Introduction
Alzheimer's disease (AD) is a chronic
neurodegenerative disease characterized by memory loss and
progressive cognitive decline, which results from neuronal death in
numerous brain regions, including the hippocampus, entorhinal
areas, temporal and parietal lobes and restricted regions within
the frontal cortex and cingulate gyrus (1). According to a previous study, there
were 50 million patients with AD worldwide in 2020 and this number
is projected to reach 152 million by 2050 (2). AD pathology is characterized by
extracellular plaques formed by β-amyloid protein deposition,
neurofibrillary tangles formed by τ hyperphosphorylation and
neuronal loss caused by proliferation of glial cells (3). Numerous factors may influence the
incidence of AD, including aging, genetic and environmental factors
(4). However, the pathogenesis of
AD is still unclear. Previous studies have reported that aging and
environmental factors can affect the incidence of AD via epigenetic
modification and evidence suggests that epigenetic mechanisms,
including DNA methylation and histone modifications, could serve an
important role in the pathogenesis of AD (5–8).
RNA modification is a form of post-transcriptional
regulation and methylation is the primary form of this modification
(9). m6A is the most common type
of RNA modification of mRNA and is involved in the regulation of
numerous important biological processes, such as RNA degradation,
translation, RNA splicing and nuclear export (10). In the nervous system, m6A
methylation affects neuronal function, neurogenesis and neuronal
differentiation via regulation of mRNA splicing (11) and participates in the regulation of
the transcription of genes associated with brain disease risk
(12). Increasing studies have
reported that m6A serves an important regulatory role in brain
development, synaptic plasticity, learning and memory (13,14).
According to a previous report, specific m6A methylation exists in
the mouse cerebral cortex and cerebellum tissue (15). A previous study reported that
protein expression levels of METTL3, which encodes the m6A
methyltransferase, is downregulated in the hippocampus (16). The accumulation of METTL3
has been reported to be positively correlated with levels of
insoluble τ protein in postmortem human AD samples (16). Furthermore, overexpression of
METTL3 has been reported to have rescued β-amyloid
(Aβ)-induced synaptic damage and cognitive impairment in
vivo (17). Our previous study
reported that total methylation level of m6A RNA in the hippocampus
of AD mouse model is significantly higher compared with that of the
control group, which indicates that m6A methylation of RNA promotes
the development of AD (18).
Circular (circ)RNAs are evolutionarily conserved
non-coding RNAs without 5′caps and 3′polyadenylation tails
(19). circRNAs have been reported
to serve a role in the regulation of neurodegenerative disease
through their interaction with disease-related miRNAs in numerous
studies (20,21). circRNAs have received growing
attention for their involvement in the pathogenesis of AD (22,23).
A circRNA microarray study reported that AD mouse models had
abnormal circRNA in the hippocampus (24,25).
m6A RNA methylation has also been reported in circRNAs (26). A review by Li and Jin (27) reported that circRNAs serve an
important role in the production and clearance of Aβ, AD
neuroinflammation, oxidative stress and autophagy, and that
circRNAs are widely involved in regulation of AD physiological and
pathophysiological processes and may have the potential to be new
biomarkers and novel therapeutic targets. Furthermore, circRNA
ciRS-7 has been reported to act as an endogenous, anticomplementary
microRNA (miRNA or miR) ‘sponge’ to adsorb and affect normal
miRNA-7 function (28).
Downregulation of ciRS-7 might increase endogenous miRNA-7 levels
in AD. Due to inhibition by the ‘sponging’ effects of ciRS-7,
expression of AD-associated targets, such as ubiquitin protein
ligase UBE2A, an autophagic, phagocytic protein essential in the
clearance of amyloid peptides in the AD brain, may be downregulated
(28).
In the present study, high-throughput sequencing was
used to assess dysregulation of the circRNA m6A profile in the
brains of APP/PS1 and control mice, and evaluate the differences.
Furthermore, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) analysis was used to evaluate the biological roles
and signaling pathways of these differentially modified circRNA
m6A. The methylation degree of circRNA m6A was detected using
methylated RNA immunoprecipitation reverse
transcription-quantitative PCR (MeRIP-RT-qPCR).
Materials and methods
Mouse model
APP/PS1 double-transgenic male mice (age, 9 months)
were used as AD models (10 per group) and age-matched C57BL/6
wild-type mice (9 months old; n=20) were used as the control group.
All 20 mice were purchased from Beijing Huafukang Biotechnology
Co., Ltd. All mice were housed at a controlled room temperature
(25±2°C), in 50–60% humidity, with 12-h light and dark cycles, and
free access to water and food for 2 weeks. The mice were sacrificed
using cervical dislocation; lack of breathing and cardiac arrest
were considered to confirm death. The cortex, hippocampus and
cerebellum of mice were dissected and immediately frozen using
liquid nitrogen before being stored at −80°C for use in future
experiments. All procedures were performed according to the
guidelines of the Ethical Committee for Animal Experiments of
Shandong University (Jinan, Shandong, China) and the study protocol
was approved by the Ethical Committee for Animal Experiments of
Shandong University [approval no. KYLL-2020(KJ)A-0098].
All of the 20 mice were eventually euthanized. The
humane endpoints used in the present study were as follows: Labored
breathing, nasal discharge, lethargy or persistent recumbency,
difficulty with ambulation or inability to obtain food or
water.
RNA extraction
Total RNA was isolated from frozen mouse brain
samples using TRIzol® reagent (Thermo Fisher Scientific,
Inc.). A DeNovix spectrophotometer (DS-11) was used to determine
RNA quality. Samples with an A260/A280 ratio 1.9-2.2 were used for
further experiments.
MeRIP high-throughput sequencing
circRNA m6A high-throughput sequencing (circRNA
m6A-seq) was performed by Cloudseq Biotech, Inc. High-throughput
sequencing of circRNA m6A methylation was performed on hippocampal
samples from APP/PS1 and C57BL/6 control mice (3 per group). All
the following steps were performed by Cloudseq Biotech Inc.
