Introduction
Renal cell carcinoma (RCC) is a common malignant
neoplasm in the urinary system and the sixth leading cause of
cancer mortality in Western countries (1). It is estimated that 63,920 new cases of
kidney cancer will occur in 2014 in the USA, with 13,860
mortalities due to this disease (1).
Clear cell renal cell carcinoma (CCRCC), accounting for 70% of all
RCC cases, and papillary (pRCC; 10–15% of all RCC cases)
carcinomas, constitute the two most frequent subtypes of RCC
(2). Although clinical management of
RCC has changed in previous years, with increased incidental
diagnosis and initiation of therapy in localized stages, 20–40% of
patients with RCC succumb to cancer progression (3). Due to the high resistance of the tumors
to chemotherapy and radiotherapy (4),
it is important to identify novel approaches that can effectively
inhibit RCC cell growth and metastasis. Molecularly-targeted
therapies have shown potential in achieving these goals. CKLF-like
MARVEL transmembrane domain-containing 5 (CMTM5) is a member of the
CMTM protein family and was originally reported by the Human
Genomics Center of Peking University (Beijing, China) in 2003
(5). CMTM5 is located at 14q11.2, and
has been reported to act as a tumor-suppressor gene (TSG), since it
was specifically downregulated in numerous human cancers, including
cervical carcinoma, ovarian cancer, pancreatic cancer, myeloid
leukemia and prostate cancer (6–11).
However, the role of CMTM5 in human RCC remains unclear. The
present study aimed to detect the expression pattern of CMTM5 and
its association with clinicopathological characteristics in
patients with RCC. The effect of CMTM5 on cell viability and
apoptosis was also explored using an RCC cell line in
vitro.
Materials and methods
Tissue samples and cancer cells
The present study was approved by the Human Study
Ethics Committees of the Ministry of Public Health (the First
Affiliated Hospital of Wenzhou Medical University, Wenzhou, China).
All specimens were handled and made anonymous according to the
ethical and legal standards. A tissue microarray (TMA) of 75
patients with RCC and adjacent normal kidney tissues was purchased
from Shanghai Outdo Biotech Co., Ltd. (Shanghai, China). The mean
age of the patients with RCC was 58.7 years. Tissue samples were
collected from 75 patients with RCC who underwent radical
nephrectomy at the Department of Urology, the First Affiliated
Hospital of Wenzhou Medical University between September 2006 and
December 2008. None of the patients had received chemotherapy or
radiotherapy prior to surgery. The RCC tissues consisted of 34
stage I, 23 stage II, 14 stage III and 4 stage IV, according to the
World Health Organization criteria from 2004 (12). The clinicopathological features of
this patient cohort are summarized in Table I. The metastatic clear renal cell
adenocarcinoma CAKII, papillary renal cell carcinoma ACHN, the
metastatic renal cell adenocarcinoma OS-RC-2 and the human kidney
HK2 cell lines were purchased from the cell bank of the Chinese
Academy of Sciences (Beijing, China), and were maintained in RPMI
1640 medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA USA) at 37°C with a 5% CO2
humidified atmosphere.
 | Table I.Associations between CMTM5 expression
and various clinicopathological parameters of human renal cell
carcinoma. |
Table I.
Associations between CMTM5 expression
and various clinicopathological parameters of human renal cell
carcinoma.
| Clinicopathological
parameters | Cases, n | CMTM5-low, n (%) | CMTM5-high, n
(%) | P-value |
|---|
| Sex |
|
|
| 0.80 |
| Male | 50 | 32 (42.7) | 18 (24.0) |
|
|
Female | 25 | 17 (22.7) | 8 (10.7) |
|
| Age (years) |
|
|
| 0.32 |
|
<60 | 45 | 27 (36.0) | 18 (24.0) |
|
| ≥60 | 30 | 22 (29.3) | 8 (10.7) |
|
| Clinical stage |
|
|
| 0.58 |
| I–II | 57 | 36 (48.0) | 21 (28) |
|
|
III–IV | 18 | 13 (17.3) | 5 (6.7) |
|
| Histological
grade |
|
|
| 0.63 |
| I–II | 40 | 25 (33.3) | 15 (20.0) |
|
|
III–IV | 35 | 24 (32.0) | 11 (14.7) |
|
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR analysis was performed to detect the
expression of CMTM5 mRNA in RCC and adjacent normal kidney tissues,
as well as in RCC and human kidney cells. Briefly, the 20-µl RT
reaction system (K1622, Fermentas; Thermo Fisher Scientific, Inc.)
containing 1 µg RNA and diethyl pyrocarbonate-treated water was
placed in an RNase-free PCR tube. The components were mixed and
heated to 65°C for 5 min before being transferred to an ice bath.
Then, 4.1 µl 5X reaction buffer, 0.6 µl RiboLock RNase inhibitor
(20 U/µl), 2 µl deoxynucleotide triphosphate mix (10 mM of each
deoxynucleotide triphosphate) and 1.1 µl ReverTraAce were added
(K1622; Fermentas; Thermo Fisher Scientific, Inc.). The mixture was
heated to 72°C for 60 min, followed by 72°C for 10 min. The
complementary DNAs were either stored at −20°C or immediately used
for qPCR, which was performed using the AceQ qPCR SYBR-Green Master
mix, Vazyme Biotech Co., Ltd., China) on the ABI 2720 Thermal
Cycler; Thermo Fisher Scientific, Inc.). The CMTM5 primers used for
qPCR were forward, 5′-TGATGCCTTCAAGATCTACCG-3′ and reverse,
5′-TCCACAGATAAGTCCCCAGTG-3′. As an internal control, β-actin was
used, and its primers were forward, 5′-GGCACTCTTCCAGCCTTCC-3′ and
reverse, 5′-GAGCCGCCGATCCACAC-3′. The thermocycler conditions were
as follows: Step 1: 95°C for 2 min; then Step 2: 60–95°C for 2 min
for 40 cycles; and then Step 3: 72°C for 2 min for 40 cycles. The
2−ΔΔCq method was used to quantify the results (13).
