.89 | 0.95
(0.46–1.97) |
| RPL14 | GSE17537 | HG-U133_Plus_2 | 213588_x_at | 55 | 0.86 | 0.88
(0.23–3.43) |
| RPL27A | GSE12945 | HG-U133A | 212044_s_at | 62 | 0.88 | 1.06
(0.48–2.33) |
| RPL27A | GSE12945 | HG-U133A | 203034_s_at | 62 | 0.72 | 0.80
(0.23–2.71) |
| RPL27A | GSE17536 | HG-U133_Plus_2 | 223707_at | 177 | 0.43 | 0.77
(0.40–1.49) |
| RPL27A | GSE17536 | HG-U133_Plus_2 | 212044_s_at | 177 | 0.69 | 0.87
(0.42–1.78) |
| RPL27A | GSE17536 | HG-U133_Plus_2 | 203034_s_at | 177 | 0.48 | 2.43
(0.21–27.88) |
| RPL27A | GSE17537 | HG-U133_Plus_2 | 212044_s_at | 55 | 0.23 | 0.75
(0.47–1.20) |
| RPL27A | GSE17537 | HG-U133_Plus_2 | 203034_s_at | 55 | 0.83 | 1.46
(0.05–41.30) |
| RPL27A | GSE17537 | HG-U133_Plus_2 | 223707_at | 55 | 0.28 | 0.50
(0.14–1.77) |
| RPL28 | GSE12945 | HG-U133A | 213223_at | 62 | 0.24 | 2.03
(0.62–6.63) |
| RPL28 | GSE12945 | HG-U133A | 200003_s_at | 62 | 0.58 | 1.34
(0.48–3.73) |
| RPL28 | GSE17536 | HG-U133_Plus_2 | 213223_at | 177 | 0.95 | 0.98
(0.53–1.81) |
| RPL28 | GSE17536 | HG-U133_Plus_2 | 200003_s_at | 177 | 0.81 | 0.83
(0.18–3.74) |
| RPL28 | GSE17537 | HG-U133_Plus_2 | 200003_s_at | 55 | 0.19 | 2.72
(0.61–12.16) |
| RPL28 | GSE17537 | HG-U133_Plus_2 | 213223_at | 55 | 0.99 | 1.01
(0.26–3.88) |
| RPL7 | GSE12945 | HG-U133A | 212042_x_at | 62 | 0.59 | 1.98
(0.17–23.44) |
| RPL7 | GSE12945 | HG-U133A | 200717_x_at | 62 | 0.42 | 3650.83 |
|
|
|
|
|
|
|
(0.00–1740006561282.69) |
| RPL7 | GSE17536 | HG-U133_Plus_2 | 239493_at | 177 | 0.071 | 1.69
(0.96–3.00) |
| RPL7 | GSE17536 | HG-U133_Plus_2 | 212042_x_at | 177 | 0.16 | 3.10
(0.63–15.23) |
| RPL7 | GSE17536 | HG-U133_Plus_2 | 200717_x_at | 177 | 0.14 | 4.24
(0.63–28.50) |
| RPL7 | GSE17537 | HG-U133_Plus_2 | 239493_at | 55 | 0.0050 | 0.09
(0.02–0.48) |
| RPL7 | GSE17537 | HG-U133_Plus_2 | 212042_x_at | 55 | 0.45 | 0.38
(0.03–4.76) |
| RPL7 | GSE17537 | HG-U133_Plus_2 | 200717_x_at | 55 | 0.53 | 0.47
(0.04–4.97) |
| RPS17 | GSE12945 | HG-U133A | 201665_x_at | 62 | 0.86 | 0.82
(0.08–8.15) |
| RPS17 | GSE12945 | HG-U133A | 212578_x_at | 62 | 0.66 | 1.91
(0.11–33.65) |
| RPS17 | GSE17536 | HG-U133_Plus_2 | 212578_x_at | 177 | 0.082 | 4.17
(0.84–20.77) |
| RPS17 | GSE17536 | HG-U133_Plus_2 | 201665_x_at | 177 | 0.17 | 2.96
(0.64–13.76) |
| RPS17 | GSE17537 | HG-U133_Plus_2 | 212578_x_at | 55 | 0.53 | 2.16
(0.20–23.51) |
| RPS17 | GSE17537 | HG-U133_Plus_2 | 201665_x_at | 55 | 0.37 | 2.99
(0.27–32.95) |
| RPS3A | GSE12945 | HG-U133A | 201257_x_at | 62 | 0.80 | 25.91 |
|
|
|
|
|
|
|
(0.00–2227310678806.87) |
| RPS3A | GSE17536 | HG-U133_Plus_2 | 201257_x_at | 177 | 0.97 | 1.04
(0.11–9.64) |
| RPS3A | GSE17537 | HG-U133_Plus_2 | 201257_x_at | 55 | 0.65 | 0.48
(0.02–11.00) |
| RPS9 | GSE12945 | HG-U133A | 217747_s_at | 62 | 0.85 | 0.90
(0.31–2.63) |
| RPS9 | GSE12945 | HG-U133A | 214317_x_at | 62 | 0.77 | 5.03
(0.00–240092.23) |
| RPS9 | GSE17536 | HG-U133_Plus_2 | 1557981_at | 177 | 0.0018 | 0.06
(0.01–0.35) |
| RPS9 | GSE17536 | HG-U133_Plus_2 | 214317_x_at | 177 | 0.81 | 1.22
(0.24–6.07) |
| RPS9 | GSE17536 | HG-U133_Plus_2 | 217747_s_at | 177 | 0.66 | 1.30
(0.40–4.17) |