Briefly, RNA fragments were treated with anti-m6A polyclonal
antibody (cat. no. 202003; Synaptic Systems GmbH) in
immunoprecipitation (IP) buffer [150 mM NaCl, 0.1% NP-40 and 10 mM
Tris-HCl (pH 7.4)] for 2 h at 4°C. After that, the mixture was
immunoprecipitated by incubating it for 2 h at 4°C with 50 µl
protein A beads (Thermo Fisher Scientific, Inc.). The bound RNA was
eluted from the beads using m6A (Berry & Associates; Biosearch
Technologies) in IP buffer and extracted using TRIzol reagent
(Thermo Fisher Scientific, Inc.). The NEBNext® Ultra II
Directional RNA Library Prep kit (cat. no. NEB#E7760; New England
Biolabs, Inc.) was used to obtain purified RNA for the creation of
an RNA-seq library. RNA quantification and quality assurance with
the NanoDrop ND-1000 (Thermo). The concentration of the control
group was 561.6 ng/µl and the concentration of the experimental
group was 1,240.23 ng/µl. GenSeq® m6A MeRIP kit
(GS-ET-001) via Illumina HiSeq 4000 sequencer was used for 150-bp
paired-end sequencing of input samples without IP and the m6A IP
samples with 6 ng of total RNA input. The differentially expressed
genes of circRNA m6A methylation of the samples from the two groups
were then examined. Methylated sites were identified by MACS2
(1.4.2) software. Differentially methylated sites were identified
by diffReps (1.55.6). The distinct physiological activities of
these genes were analyzed using GO and KEGG pathway analysis, where
P<0.05 was considered to indicate a statistically significant
difference. The generated data are available under accession number
GSE216901 from the Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/).
MeRIP-RT-qPCR
Total RNA (1 µg) was extracted from mouse
hippocampal samples and cDNA was synthesized using the PrimeScript
First Strand cDNA Synthesis Kit (cat. no. RR047A; Takara Bio, Inc.)
according to the manufacturer's instructions. RT-qPCR was performed
using the SYBR-Green assay (cat. no. RR041A; Takara Bio, Inc.) and
a StepOnePlus Real-Time PCR system (Mastercycler; Eppendorf) in
order to determine the relative RNA levels of target genes. RT-qPCR
using gene-specific primers was performed on the input control and
the m6A-IP samples. The thermocycling conditions were as follows:
40 cycles of 95°C for 30 sec, 95°C for 15 sec and 55°C for 15 sec.
The primers used for RT-qPCR are presented in Table I.
 | Table I.Sequences of primers used for reverse
transcription-quantitative PCR. |
Table I.
Sequences of primers used for reverse
transcription-quantitative PCR.
| Target | Sequence
(5′-3′) |
|---|
|
chr16:85056324-85120697- | F:
TTAGTGAATGCCGGTGATGC |
|
| R:
ACCACAACCACCACTGAGTC |
|
chr16:85013627-85030365+ | F:
AAATGGGCATGCTCGTTCTC |
|
| R:
TGTGTGTGTGTGTGTGTTGG |
|
chr12:16977542-16978102+ | F:
CGACAGCTTGCTCCAAACAA |
|
| R:
TTTAATGTGGCACCTACGGC |
|
chr16:8708859-8716619- | F:
CCTCTTCTGCGTTGCTGTG |
|
| R:
GCGCCCCATTACATTTTGAAG |
|
chr11:108923152-108931579+ | F:
ACTGACCGACGATTCCATGT |
|
| R:
AGGAGAGTCACTAACACGGC |
|
chrX:143709648-143744001+ | F:
TCAGGAAGCATTGTGAGTGT |
|
| R:
AGTGAATTCCCCGGTGACTG |
|
chr5:137291102-137291478+ | F:
TTTCTTCCTCCACGCCTTGT |
|
| R:
AGGACGGATGAAACCCAGAA |
|
chr1:68305845-68396315- | F:
ATGAACCATGACACACAGGC |
|
| R:
AGATGGACTCCTCACATGCC |
Statistical analysis
Data are presented as the mean ± SEM of at least
three independent experiments. Unpaired student's t-test was used
to assess statistical differences between groups. P<0.05 was
considered to indicate a statistically significant difference. SPSS
17.0 software (SPSS Inc.) was used for data analysis.
Results
Differential expression between
APP/PS1 and wild-type mice
Compared with the control group, high-throughput
sequencing demonstrated that certain circRNA m6A genes were
methylated at higher or lower levels in APP/PS1 AD mice. A total of
537 genes demonstrated an increase in methylation in the AD group;
however, a decrease in methylation was demonstrated in 1,359 genes.
The results demonstrated that there was a total of 4,918
methylation sites; upregulated genes accounted for 10.92% of
possible sites and the downregulated genes accounted for 27.63% of
possible sites. A total of 50 upregulated and 50 downregulated
genes (ranked according to fold-change) are presented in Table SI.
