Introduction
Medulloblastoma (MB) is the most common malignant
brain tumor in childhood and is thought to arise from precursor
cells in the cerebellar granule cell lineage (1). MB patients are now divided into
stratification groups according to age, degree of resection and
disease dissemination, and are treated depending upon risk.
Although the use of multidisciplinary approaches and stratification
management of the disease have improved prognosis, 50% of patients,
particularly in the high-risk group, experience disease recurrence,
dissemination to the cerebrospinal fluid space, and/or a high
incidence of sequelae (2).
The concept regarding the existence of cancer stem
cells or cancer initiating cells (CICs) is currently a focal point.
The hypothesis that cancerous cells originate from rare populations
of CICs that are more tumorigenic than other cancer cells has
gained increasing credence (3).
CICs are thought to persist in tumors as a distinct population that
can cause tumor recurrence and distant metastasis. The existence of
CICs in MB has also been reported (4,5).
Prominin-1 antigen (CD133) was identified in hematopoietic stem
cells (6,7) and neuroepithelial stem cells (8) and has generally been used as a marker
for CICs (9). Although some
investigators assert that CD133 is not an adequate marker of CICs
since both CD133-positive and -negative cells are able to initiate
tumors (10), it is also true that
CIC-like cells that exhibit self-renewal and multipotential
properties are restricted in the CD133-positive cell fractions. In
the present study, CD133-positive cells were isolated from the
human Daoy MB cell line using magnetic-activated cell sorting
(MACS) beads and the transcript profiles of CD133-positive Daoy MB
cells were investigated using DNA microarray analysis in order to
obtain a better understanding of the molecular properties of CICs
involved in MB tumorigenesis.
Materials and methods
Cell culture
The human Daoy medulloblastoma (MB) cell line was
purchased from the American Type Culture Collection (ATCC) and
cultured in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich, Inc., St. Louis, MO, USA) with 10% fetal bovine
serum (FBS; Sigma-Aldrich, Inc.), 100 U/ml penicillin, and 100
μg/ml streptomycin at 37°C under 5% CO2.
Flow cytometry
Cells were detached in phosphate-buffered saline
(PBS) containing 0.25% trypsin and 0.02% EDTA for 3 min at 37°C
under 5% CO2, and the reaction was stopped by adding
complete medium (DMEM with 10% FBS). Following centrifugation at
1000 rpm for 5 min, the cells were washed and resuspended in bovine
serum albumin (BSA)/PBS buffer (PBS with 0.1% BSA and 2 mM EDTA).
Half of the cells were incubated with FcR blocking reagent
(Miltenyi Biotec Inc., Auburn, CA, USA) and anti-CD133-PE (Miltenyi
Biotec Inc.) for 10 min at 4°C, and the remaining cells were
incubated with IgG-PE (BD Biosciences, San Jose, CA, USA) as
controls. After washing, the cells were resuspended in BSA/PBS
buffer and analyzed using the Beckman Coulter Epics XL system
(Beckman Coulter, Inc., Chaska, MN, USA). The data were analyzed
using FlowJo software (Tree Star Inc., Ashland, OR, USA).
Cell sorting
CD133-positive Daoy cells were sorted using the
CD133 cell isolation kit (Miltenyi Biotec Inc.). Briefly, cells
were suspended in BSA/PBS buffer, incubated with FcR blocking
reagent and CD133 microbeads (Miltenyi Biotec Inc.) for 30 min at
4°C. To determine the sorting efficiency, the cells were incubated
with anti-CD133/2-PE for 10 min. Following washing and
centrifugation, the cells were resuspended in BSA/PBS buffer,
loaded onto a magnetic separation column (Miltenyi Biotec Inc.) and
placed in a magnetic cell separator. The column was rinsed, and the
magnetically labeled cells were flushed out with elution buffer and
collected. These cells were used in the subsequent experiments.
DNA microarray analysis
Total RNAs were isolated from the CD133-positive
Daoy cells (sorted and control) using TRIzol™ (Invitrogen,
Carlsbad, CA, USA). Synthesis and labeling of cRNAs and
hybridization of biotin-labeled cRNA probes to the Human Genome
U133A 2.0 expression Chip arrays (Affymetrix, Santa Clara, CA, USA)
were performed according to the manufacturer's protocol. The
imaging screens were scanned and analyzed using the Affymetrix
Microarray Suite and GeneSpring GX (Agilent Technologies, Santa
Clara, CA, USA).