| RPS9 | GSE17537 | HG-U133_Plus_2 | 217747_s_at | 55 | 0.99 | 1.01
(0.36–2.84) |
| RPS9 | GSE17537 | HG-U133_Plus_2 | 1557981_at | 55 | 0.25 | 0.08
(0.00–5.87) |
| RPS9 | GSE17537 | HG-U133_Plus_2 | 214317_x_at | 55 | 0.70 | 1.50
(0.19–11.58) |
Discussion
The present study is the first, to the best of our
knowledge, to use multiple bioinformatics analysis approaches to
demonstrate the potential key genes and pathways associated with
MSI status in patients with CRC. Among the significant GO terms,
RNA metabolic process, endoplasmic reticulum and chemokine receptor
binding were the top-ranked terms. No significant KEGG pathway was
identified, however, using GSEA, the MTORC1 signaling pathway was
significantly enriched. MTOR signaling pathways, consisting of at
least two complexes, mTORC1 and mTORC2, receive a plethora of input
factors and modulate a broad spectrum of downstream molecules
(35,36). Noteworthy, the inhibition of mTOR1
only leads to mild protein synthesis reduction and potential
influence upon the cell cycle process (35,37).
Previously, Choi et al investigated somatic
mutational, intratumoral heterogeneity and expressional alterations
of mTOR pathway-related genes in cancer with MSI, including PIK3CB,
insulin receptor substrate 1/2 (IRS1), RPS6, eukaryotic translation
initiation factor 4B (EIF4B), RPS6KA5 and PRKAA2 (38). Of the patients with MSI-H CRC, 8.9%
harbored IRS1 frameshift mutations, whereas 10.1% harbored
mutations in EIF4B and 3.8% in RPS6KA5. Noteworthy, no mutations
was identified in MSS or MSI-L (38). The study by Choi et al and
the present study demonstrated the potential roles of the ribosomal
protein family associated with MSI status. In addition, differing
from previous search strategies in published results (38), the present study illustrated how
WGCNA can be implemented to predict new genes in the regulation of
MSI in CRC.
Lin et al analyzed the mutations of 113 MSS
and 29 MSI-H cases of CRC (39).
Mutations of PIK3CA, phosphatase and tensin homolog and/or AKT1 in
the mTOR pathway were found in 59% of the MSI-H patients compared
with 19% of the patients with MSS (39). Methodologically, a 50-gene AmpliSeq
Cancer Hotspot Panel was introduced by Lin et al, whereas
the expression profile generated using the ABI Human Genome Survey
Microarray was analyzed in the present study for GSEA and WGCNA.
Collectively, the present study provided an insightful target and
further complemented the results reported by Choi et al and
Lin et al with regards to the multiple bioinformatics
strategies.
In the co-expressed gene network, ZNF813, ZNF426,
ZNF611, ZNF320 and ZNF573 were the top-ranked hub genes closely
associated with MSI-H. Reduced expression levels of ZNF813 and
ZNF573 were found in the MSI-H group and pembrolizumab no-response
group. Of note, the check-point inhibitor pembrolizumab
significantly prolonged the prognostic outcomes of patients with
MSI-H CRC compared with those in the MSS group (7). This indicated that potential
mechanisms exist between MSI status and the outcomes of check-point
inhibitor treatment, further highlighting the predictive role of
ZNFs.
