Introduction
Human hepatocellular carcinoma (HCC) is the third
most common cause of mortality due to cancer and the fifth most
common malignancy worldwide, with a particularly high incidence
observed in Africa and Asia (1).
Liver carcinogenesis is generally considered to be associated with
inflammation caused by environmental factors, including hepatitis B
and C viral infections, which are the major risk factors of HCC
(2,3).
Toll-like receptor 4 (TLR4) is a well-studied member
of the TLR protein family. Lipopolysaccharide (LPS) released from
bacteria can bind to TLR4, leading to the activation of TLR4 and
myeloid differentiation primary response 88 (MyD88). Signaling via
TLR4/MyD88 initiates the activation of nuclear factor-κB (NF-κB)
and a subsequent expression of pro-inflammatory genes that trigger
the host inflammatory response to infection (4,5). TLR4 is
overexpressed in various types of human cancer, including breast
cancer, melanoma, colon cancer, ovary cancer and prostate cancer
(6–10). TLR4 expression and activation of the
TLR4/MyD88-mediated signaling pathways promote carcinogenesis and
metastasis (11,12). Furthermore, it has been demonstrated
that TLR4 contributes to the growth and migration of hepatocellular
tumors (13,14).
An inflammatory microenvironment is a hallmark of
tumor-associated inflammation (15).
Chronic inflammation induces DNA damage and oncogenic mutations,
causing cancer formation and progression (16–18).
Interleukin (IL)-23, a member of the IL-12 family, is a
heterodimeric pro-inflammatory cytokine that is mainly produced by
inflammatory myeloid cells. IL-23 stimulates T cells to
differentiate into T helper 17 (Th17) cells in the presence of IL-6
and transforming growth factor-β (19). Th17 cells are a population of T helper
lymphocytes, and include the subtypes of IL-17A, IL-17B, IL-17C,
IL-17D, IL-17E/IL-25 and IL-17F. These cells are able to produce
another pro-inflammatory cytokine, IL-17 (20). IL-17 recruits neutrophils, enhances
dendritic cell maturation and stimulates macrophages to produce
IL-1β and tumor necrosis factor (TNF)-α, which then induce and
mediate pro-inflammatory responses (21). An increasing number of studies
demonstrated that the IL-23/IL-17A axis contributes to cancer
growth and metastasis. Furthermore, this axis can reduce the
infiltration of CD8+ cells in tumors and enhance the
immunosuppressive activity of regulatory T cells (22,23).
In the present study, the expression of TLR4 in HCC
tissues and its correlation with the clinical features of HCC was
analyzed. In addition, the correlation between TLR4 expression and
the IL-23/IL-17A axis in HCC were examined in order to determine
whether IL-23 and IL-17A serve a role in its carcinogenesis.
Materials and methods
Chemicals
Primers were synthesized by SBS Genetech Co., Ltd.
(Beijing, China). The primary antibodies against TLR4 (dilution
1:400; cat. no. ab13556), IL-23 (dilution 1:400; cat. no. ab45420),
IL-17A (dilution 1:200; cat. no. ab136668) and β-actin (dilution
1:1,000; cat. no. ab8227) were all purchased from Abcam (Cambridge,
UK). ELISA kits for the detection of IL-23 (cat. no. F0153) and
IL-17A (cat. no. F01451) levels were provided by Westang Biological
Technology Co., Ltd. (Shanghai, China). LPS was obtained from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany), while the MyD88
inhibitor ST2825 was from MedChemExpress (Monmouth Junction, NJ,
USA). TRIzol® reagent, fetal bovine serum (FBS),
Dulbecco's modified Eagle's medium (DMEM), and the SYBR-Green PCR
Master Mix, bicinchoninic acid (BCA) and enhanced chemiluminescence
(ECL) kits were purchased from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The reverse transcription (RT) kit was supplied
by Promega Corporation (Madison, WI, USA).
Cell culture
Human liver cancer HepG2 cells were obtained from
the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in DMEM supplemented
with 10% FBS, penicillin (100 U/ml) and streptomycin (100 g/ml) in
a humidified atmosphere containing 5% CO2 at 37°C.
