Introduction
Acute lung injury (ALI) is characterized by serious
pulmonary inflammatory responses with a high incidence of morbidity
and mortality (1). Some progress
has been made in developing a therapeutic strategy and etiology,
although the mortality rate of patients with ALI remains high
(2). Consequently, there is a need
to identify innovative therapeutic strategies and effective
medications for the benefit of patients with ALI.
MicroRNAs (miRNAs; miRs) are a class of short
non-protein coding RNAs with lengths of 19–25 nucleotides, which
modulate target gene expression at the posttranscriptional level
(3). An increasing number of
studies have reported that miRNAs play critical roles in the
progression of several lung diseases, including ALI (4–6). For
example, Yang et al (5)
reported that the upregulation of miR-140-5p protected mice from
lipopolysaccharide (LPS)-induced ALI via the myeloid
differentiation primary response 88 (MyD88)/NF-κB pathway by
targeting Toll-like receptor (TLR)4. Xie et al (6) revealed that the downregulation of
miR-34b-5p attenuated inflammatory responses and apoptosis in
LPS-induced ALI of mice through targeting progranulin. However, the
effect of miR-26a-5p on LPS-induced ALI and its related molecular
mechanisms have not yet been reported.
Connective tissue growth factor (CTGF) is a
multifunctional matricellular protein that belongs to the
CTGF-Cyr61/Cef10-Nov (CCN) family. It was first reported in 1991 by
Bradham et al (7).
Increasing evidence has demonstrated that CTGF, a downstream
effector of transforming growth factor-β1 (TGF-β1), can induce lung
injury and contribute to pulmonary fibrosis (8–10). In
addition, CTGF has been reported to have an important role in
inflammatory diseases including rheumatoid arthritis, inflammatory
kidney diseases and silicosis (11). Previous studies have suggested that
miRNAs, such as miR-18a, regulates lung injury by modulating the
expression of CTGF and the TGF-β1 pathway (12). However, whether miR-26a-5p exerts
beneficial effects on the progression of LPS-induced ALI by
modulating CTGF remains unclear.
The present research aimed to identify the potential
effects of miRNA-26a-5p on LPS-induced ALI and its related
molecular mechanisms in vitro and in vivo. These data
confirmed that miR-26a-5p could attenuate lung inflammation and
apoptosis in LPS-induced ALI by targeting CTGF. Thus, miR-26a-5p
may serve as an important target for the treatment of ALI.
Materials and methods
Animals
A total of 230 male C57BL/6 mice (6–8 weeks; 20–25
g) were obtained from Beijing Vital River Laboratory Animal
Technology Co., Ltd. All mice were housed in climate-controlled
quarters with a 12-h light/dark cycle and provided water and
standard laboratory chow ad libitum. The experimental
procedures were performed in accordance with the Guide for the Care
and Use of Laboratory Animals and approved by the Ethics committee
of Zibo Women & Children Hospital.
Experimental protocol and animal
models of ALI
The mice were intratracheally injected with 5 mg/kg
LPS (Beijing Solabio Science & Technology C., Ltd.), dissolved
in phosphate buffer saline (PBS). A total of 24 h prior to LPS
treatment, the mice were administered with intraperitoneal
injections of miR-26a-5p mimic (20 mg/kg) or miR-26a-5p mimic
negative control (NC) (20 mg/kg) twice per day for 2 days
consecutively. Subsequently, the mice were randomly divided into
five groups (n=38/group): Control group (normal mice without any
treatment), Sham group (received PBS), LPS group (received LPS),
LPS + mimic negative control (NC) group (received LPS and
miR-26a-5p mimic NC) and LPS + miR-26a-5p mimic group (received LPS
and miR-26a-5p mimic). All mice were sacrificed by cervical
dislocation under anesthesia via intraperitoneal injection of 50
mg/kg pentobarbital sodium at 24 h after LPS treatment for the
subsequent experiment. The criteria used to confirm death were as
follows: Cessation of heartbeat for more than 5 min and no
pupillary reflex to strong light. The miR-26a-5p mimic and
miR-26a-5p mimic NC were purchased from Takara Biotechnology Co.,
Ltd. The sequences used were as follows: miR-26a-5p mimic,
5′-UUCAAGUAAUCCAGGAUAGGCU-3′ (sense); 5′-CCUAUCCUGGAUUACUUGAAUU-3′
(antisense); and miR-26a-5p mimic NC, 5′-UUCUCCGAACGUGUCACGUTT-3′
(sense); 5′-ACGUGACACGUUCGGAGAATT-3′ (antisense).
In addition, survival experiments were performed in
the four mice groups (n=10/group; Sham, LPS, LPS + mimic NC, LPS +
miR-26a-5p mimic). The survival of mice was monitored for 84 h.
Lung wet/dry (W/D) weight ratio
Lung edema was evaluated according to the wet/dry
weight ratio of lung tissues. After the mice (n=6) were sacrificed,
the right lungs were excised and immediately weighed to obtain the
wet weight. Subsequently, the lung samples were dried at 80°C for
48 h to obtain the dry weight (Lung wet/dry ratio=wet weight/dry
weight).
Myeloperoxidase (MPO) and
malondialdehyde (MDA) assay
After the mice (n=6) were sacrificed, the right lung
tissues were collected and homogenized in
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid containing 0.5%
cetyltrimethyl ammonium bromide. Then, the activity of MPO (cat.
no. A044) and the content of MDA (cat. no. A003-1) were measured by
using the corresponding test kits obtained from Nanjing Jiancheng
Bioengineering Institute in accordance with the manufacturer's
instructions.
Protein concentration and cell counts
in bronchoalveolar lavage fluid (BALF)
After the mice (n=6) were sacrificed, the BALF was
collected by injection and retraction of 1.5 ml PBS three times,
and was centrifuged at 1,200 × g for 10 min at 4°C. Subsequently,
the supernatants were collected for the assessment of total protein
concentration by the bicinchoninic acid (BCA) method. The cell
pellet was resuspended in PBS, and the neutrophil and lymphocyte
counts were determined using a hemocytometer following
Wright-Giemsa staining for 10 min at room temperature.
Hematoxylin and eosin (H&E)
staining
After the mice (n=5) were sacrificed, the right lung
tissues were collected and fixed in 4% formaldehyde at room
temperature for 24 h. Then, the lung tissues were dehydrated in
graded concentrations of ethanol, embedded in paraffin, and
transversely cut into 5-µm thick sections. Finally, the
histological changes in lung tissues were analyzed by H&E
staining under an optical microscope (magnification, ×100). In
addition, the histological score of the lung tissues was calculated
by assessing the alveolar congestion, hemorrhage, inflammatory cell
infiltration and alveolar wall thickness, and graded according to a
five-point scale from 0 to 4 as follows: 0=no damage, l=mild
damage, 2=moderate damage, 3=severe damage and 4=very severe damage
(13).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labelling (TUNEL) staining
After the mice (n=5) were sacrificed, the apoptotic
cells in the harvested lung tissues were detected by using the
TUNEL staining kit supplied by Shanghai Ruisai Biotechnology Co.,
Ltd. following the manufacturer's protocol. Five random fields were
viewed in each section and the rate of positive cells was analyzed
under a fluorescence microscope at a magnification of ×400.
Cell culture and transfection
The human type II alveolar epithelial cells (A549),
supplied by American Type Culture Collection, were maintained in
Dulbecco's Modified Eagle's Medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.), supplemented with 10% fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.) at 37°C in a humidified atmosphere with 5% CO2.