Western blot analysis
Total ACHN cell protein was extracted using
radioimmunoprecipitation protein lysis buffer (Beyotime Institute
of Biotechnology, Shanghai, China) with added 1% protease inhibitor
cocktail and 1 mM phenylmethylsulfonyl fluoride (PMSF). The protein
concentration was measured by BCA Protein Assay kit (Beyotime
Institute of Biotechnology). The 50 µg protein samples were
separated by electrophoresis in 10% SDS-PAGE and then transferred
onto a nitrocellulose membrane (EMD Millipore, Billerica, MA, USA).
The membranes were probed with ananti-CMTM5 antibody (dilution
1:500; kindly provided by Professor Wenling Han, Peking University)
at 4°C overnight, followed by incubation with a horseradish
peroxidase-conjugated secondary antibody (dilution, 1:5,000;
sc2030; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at room
temperature for 1 h. Then, the protein bands were visualized with
SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher
Scientific, Inc.). An anti-a-tubulin antibody (dilution 1:3,000;
sc-73242, Santa Cruz Biotechnology, Inc.) was used as a loading
control.
Cell transfection
Recombinant pLenti6.3-CMTM5-IRES-EGFP lentivirus and
negative control lentivirus Lenti-EGFP were purchased from Shanghai
R&S Biotechnology Co., Ltd. (Shanghai, China). ACHN cells
(1×105 cells/dish) were seeded in a 24-well plate. The
Lenti-EGFP and CMTM5 lentiviruses were infected into ACHN cells
using Polo3000 (Shanghai R&S Biotechnology Co., Ltd.),
according to the manufacturer's protocol. ACHN cells were infected
with lentiviruses (1×109 TU/ml; Shanghai R&S
Biotechnology Co., Ltd.) in the presence of 8 µg/ml Polybrene
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The efficiency of
transfection was confirmed as >70% of cells were expressing
EGFP.
Immunohistochemistry (IHC)
The subcellular localization and expression level of
CMTM5 protein in TMA (tissue thickness, 4 µm) was analyzed by the
streptavidin-biotin complex method. Upon deparaffinization in
xylene and rehydration in a 100-95-75–0% graded ethanol series,
antigens were retrieved in citrate (0.1 M, pH 6.0) for 15 min at
95°C. The TMA samples were incubated with 3% hydrogen peroxide for
20 min at room temperature, washed with PBS, and then incubated
with 3% goat serum (Shanghai Mengry Bio-Technology Co., Ltd.,
Shanghai, China) for 30 min at room temperature to reduce
non-specific binding. Subsequently, the TMA were incubated with a
primary polyclonal rabbit anti-human CMTM5 antibody (dilution
1:1,000; kindly provided by Professor Wenling Han), or with goat
serum instead of anti-CMTM5 antibody as a negative control, at 4°C
overnight, followed by incubation with a horseradish
peroxidase-conjugated secondary antibody (dilution, 1:5,000;
SC2030; Santa Cruz Biotechnology, Inc.) for 30 min at room
temperature. The immunostaining of CMTM5 protein was visualized
with 3,3′-diaminobenzidine (Dako; Agilent Technologies, Inc., Santa
Clara, CA, USA) subsequent to washing with PBS. The IHC results
were evaluated by two independent investigators who were blinded to
the clinical background of the specimens. The immunoreactive score
of CMTM5 protein expression was semi-quantified by staining
intensity and percentage of tumor cell positivity. The percentage
of positive cells was scored as follows: 0 (0%), 1 (<10%), 2
(11–50%), 3 (51–80%) and 4 (>80%). The staining intensity was
scored as follows: Below the level of detection =0; weak expression
=1; moderate expression =2; and intense expression =3. The
percentage of positive cells was multiplied with the staining
intensity to obtain the immunoreactive score (scale, 0–12). Scores
>6 were considered to indicate high expression, while scores
=0–6 were considered to indicate low expression.
Cell viability analysis
The anti-proliferative effects of CMTM5 were
determined by Cell Counting kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) according to the
manufacturer's protocol. Following transfection with
CMTM5-overexpressing lentivirus (CMTM5 OE) and negative control
lentivirus (NC) ACHN cells were seeded in 96-well microplates at a
density of ~3,000 cells per well for 24, 48 and 72 h, and then
incubated at 37°C for another 2 h with CCK-8 (10 µl per well). The
absorption of each well at 450 nm was measured under a microplate
reader.