ZNFs are one of the most common proteins in the
eukaryotic system, with a broad range of biological functions,
including DNA recognition, RNA transcription, apoptosis and protein
structure (40). The five hub
genes are mainly located in the nucleus and are involved in
DNA-binding and transcription regulation. Novel topologies of
numerous ZNF domains have provided evidence for structure/function
relationships (40). To the best
of our knowledge, the present study is the first highlighting the
potential association between ZNFs and MSI in CRC.
The genes in the pink module of the WGCNA
demonstrated the highest correlation with MSI. By performing
further PPI network analysis, the top 10 hub genes were identified,
including RPL12, RPS3A, RPS9, RPL27A, RPL7, RPL28, RPL14, RPS17,
MRPL16 and GFM2. Given the high proportion of RPs in the hub gene
list, RPs are of interest for further discussion and may be of
significance to the mechanism underlying the MSI in CRC. The
synthesis of RPs is the basis for the biological processes in each
cell (41). The newly decoded
crystal structures of ribosomes provide multiple traits associated
with PPIs, RNA-protein and protein-drugs interactions (41). Noteworthy, the mutation of RPS20,
part of the small ribosome subunit, can render individuals with MSS
predisposition (42), highlighting
the association between ribosome and MSI/MSS status. In addition,
RPs are associated with biological RNA synthesis, one of the
predominant features of cancer cells exposed to 5-FU treatment
(43,44). Therefore, this clarified the role
of MSI in FU-non-responders, at least in part. Of note, the
prognostic evaluation of the hub genes indicated that their
potential roles may not be directly associated with survival
status.
Previously, Timmermann et al fully
investigated the whole exome next generation sequencing of 454
patients with CRC and identify the significant 359 mutations in MSI
and 45 mutations in MSS (45). In
addition to the MSI and CIN subtypes, a third subtype associated
with sessile-serrated adenomas was proposed (46). This newly added third subtype may
partially clarify the limited DEGs identified in MSI-L vs. MSS and
MSI vs. MSS in the present study.
The limitations of the present study include the
comparably small sample size in MSI-H and lack of experimental
validation for hub genes. Larger CRC samples with MSI/MSS and
molecular biological experiments are required to specifically
confirm the functions and mechanisms of the hub genes underlying
MSI in CRC. However, to reduce the potential confounding factors
produced by a single bioinformatics approach, the present study
employed multiple bioinformatics patterns, including DEG analysis,
GSEA and WGCNA. In addition, due to the limited number of patients
and lack of survival data in original files, the prognostic values
of hub genes identified by WGCNA were examined using the PrognoScan
database.
In conclusion, the bioinformatics analysis performed
in the present study identified key genes and pathways associated
with MSI, and further elucidated insightful traits for potential
mechanisms.
Acknowledgements
The authors would like to thank the Shanghai
Institute of Digestive Surgery, Ruijin Hospital, Shanghai Jiao Tong
University School of Medicine (Shanghai, China) for their academic
support. The authors would also like to thank Dr Ernest Johann
Helwig (Tongji Medical College, Huazhong University of Science and
Technology, Wuhan, China) for his helpful contributions with
discussions and language editing.
Funding
This study was financially supported by the National
Natural Science Foundation of China (grant nos. 81402423 and
81572818) and the Shanghai Municipal Commission of Health and
Family Planning (grant no. 2017YQ062).
Availability of data and materials
The datasets supporting the conclusion of this
article are included within the article.