The miR-26a-5p mimic, pcDNA3.1-CTGF and their negative controls
were obtained from Takara Biotechnology Co., Ltd. Briefly, 10 nM
miR-26a-5p mimic or pcDNA3.1-CTGF were transfected into A549 cells
(6×106) using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), at room temperature
for 48 h according to the manufacturer's protocol. The transfected
A549 cells were randomly divided into the Control group (no
treatment), LPS group (treated with LPS), LPS + mimic NC group
(treated with LPS and miR-26a-5p mimic negative control), LPS +
miR-26a-5p mimic group (treated with LPS and miR-26a-5p mimic), LPS
+ miR-26a-5p mimic + oe-CTGF NC group (treated with LPS, miR-26a-5p
mimic and pcDNA3.1-CTGF NC) and LPS + miR-26a-5p mimic + oe-CTGF
(treated with LPS, miR-26a-5p mimic and pcDNA3.1-CTGF). After
transfection for 48 h, the A549 cells were treated with LPS
(Sigma-Aldrich; Merck KGaA) for 24 h and analyzed.
Cell viability assay
To assess cell viability,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was used. The transfected A549 cells were seeded into 96-well
plates at a density of 1×103 cells/well. After 24 h of
LPS stimulation, A549 cells were incubated with 20 µl of MTT at a
final concentration of 5 mg/ml (Sigma-Aldrich; Merck KGaA) in the
dark for 4 h. Subsequently, dimethyl sulfoxide (DMSO, 150 µl) was
added to each well. Finally, the cell viability was determined by
measuring absorbances at 490 nm by using a microplate reader.
Flow cytometry
The A549 cells undergoing apoptosis were detected
with an Annexin V-fluorescein isothiocyanate (FITC) apoptosis
detection kit (BD Biosciences) following the manufacturer's
instructions. Briefly, 24 h after LPS stimulation, the A549 cells
from different groups were harvested, washed with PBS and
resuspended in Annexin-binding buffer. Propidium iodide (PI) and
FITC-conjugated Annexin V were added into the cell suspensions and
maintained in the dark for 15 min at 37°C. The apoptotic cells were
then analyzed using a flow cytometer (BD FACScalibur; BD
Biosciences).
Enzyme-linked immunosorbent assay
(ELISA)
The levels of interleukin (IL)-1β, IL-6 and tumor
necrosis factor (TNF)-α in mice BALF (n=6) and A549 cells were
determined using an ELISA kit (BioLegend, Inc.) in accordance with
the manufacturer's instructions. The ELISA kits for mice BALF were
as follows: IL-1β (cat. no. 432601), IL-6 (cat. no. 431304) and
TNF-α (cat. no. 430904). The ELISA kits for A549 cells were as
follows: IL-1β (cat. no. 579409), IL-6 (cat. no. 430504) and TNF-α
(cat. no. 430204).
Dual luciferase reporter gene
assay
TargetScan (14) was
used to predict the targeting relationship between miR-26a-5p and
CTGF. The 3′-untranslated region (3′-UTR) of CTGF, with wild-type
or mutant binding sites for miR-26a-5p, was amplified and cloned
into the pGL3 luciferase vector (Promega Corporation) to generate
the plasmid pGL3-WT-CTGF-3′-UTR (CTGF-WT) or pGL3-Mut-CTGF-3′-UTR
(CTGF-MT). For the luciferase reporter assay, A549 cells were
co-transfected with reporter vectors and miR-26a-5p mimics or
miR-26a-5p mimics NC using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.). A total of 48 h after
the transfection, luciferase activity was detected by using a dual
luciferase kit (Promega Corporation). Firefly luciferase activity
was normalized to Renilla luciferase activity.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Lung tissue was collected after the mice (n=5) were
sacrificed. Total RNA was extracted from A549 cells and lung
tissues by TRIzol® Reagent (Thermo Fisher Scientific,
Inc.), based on the manufacturer's protocols. Total RNA (2 µg) was
added to the reverse transcription reaction to synthesize
complementary DNA (cDNA) using SuperScript™ IV First-Strand
Synthesis System (Invitrogen; Thermo Fisher Scientific, Inc.).
Thereafter, RT-qPCR was performed on a 7500 Real-time PCR System
with MirVana™ qRT-PCR miRNA (Invitrogen; Thermo Fisher Scientific,
Inc.). The PCR reaction conditions were as follows: 95°C for 10 min
followed by 40 cycles at 95°C for 10 sec, 60°C for 20 sec, and 72°C
for 30 sec. The primers used in the present study were as follows:
miR-26a-5p forward (F), 5′-GGATCCGCAGAAACTCCAGAGA-3′ and reverse
(R), 5′-TTGGAGGAAAGACGATTTCCGT-3′; U6 F, 5′-GCGCGTCGTGTAAAGCGTTC-3′
and R, 5′-GTGCAGGGTCCGAGGT-3′; CTGF F, 5′-CAAGGGCCTCTTCTGTGACT-3′
and R, 5′-ACGTGCACTGGTACTTGCAG-3′; B-cell lymphoma-2 (Bcl-2) F,
5′-ATGTGTGTGGAGAGCGTCAA-3′ and R, 5′-GCCGGTTCAGGTACTCAGTC-3′; Bax
F: 5′-TCTGACGGCAACTTCAACTG-3′, R: 5′-GGAGGAAGTCCAATGTCCAG-3′;
cleaved caspase-3 F: 5′-CTCGGTCTGGTACAGATGTCG-3′, R:
5′-TGGCTCAGAAGCACACAAAC-3′; GAPDH F, 5′-GCACCGTCAAGGCTGAGAAC-3′ and
R, 5′-ATGGTGGTGAAGACGCCAGT-3′. For the normalization of the
expression levels, U6 and GAPDH were used as internal controls. The
relative expression levels were calculated using the
2−ΔΔCq method (15).
Western blot analysis
Lung tissue was collected after the mice (n=5) were
sacrificed. Total proteins were extracted from A549 cells and lung
tissues by using lysis buffer (Beijing Solarbio Science &
Technology Co., Ltd.) containing phenylmethylsulfonyl fluoride
(PMSF). The concentration of proteins was measured with a BCA
protein assay kit (Beyotime Institute of Biotechnology). Protein
samples (50 µg) were loaded on 10% sodium dodecyl
sulfate-polyacrylamide gel for electrophoresis and then transferred
onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA). After
blocking with 5% skimmed milk for 2 h at room temperature, the
membranes were incubated at 4°C overnight with the following
primary antibodies supplied by Cell Signaling, Inc.: Bax (1:1,000;
product no. 14796), cleaved caspase-3 (1:1,000; product no. 9661),
Bcl-2 (1:1,000; product no. 3498), CTGF (1:1,000; product no.
86641) and GAPDH (1:1,000; product no. 5174). After washing with
TBST (20% Tween-20) three times, the membranes were incubated with
the horseradish peroxidase-conjugated anti-rabbit IgG secondary
antibody (1:5,000; cat. no. 7040; Cell Signaling Technology, Inc.)
at room temperature for 1 h. Finally, the protein bands were
visualized with an ECL system (Thermo Fisher Scientific, Inc.) and
then analyzed by using Image Lab™ Software (version 3.0; Bio-Rad
Laboratories, Inc.).
Statistical analysis
The statistical analysis was performed using SPSS
23.0 software (IBM Corp.). Data in the present study are presented
as the mean ± standard deviation (SD). Statistical differences were
analyzed using the unpaired Student's t-test or one-way analysis of
variance with Tukey's multiple comparison post-hoc test. Survival
rates were assessed by the Kaplan-Meier method, and survival curves
were compared by log-rank tests. P<0.05 was considered to
indicate a statistically significant difference. All experimental
data were obtained from at least three independent experiments.