Cell apoptosis and cell cycle
analyses
Cell apoptosis was measured by flow cytometry using
the Annexin V-FITC/PI Apoptosis Detection kit (BD556547; BD
Biosciences, Franklin Lakes, NJ, USA) according to the
manufacturer's protocol. At 72 h post-transfection, ACHN cells were
harvested and incubated with fluorescein isothiocyanate
(FITC)-conjugated Annexin V for 30 min at room temperature in the
dark. Then samples were analyzed on a FACScan flow cytometer (BD
Biosciences). The cell cycle distribution was analyzed 48 h after
transfection. ACHN cells were washed in PBS, trypsinized, washed
again in PBS and fixed in 70% ethanol at −20°C. Subsequently, 10
µg/ml propidium iodide (PI) was added to each sample, and the cell
cycle was analyzed with a FACScan flow cytometer. The obtained data
were analyzed using CellQuest software (version 7.5.3, BD
Biosciences).
In vitro migration and invasion
assays
Migration and invasion assays were performed using
Transwell chamber inserts (Corning Incorporated, Corning, NY, USA)
without or with Matrigel (for invasion) according to the
manufacturer's protocol. An equal number of ACHN cells infected
with CMTM5 OE and NC (4.0×105) and cells were seeded
onto the inserts. The lower chamber was incubated in RPMI 1640
medium containing 20% FBS. The cells were incubated for 24 h before
the inserts on the upper chamber were removed. Cells that had
migrated to or invaded the lower membrane were stained with 0.1%
crystal violet and counted under a light microscope in five random
fields.
Statistical analysis
A χ2 test was used to analyze the
association between CMTM5 expression and different
clinicopathological characteristics. For the in vitro
studies, the statistical significance of the differences between
groups was determined using one-way analysis of variance. Data were
presented as the mean ± standard error. All tests were two-sided,
and P<0.05 was considered to indicate a statistically
significant difference. SPSS 19.0 package (IBM Corp., Armonk, NY,
USA) was used for all statistical evaluations. All experiments were
performed in triplicate.
Results
Downregulation of CMTM5 mRNA and
protein expression in human RCC tissues
The expression patterns of CMTM5 protein in RCC and
adjacent non-cancerous kidney tissues were examined using a TMA by
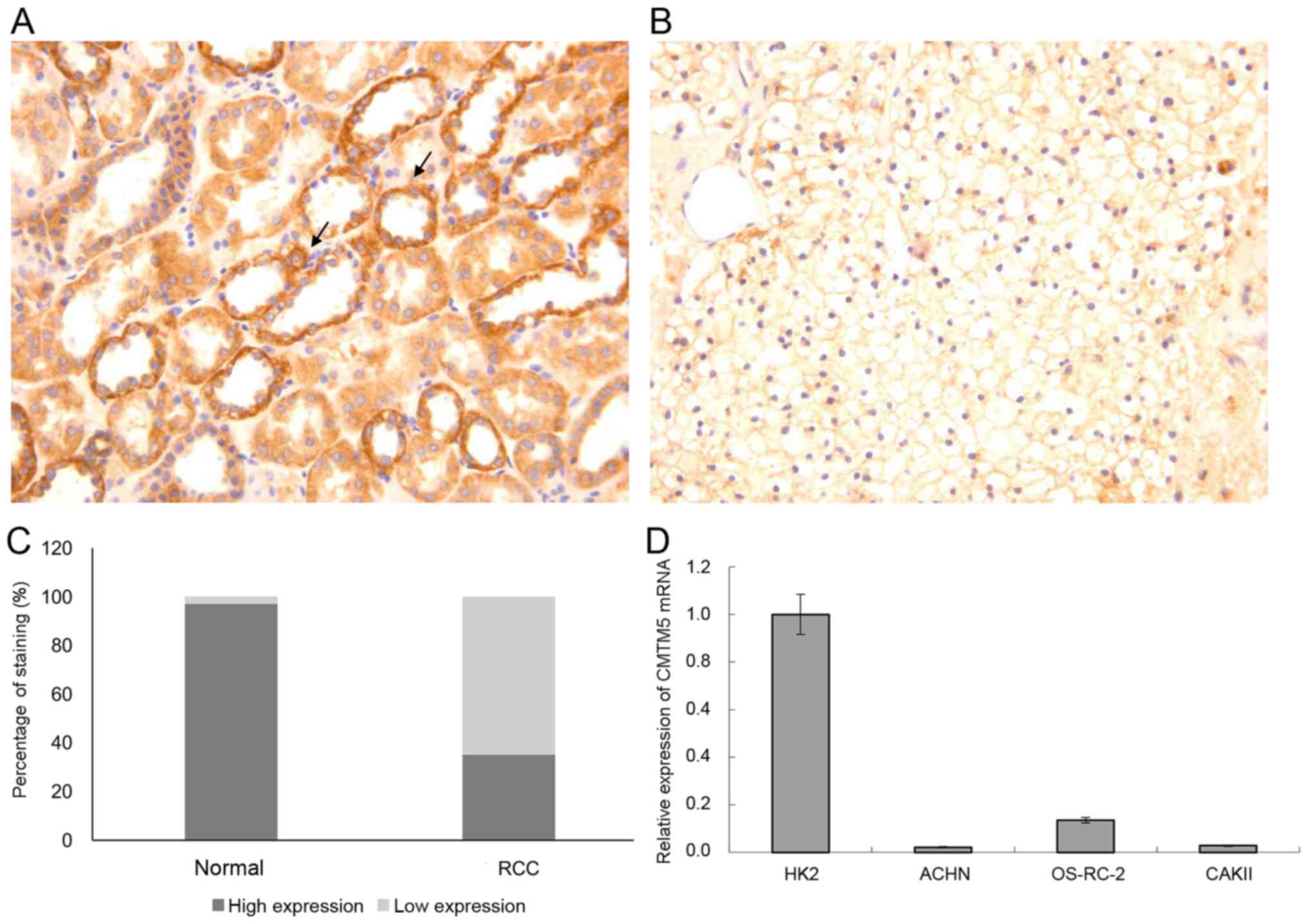
IHC. As shown in Fig. 1A, the
positive immunostaining of CMTM5 protein was mainly localized in
the membrane and cytoplasm of renal epithelial cells, and was
detected in 97% (73/75) of normal kidney tissues. However, CMTM5
protein was either absent or weakly expressed in 65% (48/75) of RCC
tissues (Fig. 1B). There was a
significant difference in the percentage of positive immunostaining
expression of CMTM5 protein between RCC (26/75, 35%) and adjacent
normal kidney tissues (73/75, 97%) (P<0.01; Fig. 1C).
