Introduction
Lung cancer is currently the most common type of
cancer worldwide in terms of incidence and mortality, and non-small
cell lung cancer (NSCLC) accounts for 85% of all lung cancer cases
(1). There remains a lack of
effective biomarkers or indicators for diagnosis, which often leads
to delayed diagnosis and thus the majority of patients with NSCLC
present with an advanced stage. Therefore, the development of novel
techniques for early diagnosis of NSCLC is required.
MicroRNAs (miRNAs) are a class of small noncoding
RNAs that are ∼19–24 nucleotides in length, and they regulate
∼20–30% of the genes in the human genome (2). The coding sequences of miRNAs are
located in the regions between genes or in the introns (3). Previous studies have demonstrated that
miRNAs are involved in a variety of biological processes, including
cell proliferation, differentiation, apoptosis and the development
and differentiation of tissues and organs (4,5); and
pathological processes, including tumorigenesis and tumor
progression (6). Such regulation is
achieved primarily by their partial recombination with the
3′-untranslated region (3′-UTR) of the target mRNA (7,8).
A previous study revealed that molecular markers of
cancer stem-like cells were connected with malignancies by using
gene microarray and sequencing analysis (9). Another study demonstrated that miRNAs
are critical in the carcinogenesis of lung cancer (10). miR-203 was first identified as a
keratinocyte-specific miRNA in the skin, but it is also expressed
in the squamous epithelium of the cervix and esophagus (11). miR-203 regulates embryonic epidermal
differentiation and has been implicated in skin diseases, but it
also serves as a tumor suppressor or oncogene by regulating
proliferation, differentiation, invasion, metastasis and apoptosis
in certain types of human cancer (12,13).
B-cell-specific moloney murine leukemia virus insertion site 1
(Bmi1) is a member of the polycomb repressive complex 1 (PRC1) and
is highly expressed in several types of cancer, including lung
neoplasm (14,15). Yu et al (9) demonstrated that miR-203 inhibited the
proliferation of esophageal cancer cells by suppressing Bmi1
directly. However, the level of miR-203 expression and its role in
NSCLC remains unclear.
In the present study, the interaction between
miR-203 and Bmi1 expression levels was investigated, in addition to
the mechanistic role of miR-203 in NSCLC.
Materials and methods
Patient sample collection
A total of 21 paired NSCLC samples were obtained
from the First Affiliated Hospital of Soochow University (Suzhou,
China) between January 2013 and April 2014. All patients provided
written informed consent prior to tissue donation for research
purposes. The present study was approved by the ethics committee of
Soochow University. All tissues were frozen in liquid nitrogen
immediately following the operation and stored at −70°C until
required.
Cell culture
The human NSCLC cell lines A549, H1299, H226, H1650,
H460 and LTEP-α-2 were obtained from the Cell bank of the Chinese
Academy of Sciences (Shanghai, China). The human bronchial
epithelial (HBE) cell line was obtained from Bogoo Biological
Technology Co. Ltd. (Shanghai, China). Cells were cultured in
RPMI-1640 medium containing 10% fetal bovine serum (FBS; Gibco Life
Technologies, Carlsbad, CA, USA). All the cells were cultured in
humidified sterile conditions containing 5% CO2 at
37°C.
Construction of the luciferase
reporter plasmids, transfection and dual-luciferase assay
The psiCHECK-2 dual luciferase vector (Promega
Corporation, Madison, WI, USA) was used to construct the plasmid
containing the 3′-untranslated region (3′-UTR) of Bmi1. The
fragments containing the predicted wild and mutant sites were
directly synthesized (Genewiz, Inc., Suzhou, China) and then
subcloned into the psiCHECK-2 vector following digestion with
XhoI and NotI restriction enzymes (Thermo Fisher
Scientific, Inc., Pittsburgh, PA, USA) to generate the
Bmi1-3′-UTR-wild and Bmi1-3′-UTR-mutant vectors. Subsequently,
LTEP-α-2 cells (1×105/well) were seeded into a 24-well
plate and co-transfected with 50 ng/well Bmi1-3′-UTR-wild or
Bmi1-3′-UTR-mutant vector and 50 nM/well miR-203 mimics (5′-UUG UGC
UUG AUC UAA CCA UGU-3′) or scrambled microRNA negative control
(miR-NC; 5-UUC UCC GAA CGU GUC ACG UTT-3′). Following culture for
48 h, the LTEP-α-2 cells were collected and the luciferase
activities were measured by the Dual-Luciferase Reporter Assay kit
(Promega Corporation) on a TD20/20 Luminometer (Turner Designs,
Westport, MA, USA). Each experiment was performed in triplicate.