GO and KEGG pathway analysis of
differentially methylated circRNA m6A genes
To evaluate the biological role of circRNA m6A
methylation in AD mice, the genes containing significantly altered
m6A peaks, which indicated differentially methylated genes (DMGs),
were analyzed using GO and KEGG pathway analysis. There was a
marked difference in degree of methylation between the AD and
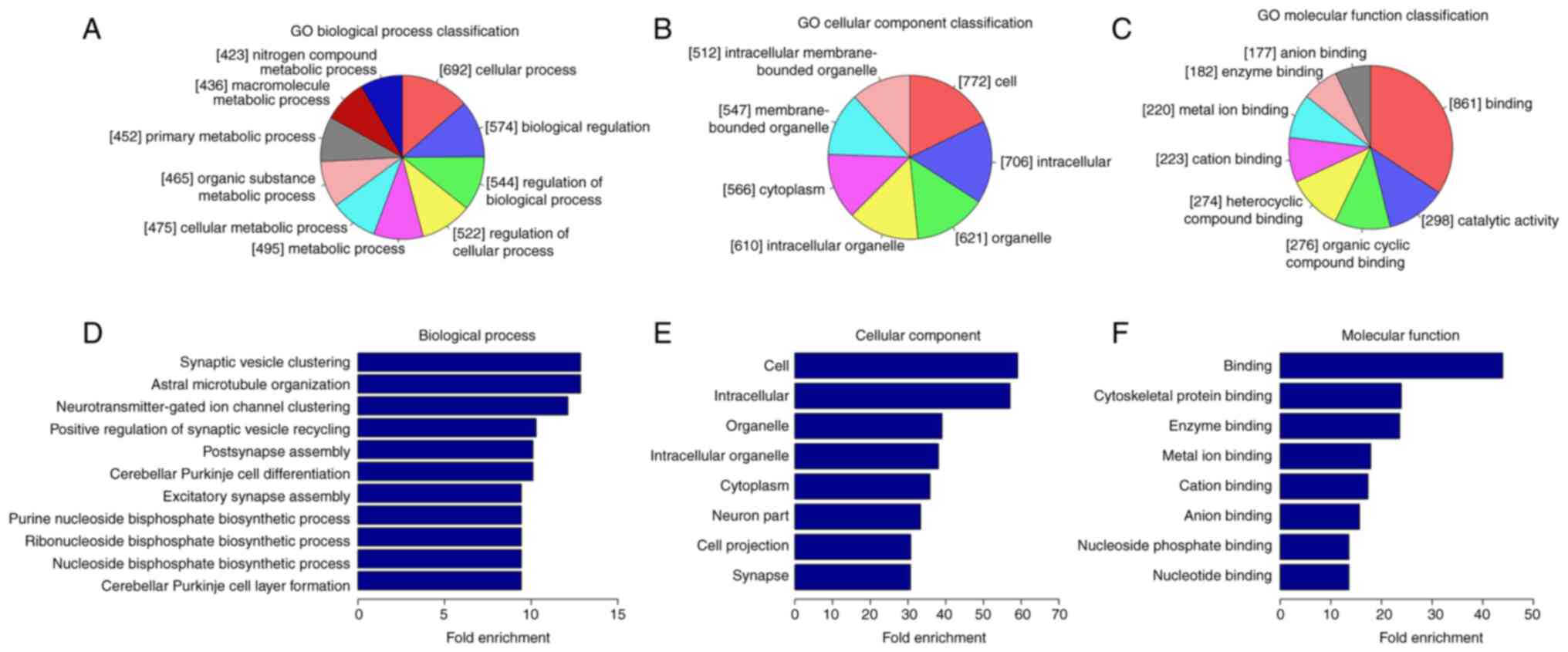
control groups. The three parts of GO analysis (Fig. 1) are biological process (BP), cell
component (CC) and molecular function (MF). BP analysis
demonstrated that the genes with lower circRNA m6A methylation in
the AD group were significantly linked with ‘synaptic vesicle
clustering’, ‘astral microtubule organization’,
‘neurotransmitter-gated ion channel clustering’, ‘positive
regulation of synaptic vesicle recycling’, ‘postsynapse assembly’
and ‘cerebellar Purkinje cell differentiation’. Significant GO CC
terms for differentially modified RNAs demonstrated that these
circRNAs were associated with ‘cell’, ‘intracellular’, ‘organelle’
and ‘cytoplasm’. For MF, these circRNAs were associated with