Semi-quantitative reverse-transcriptase
polymerase chain reaction (RT-PCR) analyses
Total RNAs were prepared and used as templates for
cDNA synthesis with random hexa-nucleotide primers and SuperScript
reverse transcriptase II (Invitrogen). Real-time PCR analyses were
performed using a QuantiTect SYBR-Green PCR kit (Takara, Kyoto,
Japan) and a LightCycler System (Roche, Basel, Switzerland). The
PCR primer sequences were determined using WWW primer tool, Primer3
(http://biotools.umassmed.edu/bioapps/primer3_www.cgi)
(Table I). The transcript abundance
of the genes of interest was normalized to that of
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA as an
internal standard. At least 3 independent analyses were performed
for each sample and for each gene.
 | Table IRT-PCR primer sequences of the genes
of interest. |
Table I
RT-PCR primer sequences of the genes
of interest.
| Gene symbol | Genebank | Sense | Antisense |
|---|
| NRG1 | NM013959 |
TTGGTGCTGCTTTCTTGTTG |
CGGAGCCTCACACACCTATT |
| CCND1 | BC000076 |
TCCTCTCCAAAATGCCAGAG |
TGAGGCGGTAGTAGGACAGG |
| CDK6 | NM001259 |
AGGGTGCAGTCAAAACAACC |
TCCCATCCACTTCAAAGGAG |
| VEGF | AF091352 |
TGCAGATTATGCGGATCAAA |
GCGAGTCTGTGTTTTTGCAG |
| INHBA | M1343 |
AGACGCTGCACTTCGAGATT |
CCCTTTAAGCCCACTTCCTC |
| JAG1 | U73936 |
AGCTGGCTTACACTGGCAAT |
AAGTGGGAGCTCAAAGACCA |
| MYC | NM002467 |
CTCCTGGCAAAAGGTCAGAG |
TCGGTTGTTGCTGATCTGTC |
| HES1 | NM005524.2 |
CTCTCTTCCCTCCGGACTCT |
AGGCGCAATCCAATATGAAC |
| PML | AF230411 |
GCAGCAGTGAGTCCAGTGA |
GCTCTGCCTGCACTTCTTT |
| NFASC | NM015090.2 |
TGCCTTGCTTTTGAGGAGAT |
GGCTGTGGTCAGGGAAACTA |
| APOE | NM000041 |
CCAATCACAGGCAGGAAGAT |
AGCGCAGGTAATCCCAAAAG |
| ASTN | AB0006627 |
ACAACACCCTCCTGGATCTG |
AAGGAGTCCATTGCACCAAC |
| BMP2 | NM001200 |
GGAGAATGCCCTTTTCCTCT |
ACAACCCTCCACAACCATGT |
| NEFL | NM006158.2 |
TCTGTTTGCTTGCAGAGTGG |
GCTAACCACCGAAGGTTCAA |
| MAP2 | U89330 |
AAGAAGGTCGCCATCATACG |
GGCGGATGTTCTTCAGAGAG |
| GAPDH | |
TGCACCACCAACTGCTTAG |
GAGGCAGGGATGATGTTC |
Results
CD133-positive Daoy MB cells were highly
enriched by MACS
Flow cytometry showed that 3–5% of Daoy cells
expressed prominin-1 antigen (CD133) and CD133/2 antigens. After
MACS was applied, the CD133-positive cells were highly enriched
(>60%). These cells were then used in the DNA microarray gene
expression analyses.