The results were expressed as relative Renilla luciferase
activities, which were obtained following normalization to firefly
luciferase activities. All the transient transfections were
performed using Lipofectamine 2000 (Invitrogen Life Technologies,
Carlsbad, CA, USA).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
The total RNA from cell lines and NSCLC tissues was
extracted using TRIzol reagent (Invitrogen Life Technologies) and
measured on a NanoDrop (ND-2000 spectrophotometer; Thermo Fisher
Scientific, Inc., Wilmington, DE, USA) according to the
manufacturer's protocol. The synthesis of cDNA was performed using
an M-MLV First Strand kit (Invitrogen Life Technologies). The RT
primers for mature miR-203 and U6 were designed as RT stem-loop
primers ((Table I)). Quantitative PCR
(qPCR) was performed using a SYBR Green PCR kit (Takara
Biotechnology Co., Ltd., Dalian, China) and an ABI7500 Real-Time
system (Applied Biosystems Life Technologies, Foster City, CA, USA)
were used to quantify the expression levels of RNA. U6 small
nuclear RNA (snRNA) and GAPDH mRNA were used as endogenous controls
to normalize miR-203 and Bmi1 expression levels. The primer
sequences for miR-203, U6, Bmi1 and GAPDH detection are listed in
(Table I). The relative expression
levels were calculated using the ∆∆Ct method.
 | Table I.Primers for RT or amplification of the
mature miR-203, U6, Bmi1 and GAPDH mRNA. |
Table I.
Primers for RT or amplification of the
mature miR-203, U6, Bmi1 and GAPDH mRNA.
| Name | Sequence, 5′-3′ |
|---|
| RT primers |
|
| U6 |
CGAGCACAGAATCGCTTCACGAATTTGCGTGTCAT |
|
miR-2031 |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTAGTGGTC |
| RT-qPCR primers |
|
| U6 | F,
CGAGCACAGAATCGCTTCA; and R, CTCGCTTCGGCAGCACATAT |
|
miR-203 | F,
GTATCCAGTGCAGGGTCCGA; and R, CGACGGTGAAATGTTTAG |
| Bmi1 | F,
GTGCTTTGTGGAGGGTACTTCAT; and R, TTGGACATCACAAATAGGACAATACTT |
|
GAPDH | F,
GAAGGTGAAGGTCGGAGTC; and R, GAAGATGGTGATGGGATTTC |
Western blot analysis
The cells were lysed in RIPA buffer (Cell Signaling
Technology, Inc., Danvers, MA, USA) with protease inhibitor
(Sigma-Aldrich, St. Louis, MO, USA) at 72 h post-transfection. The
total proteins were separated by 10% SDS-PAGE (Sangon Biotech, Co.,
Ltd., Shanghai, China) and run at a constant voltage of 110 V for 2
h, and subsequently transferred onto polyvinylidene difluoride
membranes (Bio-Rad Laboratories, Inc., Hercules, California, USA).
The membranes were blocked using 1% bovine serum albumin
(Sigma-Aldrich) for 30 min and incubated with the primary antibody
Bmi1 (monoclonal, rabbit anti-human; dilution, 1:3000; catalog no.
5856S; Cell Signaling Technology, Danvers, MA, USA) or GAPDH
(polyclonal, rabbit anti-human; dilution, 1:5000; catalog no.
AP0063; Bioworld Technology, Minneapolis, MN, USA) with agitation
overnight at 4°C. Following 3 washes with Tris-buffered saline and
Tween-20 (TBST), the membranes were incubated with secondary
antibodies (goat anti-rabbit, dilution, 1:3000; catalog no.
sc-2004; Santa Cruz Biotechnology, Dallas, TX, USA) at room
temperature for 2 h and then the washes were repeated. The result
was visualized using an ECL detection system (Pierce Biotechnology,
Inc., Rockford, IL, USA). The Bmi1 protein expression levels were
normalized against the GAPDH protein expression levels and the
density was quantified using Quantity One software, version 4.6
(Bio-Rad Laboratories, Inc.).
Cell proliferation assay
When the cells were in the logarithmic growth phase,
they were seeded into 96-well plates at a density of
5×103 cells/well. Cell proliferation was detected using
the Cell Counting kit-8 (CCK8) and 5-ethynyl-2′-deoxyuridine (EdU)
assays. A CCK8 (Beyotime Institute of Biotechnology, Shanghai,
China) was used following the manufacturer's instructions. To
estimate the number of viable cells, the optical density at a
wavelength of 450 nm (OD450) was measured daily over 4 consecutive
days. The EdU assay (Guangzhou RiboBio, Co., Ltd., Guangzhou,
China) was used to label cells undergoing DNA replication. At 48 h
post-transfection, the cells were cultured with EdU reagent
(Guangzhou RiboBio Co., Ltd.) for 2 h to identify those cells in
the S-phase of the cell cycle. The EdU medium mixture was then
discarded and the cells were fixed with 4% paraformaldehyde
(Sigma-Aldrich) for 30 min at room temperature. Following washes
with glycine (2 mg/ml; Sigma-Aldrich) and phosphate-buffered saline
for 5 min each, 0.5% Triton X-100 (Sigma-Aldrich) was added to each
well. Apollo dyeing reaction buffer (Guangzhou RiboBio, Co., Ltd.)
was then added to each well and the plate was shaken in the dark
for 30 min at room temperature. The DNA was stained by Hoechst
33342 (Guangzhou RiboBio, Co., Ltd.) dye for 30 min and the
proportion of nucleated cells coalescent with EdU were observed
using fluorescence microscopy (Olympus IX71; Olympus, Tokyo,
Japan).