‘cytoskeletal protein binding’, ‘enzyme binding’, ‘metal ion
binding’, ‘cation binding’ and ‘anion binding’.
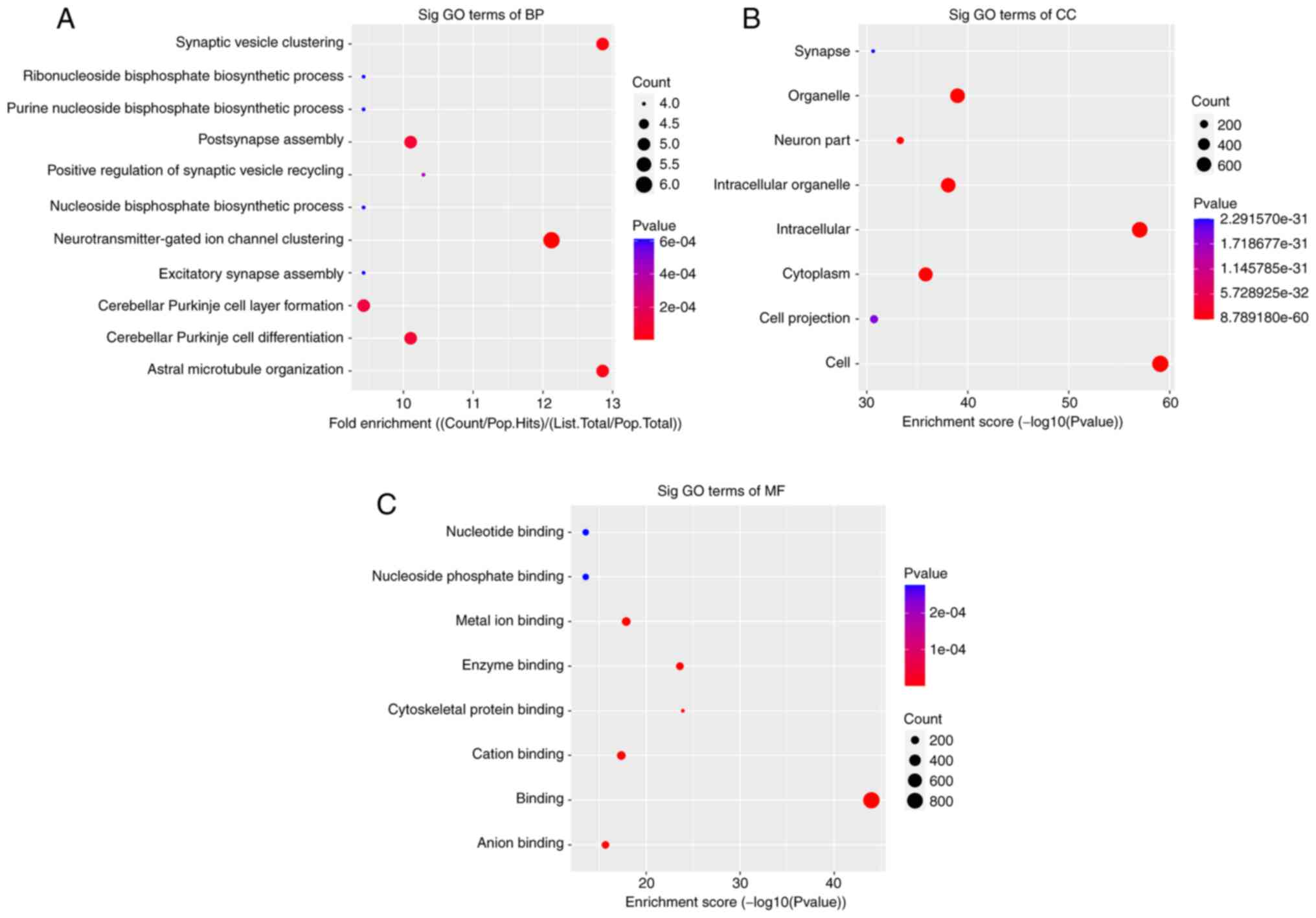
The significant enrichment pathways with the top 10
enrichment scores [log10(P-value)] were assessed using the KEGG
pathway dot-plot. The KEGG pathway for BP was associated with
‘synaptic vesicle clustering’, ‘postsynapse assembly’,
‘neurotransmitter-gated ion channel clustering’, ‘cerebellar
Purkinje cell layer formation’, ‘cerebellar Purkinje cell
differentiation’ and ‘astral microtubule organization’ (Fig. 2A). For CC, the KEGG pathway was
associated with ‘organelle’, ‘intracellular part’, ‘intracellular
organelle’, ‘cytoplasm’ and ‘cell part’ (Fig. 2B). In MR, the KEGG pathways were
enriched for ‘metal ion binding’, ‘enzyme binding’, ‘cation
binding’ and ‘anion binding’ (Fig.
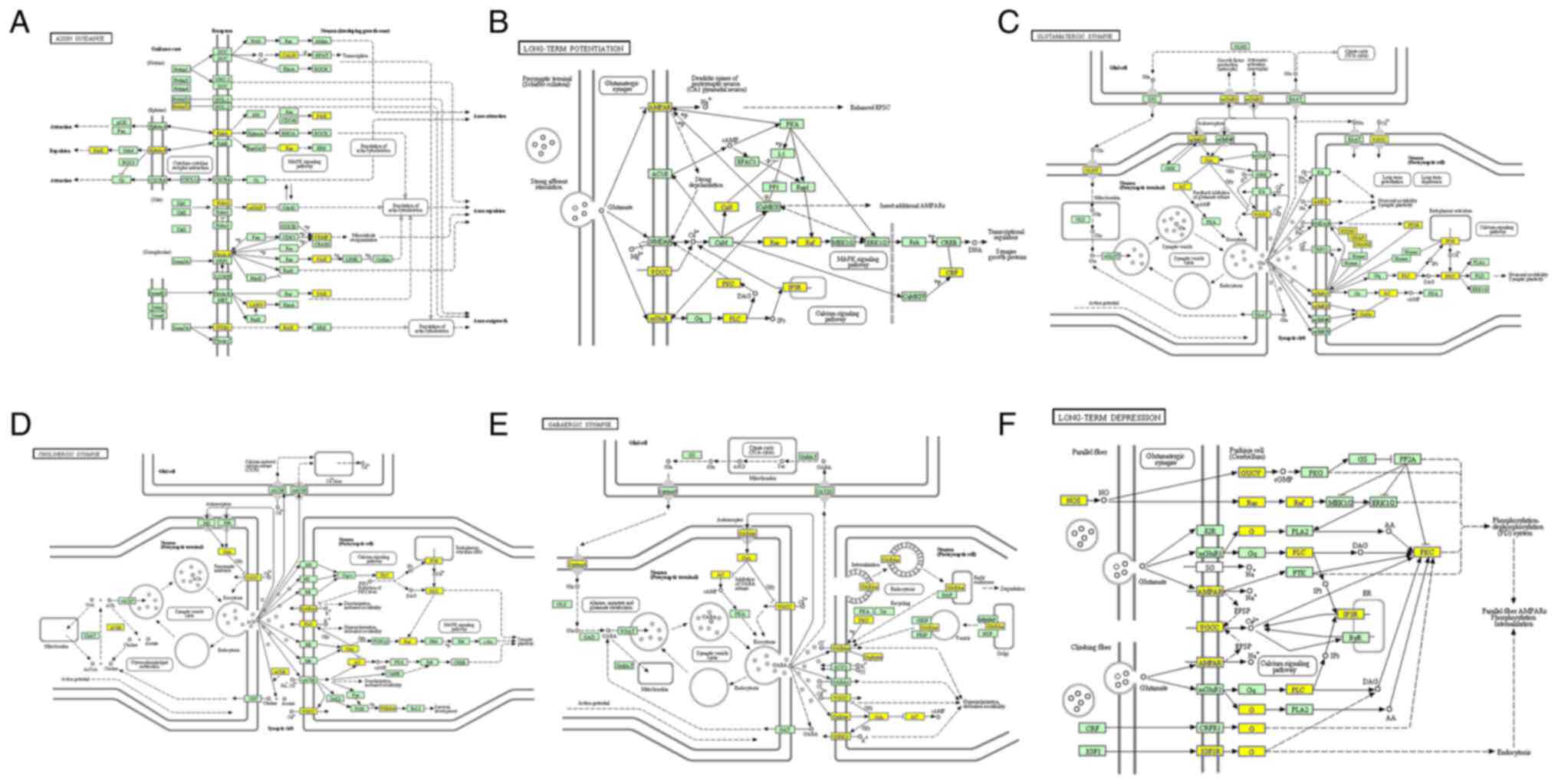
2C). Furthermore, the six primary pathways were associated with
‘axon guidance’, ‘long-term potentiation’, ‘glutamatergic synapse,
‘cholinergic synapse’, ‘GABAergic synapse’ and ‘long-term
depression’ (Fig. 3).