Next, the expression levels of CMTM5 mRNA were
detected in metastatic renal cell adenocarcinoma cell lines ACHN
and CAKII and metastatic clear renal cell adenocarcinoma cell line
OS-RC-2. Compared with that in the normal renal tubular epithelial
cell line HK2, the expression level of CMTM5 mRNA was significantly
downregulated in metastatic renal cell adenocarcinoma cell lines
ACHN and CAKII and metastatic clear renal cell adenocarcinoma cell
line OS-RC-2 (Fig. 1D). The ACHN cell
line was selected for further functional experiments as it
exhibited the lowest expression of CMTM5 mRNA.
Furthermore, no statistically significant
associations were identified between various clinicopathological
features, including sex, age, clinical staging or histological
grade, and the expression of CMTM5 protein (Table I).
CMTM5 suppresses RCC cell
proliferation
The downregulation of CMTM5 in RCC suggested that
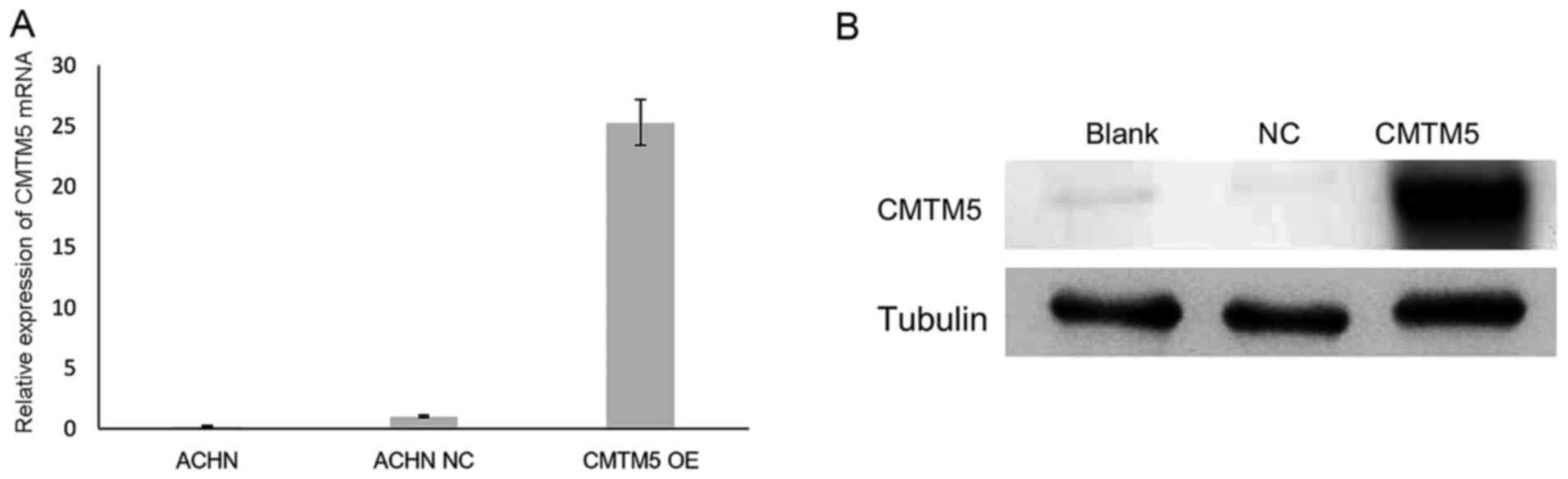
CMTM5 may act as a TSG in this cancer type. To investigate its
anti-proliferative effect, the restoration of CMTM5 mRNA and
protein expression in ACHN cells was confirmed by RT-qPCR and
western blotting, respectively (Fig.