Results
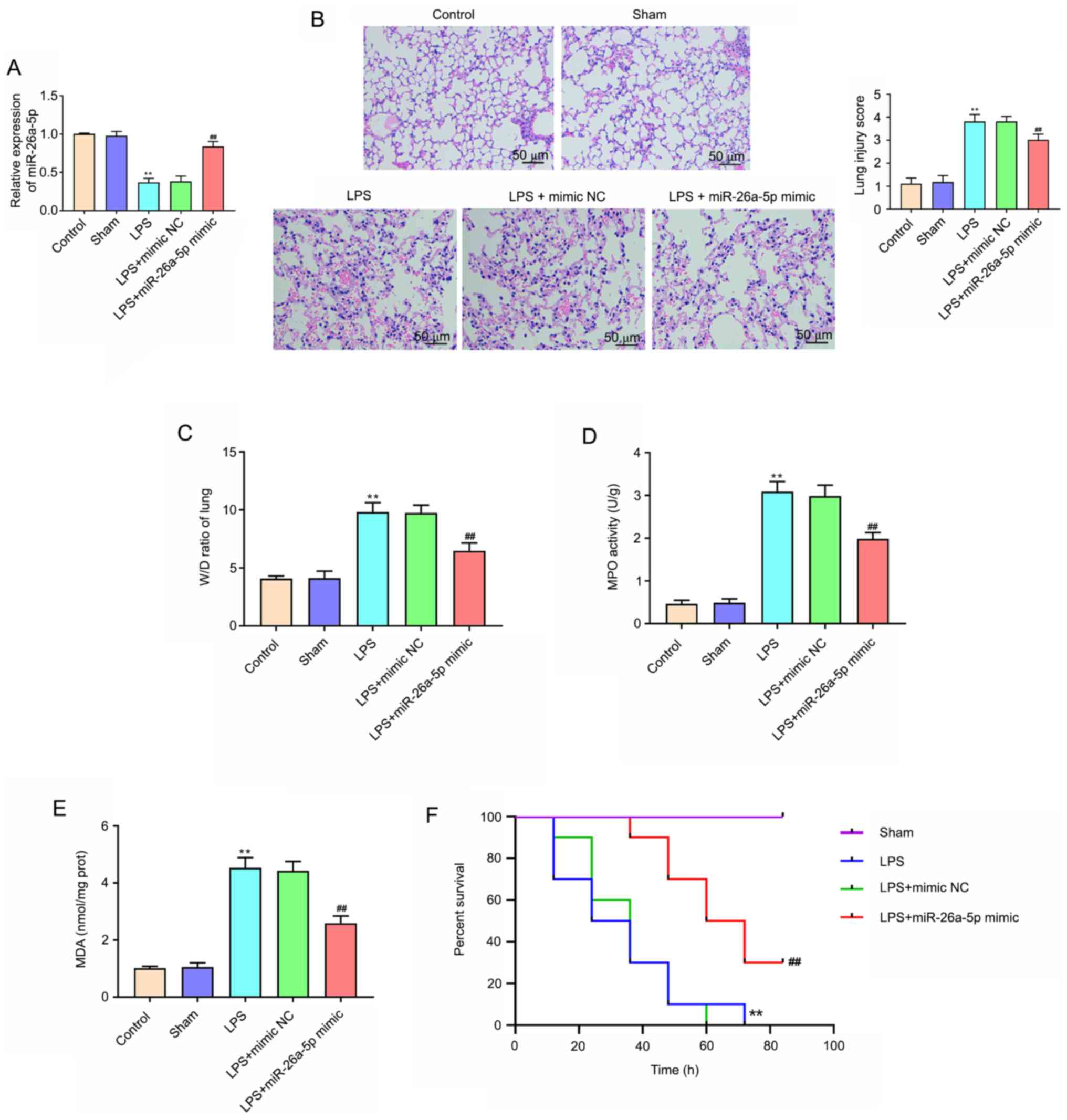
Overexpression of miR-26a-5p
alleviates LPS-induced acute lung injury in mice
The results of RT-qPCR revealed that the expression
of miR-26a-5p was significantly decreased in the LPS group compared
with the Sham group (P<0.01; Fig.
1A). Compared with the LPS + mimic NC group, the expression of
miR-26a-5p was significantly increased in the LPS + miR-26a-5p
mimic group (P<0.01; Fig. 1A).
In addition, to explore the effect of miR-26a-5p on LPS-induced
ALI, H&E staining and the W/D ratio of lung tissue were
performed. As revealed in Fig. 1B,
after the administration of LPS, the pulmonary edema, hemorrhage,
alveolar wall thickening and inflammatory cell infiltration were
markedly increased. Additionally, miR-26a-5p overexpression
significantly ameliorated the histological changes induced by LPS
(Fig. 1B). Lung injury scores also
confirmed the result. Moreover, Fig.
1C revealed that the lung wet/dry weight ratio was
significantly increased in the group treated with LPS compared with
the Sham group (P<0.01). When compared with the LPS + mimic NC
group, miR-26a-5p mimic significantly decreased the lung W/D ratio
(P<0.01). As revealed in Fig. 1D and
E, the activity of MPO and MDA content was significantly
increased in LPS group compared with that of the Sham group
(P<0.01). Correspondingly, upregulation of MPO and MDA was
reversed by the overexpression of miR-26a-5p (P<0.01). In
addition, the results revealed that LPS group mice survival rate
was significantly reduced compared with the Sham group mice
(P<0.01; Fig. 1F). Conversely,
the mice survival rate in the LPS + miR-26a-5p mimic group was
significantly longer than the LPS + mimic NC group (P<0.01;
Fig. 1F). Collectively, the results
indicated that the overexpression of miR-26a-5p could alleviate
LPS-induced ALI in mice.
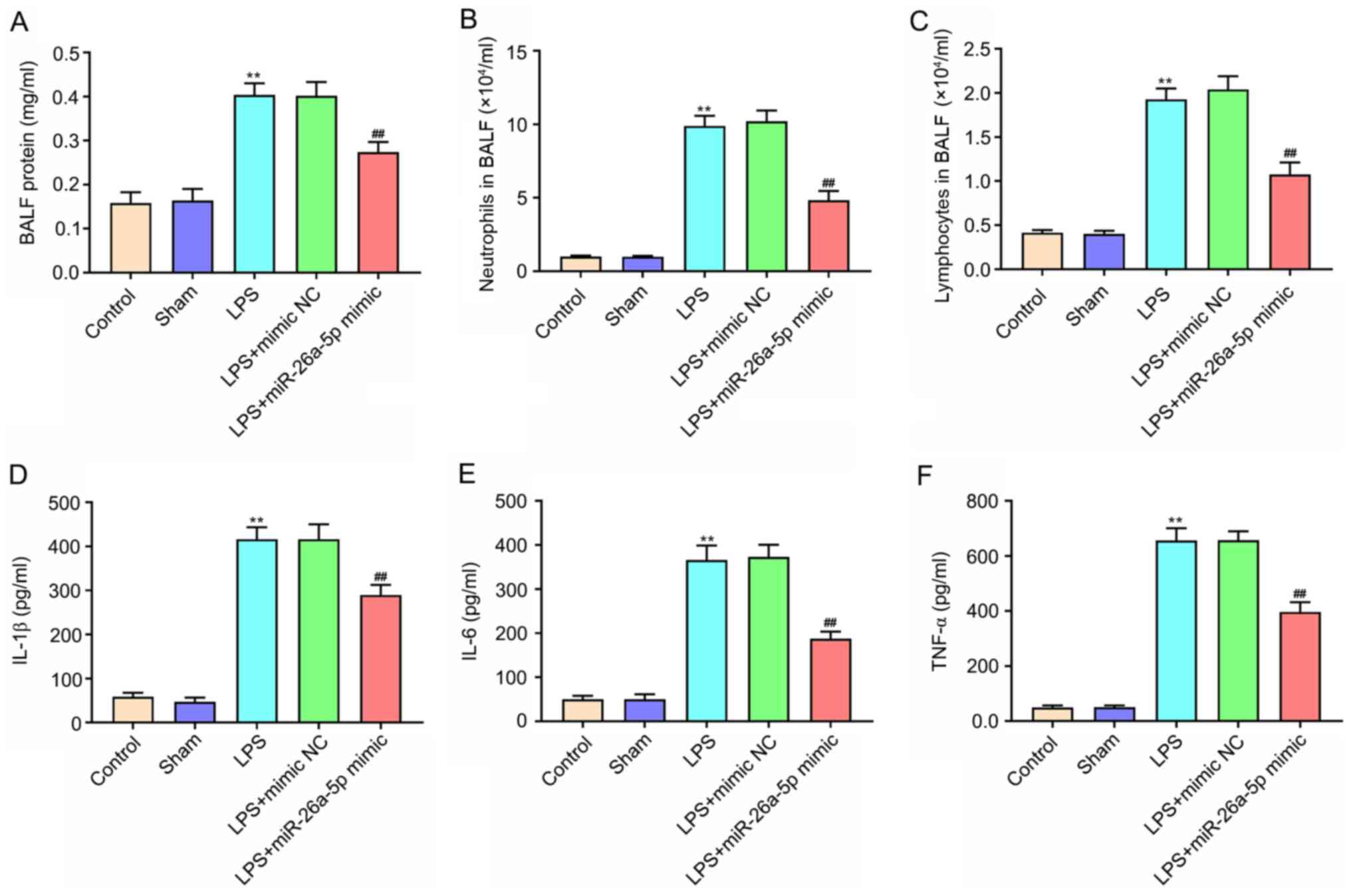
Overexpression of miR-26a-5p
alleviates lung tissue inflammation in LPS-induced ALI mice
The protein expression levels and the number of
neutrophils and lymphocytes in BALF were significantly increased in
the LPS group compared with the Sham group (P<0.01; Fig. 2A-C). In contrast, miR-26a-5p
overexpression significantly decreased the upregulation induced by
LPS (P<0.01). The ELISA results confirmed that the expression of
IL-1β, IL-6 and TNF-α were significantly increased after the
administration of LPS (P<0.01; Fig.