circRNA m6A methylation of gene
expression is different
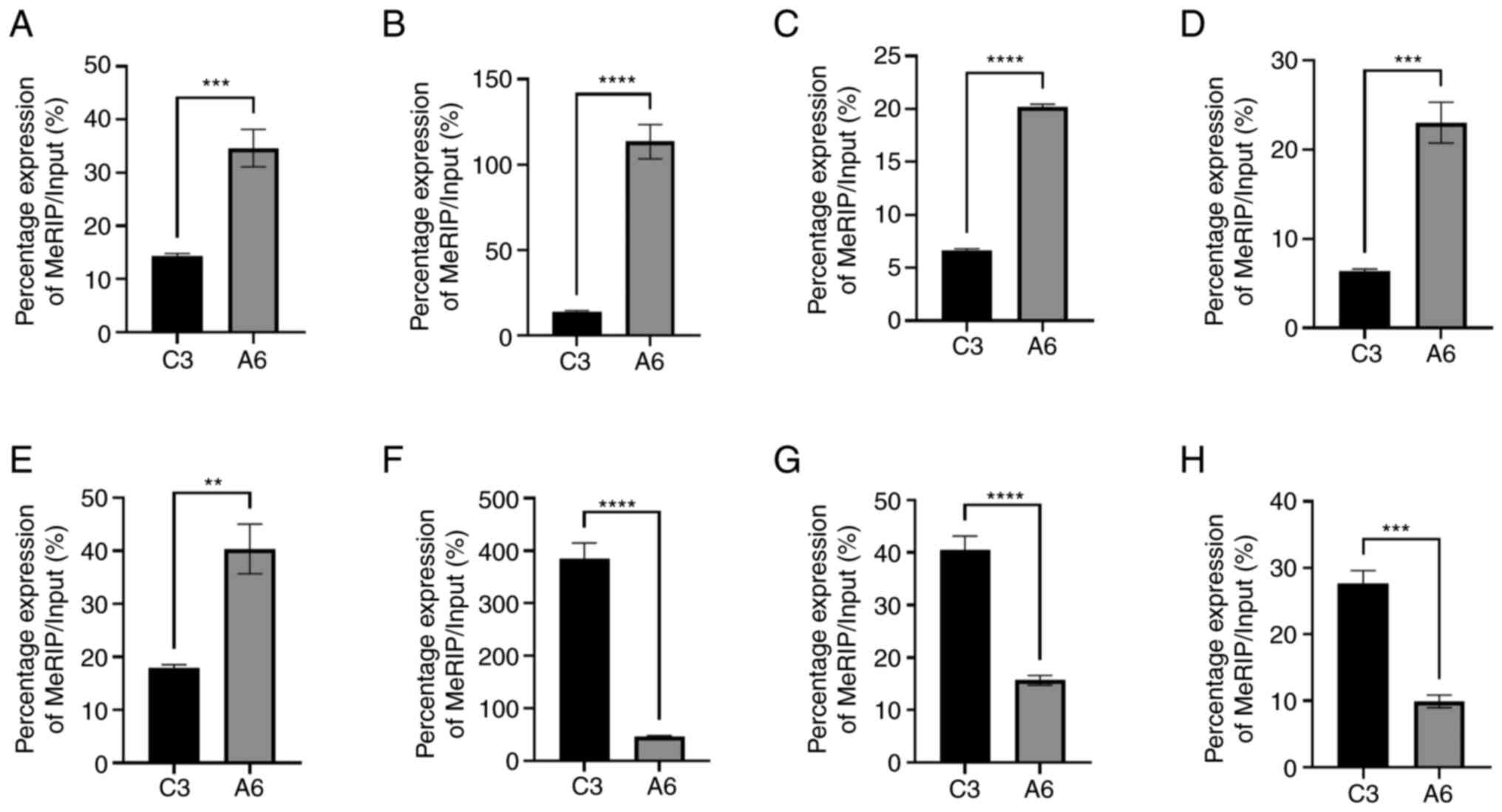
The results of high-throughput sequencing analysis
demonstrated that the degree of methylation in the AD group was
different from that in the control group. Eight differentially
expressed genes were selected and methylation levels were assessed
using RT-qPCR. Compared with the control group, among the eight
selected genes, 5 demonstrated significantly higher methylation
levels, and the remaining three demonstrated significantly lower
methylation levels (Fig. 4).
Combined with the aforementioned high-throughput sequencing
results, these results suggested that circRNA m6A methylation was
different in the brain of the AD mice compared with the normal
mice.
Discussion
A perfect mouse model of AD would develop
neuropathology as well as behavioral changes associated with AD
(29). Double transgenic mice
(APP/PS1) have been widely used to study the pathogenesis of AD and
assess the effectiveness of AD treatments (30–32).
The APP/PS1 mouse model of AD demonstrates a significant increase
in β-amyloid production associated with certain behavioral
abnormalities. Behavioral tests have previously demonstrated that
compared with wild-type mice, transgenic APP/PS1 mice have
significant memory deficits at 6 months of age and that this
deficit is exacerbated and more errors occurred at 12 months of
age. Cognitive deficits and reference memory impairments of APP/PS1
mouse appear before 6 months of age and lasted for the rest of the
life (30,33,34).
Deficits in associative learning have also been described in the
fear conditioning task at 6–8 months of age and passive avoidance
deficits were demonstrated at 12 months of age (35). In the present study, the APP/PS1
mouse was used as an animal model of AD.
In eukaryotes, m6A is the most prevalent internal
RNA modification and regulates all aspects of RNA metabolism
(16). m6A has been associated
with neurogenesis (36), learning
and memory (37), brain
development (12,38–40)
and axon regeneration (41) in
previous studies. The presence of specific m6A methylation in mouse
cerebral cortex and cerebellum tissue was also reported in a
previous study (38). In our
previous study, it was reported that the methylation genes in the
AD group were associated with presynaptic membranes, postsynaptic
membranes and synaptic growth, which suggested that m6A was likely
involved in AD (18). All these
results indicated that m6A is associated with AD.
circRNAs are a type of RNA made through reverse
splicing, in which the 3′ and 5′ ends of a transcript are
chemically spliced together to form a continuous loop. In
eukaryotes, many circRNAs have been reported and the nervous system
and synapses are reported to be particularly rich in these circRNAs
(42). A recent study reported
aberrant regulation of circRNAs in AD and three circRNAs that may
provide clinical insight into AD risk and progression (43). circRNAs have been reported to
demonstrate the same m6A modification as mRNAs but are frequently
found in distinct places; the m6A readers YTHDF1 and YTHDF2
interact with m6A circRNAs, while the m6A writer METTL3 regulates
m6A levels (44).