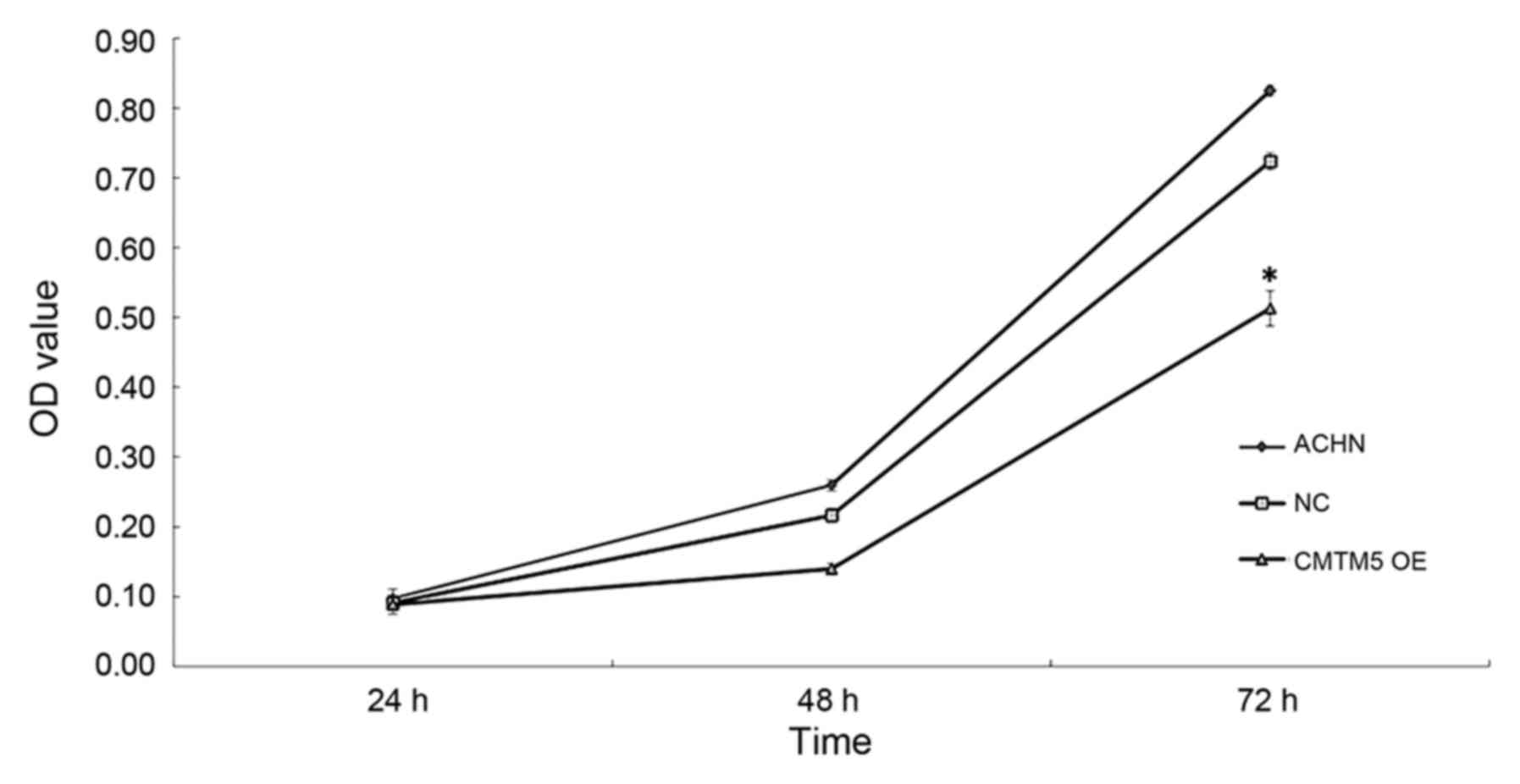
2). In the CCK-8 assay, the viability of ACHN-CMTM5 cells was
decreased significantly compared with that of the untreated and
null-infected controls at 72 h (all P<0.05; Fig. 3).
Restoration of CMTM5 expression
induces cell cycle arrest in G0/G1 phase and apoptosis
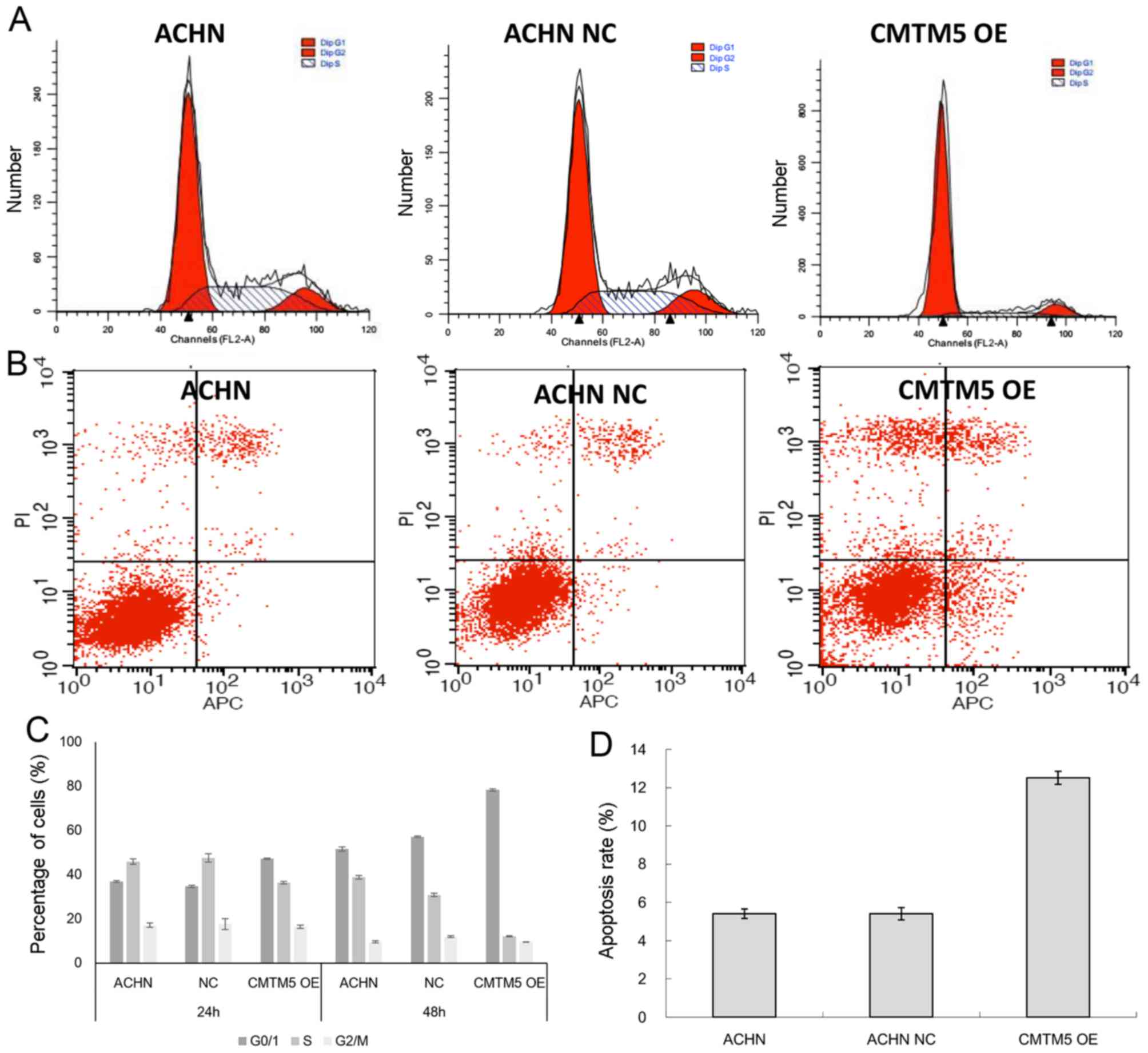
The cell cycle analysis results generated by flow
cytometry revealed that the CMTM5-infected group contained more
cells in G0/G1 phase than the control Lenti-EGFP infected cells
(cell viability index, 42.88 vs. 21.68%, respectively) (P<0.05;
Fig. 4A and C). The apoptotic effect
of CMTM5 in RCC cells was evaluated by Annexin V-FITC/PI staining
assay. Compared with that of control Lenti-EGFP infected cells, the