Apoptosis assay
The cells were collected following 48 h of culture
post-transfection with miR-203 and the cell density was adjusted to
1×106cells/ml. The cells were then stained using an
Annexin V-FITC Apoptosis Detection kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. The level
of apoptosis was assessed by flow cytometry (FC500; Beckman
Coulter, Miami, FL, USA).
Transwell invasion assay
At 24 h post-transfection with miR-203 or miR-NC,
the cells were harvested. For the invasion assay, the cells were
resuspended in RPMI-1640 medium containing 1% FBS and then seeded
at a density of 4×104 cells/well into the inserts of the
Transwell chamber (Corning Incorporated, Corning, NY, USA), which
were coated with Matrigel (27.2 µg/µl; BD Biosciences, Franklin
Lakes, NJ, USA). The inserts were subsequently cultured in wells
with 20% FBS-containing medium and the plates were incubated at
37°C for 36 h. Next, the inserts were removed and a cotton tip was
used to scrape off the non-invading cells on the upper surface.
Cells on the lower surface were then fixed with 4% paraformaldehyde
for 30 min at room temperature, and stained with 0.1% crystal
violet (Sigma-Aldrich) for a further 30 min. Following fixation, 3
randomly selected images per well were acquired using an inverted
microscope (Olympus CKX41; Olympus, Tokyo, Japan) and the average
counts were calculated.
Statistical analysis
The results were analyzed using GraphPad Prism
Software, version 5.01 (GraphPad Software Inc., La Jolla, CA, USA).
All data are presented as the mean ± standard deviation from 3
independent experiments. A 2-tailed Student's t-test was used for
the comparison of the mean between groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
Upregulation of Bmi1 and
downregulation of miR-203 in NSCLC
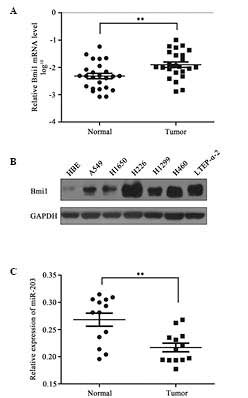
To determine whether the expression of Bmi1 was
increased in NSCLC, the Bmi1 mRNA expression in 25 paired NSCLC
tissues and adjacent non-tumor tissues was quantified by RT-qPCR.
This analysis revealed that the mRNA expression level of Bmi1 was
significantly increased in NSCLC tissues compared with paired
non-tumor tissues (P<0.01; Fig.
1A). In addition, Bmi1 protein levels were measured in 7
different cell lines. Compared with HBE cells, Bmi1 protein
expression levels were significantly increased in the NSCLC cell
lines, including the A549, H1650, H226, H1299, H460 and LTEP-α-2
cell lines (Fig. 1B). However,
miR-203 was significantly downregulated in the 13 NSCLC samples
compared with paired non-tumor tissues (Fig. 1C).