In the present study, high-throughput sequencing on
the degree of circRNA m6A methylation in the hippocampus, cortex
and cerebellum in APP/PS1 and normal mice as controls was
performed. In comparison to control mice, AD animals demonstrated
significantly different levels of m6A methylation of certain
circRNAs.
In patients with AD, synapse loss and damage are the
most likely causes of cognitive decline (45,46).
Furthermore, previous studies have reported that m6A is linked to
synaptic plasticity (47,48). According to a recent study, a novel
circRNA is linked to psychiatric disorder and regulates synaptic
gene expression and cognitive flexibility (49). In the present study, the KEGG
pathway results demonstrated that circRNA m6A methylation
alteration was associated with glutamatergic, cholinergic and
GABAergic synapses. These results indicated that circRNA m6A
methylation may have been involved in AD pathogenesis.
To evaluate the aforementioned results, 8
differentially expressed genes were selected from the
high-throughput sequencing data and MeRIP-RT-qPCR was performed.
These genes were associated with AD. chr16:85056324-85120697- and
chr16:85013627-85030365+ are part of the amyloid-β precursor
protein (APP) gene and mutations in this gene are linked to the
development of early onset (familial) AD (50). chr12: 16977542-16978102+ is a
fragment of the Rock2 gene, which has been reported to be
correlated with Aβ production (51). ch16:8708859-8716619- is associated
with the ubiquitin specific peptidase 7 gene and has an
anti-neuroinflammatory effect (52). chr11:108923152-108931579+ is part
of the Axin2 gene, which serves an important role in the
Wnt/β-catenin signaling pathway, which inhibits Aβ production and τ
protein hyperphosphorylation in the brain (53). chrX:143709648-143744001+ is part of
the p21 activated kinase gene, which regulates DNA synthesis and
neuronal apoptosis caused by familial AD mutants of APP (54). chr5:137291102-137291478+, part of
the acetylcholinesterase gene, has been reported to be linked to
pathogenesis either by increasing cholinergic deficit or
exacerbating Aβ fibril formation and toxicity in AD (55). Furthermore, chr1:68305845-68396315-
is reported to be associated with Erbb4, which mediates amyloid
β-induced neurotoxicity via JNK/τ signaling pathway activation
(56). The results demonstrated
that compared with the control group, AD mouse brain samples
demonstrated increased methylation levels of five circRNAs and
decreased methylation levels of three circRNAs. This suggested that
there were differences in levels of methylation in AD mice and that
circRNA m6A methylation was related to AD.
In summary, the present study demonstrated that
compared with the control group, AD mouse brain samples had
different levels of circRNA m6A methylation. MeRIP-RT-qPCR
supported these data and indicated that circRNA m6A methylation may
have been involved in AD pathogenesis. However, the specific
molecular mechanism of the differentially expressed circRNA m6A
genes in AD was not thoroughly studied and the potential binding
miRNAs were not elucidated. These data suggested that circRNA m6A
methylation was associated with AD but more research is needed to
confirm this association. In the future, the circRNA of the APP
gene associated with Aβ production should be evaluated
(chr16:85056324-85120697-) and MeRIP-RT-qPCR verification of
methylation level of circAPP m6A in the hippocampus of APP/PS1 mice
should be performed. APP/PS1 cells could be used to study the
specific regulation mechanism of circAPP m6A on Aβ production in an
AD cell model via regulation of APP.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was support by The Special Scientific Research
Project of Health Young Medical Science and Technology Talents in
Xinjiang Uygur Autonomous Region (grant no. WJWY-202210), The
National Natural Science Foundation of China (grant no. 82171410
and 81870848) and The Key Research and Development Program of
Shandong Province (grant no. 2017GSF218046).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request. The datasets generated and/or analyzed during the current
study are available in the GEO repository (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE216901).
Authors' contributions
XZ and YWa designed and performed the experiments,
collected and assembled the data, and wrote the manuscript. YS, MH,
FL and YWe analyzed data. SH, SY and XLZ performed data analysis
and interpretation. YWa and JB were responsible for the conception
and design of the study, and the revision and final approval of the
manuscript. All authors have read and approved the final
manuscript. XZ and YWa confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
The present study protocol was reviewed and approved
by The Ethical Committee for Animal Experiments of Shandong
University [approval no. KYLL-2020(KJ)A-0098].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lane CA, Hardy J and Schott JM:
Alzheimer's disease. Eur J Neurol. 25:59–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yiannopoulou KG and Papageorgiou SG:
Current and future treatments in Alzheimer disease: An update. J
Cent Nerv Syst Dis. 12:11795735209073972020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang JZ, Wang ZH and Tian Q: Tau
hyperphosphorylation induces apoptotic escape and triggers
neurodegeneration in Alzheimer's disease. Neurosci Bull.
30:359–366. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kwok JB: Role of epigenetics in
Alzheimer's and Parkinson's disease. Epigenomics. 2:671–682. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma X, Liu L and Meng J: MicroRNA-125b
promotes neurons cell apoptosis and tau phosphorylation in
Alzheimer's disease. Neurosci Lett. 661:57–62. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Patrick E, Rajagopal S, Wong HA, McCabe C,
Xu J, Tang A, Imboywa SH, Schneider JA, Pochet N, Krichevsky AM, et
al: Dissecting the role of non-coding RNAs in the accumulation of
amyloid and tau neuropathologies in Alzheimer's disease. Mol
Neurodegener. 12:512017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Irier HA and Jin P: Dynamics of DNA
methylation in aging and Alzheimer's disease. DNA Cell Biol. 31
(Suppl 1):S42–S48. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Unnikrishnan A, Freeman WM, Jackson J,
Wren JD, Porter H and Richardson A: The role of DNA methylation in
epigenetics of aging. Pharmacol Ther. 195:172–185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Y, Chen YS, Sun BF and Yang YG: RNA
methylation: Regulations and mechanisms. Yi Chuan. 40:964–976.