average apoptosis rate of CMTM5-ACHN cells was significantly higher
at 72 h post-transfection (5.41 vs. 12.53%, respectively; Fig. 4B and D). These results suggest that
the growth inhibition caused by CMTM5 may be mediated by G0/G1
arrest.
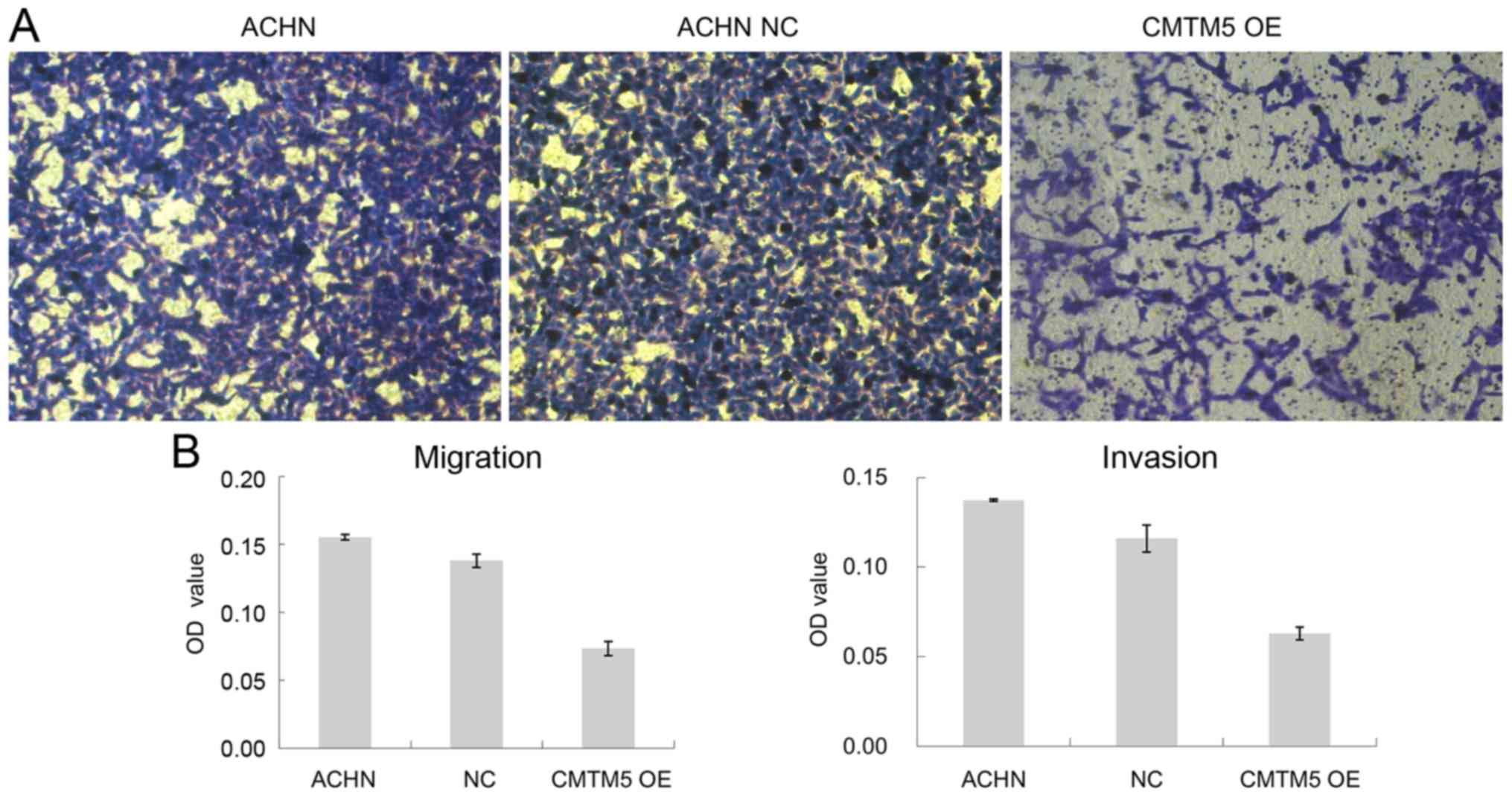
Restoration of CMTM5 expression
reduces migration and invasion of RCC
Cell migration and invasion are both key steps for
tumor development and metastasis. Using a Transwell assay, it was
observed that the restoration of CMTM5 expression reduced the
penetration rate of ACHN cells through the polycarbonate membrane
compared with that of the Lenti-EGFP-infected ACHN cells in both
migration and invasion assays (both P<0.05; Fig. 5).