Transcript analysis in CD133-positive MB
cells
Transcript analysis using DNA microarrays was
performed, and the acquired data were filtered according to the
gene expression level. In comparison with the control Daoy cells,
the CD133-positive cell-enriched fractions exhibited a >2-fold
increase in the expression of 398 genes, and a <50% decrease in
the expression of 318 genes. A number of molecules involved in the
growth signaling pathways, which play important roles both in MB
oncogenesis and stem cell proliferation, were up-regulated in the
CD133-positive cell-enriched fractions. These molecules included
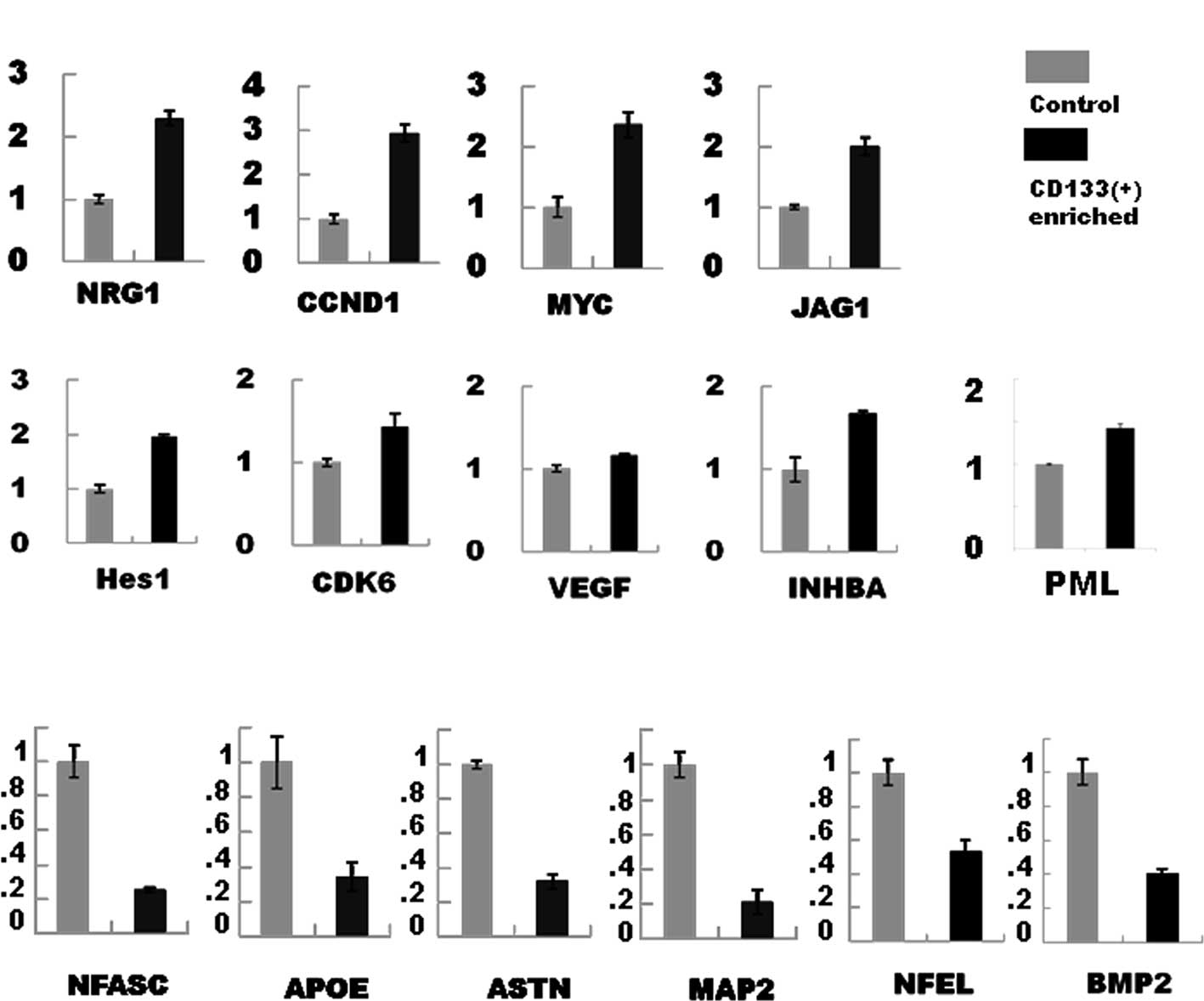
neuregulin-1 (NRG1; which showed a 6.818-fold increase), cyclin D1
(CCND1; 5.636), cyclin-dependent kinase 6 (CDK6; 3.564), vascular
endothelial growth factor (VEGF; 3.186), inhibin β A (INHBA;
3.115), Jagged 1 (JAG1; 2.702), promyelocytic leukemia gene (PML;
2.538), MYC (2.479), and hairy enhancer of split-1 (HES1; 2.078)
(Table II). On the other hand,
neural differentiation markers or developmentally regulated genes,
expressed in the granule cell lineage, such as neurofascin (NFASC;
0.0608), apolipoprotein E (APOE; 0.296), astrotactin (ASTN; 0.392),
neurofilament light polypeptide 68 kDa (NEFL; 0.418), and
microtubule-associated protein 2 (MAP2; 0.49) were down-regulated
(Table II). Semi-quantitative
RT-PCR analyses were then performed in the selected genes
(up-regulated genes, Fig. 1A;
down-regulated genes, Fig. 1B) and
the gene expression changes were confirmed to be significant.