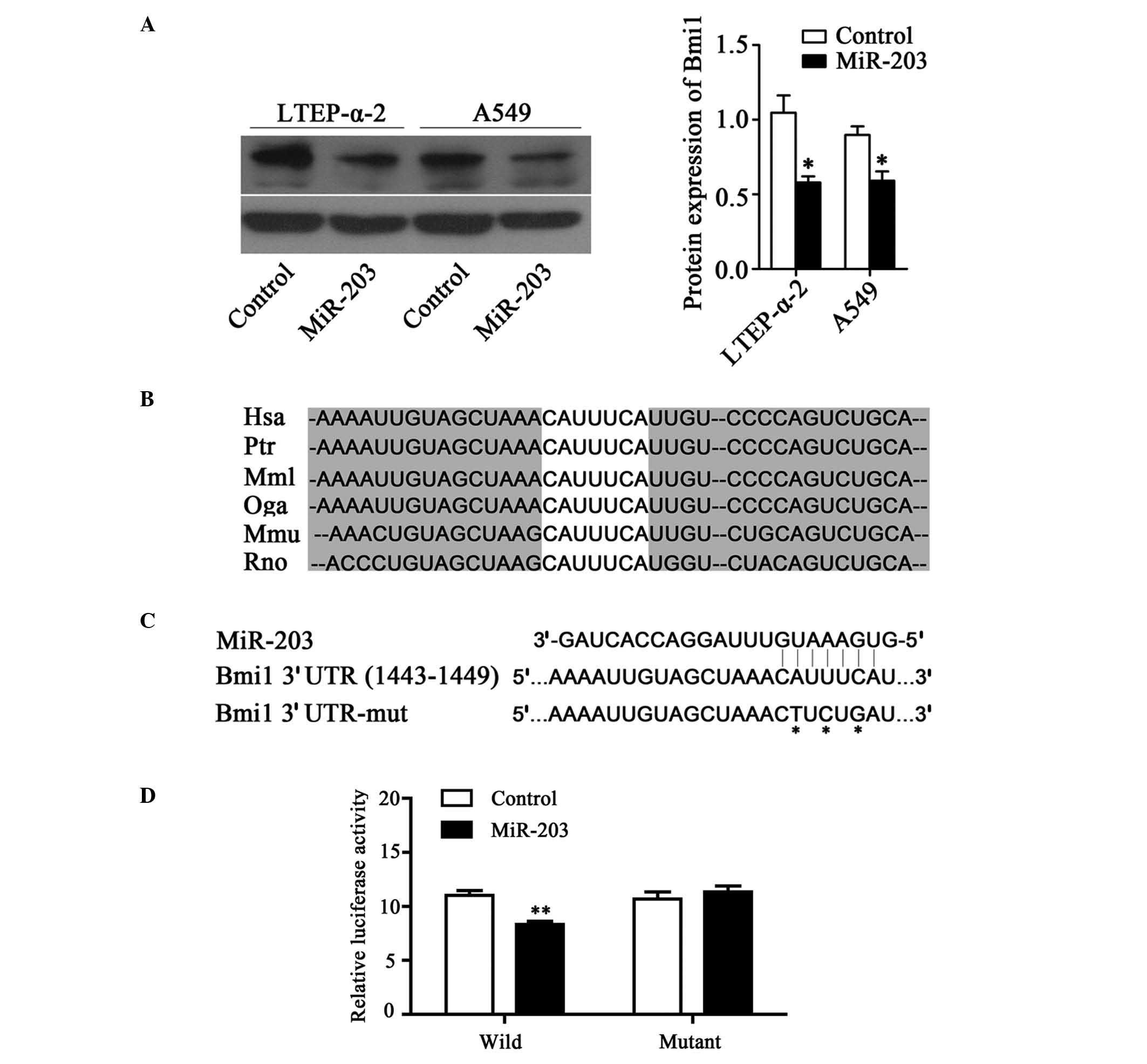
Bmi1 is a functional target of miR-203
in NSCLC
Bmi1 is a functional target of miR-203 in NSCLC.
Using TargetScan software, the potential targets of miR-203
including SNAI2, PKCα, Bmi1, PLD2, LASP1 and c-myc were identified.
To evaluate whether miR-203 regulates Bmi1 expression, miR-203
mimics were transiently transfected into A549 and LTEP-α-2 cells.
The results demonstrated that the overexpression of miR-203
markedly reduced Bmi1 protein expression levels in LTEP-α-2 and
A549 cells (P<0.05; Fig. 2A). In
accordance with these findings, it is possible that miR-203
directly targets Bmi1 to inhibit its expression. To investigate
this hypothesis, potential targets of miR-203 were identified using
TargetScan software (www.targetscan.org) and a highly conserved predicted
target site on the 3′UTR of Bmi1 mRNA was identified (Fig. 2B). This region of the Bmi1 3′UTR,
including the predicted miR-203 binding site, was then cloned into
the psiCHECK-2 vector (Fig. 2C). The
results of the dual-luciferase assay demonstrated that in LTEP-α-2
cells co-transfected with the Bmi1-3′-UTR-wild vector, miR-203
significantly suppressed the luciferase activity compared with the
control group. However, this result was not observed with the
Bmi1-3′-UTR-mutant vector. These results indicate that miR-203
binds to the 3′-UTR of Bmi1 mRNA and thus regulates its expression
(Fig. 2D).