Discussion
In the present study, downregulation of CMTM5 at the
mRNA and protein level was observed in RCC tissues and cell lines.
Functional analyses demonstrated the tumor-suppressive roles of
CMTM5 in RCC cells in vitro, including inhibition of cell
proliferation, promotion of cell apoptosis, and suppression of cell
migration and invasion.
DNA methylation is one of the most common epigenetic
modifications in the regulation of gene expression (14). CMTM5 inactivated by promoter CpG
methylation has been detected in numerous carcinomas (6,7,15). Our group also investigated whether
CMTM5 was silenced by promoter methylation in RCC by bisulfite
genomic sequencing (data not shown). However, similarly to the case
of CMTM3, no promoter methylation of CMTM5 was observed in RCC cell
lines (16). It has been previously
indicated that the expression of CMTM5 in multiple other types of
tumor was downregulated in RCC mainly by different epigenetic or
genetic mechanisms (6–11).
The evidence of CMTM5 downregulation in RCC cells
prompted us to investigate the biological function of CMTM5 on RCC
cell lines in vitro. Previous studies revealed that CMTM
family proteins, including CMTM3, CMTM4 andCMTM5, act as TSGs that
can inhibit cell growth and induce apoptosis (5,15–18). The present study investigated the role
of CMTM5 on RCC cell growth and apoptosis by restoration of CMTM5
expression in the RCC cell line ACHN. The growth of ACHN-CMTM5
cells was significantly inhibited, and the cell apoptotic rate was
markedly increased, following the transfection of the CMTM5 vector.
In addition, it was observed that the restoration of CMTM5
expression suppressed both the motility and invasiveness of ACHN
cells, which is in line with previous findings in other tumors
(6,7,10,11). In the present study, the ACHN line was
selected for functional studies as the CMTM5 mRNA expression level
was the lowest of the cell lines tested. However, a previous study
by Brodaczewska et al (2) has
indicated that the ACHN cell line is not suitable for studies of
clear cell RCC, which accounts for 70–75% of all RCC cases.
Therefore, further studies in clear cell RCC are required in the
future to further validate the data of the present study.
The molecular mechanisms underlying the
tumor-suppressive effects of CMTM5 on RCC cell growth and migration
remain unclear. As a member of the CMTM family, the CMTM5 protein
has a predicted MARVEL domain, which is a common feature of the
occludin, physin, gyrin, MAL and CMTM families, and is a
membrane-associating domain that may form part of the machinery of
membrane apposition events (19). For
instance, CMTM3, another member of the CMTM family, may inhibit
tumor growth and invasion through promoting the transcription of
p21, downregulating the expression of matrix metalloproteinase 2
and phosphorylating extracellular signal-regulated kinase 1/2
(Erk1/2) (16,20). CMTM5 induces the apoptosis of
pancreatic cancer cells not only by activation of caspases 3, 8 and
9, but also by exerting synergistic effects with tumor necrosis
factor-α (6). CMTM7 can repress
oncogenic epidermal growth factor receptor (EGFR) signaling through
promoting EGFR internalization (21).
CMTM8 is also a regulator of the Erk1/2 signaling pathway (22). Thus, it can be proposed that the
mechanism of action of CMTM5 in RCC cells may be associated with
membrane protein sorting and trafficking, such as EGFR and vascular
endothelial growth factor receptor signaling, which are both
approved targets for the treatment of advanced RCC (23,24).
Further evaluation of this hypothesis will provide additional
information on molecular-targeted therapy for RCC.
In conclusion, the present data suggest that CMTM5
may function as a tumor suppressor in human RCC by suppressing the
viability of RCC cells, implying its potential as a therapeutic
target for this malignancy.