| Table IIGene changes in the CD133-positive
Daoy cells. |
Table II
Gene changes in the CD133-positive
Daoy cells.
| Symbol | Genebank | Map | Fold change | Gene name |
|---|
| Up-regulated
genes |
| RGS16 | U94829 | 1q25-q31 | 10.87 | Regulator of
G-protein signaling 16 |
| NRG1 | NM_013959 | 8p21-p12 | 6.818 | Neuregulin 1; a
ligand for the NEU/ERBB2 |
| CCND1 |
BC000076 | 11q13 | 5.636 | Cyclin
D1 |
| JUN | BC002646 | 1p32-p31 | 5.161 | V-jun sarcoma virus
17 oncogene homolog (avian) |
| CASP2 | BC002427 | 7q34-q35 | 5.081 | Caspase 2,
apoptosis-related cysteine peptidase |
| EGR1 | NM_001964 | 5q31.1 | 3.963 | Early growth
response 1 |
| MET | AA005141 | 7q31 | 3.886 | Met proto-oncogene
(hepatocyte growth factor receptor) |
| Cep290 | AF317887 | 12q21.33 | 3.883 | Centrosome protein
cep290 |
| CDK6 |
NM_001259 |
7q21-q22 | 3.564 | Cyclin-dependent
kinase 6 |
| MAX | NM_002382 | 14q23 | 3.531 | MYC associated
factor X |
| DKK1 | NM_012242 | 10q11.2 | 3.277 | Dickkopf homolog 1
(Xenopus laevis) |
| VEGF |
AF091352 | 6p12 | 3.186 | Vascular
endothelial growth factor |
| INHBA | M13436 |
7p15-p13 | 3.115 | Inhibin, β A
(activin A, activin AB α polypeptide) |
| PYGO1 | AL049925 | 15q21.1 | 3.104 | Pygopus homolog 1
(Drosophila) |
| JAG1 | U73936 |
20p12.1-p11.23 | 2.702 | Jagged 1
(Alagille syndrome) |
| HDAC9 | NM_014707 | 7p21.1 | 2.651 | Histone deacetylase
9 |
| KHSRP | AI933301 | 19p13.3 | 2.643 | KH-type splicing
regulatory protein (FUSE binding protein 2) |
| NPAT | U58852 | 11q22-q23 | 2.564 | Nuclear protein,
ataxia-telangiectasia locus |
| GADD45B | NM_015675 | 19p13.3 | 2.563 | Growth arrest and
DNA-damage-inducible, β |
| PML |
AF230411 | 15q22 | 2.538 | Promyelocytic
leukemia |
| GREM1 | NM_013372 | 15q13-q15 | 2.508 | Gremlin 1, cysteine
knot superfamily, homolog |
| MYC |
NM_002467 |
8q24.12-q24.13 | 2.479 | |
| CCNT1 | NM_001240 | 12pter-qter | 2.41 | Cyclin T1 |
| TGFBR1 | NM_004612 | 9q22 | 2.408 | Transforming growth
factor, β receptor I |
| EGFR | U95089 | 7p12 | 2.299 | Epidermal growth
factor receptor |
| CCNE2 | AF112857 | 8q22.1 | 2.162 | Cyclin E2 |
| SMAD5 | AF010601 | 5q31 | 2.125 | SMAD, mothers
against DPP homolog 5 (Drosophila) |
| HES1 |
BE973687 |
3q28-q29 | 2.078 | Hairy and
enhancer of split 1 (Drosophila) |
| SMAD3 | NM_005902 | 15q21-q22 | 2.016 | SMAD, mothers
against DPP homolog 3 (Drosophila) |
| Down-regulated
genes |
| NFASC |
AI821777 | – | 0.0608 | Neurofascin
homolog (chicken) |
| IGF1 | AI972496 | 12q22-q23 | 0.166 | Insulin-like growth
factor 1 (somatomedin C) |
| IGFBP5 | AW007532 | 2q33-q36 | 0.175 | Insulin-like growth
factor binding protein 5 |
| SEMA3E | NM_012431 | 7q21.11 | 0.197 | Semaphorin 3E |
| APOE |
NM_000041 | 19q13.2 | 0.296 | Apolipoprotein
E |
| BBP | AA012917 | 1p32.1 | 0.356 | TM2 domain
containing 1 |
| VCAM1 | NM_001078 | 1p32-p31 | 0.359 | Vascular cell