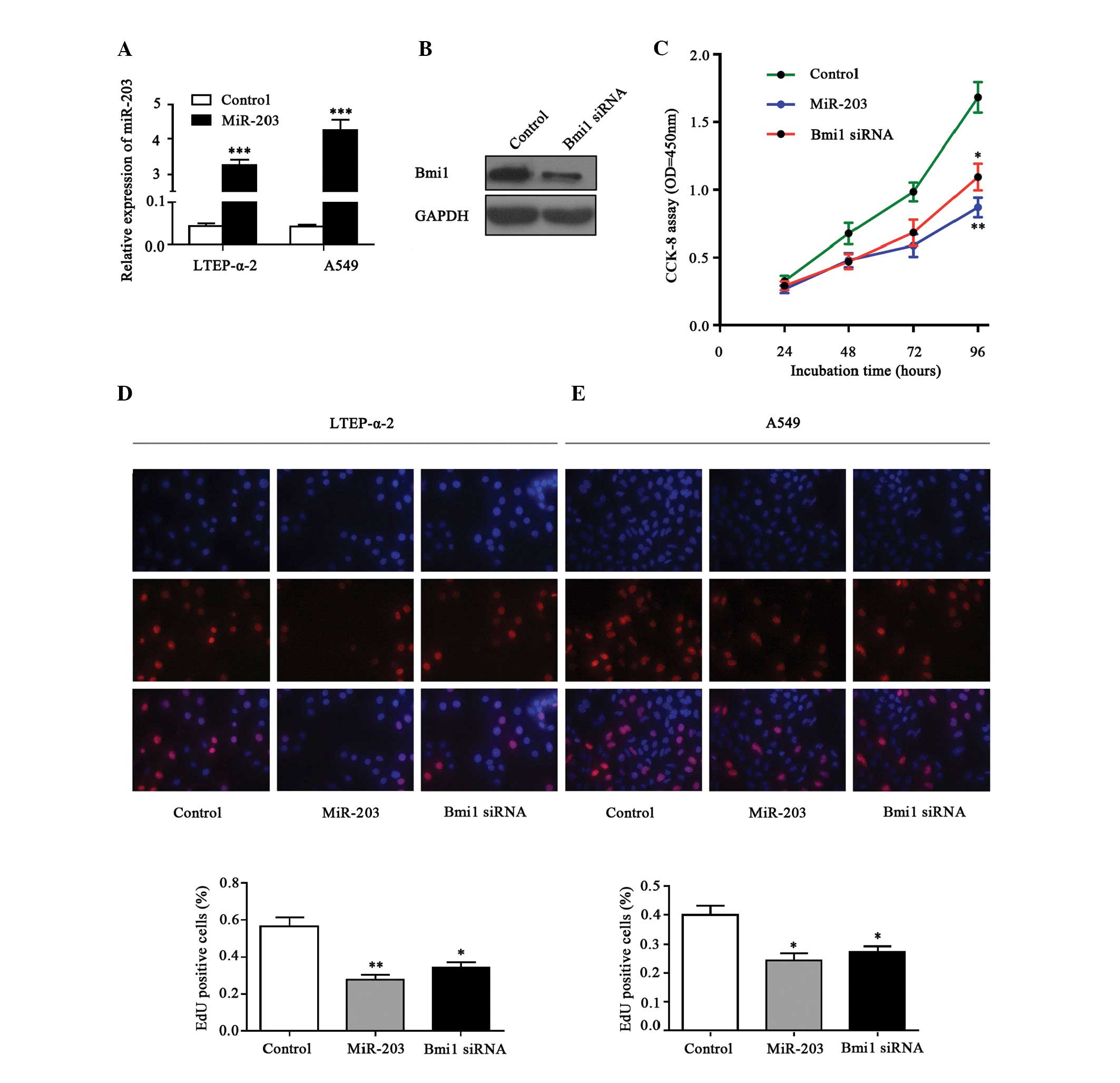
miR-203 inhibits cell proliferation in
NSCLC cells
The effects of miR-203 expression on malignant
phenotypes of NSCLC cells were observed. The efficiency of
transfection was determined by RT-qPCR, demonstrating increased
expression levels of miR-203 in the mimics-transfected cells
compared with the control cells (P<0.01; Fig. 3A). In addition, the protein level of
Bmi1 was successfully suppressed by Bmi1-siRNA, as demonstrated by
western blot analysis (Fig. 3B). The
results of the CCK8 assays demonstrated that ectopic miR-203
expression significantly inhibited LTEP-α-2 cell growth compared
with the control group (P<0.01; Fig.
3C). A similar result was observed following the suppression of
Bmi1 expression using Bmi1-siRNA (P<0.05; Fig. 3C). The EdU incorporation assay also
indicated that miR-203 inhibited NSCLC cell proliferation. Ectopic
expression of miR-203 significantly reduced EdU incorporation in
LTEP-α-2 (P<0.01; Fig. 3D) and
A549 (P<0.05; Fig. 3E) cells,
which is in accordance with the reduction in EdU incorporation
induced by attenuated expression of Bmi1 with specific siRNA in
LTEP-α-2 (P<0.05; Fig. 3D) and
A549 (P<0.05; Fig. 3E) cells.
These results indicate that forced expression of miR-203 resulted
in significant inhibition of cell growth in NSCLC cells in
vitro.