Authors' contributions
CY, HH, SZ and JS performed experiments and data
analysis; CY, HH, YZ, JM and MZ drafted the manuscript; CY, AL, JS,
MZ, YZ and JM participated in the study design, data collection and
revision process. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CI
|
confidence intervals
|
|
GEO
|
Gene Expression Omnibus
|
|
HR
|
hazard ratio
|
|
CRC
|
colorectal cancer
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
PPI
|
protein-protein interaction
|
|
STRING
|
Search Tool for the Retrieval of
Interacting Genes
|
|
MCODE
|
Molecular Complex Detection
|
|
BP
|
biological processes
|
|
CC
|
cellular components
|
|
MF
|
molecular functions
|
|
WGCNA
|
weighted gene correlation network
analysis
|
|
MSS
|
microsatellite stable
|
|
MSI-H
|
microsatellite instability high
|
|
MSI-L
|
microsatellite instability low
|
|
MMR
|
mismatch repair
|
|
FU
|
fluorouracil
|
|
HNPCC
|
hereditary nonpolyposis colorectal
cancer
|
|
UICC
|
International Union Against Cancer
|
|
ZNF
|
zinc finger protein
|
|
RP
|
ribosomal protein
|
|
CIN
|
chromosomal instability
|
References
|
1
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RGS, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Popat S, Hubner R and Houlston RS:
Systematic review of microsatellite instability and colorectal
cancer prognosis. J Clin Oncol. 23:609–618. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Markowitz SD and Bertagnolli MM: Molecular
origins of cancer: Molecular basis of colorectal cancer. N Engl J
Med. 361:2449–2460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Budinska E, Popovici V, Tejpar S, D'Ario
G, Lapique N, Sikora KO, Di Narzo AF, Yan P, Hodgson JG, Weinrich
S, et al: Gene expression patterns unveil a new level of molecular
heterogeneity in colorectal cancer. J Pathol. 231:63–76. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Roock W, De Vriendt V, Normanno N,
Ciardiello F and Tejpar S: KRAS, BRAF, PIK3CA, and PTEN mutations:
Implications for targeted therapies in metastatic colorectal
cancer. Lancet Oncol. 12:594–603. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Diaz LA Jr and Le DT: PD-1 blockade in
tumors with mismatch-repair deficiency. N Engl J Med. 373:19792015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Benson AB Jr, Venook AP, Cederquist L,
Chan E, Chen YJ, Cooper HS, Deming D, Engstrom PF, Enzinger PC,
Fichera A, et al: Colon cancer, version 1.2017, NCCN clinical
practice guidelines in oncology. J Natl Compr Canc Netw.
15:370–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peltomaki P: Role of DNA mismatch repair
defects in the pathogenesis of human cancer. J Clin Oncol.
21:1174–1179. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cottrell S, Bodmer WF, Bicknell D and
Kaklamanis L: Molecular analysis of APC mutations in familial
adenomatous polyposis and sporadic colon carcinomas. Lancet.
340:626–630. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cancer Genome Atlas Network: Comprehensive
molecular characterization of human colon and rectal cancer.
Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
de la Chapelle A and Hampel H: Clinical
relevance of microsatellite instability in colorectal cancer. J
Clin Oncol. 28:3380–3087. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aaltonen LA, Salovaara R, Kristo P,
Canzian F, Hemminki A, Peltomäki P, Chadwick RB, Kääriäinen H,
Eskelinen M, Järvinen H, et al: Incidence of hereditary
nonpolyposis colorectal cancer and the feasibility of molecular
screening for the disease. N Engl J Med. 338:1481–1487. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pinol V, Castells A, Andreu M,
Castellví-Bel S, Alenda C, Llor X, Xicola RM, Rodríguez-Moranta F,
Payá A, Jover R, et al: Accuracy of revised Bethesda guidelines,
microsatellite instability, and immunohistochemistry for the
identification of patients with hereditary nonpolyposis colorectal
cancer. JAMA. 293:1986–1994. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Walther A, Houlston R and Tomlinson I:
Association between chromosomal instability and prognosis in
colorectal cancer: A meta-analysis. Gut. 57:941–950. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ribic CM, Sargent DJ, Moore MJ, Thibodeau
SN, French AJ, Goldberg RM, Hamilton SR, Laurent-Puig P, Gryfe R,
Shepherd LE, et al: Tumor microsatellite-instability status as a
predictor of benefit from fluorouracil-based adjuvant chemotherapy
for colon cancer. N Engl J Med. 349:247–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ågesen TH, Berg M, Clancy T, Thiis-Evensen
E, Cekaite L, Lind GE, Nesland JM, Bakka A, Mala T, Hauss HJ, et
al: CLC and IFNAR1 are differentially expressed and a global
immunity score is distinct between early-and late-onset colorectal
cancer. Genes Immun. 12:653–662. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Edgar R, Domrachev M and Lash AE: Gene
expression omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gröne J, Lenze D, Jurinovic V, Hummel M,
Seidel H, Leder G, Beckmann G, Sommer A, Grützmann R, Pilarsky C,
et al: Molecular profiles and clinical outcome of stage UICC II
colon cancer patients. Int J Colorectal Dis. 26:847–858. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jorissen RN, Lipton L, Gibbs P, Chapman M,
Desai J, Jones IT, Yeatman TJ, East P, Tomlinson IP, Verspaget HW,
et al: DNA copy-number alterations underlie gene expression
differences between microsatellite stable and unstable colorectal
cancers. Clin Cancer Res. 14:8061–8069. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hugo W, Zaretsky JM, Sun L, Song C, Moreno
BH, Hu-Lieskovan S, Berent-Maoz B, Pang J, Chmielowski B, Cherry G,
et al: Genomic and transcriptomic features of response to anti-PD-1
therapy in metastatic melanoma. Cell. 165:35–44. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Davis S and Meltzer PS: GEOquery: A bridge
between the gene expression omnibus (GEO) and BioConductor.