adhesion molecule 1 |
| SLIT3 | AB011538 | 5q35 | 0.328 | Slit homolog 3
(Drosophila) |
| RARRES2 | BC000069 | 7q36.1 | 0.365 | Retinoic acid
receptor responder (tazarotene induced) 2 |
| ASTN |
AB006627 | 1q25.2 | 0.392 |
Astrotactin |
| BMP2 |
AA583044 | 20p12 | 0.397 | Bone
morphogenetic protein 2 |
| TNC | BF434846 | 9q33 | 0.404 | Tenascin C
(hexabrachion) |
| UNC5B | AA127885 | 10q22.2 | 0.406 | Unc-5 homolog B
(C. elegans) |
| NEFL |
AL537457 | 8p21 | 0.418 | Neurofilament,
light polypeptide 68 kDa |
| RBP1 | NM_002899 | 3q23 | 0.448 | Retinol binding
protein 1, cellular |
| CDH11 | AU144378 | – | 0.459 | Cadherin 11, type
2, OB-cadherin (osteoblast) |
| RAI16 | NM_022749 | 8p21.3 | 0.461 | Retinoic acid
induced 16 |
| CASP4 | AL050391 | 11q22.2-q22.3 | 0.48 | Caspase 4,
apoptosis-related cysteine peptidase |
| MAP2 | U89330 |
2q34-q35 | 0.49 |
Microtubule-associated protein
2 |
Discussion
In the present study, we first isolated
CD133-positive cells in the human Daoy medulloblastoma (MB) cell
line. The percentage of CD133-positive cells was approximately
3–5%, which was in accordance with previous studies (0.5–10%)
(11,12). After MACS was applied, the
percentage of CD133-positive cells was noted to be greater than
60%. These enriched cell fractions were subsequently subjected to
transcript analysis using DNA microarrays.
Transcript analysis using DNA microarrays identified
various molecules that were components of the growth signaling
pathways, which play important roles both in MB oncogenesis and
stem cell proliferation. The genes which exhibited up-regulated
expression included the activator of MAP kinase signal (RGS16), a
ligand of ERBB (EGF signal component; NRG1), Wnt signal targets
CCND1 and c-myc, a ligand of Notch signal (JAG1), and its target
(HES1) (13–15) (Table
II). c-myc is known to play a key role in stem cell
self-renewal and was used to produce induced pluripotent stem cells
(16). The Wnt and Notch pathways
are involved in the maintenance of stem cell properties and in MB
oncogenesis (17–19). The remaining up-regulated genes
included INHBA, an inhibitor of differentiation factors, such as
activin and TGF β, and VEGF which plays a role in the
neovascularization of tumors (20,21).
These genes may be involved in tumor recurrence or distant
metastasis.
In contrast, the genes whose expression decreased to
less than 50% included the neural markers (MAP2 and NEFL) (22,23),
developmentally regulated genes in the cerebellar granule cell
lineage (NFASC, UNC5B, ASTAN, SLIT3 and APOE) (24–27),
and molecules involved in retinoic acid-induced apoptosis in
neuroblastoma (RBP1, BMP2, RARRES2 and CASP4) (28,29).
Down-regulation of these genes may result in the inhibition of
differentiation and maintenance of undifferentiated properties of
CICs or may contribute to the inhibition of cell death, thereby
providing infertility to CICs.