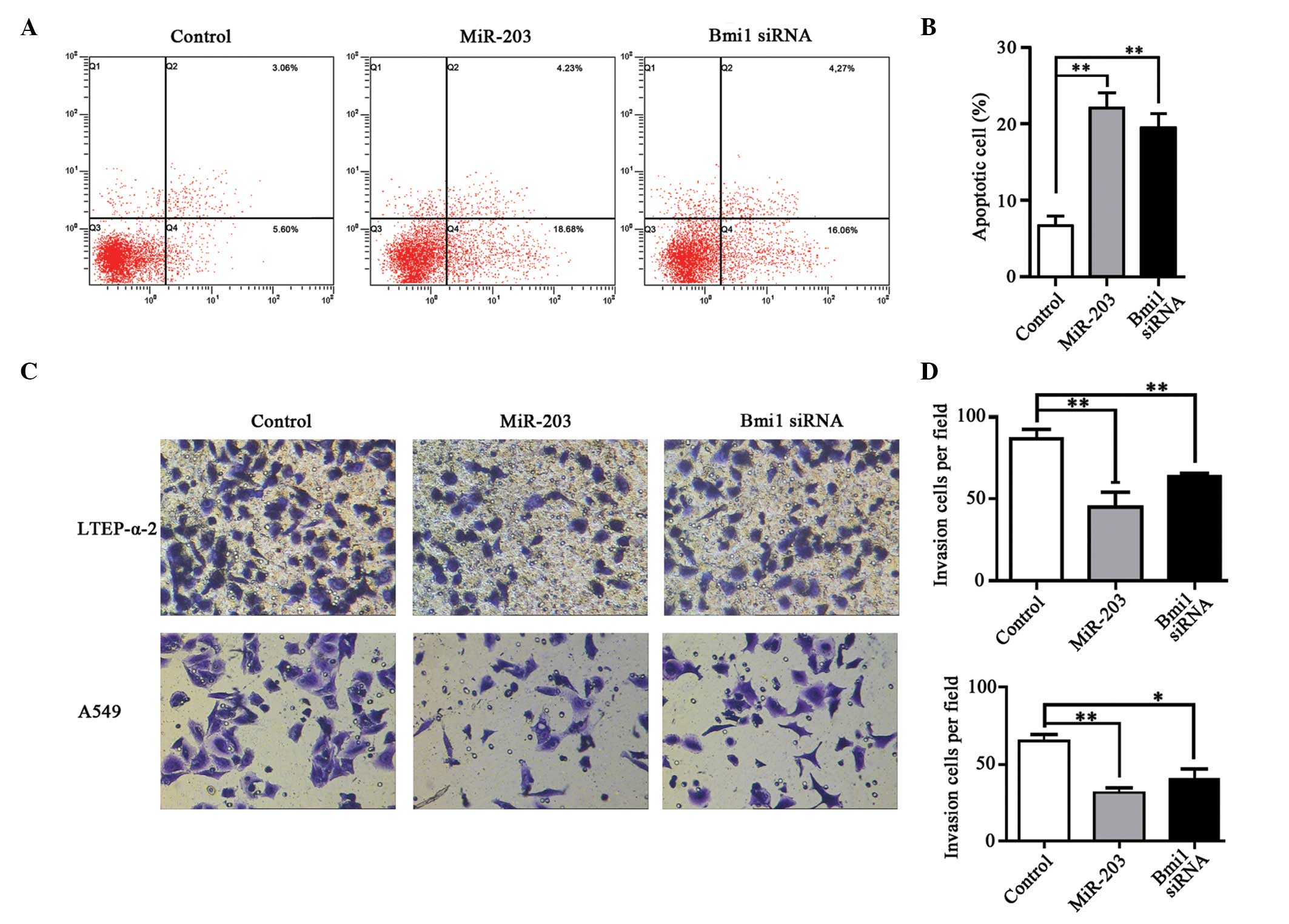
Restoration of miR-203 induces
apoptosis in NSCLC cells
An apoptosis assay was performed in LTEP-α-2 cells
with increased miR-203 expression or silenced Bmi1 (Fig. 4A). The percentage of apoptotic cells
in the miR-203 transfection group was significantly increased
compared with the control group (21.7 vs. 8.7%, P<0.01; Fig. 4B). Similar to the results obtained
from the miR-203 transfection, comparable results were obtained
from downregulation of Bmi1, in which the percentage of apoptotic
cells was also significantly increased compared with the control
group (20.3 vs. 8.7%, P<0.01; Fig.
4B). The flow cytometry results demonstrated that ectopic
expression of miR-203 and downregulation of Bmi1 by siRNA induced
cell apoptosis.
Upregulation of miR-203 inhibited
metastatic ability of NSCLC cells
To demonstrate whether miR-203 expression affects
the metastatic capability of NSCLC cells, a Transwell invasion
assay was performed (Fig. 4C). The
quantity of LTEP-α-2 and A549 cells that invaded the filter in the
miR-203 transfected group was markedly reduced compared with the
control group in the two cell types (P<0.01; Fig. 4D). Similarly, cell invasion was
significantly reduced following transfection of Bmi1-specific siRNA
(P<0.01 and P<0.05 in LTEP-α-2 and A549 cells, respectively;
Fig. 4D).