2D-F). Furthermore, the upregulation of IL-1β, IL-6 and TNF-α
was reversed by the overexpression of miR-26a-5p (P<0.01;
Fig. 2D-F). These results revealed
that the overexpression of miR-26a-5p could alleviate lung tissue
inflammation in LPS-induced ALI in mice.
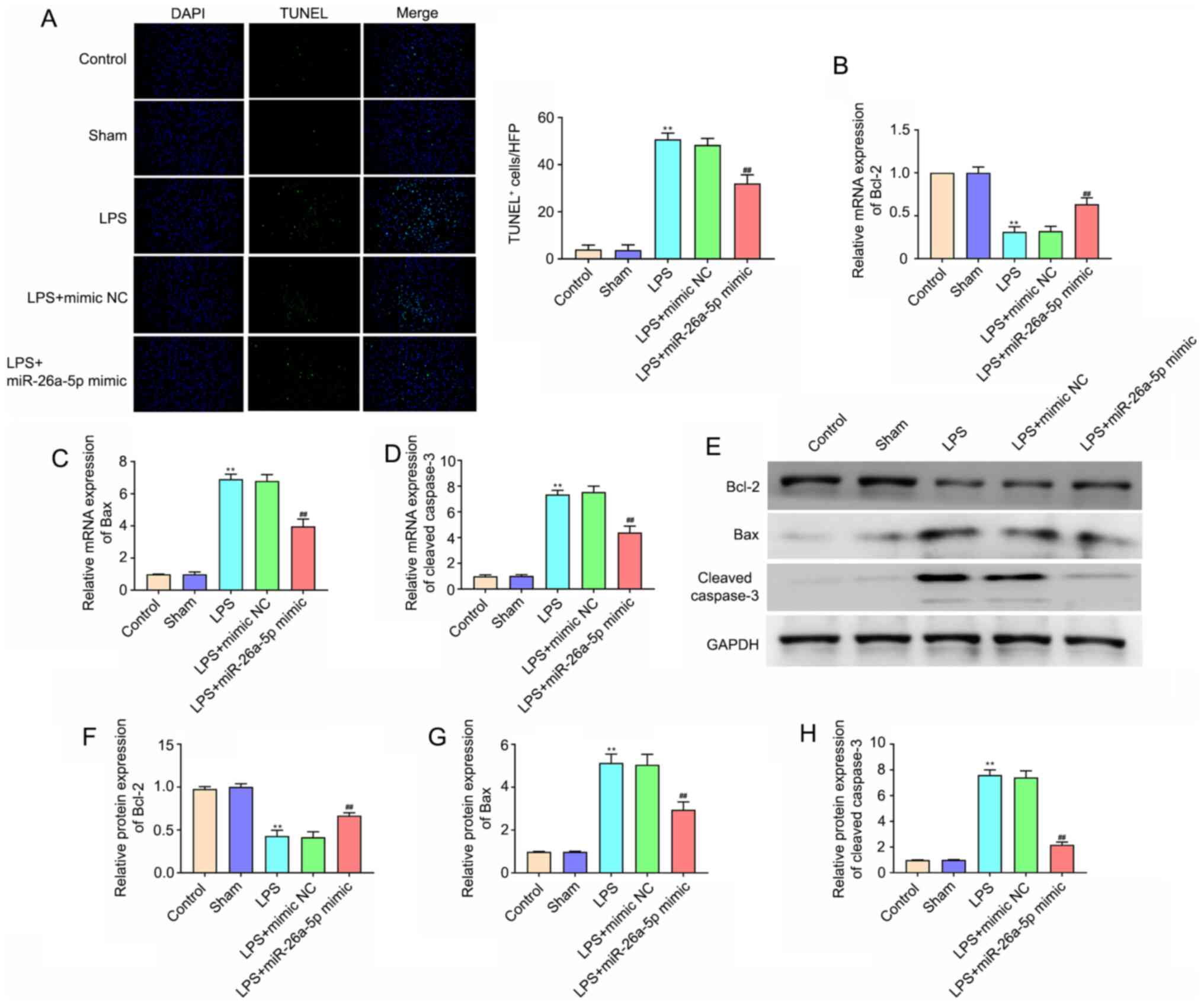
Overexpression of miR-26a-5p decreases
lung tissue cell apoptosis in LPS-induced ALI mice
To evaluate the cell apoptosis in lung tissues,
TUNEL staining was performed (Fig.
3A). The number of apoptotic cells in the lung tissues in the
LPS group was significantly increased compared with that in the
Sham group (P<0.01). miR-26a-5p overexpression also
significantly reduced the LPS-induced apoptosis of lung cells
(P<0.01). In addition, apoptosis-related proteins were detected
via RT-qPCR and western blotting. The results of Fig. 3B-H demonstrated that the mRNA and
protein expression levels of Bcl-2 in the LPS treatment group were
lower than those in the Sham group (P<0.01), whereas the
expression levels of Bax and cleaved caspase-3 were significantly
higher (P<0.01). In addition, the overexpression of miR-26a-5p
significantly reversed these effects on Bcl-2, Bax and cleaved
caspase-3 (P<0.01). All data indicated that the overexpression
of miR-26a-5p could decrease lung tissue cell apoptosis in
LPS-induced ALI in mice.