An understanding of the molecular pathway involved
in MB oncogenesis has been advanced by analyses of the Turcot- and
Gorlin-inherited syndromes which are associated with the
development of MB. The Wnt and sonic hedgehog (SHH) signal pathways
are involved in MB oncogenesis in the Turcot and Gorlin syndromes,
respectively (30–33). In addition, the Notch, epidermal
growth factor receptor ERBB, and platelet-derived growth factor
(PDGF) signaling pathways are involved in MB oncogenesis or
prognosis (33–37). These pathways play crucial roles in
the proliferation and/or differentiation of the cerebellar granule
cell lineage where MB originates. Furthermore, molecular studies
have shown that developmentally regulated signals, such as Wnt, SHH
and Notch, play important roles in self-renewal, proliferation
and/or the multipotency of stem cells, and are also involved in MB
oncogenesis (17–19). These molecular studies and the
results of the present study indicate that further understanding of
the molecular properties and fundamental signaling pathways of CICs
involved in MB oncogenesis may lead to the development of new, more
effective, and less toxic treatment modalities for MB, thereby
improving the quality of life of children with MB.
Acknowledgements
We express sincere appreciation to Professor Y.
Koide, Department of Microbiology and Immunology, Hamamatsu
University School of Medicine, Professor T. Nagata, Dr S. Seto and
Dr M. Uchijima and other members of Professor Koide's Laboratory
for helpful advice, technical assistance and valuable discussions.
This study was supported by a fund from the Japanese Ministry of
Education, Culture, Sports, Science and Technology (no.
17501509).
References
|
1
|
Louis DN, Ohgaki H, Wiestler OD and
Cavenee WK: WHO Classification of Tumours of the Central Nervous
System. 4th edition. International Agency for Research on Cancer;
Lyon: 2007
|
|
2
|
Crawford JR, MacDonald TJ and Packer RJ:
Medulloblastoma in childhood: new biological advances. Lancet
Neurol. 6:1073–1085. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clarke MF and Fuller M: Stem cells and
cancer: two faces of eve. Cell. 124:1111–1115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hemmati HD, Nakano I, Lazareff JA, et al:
Cancerous stem cells can arise from pediatric brain tumors. Proc
Natl Acad Sci USA. 100:15178–15183. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miraglia S, Godfrey W, Yin AH, et al: A
novel five-transmembrane hematopoietic stem cell antigen:
isolation, characterization, and molecular cloning. Blood.
90:5013–5021. 1997.PubMed/NCBI
|
|
7
|
Yu Y, Flint A, Dvorin EL and Bischoff J:
AC133–2, a novel isoform of human AC133 stem cell antigen. J Biol
Chem. 277:20711–20716. 2002.
|
|
8
|
Fargeas CA, Corbeil D and Huttner WB:
AC133 antigen, CD133, prominin-1, prominin-2, etc.: prominin family
gene products in need of a rational nomenclature. Stem Cells.
21:506–508. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mizrak D, Brittan M and Alison MR: CD133:
molecule of the moment. J Pathol. 214:3–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shmelkov SV, Butler JM, Hooper AT, et al:
CD133 expression is not restricted to stem cells, and both
CD133+ and CD133− metastatic colon cancer
cells initiate tumors. J Clin Invest. 118:2111–2120.
2008.PubMed/NCBI
|
|
11
|
Blazek ER, Foutch JL and Maki G: Daoy
medulloblastoma cells that express CD133 are radioresistant
relative to CD133− cells, and the CD133+
sector is enlarged by hypoxia. Int J Radiat Oncol Biol Phys.
67:1–5. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srivastava VK and Nalbantoglu J: Flow
cytometric characterization of the DAOY medulloblastoma cell line
for the cancer stem-like phenotype. Cytometry A. 73:940–948. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Buckbinder L, Velasco-Miguel S, Chen Y, et
al: The p53 tumor suppressor targets a novel regulator of G protein
signaling. Proc Natl Acad Sci USA. 94:7868–7872. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guarnaccia C, Pintar A and Pongor S: Exon
6 of human Jagged-1 encodes an autonomously folding unit. FEBS
Lett. 574:156–160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gilbertson RJ, Clifford SC, MacMeekin W,
et al: Expression of the ErbB-neuregulin signaling network during
human cerebellar development: implications for the biology of
medulloblastoma. Cancer Res. 58:3932–3941. 1998.
|
|
16
|
Takahashi K, Tanabe K, Ohnuki M, et al:
Induction of pluri-potent stem cells from adult human fibroblasts
by defined factors. Cell. 131:861–872. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reya T and Clevers H: Wnt signalling in
stem cells and cancer. Nature. 434:843–850. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ruiz i Altaba A, Sanchez P and Dahmane N:
Gli and hedgehog in cancer: tumours, embryos and stem cells. Nat
Rev Cancer. 2:361–372. 2002.PubMed/NCBI
|
|
19
|
Gilbertson RJ: Medulloblastoma: signalling
a change in treatment. Lancet Oncol. 5:209–218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brown CW, Houston-Hawkins DE, Woodruff TK
and Matzuk MM: Insertion of Inhbb into the Inhba locus rescues the
Inhba-null phenotype and reveals new activin functions. Nat Genet.