2018.(In Chinese). PubMed/NCBI
|
|
10
|
Shafik AM, Zhang F, Guo Z, Dai Q, Pajdzik
K, Li Y, Kang Y, Yao B, Wu H, He C, et al: N6-methyladenosine
dynamics in neurodevelopment and aging, and its potential role in
Alzheimer's disease. Genome Biol. 22:172021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haussmann IU, Bodi Z, Sanchez-Moran E,
Mongan NP, Archer N, Fray RG and Soller M: m6A
potentiates Sxl alternative pre-mRNA splicing for robust Drosophila
sex determination. Nature. 540:301–304. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoon KJ, Ringeling FR, Vissers C, Jacob F,
Pokrass M, Jimenez-Cyrus D, Su Y, Kim NS, Zhu Y, Zheng L, et al:
Temporal control of mammalian cortical neurogenesis by
m6A methylation. Cell. 171:877–889.e17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Widagdo J and Anggono V: The
m6A-epitranscriptomic signature in neurobiology: From
neurodevelopment to brain plasticity. J Neurochem. 147:137–152.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Flamand MN and Meyer KD: The
epitranscriptome and synaptic plasticity. Curr Opin Neurobiol.
59:41–48. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L, Du K, Wang J, Nie Y, Lee T and
Sun T: Unique and specific m6A RNA methylation in mouse
embryonic and postnatal cerebral cortices. Genes (Basel).
11:11392020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang H, Camats-Perna J, Medeiros R,
Anggono V and Widagdo J: Altered expression of the m6A
methyltransferase METTL3 in Alzheimer's disease. eNeuro.
7:ENEURO.0125–20.2020. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao F, Xu Y, Gao S, Qin L, Austria Q,
Siedlak SL, Pajdzik K, Dai Q, He C, Wang W, et al: METTL3-dependent
RNA m6A dysregulation contributes to neurodegeneration
in Alzheimer's disease through aberrant cell cycle events. Mol
Neurodegener. 16:702021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han M, Liu Z, Xu Y, Liu X, Wang D, Li F,
Wang Y and Bi J: Abnormality of m6A mRNA methylation is involved in
Alzheimer's disease. Front Neurosci. 14:982020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen LL: The biogenesis and emerging roles
of circular RNAs. Nat Rev Mol Cell Biol. 17:205–211. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsuzaki M, Honkura N, Ellis-Davies GC
and Kasai H: Structural basis of long-term potentiation in single
dendritic spines. Nature. 429:761–766. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Noureddini M and Bagheri-Mohammadi S:
Adult hippocampal neurogenesis and Alzheimer's disease: Novel
application of mesenchymal stem cells and their role in hippocampal
neurogenesis. Int J Mol Cell Med. 10:1–10. 2021.PubMed/NCBI
|
|
22
|
Lukiw WJ: Circular RNA (circRNA) in
Alzheimer's disease (AD). Front Genet. 4:3072013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang JL, Su M and Wu DP: Functional roles
of circular RNAs in Alzheimer's disease. Ageing Res Rev.
60:1010582020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang JL, Qin MC, Zhou Y, Xu ZH, Yang SM,
Zhang F, Zhong J, Liang MK, Chen B, Zhang WY, et al: Comprehensive
analysis of differentially expressed profiles of Alzheimer's
disease associated circular RNAs in an Alzheimer's disease mouse
model. Aging (Albany NY). 10:253–265. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Z, Xu P, Chen B, Zhang Z, Zhang C,
Zhan Q, Huang S, Xia ZA and Peng W: Identifying
circRNA-associated-ceRNA networks in the hippocampus of
Aβ1-42-induced Alzheimer's disease-like rats using microarray
analysis. Aging (Albany NY). 10:775–788. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang L, Hou C, Chen C, Guo Y, Yuan W, Yin
D, Liu J and Sun Z: The role of N6-methyladenosine
(m6A) modification in the regulation of circRNAs. Mol
Cancer. 19:1052020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li W and Jin G: The circRNA and role in
Alzheimer's disease: From regulation to therapeutic and diagnostic
targets. Hippocampus. 2022.PubMed/NCBI
|
|
28
|
Akhter R: Circular RNA and Alzheimer's
disease. Adv Exp Med Biol. 1087:239–243. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lok K, Zhao H, Shen H, Wang Z, Gao X, Zhao
W and Yin M: Characterization of the APP/PS1 mouse model of
Alzheimer's disease in senescence accelerated background. Neurosci
Lett. 557(Pt B): 84–89. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiong H, Callaghan D, Wodzinska J, Xu J,
Premyslova M, Liu QY, Connelly J and Zhang W: Biochemical and
behavioral characterization of the double transgenic mouse model
(APPswe/PS1dE9) of Alzheimer's disease. Neurosci Bull. 27:221–232.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Heneka MT, Kummer MP, Stutz A, Delekate A,
Schwartz S, Vieira-Saecker A, Griep A, Axt D, Remus A, Tzeng TC, et
al: NLRP3 is activated in Alzheimer's disease and contributes to
pathology in APP/PS1 mice. Nature. 493:674–678. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Izco M, Martínez P, Corrales A, Fandos N,
García S, Insua D, Montañes M, Pérez-Grijalba V, Rueda N, Vidal V,
et al: Changes in the brain and plasma Aβ peptide levels with age
and its relationship with cognitive impairment in the APPswe/PS1dE9
mouse model of Alzheimer's disease. Neuroscience. 263:269–279.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Denver P, English A and McClean PL:
Inflammation, insulin signaling and cognitive function in aged
APP/PS1 mice. Brain Behav Immun. 70:423–434. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Götz J, Bodea LG and Goedert M: Rodent
models for Alzheimer disease. Nat Rev Neurosci. 19:583–598. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Webster SJ, Bachstetter AD, Nelson PT,
Schmitt FA and Van Eldik LJ: Using mice to model Alzheimer's
dementia: An overview of the clinical disease and the preclinical
behavioral changes in 10 mouse models. Front Genet. 5:882014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Li Y, Yue M, Wang J, Kumar S,
Wechsler-Reya RJ, Zhang Z, Ogawa Y, Kellis M, Duester G and Zhao
JC: N6-methyladenosine RNA modification regulates
embryonic neural stem cell self-renewal through histone
modifications. Nat Neurosci. 21:195–206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shi H, Zhang X, Weng YL, Lu Z, Liu Y, Lu
Z, Li J, Hao P, Zhang Y, Zhang F, et al: m6A facilitates
hippocampus-dependent learning and memory through YTHDF1. Nature.