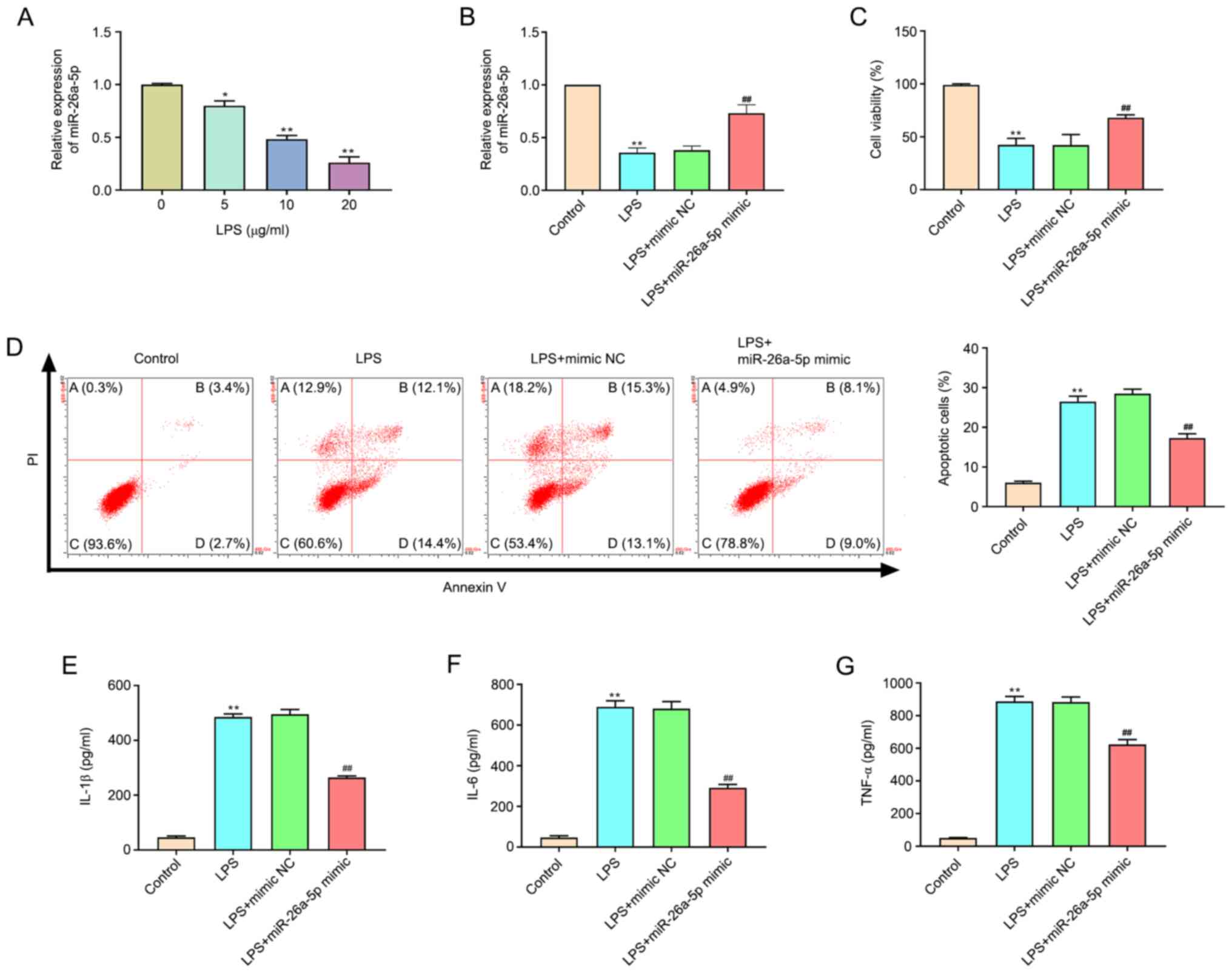
Overexpression of miR-26a-5p decreases
apoptosis and inflammatory responses in LPS-induced A549 cells
As revealed in Fig.
4A, the expression of miR-26a-5p in LPS-treated cells was
significantly decreased compared with the untreated cells in a
dose-dependent manner (P<0.05, P<0.01). A concentration of 20
µg/ml LPS was selected for subsequent experiments. Transfection
efficacy was confirmed by the significantly augmented expression of
miR-26a-5p at the mRNA expression level in A549 cells transfected
with the miR-26a-5p mimic (Fig.
4B). MTT assay results confirmed that the administration of LPS
significantly restricted cell viability (P<0.01; Fig. 4C). Correspondingly, miR-26a-5p
overexpression significantly increased the cell viability of A549
cells relative to the LPS + mimic NC group (P<0.01), which
indicated that miR-26a-5p overexpression increased the viability of
LPS-induced A549 cells. In addition, the results of flow cytometry
(Fig. 4D) revealed that the
administration of LPS significantly promoted A549 cell apoptosis
(P<0.01). However, the overexpression of miR-26a-5p
significantly reduced the cell apoptosis induced by LPS
(P<0.05). The present study also detected inflammatory cytokines
in A549 cells and determined that the expression levels of IL-1β,
IL-6 and TNF-α were significantly increased after LPS
administration (P<0.01; Fig.
4E-G). The upregulation of IL-1β, IL-6 and TNF-α was
significantly reversed by the overexpression of miR-26a-5p
(P<0.01). Altogether, the present data indicated that the
overexpression of miR-26a-5p could decrease apoptosis and
inflammatory responses in LPS-induced A549 cells.
CTGF is the target gene of
miR-26a-5p
TargetScan predicted that the binding site of
miR-26a-5p was the 3′-UTR region in CTGF (Fig. 5A). The dual luciferase reporter gene
assay results revealed that the overexpression of miR-26a-5p
significantly decreased the luciferase activity in the CTGF-WT
group (P<0.01; Fig. 5B);
however, it had no significant effect on the luciferase activity in
CTGF-MT group (P>0.05; Fig. 5B).
Furthermore, the study further elucidated whether miR-26a-5p
negatively regulated CTGF in A549 cells by RT-qPCR (Fig. 5C) and western blotting (Fig. 5D). The results revealed that the
mRNA and protein expression levels of CTGF in the LPS group were
higher than those in the Control group (P<0.01). miR-26a-5p also
significantly reversed the effect of LPS on CTGF in a
dose-dependent manner. These results indicated that CTGF was a
target gene of miR-26a-5p.