Discussion
MicroRNAs are a class of small noncoding RNAs that
serve important functions in the regulation of the biological
processes of cells, including differentiation, proliferation and
apoptosis (16). Previous studies
have demonstrated that miRNA dysregulation occurs in various types
of human cancer and these aberrantly expressed miRNAs are
considered to serve pivotal roles in carcinogenesis (17–19).
Whether miRNAs act as oncogenes or tumor suppressor genes depends
on the context. For example, miR-203 behaves as a tumor suppressor
and is downregulated in pancreatic, glioma and esophageal cancer,
while it is upregulated in epithelial ovarian cancer and acts as an
oncogene (9,20–22).
The present study demonstrated that miR-203 was
downregulated in NSCLC specimens, which is in agreement with a
previous study (23). The inhibitory
effect of miR-203 expression on the proliferation and invasion of
NSCLC cells that was observed in the present study indicates that
miR-203 may be involved in the tumorigenesis of lung cancer. Future
studies should aim to investigate whether miR-203 expression
affects cell cycle progression and can induce cell apoptosis in
NSCLC.
A previous study demonstrated that downregulated
miR-203 levels were significantly associated with lymph node
metastasis in laryngeal squamous cell carcinoma (24). However, to the best of our knowledge,
no studies have yet compared the association between miR-203 and
lymph node metastasis in lung cancer. The ability of tumor cells to
migrate and invade is associated with their cell adhesion
properties (25,26). The results of the present study
indicate that the restoration of miR-203 expression may serve an
important role in the progress of NSCLC treatment through
inhibiting cell invasion, though further in vivo studies are
required to confirm this.
To understand the mechanisms by which miR-203
suppresses cell proliferation, TargetScan software was used to
predict the target genes of miR-203. SNAI2, PKCα, Bmi1, PLD2, LASP1
and c-myc were identified in the present study as potential
targets. A proportion of these targets have already been
demonstrated to be directly regulated by miRNAs, for example SNAI2
expression is reported to be repressed by miR-203 in prostate cells
(27). SNAI2 is a transcription
factor that belongs to the E-box-motif family. It inhibits
apoptosis by repressing p53-mediated transcription and promotes
epithelial-mesenchymal transition by directly repressing E-cadherin
(28,29). However, it is not clear whether there
is an interaction between miR-203 and Bmi1 in NSCLC. Bmi1, a
polycomb gene family member, is expressed in almost all tissues and
is upregulated in various types of human cancer indicating that
Bmi1 may be involved in tumor progression (14). A previous study demonstrated that Bmi1
is a direct target of miR-128 in glioma and that it serves an
important function in glioma proliferation and self-renewal
(30). Another study demonstrated
that Bmi1 was overexpressed in lung cancer (15), which is in accordance with the results
of the present study. The present study also demonstrated Bmi1
protein expression levels were significantly increased in the NSCLC
cell lines, A549, H1299, H226, H1650, H460 and LTEP-α-2, compared
with healthy HBE cells.
To assess whether miR-203 directly represses Bmi1
expression, western blot analysis and a luciferase assay were
performed. These analyses corroborated that miR-203 specifically
downregulated endogenous Bmi1 expression at the
post-transcriptional level.
There are numerous genes predicted to be targets of
miR-203, and Bmi1 was the only one confirmed by the present study.
There may be other potential targets of miR-203 that function in
the tumorigenesis of lung cancer, such as c-myc, therefore
further studies focusing on these novel target genes are required.
In addition, the sample size in the present study was small, and
further studies from different specimens of NSCLC patients are
required to strengthen the findings of the present study.
In conclusion, the present study demonstrated that
miR-203 represses Bmi1 protein expression by directly targeting the
Bmi1 3′UTR, which may reduce the proliferative and invasive
abilities of NSCLC cells. These findings indicate that miR-203 may
be a novel target for the diagnosis and therapy of NSCLC.
Acknowledgements
The authors would like to thank the patients for
their participation and cooperation and Dr Hong-Tao Zhang for his
helpful criticism. The present study was supported in part by the
grants from the Jiangsu Province Key Provincial Talents Program
(grant no. RC2011106 to Dr Jun Zhao), the Provincial Talents of Six
Summits Program (the eighth, grant no. 2011-WS-056 to Dr Jun Zhao),
the Natural Science Foundation of Jiangsu Province (grant no.
BK20131159 to Dr Jun Zhao), Suzhou city Municipal Youth Fund of
Science and Education (grant no. KJXW2011007 to Dr Chang Li) and
the Suzhou Key Laboratory for Molecular Cancer Genetics (grant no.
SZS201209 to Dr Jun Zhao).