Acknowledgements
The present study was supported by the Wenzhou
Natural Science Foundation (grant no. Y20140579). The authors thank
Professor Wenling Han for generously providing the anti-CMTM5
antibody, as well as the Department of Pathology of The First
Affiliated Hospital of Wenzhou Medical University (Wenzhou, China),
for their support in IHC and staining evaluation.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brodaczewska KK, Szczylik C, Fiedorowicz
M, Porta C and Czarnecka AM: Choosing the right cell line for renal
cell cancer research. Mol Cancer. 15:832016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leibovich BC, Lohse CM, Crispen PL,
Boorjian SA, Thompson RH, Blute ML and Cheville JC: Histological
subtype is an independent predictor of outcome for patients with
renal cell carcinoma. J Urol. 183:1309–1315. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grimm MO, Wolff I, Zastrow S, Frohner M
and Wirth M: Advances in renal cell carcinoma treatment. Ther Adv
Urol. 2:11–17. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao W, Wang J, Li H, Guan W, Xia D, Yu G,
Xiao H, Lang B, Ma X, Liu J, et al: Fibulin-1 is down-regulated
through promoter hypermethylation and suppresses renal cell
carcinoma progression. J Urol. 190:291–301. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo X, Li T, Wang Y, Shao L, Zhang Y, Ma D
and Han W: CMTM5 induces apoptosis of pancreatic cancer cells and
has synergistic effects with TNF-alpha. Biochem Biophys Res Commun.
387:139–142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li P, Liu K, Li L, Yang M, Gao W, Feng J,
Lv Y, Qu X and Kong B: Reduced CMTM5 expression correlates with
carcinogenesis in human epithelial ovarian cancer. Int J Gynecol
Cancer. 21:1248–1255. 2011.PubMed/NCBI
|
|
8
|
Niu J, Li H, Zhang Y, Li J, Xie M, Li L,
Qin X, Qin Y, Guo X, Jiang Q, et al: Aberrant expression of
CKLF-like MARVEL transmembrane member 5 (CMTM5) by promoter
methylation in myeloid leukemia. Leuk Res. 35:771–776. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shao L, Cui Y, Li H, Liu Y, Zhao H, Wang
Y, Zhang Y, Ng KM, Han W, Ma D and Tao Q: Cmtm5 exhibits tumor
suppressor activities and is frequently silenced by methylation in
carcinoma cell lines. Clin Cancer Res. 13:5756–5762. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shao L, Guo X, Plate M, Li T, Wang Y, Ma D
and Han W: CMTM5-v1 induces apoptosis in cervical carcinoma cells.
Biochem Biophys Res Commun. 379:866–871. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xiao Y, Yuan Y, Zhang Y, Li J, Liu Z,
Zhang X, Sheng Z, Xu T and Wang X: CMTM5 is reduced in prostate
cancer and inhibits cancer cell growth in vitro and in vivo. Clin
Transl Oncol. 17:431–437. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
World Health Organization Classification
of Tumours: Pathology and Genetics of Tumours of the Urinary System
and Male Genital OrgansEble JN, Sauter G, Epstein JI and Sesterhenn
IA: IARC Press; Lyon: pp. 72004
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dawson MA and Kouzarides T: Cancer
epigenetics: From mechanism to therapy. Cell. 150:12–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Nan X, Li X, Chen Y, Zhang J and
Sun L: CMTM5 exhibits tumor suppressor activity through promoter
methylation in oral squamous cell carcinoma. Biochem Biophys Res
Commun. 447:304–310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xie J, Yuan Y, Liu Z, Xiao Y, Zhang X, Qin
C, Sheng Z, Xu T and Wang X: CMTM3 is frequently reduced in clear
cell renal cell carcinoma and exhibits tumor suppressor activities.
Clin Transl Oncol. 16:402–409. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su Y, Lin Y, Zhang L, Liu B, Yuan W, Mo X,
Wang X, Li H, Xing X, Cheng X, et al: CMTM3 inhibits cell migration
and invasion and correlates with favorable prognosis in gastric
cancer. Cancer Sci. 105:26–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Plate M, Li T, Wang Y, Mo X, Zhang Y, Ma D
and Han W: Identification and characterization of CMTM4, a novel
gene with inhibitory effects on hela cell growth through inducing
G2/M phase accumulation. Mol Cells. 29:355–361. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sanchez-Pulido L, Martín-Belmonte F,
Valencia A and Alonso MA: Marvel: A conserved domain involved in
membrane apposition events. Trends Biochem Sci. 27:599–601. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Z, Xie J, Wu J, Li W, Nie L, Sun X, Sun
X, Tang A, Li X, Liu R, et al: CMTM3 inhibits human testicular
cancer cell growth through inducing cell-cycle arrest and
apoptosis. PloS One. 9:e889652014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Li J, Su Y, Fan Y, Guo X, Li L, Su
X, Rong R, Ying J, Mo X, et al: A novel 3p22.3 gene CMTM7 represses
oncogenic egfr signaling and inhibits cancer cell growth. Oncogene.
33:3109–3118. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang W, Mendoza MC, Pei X, Ilter D,
Mahoney SJ, Zhang Y, Ma D, Blenis J and Wang Y: Down-regulation of
CMTM8 induces epithelial-to-mesenchymal transition-like changes via
c-MET/extracellular signal-regulated kinase (ERK) signaling. J Biol
Chem. 287:11850–11858. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Clark PE: The role of VHL in clear-cell
renal cell carcinoma and its relation to targeted therapy. Kidney
Int. 76:939–945. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li C, Liu B, Dai Z and Tao Y: Knockdown of
VEGF receptor-1 (VEGFR-1) impairs macrophage infiltration,
angiogenesis and growth of clear cell renal cell carcinoma (crcc).
Cancer Biol Ther. 12:872–380. 2011. View Article : Google Scholar : PubMed/NCBI
|