563:249–253. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang M, Lv H, Zhang W, Ma C, He X, Zhao
S, Zhang ZW, Zeng YX, Song S, Niu Y and Tong WM: Region-specific
RNA m6A methylation represents a new layer of control in
the gene regulatory network in the mouse brain. Open Biol.
7:1701662017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dominissini D, Moshitch-Moshkovitz S,
Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K,
Jacob-Hirsch J, Amariglio N, Kupiec M, et al: Topology of the human
and mouse m6A RNA methylomes revealed by m6A-seq. Nature.
485:201–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma C, Chang M, Lv H, Zhang ZW, Zhang W, He
X, Wu G, Zhao S, Zhang Y, Wang D, et al: RNA m6A
methylation participates in regulation of postnatal development of
the mouse cerebellum. Genome Biol. 19:682018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Weng YL, Wang X, An R, Cassin J, Vissers
C, Liu Y, Liu Y, Xu T, Wang X, Wong SZH, et al: Epitranscriptomic
m6A regulation of axon regeneration in the adult
mammalian nervous system. Neuron. 97:313–325.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dube U, Del-Aguila JL, Li Z, Budde JP,
Jiang S, Hsu S, Ibanez L, Fernandez MV, Farias F, Norton J, et al:
An atlas of cortical circular RNA expression in Alzheimer disease
brains demonstrates clinical and pathological associations. Nat
Neurosci. 22:1903–1912. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li Y, Fan H, Sun J, Ni M, Zhang L, Chen C,
Hong X, Fang F, Zhang W and Ma P: Circular RNA expression profile
of Alzheimer's disease and its clinical significance as biomarkers
for the disease risk and progression. Int J Biochem Cell Biol.
123:1057472020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhou C, Molinie B, Daneshvar K, Pondick
JV, Wang J, Van Wittenberghe N, Xing Y, Giallourakis CC and Mullen
AC: Genome-wide maps of m6A circRNAs identify widespread and
cell-type-specific methylation patterns that are distinct from
mRNAs. Cell Rep. 20:2262–2276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kashyap G, Bapat D, Das D, Gowaikar R,
Amritkar RE, Rangarajan G, Ravindranath V and Ambika G: Synapse
loss and progress of Alzheimer's disease-a network model. Sci Rep.
9:65552019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
John A and Reddy PH: Synaptic basis of
Alzheimer's disease: Focus on synaptic amyloid beta, P-tau and
mitochondria. Ageing Res Rev. 65:1012082021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chokkalla AK, Mehta SL and Vemuganti R:
Epitranscriptomic regulation by m6A RNA methylation in
brain development and diseases. J Cereb Blood Flow Metab.
40:2331–2349. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Madugalle SU, Meyer K, Wang DO and Bredy
TW: RNA N6-methyladenosine and the regulation of RNA
localization and function in the brain. Trends Neurosci.
43:1011–1023. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zimmerman AJ, Hafez AK, Amoah SK,
Rodriguez BA, Dell'Orco M, Lozano E, Hartley BJ, Alural B, Lalonde
J, Chander P, et al: A psychiatric disease-related circular RNA
controls synaptic gene expression and cognition. Mol Psychiatry.
25:2712–2727. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang X, Zhou X, Li G, Zhang Y, Wu Y and
Song W: Modifications and trafficking of APP in the pathogenesis of
Alzheimer's disease. Front Mol Neurosci. 10:2942017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Herskowitz JH, Feng Y, Mattheyses AL,
Hales CM, Higginbotham LA, Duong DM, Montine TJ, Troncoso JC,
Thambisetty M, Seyfried NT, et al: Pharmacologic inhibition of
ROCK2 suppresses amyloid-β production in an Alzheimer's disease
mouse model. J Neurosci. 33:19086–19098. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang XW, Feng N, Liu YC, Guo Q, Wang JK,
Bai YZ, Ye XM, Yang Z, Yang H, Liu Y, et al: Neuroinflammation
inhibition by small-molecule targeting USP7 noncatalytic domain for
neurodegenerative disease therapy. Sci Adv. 8:eabo07892022.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jia L, Piña-Crespo J and Li Y: Restoring
Wnt/β-catenin signaling is a promising therapeutic strategy for
Alzheimer's disease. Mol Brain. 12:1042019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
McPhie DL, Coopersmith R, Hines-Peralta A,
Chen Y, Ivins KJ, Manly SP, Kozlowski MR, Neve KA and Neve RL: DNA
synthesis and neuronal apoptosis caused by familial Alzheimer
disease mutants of the amyloid precursor protein are mediated by
the p21 activated kinase PAK3. J Neurosci. 23:6914–6927. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Park D, Choi EK, Cho TH, Joo SS and Kim
YB: Human neural stem cells encoding ChAT gene restore cognitive
function via acetylcholine synthesis, Aβ elimination, and
neuroregeneration in APPswe/PS1dE9 mice. Int J Mol Sci.
21:39582020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang H, Zhang L, Zhou D, Li H and Xu Y:
ErbB4 mediates amyloid β-induced neurotoxicity through JNK/tau
pathway activation: Implications for Alzheimer's disease. J Comp
Neurol. 529:3497–3512. 2021. View Article : Google Scholar : PubMed/NCBI
|