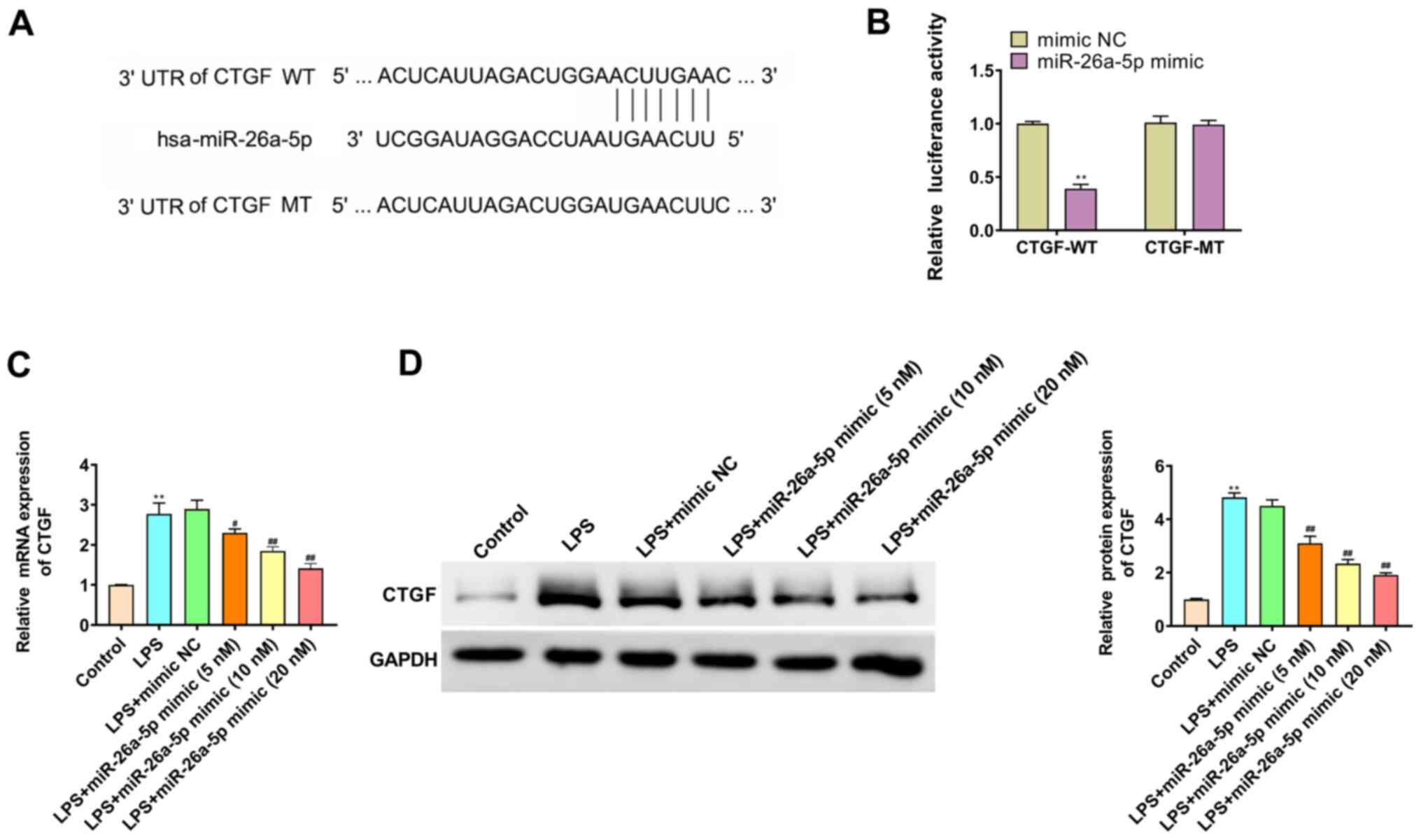 | Figure 5.CTGF is the target gene of
miR-26a-5p. (A) The binding target of miR-26a-5p and CTGF was
predicted by TargetScan software 7.2. (B) Luciferase activity of
CTGF-WT and CTGF-MT following the co-transfection of miR-26a-5p
mimic or mimic NC in A549 cells detected by dual-luciferase
reporter gene assay. (C) The mRNA expression of CTGF was detected
by reverse transcription-quantitative PCR in A549 cells transfected
with mimic NC or various concentrations (5, 10 and 20 nM) of
miR-26a-5p. (D) The protein expression of CTGF was detected by
western blotting in A549 cells transfected with mimic NC or various
concentrations (5, 10 and 20 nM) of miR-26a-5p. Data were presented
as the mean ± SD. (B) **P<0.01 vs. mimic NC group. (C and D)
**P<0.01 vs. Control group; #P<0.05,
##P<0.01 vs. LPS + mimic NC group. miR, microRNA; NC,
negative control; LPS, lipopolysaccharide; CTGF, connective tissue
growth factor; UTR, untranslated region; WT, wild-type; MT,
mutant. |
miR-26a-5p inhibits apoptosis and
inflammatory responses via targeting CTGF in A549 cells
As revealed in Fig. 6A
and B, the transfection efficacy of pcDNA3.1-CTGF, the CTGF
overexpression plasmid, was confirmed by the significantly enhanced
expression of CTGF at the mRNA and protein expression levels in
A549 cells. In addition, the results of Fig. 6C-G revealed that CTGF overexpression
significantly increased cell apoptosis and the expression levels of
IL-1β, IL-6 and TNF-α, and decreased cell viability (P<0.01).
Moreover, the overexpression of CTGF significantly reversed the
effects of miR-26a-5p mimic on cell viability, apoptosis and
inflammatory cytokines in LPS-induced A549 cells (P<0.01).
Collectively, this data indicated that miR-26a-5p could inhibit
apoptosis and inflammatory responses via targeting CTGF in A549
cells.
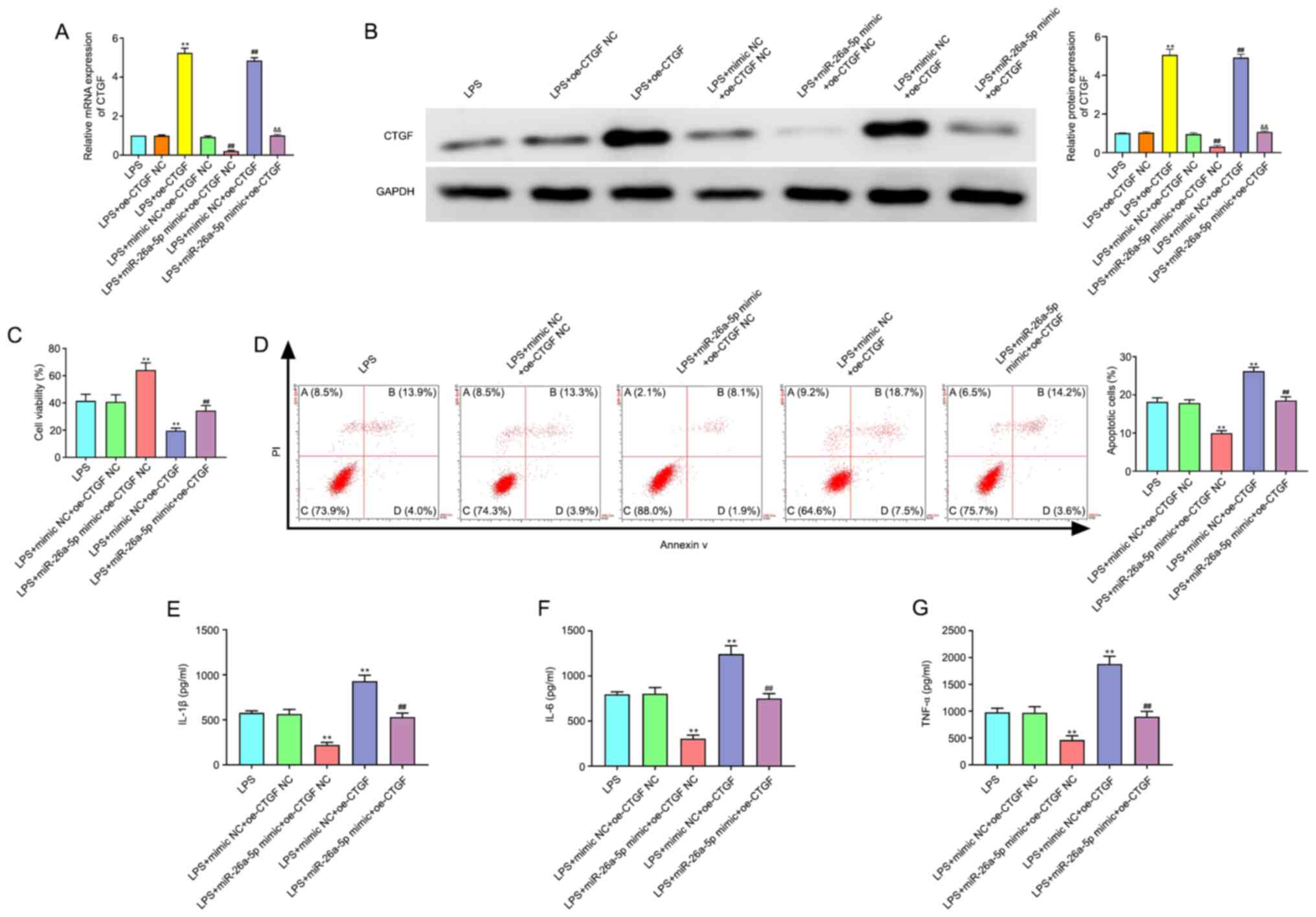 | Figure 6.CTGF overexpression reverses the
effect of miR-26a-5p on the apoptosis and inflammatory responses in
LPS-induced A549 cells. A549 cells were co-transfected with
miR-26a-5p mimic (10 nM), oe-CTGF (10 nM), mimic NC (10 nM) and
oe-CTGF NC (10 nM), and then treated with LPS for 24 h, followed by
the assessment of cell apoptosis and inflammatory response. (A) The
mRNA expression of CTGF was detected by reverse
transcription-quantitative PCR in transfected A549 cells. (B) The
protein expression of CTGF was measured by western blotting in
transfected A549 cells. (C) Cell viability was detected by MTT
assay in transfected A549 cells. (D) The cell apoptosis was
detected by flow cytometry in transfected A549 cells. The
expression levels of (E) IL-1β, (F) IL-6 and (G) TNF-α were
detected by ELISA in transfected A549 cells. Data were presented as
the mean ± SD. (A and B) **P<0.01 vs. LPS + oe-CTGF NC group;
##P<0.01 vs. LPS + mimic NC + oe-CTGF NC group;
&&P<0.01 vs. LPS + miR-26a-5p mimic + oe-CTGF
NC or LPS + mimic NC + oe-CTGF group. (C-G) **P<0.01 vs. LPS +
mimic NC + oe-CTGF NC group; ##P<0.01 vs. LPS +
miR-26a-5p mimic + oe-CTGF NC or LPS + mimic NC + oe-CTGF group.