Bioinformatics. 23:1846–1847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pathan M, Keerthikumar S, Ang CS, Gangoda
L, Quek CY, Williamson NA, Mouradov D, Sieber OM, Simpson RJ, Salim
A, et al: FunRich: An open access standalone functional enrichment
and interaction network analysis tool. Proteomics. 15:2597–2601.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. Nat
Genet. 25:25–29. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Warde-Farley D, Donaldson SL, Comes O,
Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, Kazi F, Lopes CT,
et al: The GeneMANIA prediction server: Biological network
integration for gene prioritization and predicting gene function.
Nucleic Acids Res. 38 Suppl_2:W214–W220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinform. 4:22003. View Article : Google Scholar
|
|
30
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein–protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:D447–D452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES, et al: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang B and Horvath S: A general framework
for weighted gene co-expression network analysis. Stat Appl Genet
Mol Boil. Aug 12–2005.(Epub ahead of print). View Article : Google Scholar
|
|
34
|
Mizuno H, Kitada K, Nakai K and Sarai A:
PrognoScan: A new database for meta-analysis of the prognostic
value of genes. BMC Med Genomics. 2:182009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Abraham RT and Gibbons JJ: The mammalian
target of rapamycin signaling pathway: Twists and turns in the road
to cancer therapy. Clin Cancer Res. 13:3109–3114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han JM, Jeong SJ, Park MC, Kim G, Kwon NH,
Kim HK, Ha SH, Ryu SH and Kim S: Leucyl-tRNA synthetase is an
intracellular leucine sensor for the mTORC1-signaling pathway.
Cell. 149:410–424. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fingar DC and Blenis J: Target of
rapamycin (TOR): An integrator of nutrient and growth factor
signals and coordinator of cell growth and cell cycle progression.
Oncogene. 23:3151–3171. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choi MR, Yoo NJ, An CH and Lee SH:
Frameshift mutations in mammalian target of rapamycin pathway genes
and their regional heterogeneity in sporadic colorectal cancers.
Hum Pathol. 46:753–760. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin EI, Tseng LH, Gocke CD, Reil S, Le DT,
Azad NS and Eshleman JR: Mutational profiling of colorectal cancers
with microsatellite instability. Oncotarget. 6:42334–42344. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Laity JH, Lee BM and Wright PE: Zinc
finger proteins: New insights into structural and functional
diversity. Curr Opin Struct Boil. 11:39–46. 2001. View Article : Google Scholar
|
|
41
|
Kothe U: Recent progress on understanding
ribosomal protein synthesis. Comprehensive Natural Products II
Chemistry and Biology, Section Amino Acids, Peptides and Proteins
(Oxford). Elsevier. 2010.
|
|
42
|
Nieminen TT, O'Donohue MF, Wu Y, Lohi H,
Scherer SW, Paterson AD, Ellonen P, Abdel-Rahman WM, Valo S,
Mecklin JP, et al: Germline mutation of RPS20, encoding a ribosomal
protein, causes predisposition to hereditary nonpolyposis
colorectal carcinoma without DNA mismatch repair deficiency.
Gastroenterology. 147:595–598.e5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Longley DB, Harkin DP and Johnston PG:
5-fluorouracil: Mechanisms of action and clinical strategies. Nat
Rev Cancer. 3:330–338. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kurland CG and Maaløe O: Regulation of
ribosomal and transfer RNA synthesis. J Mol Boil. 4:193–210. 1962.
View Article : Google Scholar
|
|
45
|
Timmermann B, Kerick M, Roehr C, Fischer
A, Isau M, Boerno ST, Wunderlich A, Barmeyer C, Seemann P, Koenig
J, et al: Somatic mutation profiles of MSI and MSS colorectal
cancer identified by whole exome next generation sequencing and
bioinformatics analysis. PLoS One. 5:e156612010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
De Sousa E, Melo F, Wang X, Jansen M,
Fessler E, Trinh A, de Rooij LP, de Jong JH, de Boer OJ, van
Leersum R, Bijlsma MF, et al: Poor-prognosis colon cancer is
defined by a molecularly distinct subtype and develops from
serrated precursor lesions. Nat Med. 19:614–618. 2013. View Article : Google Scholar : PubMed/NCBI
|