25:453–457. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holash J, Maisonpierre PC, Compton D, et
al: Vessel cooption, regression, and growth in tumors mediated by
angiopoietins and VEGF. Science. 284:1994–1998. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garner CC, Tucker RP and Matus A:
Selective localization of messenger RNA for cytoskeletal protein
MAP2 in dendrites. Nature. 336:674–677. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Julien JP, Grosveld F, Yazdanbaksh K,
Flavell D, Meijer D and Mushynski W: The structure of a human
neurofilament gene (NF-L): a unique exon-intron organization in the
intermediate filament gene family. Biochim Biophys Acta. 909:10–20.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ango F, di Cristo G, Higashiyama H,
Bennett V, Wu P and Huang ZJ: Ankyrin-based subcellular gradient of
neurofascin, an immunoglobulin family protein, directs GABAergic
innervation at purkinje axon initial segment. Cell. 119:257–272.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wingate RJ: The rhombic lip and early
cerebellar development. Curr Opin Neurobiol. 11:82–88. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan W, Zhou L, Chen JH, Wu JY, Rao Y and
Ornitz DM: The mouse SLIT family: secreted ligands for ROBO
expressed in patterns that suggest a role in morphogenesis and axon
guidance. Dev Biol. 212:290–306. 1999. View Article : Google Scholar
|
|
27
|
Lafarga M, Crespo P, Berciano MT, Andres
MA and Leon J: Apolipoprotein E expression in the cerebellum of
normal and hypercholesterolemic rabbits. Brain Res Mol Brain Res.
21:115–123. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hallahan AR, Pritchard JI, Chandraratna
RA, et al: BMP-2 mediates retinoid-induced apoptosis in
medulloblastoma cells through a paracrine effect. Nat Med.
9:1033–1038. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gumireddy K, Sutton LN, Phillips PC and
Reddy CD: All-transretinoic acid-induced apoptosis in human
medulloblastoma: activation of caspase-3/poly(ADP-ribose)
polymerase 1 pathway. Clin Cancer Res. 9:4052–4059. 2003.PubMed/NCBI
|
|
30
|
Goodrich LV, Milenkovic L, Higgins KM and
Scott MP: Altered neural cell fates and medulloblastoma in mouse
patched mutants. Science. 277:1109–1113. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wechsler-Reya R and Scott MP: The
developmental biology of brain tumors. Annu Rev Neurosci.
24:385–428. 2001. View Article : Google Scholar
|
|
32
|
Yokota N, Nishizawa S, Ohta S, et al: Role
of Wnt pathway in medulloblastoma oncogenesis. Int J Cancer.
101:198–201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yokota N, Mainprize TG, Taylor MD, et al:
Identification of differentially expressed and developmentally
regulated genes in medulloblastoma using suppression subtraction
hybridization. Oncogene. 23:3444–3453. 2004. View Article : Google Scholar
|
|
34
|
Fan X, Matsui W, Khaki L, et al: Notch
pathway inhibition depletes stem-like cells and blocks engraftment
in embryonal brain tumors. Cancer Res. 66:7445–7452. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gilbertson R, Wickramasinghe C, Hernan R,
et al: Clinical and molecular stratification of disease risk in
medulloblastoma. Br J Cancer. 85:705–712. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
MacDonald TJ, Brown KM, LaFleur B, et al:
Expression profiling of medulloblastoma: PDGFRA and the RAS/MAPK
pathway as therapeutic targets for metastatic disease. Nat Genet.
29:143–152. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gilbertson RJ and Clifford SC: PDGFRB is
overexpressed in metastatic medulloblastoma. Nat Genet. 35:197–198.
2003. View Article : Google Scholar : PubMed/NCBI
|