miR, microRNA; NC, negative control; LPS, lipopolysaccharide; CTGF,
connective tissue growth factor; oe, overexpression plasmid. |
Discussion
ALI is a serious lung disease that is often
accompanied by acute inflammatory responses and can lead to
respiratory failure (16). At
present, the five-year survival rate of patients with ALI is
reported to be lower than 50% (17). Therefore, further studies focusing
on the development of novel therapeutic targets for the treatment
of ALI are required. This research has confirmed that miR-26a-5p
could attenuate lung inflammation and apoptosis in LPS-induced ALI
by targeting CTGF.
Recent studies have reported that miRNAs, as gene
expression switches, have an important role in the progression of
ALI (18,19). A recent study has indicated that
miR-802 was expressed at a low level in LPS-induced ALI and
alleviated LPS-induced ALI by targeting Peli2 (20). Another recent study has revealed
that miR-539-5p was underexpressed in sepsis-induced ALI and could
protect mice from ALI (21). In the
present study, the expression of miR-26a-5p was downregulated in
LPS-induced ALI models in vivo and in vitro. It
should be noted that miR-26a-5p has an important role in several
injury models. For instance, Wei et al (22) have reported that miR-26a-5p
overexpression could improve cerebral ischemia/reperfusion injury
via attenuating cell apoptosis. Xing et al (23) have indicated that miR-26a-5p could
protect against myocardial ischemia/reperfusion injury by
modulating the PTEN/PI3K/AKT pathway. However, the function of
miR-26a-5p in LPS-induced ALI is unclear. The present data revealed
that the overexpression of miR-26a-5p markedly improved LPS-induced
ALI by reducing the histopathological changes and W/D ratio in lung
tissues, and thereby helped to improve the survival of ALI
mice.
Previous studies have indicated that ALI is featured
with the production of inflammatory factors, inflammatory cell
infiltration and pulmonary epithelial cell apoptosis (24,25).
Evidence from several clinical studies has indicated that TNF-α,
IL-1β and IL-6 are the main inflammatory cytokines actively
secreted in response to the inflammatory cascade in ALI (21,26).
Previous studies have indicated that the excess inflammatory
cytokines, such as TNF-α, IL-6 and IL-1β, ultimately cause cell
death via apoptosis and pyroptosis (27). In addition, an increasing number of
studies have demonstrated that miRNAs could exert a protective
effect against LPS-induced ALI by inhibiting lung inflammation and
pulmonary epithelial cell apoptosis. For example, miR-27a could
alleviate lung inflammation and apoptosis in LPS-induced ALI mice
by modulating the TLR4/MyD88/NF-κB pathway (28). Suo et al (29) have confirmed that miR-1246 could
suppress ALI-induced inflammation and apoptosis by modulating NF-κB
and Wnt/β-catenin pathways. In addition, miR-26a-5p is considered
to be associated with inflammation and apoptosis in numerous
diseases. For example, the overexpression of miR-26a-5p could
markedly suppress neuropathic pain and neuroinflammation in rats
with chronic sciatic nerve injury (30). Wen et al (31) have suggested that miR-26a-5p
significantly decreased myocardial cell apoptosis and expression of
inflammatory factors in acute myocardial infarction by targeting
ADAM17. The present study also demonstrated that the overexpression
of miR-26a-5p significantly decreased LPS-induced apoptosis and
inflammatory factor expression levels in both ALI mice and cell
models, which suggests that miR-26a-5p could exert a protective
effect against LPS-induced ALI by inhibiting apoptosis and
inflammatory responses.
CTGF, also referred to as CCN family protein 2
(CCN2) and one of six members of cysteine-rich, secreted,
heparin-binding proteins with a modular structure, has an important
role in tissue remodeling and fibrosis (32). Increasingly, research has indicated
that CTGF is a downstream mediator of TGF-β1 that induces
connective tissue cell proliferation and extracellular matrix
deposition (8). The overexpression
of CTGF has been observed in numerous fibrotic tissues, including
kidney, lung, heart and liver, whereas in normal adult tissues or
cells expression is at low or even undetectable levels (33). The present data also revealed that
the expression of CTGF was significantly upregulated in LPS-induced
A549 cells. Previous studies have indicated that inhibiting the
upregulation of CTGF could attenuate bleomycin-induced ALI and
pulmonary fibrosis (34).
Furthermore, previous studies have reported that anti-CTGF therapy
with neutralizing CTGF monoclonal antibodies could improve
bleomycin-induced or radiation-induced lung fibrosis (9,35).
Evidence from several studies has suggested that miRNAs, such as
miR-18a and miR-26a, could modulate lung injury and lung fibrosis
via the regulation of CTGF (12,36).
Furthermore, a study by Li et al (37) has revealed that miR-26a-5p could
alleviate pneumonia by targeting CTGF. The present data also
confirmed that miR-26a-5p could negatively modulate the expression
of CTGF. In addition, the present results also revealed that CTGF
overexpression significantly reversed the effect of miR-26a-5p on
cell apoptosis and inflammatory cytokines in LPS-induced A549
cells. Collectively, these results indicated that miR-26a-5p could
inhibit apoptosis and inflammatory responses via targeting CTGF in
LPS-induced ALI.
In summary, the present study confirmed that the
expression of miR-26a-5p was downregulated in LPS-induced ALI mice
and in A549 cells, while CTGF expression was upregulated. In
addition, miR-26a-5p was demonstrated to attenuate lung
inflammation and apoptosis in LPS-induced ALI by targeting CTGF.
The findings of the present study provide new perspectives on
miRNA-based diagnostic approaches against LPS-induced ALI. A key
limitation of the present study was that only the effect of
miR-26a-5p on CTGF expression was assessed; therefore, further
investigation is required to verify the protective effect of
miR-26a-5p against LPS-induced ALI.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZF designed and conceived the study. HL and TY
performed the research, analyzed data and wrote the manuscript. All
authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The protocol of this research has been approved by
the Ethics Committee of Zibo Women & Children Hospital. The
experimental procedures were performed in accordance with the Guide
for the Care and Use of Laboratory Animals and approved by the
Ethics Committee of Zibo Women & Children Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Needham DM, Wozniak AW, Hough CL, Morris
PE, Dinglas VD, Jackson JC, Mendez-Tellez PA, Shanholtz C, Ely EW,
Colantuoni E, et al: Risk factors for physical impairment after
acute lung injury in a national, multicenter study. Am J Respir
Crit Care Med. 189:1214–1224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pan C, Liu L, Xie JF and Qiu HB: Acute
respiratory distress syndrome: Challenge for diagnosis and therapy.