References
|
1
|
Li C, Xu C, Ma H, et al: Video-assisted
thoracoscopic lobectomy with a single utility port is feasible in
the treatment of elderly patients with peripheral lung cancer.
Thorac Cancer. 5:219–224. 2014. View Article : Google Scholar
|
|
2
|
Shi Y, Liu C, Liu X, et al: The microRNA
miR-34a inhibits non-small cell lung cancer (NSCLC) growth and the
CD44hi stem-like NSCLC cells. PLoS One. 9:e900222014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Monteys AM, Spengler RM, Wan J, et al:
Structure and activity of putative intronic miRNA promoters. RNA.
16:495–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kasinski AL and Slack FJ: Epigenetics and
genetics. MicroRNAs en route to the clinic: progress in validating
and targeting microRNAs for cancer therapy. Nat Rev Cancer.
11:849–864. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fang YX and Gao WQ: Roles of microRNAs
during prostatic tumorigenesis and tumor progression. Oncogene.
33:135–147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lagos-Quintana M, Rauhut R, et al:
Identification of novel genes coding for small expressed RNAs.
Science. 294:853–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee RC and Ambros V: An extensive class of
small RNAs in Caenorhabditis elegans. Science. 294:862–864. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu X, Jiang X, Li H, et al: miR-203
inhibits the proliferation and self-renewal of esophageal cancer
stem-like cells by suppressing stem renewal factor Bmi-1. Stem
Cells Dev. 23:576–585. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sonkoly E, Wei T, Janson PC, et al:
MicroRNAs: novel regulators involved in the pathogenesis of
psoriasis? PLoS One. 2:e6102007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu J, Li A, Hong SM, Hruban RH and Goggins
M: MicroRNA alterations of pancreatic intraepithelial neoplasias.
Clin Cancer Res. 18:981–992. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jin J, Deng J, Wang F, et al: The
expression and function of microRNA-203 in lung cancer. Tumour
Biol. 34:349–357. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jiang L, Li J and Song L: Bmi-1, stem
cells and cancer. Acta Biochim Biophys Sin (Shanghai). 41:527–534.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meng X, Wang Y, Zheng X, et al:
shRNA-mediated knockdown of Bmi-1 inhibit lung adenocarcinoma cell
migration and metastasis. Lung Cancer. 77:24–30. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Z, Yao H, Lin S, et al:
Transcriptional and epigenetic regulation of human microRNAs.
Cancer Lett. 331:1–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He L, Thomson JM, Hemann MT, et al: A
microRNA polycistron as a potential human oncogene. Nature.
435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iorio MV, Visone R, Di Leva G, et al:
MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miao L, Xiong X, Lin Y, et al: miR-203
inhibits tumor cell migration and invasion via caveolin-1 in
pancreatic cancer cells. Oncol Lett. 7:658–662. 2014.PubMed/NCBI
|
|
21
|
Dontula R, Dinasarapu A, Chetty C, et al:
MicroRNA 203 modulates glioma cell migration via Robo1/ERK/MMP-9
signaling. Genes Cancer. 4:285–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang S, Zhao X, Wang J, et al:
Upregulation of microRNA-203 is associated with advanced tumor
progression and poor prognosis in epithelial ovarian cancer. Med
Oncol. 30:6812013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang C, Wang X, Liang H, et al: miR-203
inhibits cell proliferation and migration of lung cancer cells by
targeting PKCα. PLoS One. 8:e739852013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian L, Li M, Ge J, et al: MiR-203 is
downregulated in laryngeal squamous cell carcinoma and can suppress
proliferation and induce apoptosis of tumours. Tumour Biol.
35:5953–5963. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Torres VA: Rab'ing tumor cell migration
and invasion: Focal adhesion disassembly driven by Rab5. Cell Adhes
Migr. 8:84–87. 2014. View Article : Google Scholar
|
|
26
|
Li L, Zhao F, Lu J, et al: Notch-1
signaling promotes the malignant features of human breast cancer
through NF-κB activation. PLoS One. 9:e959122014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qu Y, Li WC, Hellem MR, et al: MiR-182 and
miR-203 induce mesenchymal to epithelial transition and
self-sufficiency of growth signals via repressing SNAI2 in prostate
cells. Int J Cancer. 133:544–555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pérez-Caro M, Bermejo-Rodríguez C,
González-Herrero I, et al: Transcriptomal profiling of the cellular
response to DNA damage mediated by Slug (Snai2). Br J Cancer.
98:480–488. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: an alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Godlewski J, Nowicki MO, Bronisz A, et al:
Targeting of the Bmi-1 oncogene/stem cell renewal factor by
microRNA-128 inhibits glioma proliferation and self-renewal. Cancer
Res. 68:9125–9130. 2008. View Article : Google Scholar : PubMed/NCBI
|