Chin Med J (Engl). 131:1220–1224. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pattarayan D, Thimmulappa RK, Ravikumar V
and Rajasekaran S: Diagnostic Potential of extracellular microRNA
in respiratory diseases. Clin Rev Allergy Immunol. 54:480–492.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang Y, Liu D, Xi Y, Li J and Liu B:
Upregulation of miRNA-140-5p inhibits inflammatory cytokines in
acute lung injury through the MyD88/NF-κB signaling pathway by
targeting TLR4. Exp Ther Med. 16:3913–3920. 2018.PubMed/NCBI
|
|
6
|
Xie W, Lu Q, Wang K, Lu J, Gu X, Zhu D,
Liu F and Guo Z: miR-34b-5p inhibition attenuates lung inflammation
and apoptosis in an LPS-induced acute lung injury mouse model by
targeting progranulin. J Cell Physiol. 233:6615–6631. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bradham DM, Igarashi A, Potter RL and
Grotendorst GR: Connective tissue growth factor: A cysteine-rich
mitogen secreted by human vascular endothelial cells is related to
the SRC-induced immediate early gene product CEF-10. J Cell Biol.
114:1285–1294. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blom IE, Goldschmeding R and Leask A: Gene
regulation of connective tissue growth factor: New targets for
antifibrotic therapy? Matrix Biol. 21:473–482. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Alapati D, Rong M, Chen S, Hehre D,
Rodriguez MM, Lipson KE and Wu S: Connective tissue growth factor
antibody therapy attenuates hyperoxia-induced lung injury in
neonatal rats. Am J Respir Cell Mol Biol. 45:1169–1177. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo H, Ji F, Liu B, Chen X, He J and Gong
J: Peiminine ameliorates bleomycin-induced acute lung injury in
rats. Mol Med Rep. 7:1103–1110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou T, Yu Q, Lin H, Wang Z, Fu G, Lei L,
Shi Y, Zhang L, Qin L and Liu Y: The Role of CTGF in inflammatory
responses induced by silica particles in human bronchial epithelial
cells. Lung. 197:783–791. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang H, Li W, Zhang Y, Li M, Gao Y, Lao C
and Shi B: Regulatory role of miR-18a to CCN2 by TGF-β1 signaling
pathway in pulmonary injury induced by nano-SiO2.
Environ Sci Pollut Res Int. 25:867–876. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qi D, He J, Wang D, Deng W, Zhao Y, Ye Y
and Feng L: 17β-estradiol suppresses lipopolysaccharide-induced
acute lung injury through PI3K/Akt/SGK1 mediated up-regulation of
epithelial sodium channel (ENaC) in vivo and in vitro. Respir Res.
15:1592014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leung WS, Yang ML, Lee SS, Kuo CW, Ho YC,
Huang-Liu R, Lin HW and Kuan YH: Protective effect of zerumbone
reduces lipopolysaccharide-induced acute lung injury via
antioxidative enzymes and Nrf2/HO-1 pathway. Int Immunopharmacol.
46:194–200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Q and Xiao L: Isochlorogenic acid A
attenuates acute lung injury induced by LPS via Nf-κB/NLRP3
signaling pathway. Am J Transl Res. 11:7018–7026. 2019.PubMed/NCBI
|
|
18
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li W, Qiu X, Jiang H, Han Y, Wei D and Liu
J: Downregulation of miR-181a protects mice from LPS-induced acute
lung injury by targeting Bcl-2. Biomed Pharmacother. 84:1375–1382.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
You Q, Wang J, Jia D, Jiang L, Chang Y and
Li W: MiR-802 alleviates lipopolysaccharide-induced acute lung
injury by targeting Peli2. Inflamm Res. 69:75–85. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meng L, Cao H, Wan C and Jiang L:
MiR-539-5p alleviates sepsis-induced acute lung injury by targeting
ROCK1. Folia Histochem Cytobiol. 57:168–178. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei R, Zhang L, Hu W, Wu J and Zhang W:
Long non-coding RNA AK038897 aggravates cerebral
ischemia/reperfusion injury via acting as a ceRNA for miR-26a-5p to
target DAPK1. Exp Neurol. 314:100–110. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xing X, Guo S, Zhang G, Liu Y, Bi S, Wang
X and Lu Q: miR-26a-5p protects against myocardial
ischemia/reperfusion injury by regulating the PTEN/PI3K/AKT
signaling pathway. Braz J Med Biol Res. 53:e91062020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cox R Jr, Phillips O, Fukumoto J, Fukumoto
I, Parthasarathy PT, Arias S, Cho Y, Lockey RF and Kolliputi N:
Enhanced resolution of hyperoxic acute lung injury as a result of
aspirin triggered resolvin D1 treatment. Am J Respir Cell Mol Biol.
53:422–435. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu F, Zhang W, Yang F, Feng T, Zhou M, Yu
Y, Yu X, Zhao W, Yi F, Tang W and Lu Y: Interleukin-6-stimulated
progranulin expression contributes to the malignancy of
hepatocellular carcinoma cells by activating mTOR signaling. Sci
Rep. 6:212602016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ganter MT, Roux J, Miyazawa B, Howard M,
Frank JA, Su G, Sheppard D, Violette SM, Weinreb PH, Horan GS, et
al: Interleukin-1beta causes acute lung injury via alphavbeta5 and
alphavbeta6 integrin-dependent mechanisms. Circ Res. 102:804–812.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lv H, Liu Q, Wen Z, Feng H, Deng X and Ci
X: Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute
lung injury via induction of AMPK/GSK3β-Nrf2 signal axis. Redox
Biol. 12:311–324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ju M, Liu B, He H, Gu Z, Liu Y, Su Y, Zhu
D, Cang J and Luo Z: MicroRNA-27a alleviates LPS-induced acute lung
injury in mice via inhibiting in flammation and apoptosis through
modulating TLR4/MyD88/NF-κB pathway. Cell Cycle. 17:2001–2018.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suo T, Chen GZ, Huang Y, Zhao KC, Wang T
and Hu K: miRNA-1246 suppresses acute lung injury-induced
inflammation and apoptosis via the NF-κB and Wnt/β-catenin signal
pathways. Biomed Pharmacother. 108:783–791. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Su Z, Liu HL, Li L, Wei M, Ge DJ
and Zhang ZJ: Effects of miR-26a-5p on neuropathic pain development
by targeting MAPK6 in in CCI rat models. Biomed Pharmacother.
107:644–649. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wen X, Yin Y, Li X, He T, Wang P, Song M
and Gao J: Effect of miR-26a-5p targeting ADAM17 gene on apoptosis,
inflammatory factors and oxidative stress response of myocardial
cells in hypoxic model. J Bioenerg Biomembr. 52:83–92. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cicha I and Goppelt-Struebe M: Connective
tissue growth factor: Context-dependent functions and mechanisms of
regulation. Biofactors. 35:200–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gressner OA and Gressner AM: Connective
tissue growth factor: A fibrogenic master switch in fibrotic liver
diseases. Liver Int. 28:1065–1079. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim JW, Rhee CK, Kim TJ, Kim YH, Lee SH,
Yoon HK, Kim SC, Lee SY, Kwon SS, Kim KH and Kim YK: Effect of
pravastatin on bleomycin-induced acute lung injury and pulmonary
fibrosis. Clin Exp Pharmacol Physiol. 37:1055–1063. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ponticos M, Holmes AM, Shi-wen X, Leoni P,
Khan K, Rajkumar VS, Hoyles RK, Bou-Gharios G, Black CM, Denton CP,
et al: Pivotal role of connective tissue growth factor in lung
fibrosis: MAPK-dependent transcriptional activation of type I
collagen. Arthritis Rheum. 60:2142–2155. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li X, Liu L, Shen Y, Wang T, Chen L, Xu D
and Wen F: MicroRNA-26a modulates transforming growth factor
beta-1-induced proliferation in human fetal lung fibroblasts.
Biochem Biophys Res Commun. 454:512–517. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li C, Han T, Li R, Fu L and Yue L:
miR-26a-5p mediates TLR signaling pathway by targeting CTGF in
LPS-induced alveolar macrophage. Biosci Rep. 40:BSR201925982020.
View Article : Google Scholar : PubMed/NCBI
|