Human tissues
The application of clinical samples was reviewed and
approved by the Ethics Committee of Ningxia Medical University
(Yinchuan, China), and patients provided written informed consent.
Based on the National Standard for Diagnosis and Treatment of
Primary Liver Cancer (2011 edition) guidelines (24), 45 paraffin-embedded tissue samples of
primary HCC from patients who had not received radiotherapy or
chemotherapy prior to surgery were collected at the General
Hospital of Ningxia Medical University between October 2012 and
December 2014. The patients were graded on the basis of the
tumor-node-metastasis (TNM) stage (25). There were 17 adjacent normal tissues
also collected and served as the controls. All the samples were
embedded in paraffin and stored at 4°C after sectioning.
Immunohistochemical assay
Tissue sections (thickness, 4 µm) were
deparaffinized in xylene, soaked in alcohol and treated in antigen
unmasking solution (Beijing Zhongshan Golden Bridge Biotechnology
Co., Ltd., Beijing, China) in the microwave for 10 min, followed by
incubation in 3% hydrogen peroxide for 10 min to inactivate the
endogenous peroxidase. The sections were then incubated with
antibodies against TLR4, IL-23, IL-17A and β-actin at 4°C
overnight. Immunostaining was performed using the
Histostain™-Plus kit (cat. no. 859073; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. Two
independent researchers evaluated each stained section.
Immunohistochemical staining was evaluated by the intensity and
distribution of cytoplasmic staining. Staining intensity was graded
as follows: 0, no staining; 1, weak staining; 2, moderate staining;
and 3, strong staining. Staining distribution was based on the
percentage of stained tumor cells: 0, 0%; 1, <25%; 2, 25–50%; 3,
51–75% and 4, >75% (26). The
final staining scores were calculated as follows: Staining
intensity × the percentage of stained tumor cells.
ELISA
Cells were seeded in 96-well plates at a density of
5×104 cells/well and cultured for 24 h, followed by
exposure to 1 mg/l LPS for 12, 24 or 36 h alone or pretreated with
40 µmol/l ST2825 for 8 h, prior to being treated with 1 mg/l LPS
for 12, 24 or 36 h. The control cells were cultured with DMEM. The
supernatant was centrifuged at 12,000 × g for 5 min at 4°C and the
concentration of IL-23 and IL-17A in the culture medium was
measured by an ELISA kit, according to the manufacturer's
instructions. The absorbance was then detected
spectrophotometrically at 450 nm using a microplate reader and
plotted against a standard curve with standard levels expressed as
pg/ml. Each sample was analyzed in triplicate.
RNA extraction and RT-quantitative
polymerase chain reaction (qPCR)
HepG2 cells were treated with LPS (1 mg/l) for 24 h,
alone or followed by ST2825 (40 µmol/l) treatment for 8 h. The
control cells were cultured with DMEM. Total RNA was extracted from
HepG2 cells by TRIzol reagent, and further purified using RNeasy
Mini columns (cat no. 74104; Qiagen GmbH, Hilden, Germany).
Complementary DNA (cDNA) was then synthesized using a high-capacity
cDNA reverse transcription kit. Subsequently, qPCR assays were
performed in triplicate using SYBR® Green PCR Master Mix
with the following thermal cycling conditions: 95°C for 10 min, 40
cycles of denaturation at 95°C for 15 sec, annealing at 60°C for 30
sec and extension at 72°C for 30 sec. The sequences of primers used
in the qPCR assay are listed in Table
I. The relative mRNA expression of HepG2 cells was calculated
using the 2−ΔΔCq method (27), and normalized using GAPDH as the
reference gene.
 | Table I.Sequences of primers used in
quantitative polymerase chain reaction assay. |
Table I.
Sequences of primers used in
quantitative polymerase chain reaction assay.
| Genes | Primer sequence
(5′-3′) |
|---|
| TLR4 | F:
GCAGTTTCTGAGCAGTCGTG |
|
| R:
CTGTCCTCCCACTCCAGGTA |
| IL-23 | F:
GCCTTCTCTGCTCCCTGATA |
|
| R:
GACTGAGGCTTGGAATCTGC |
| IL-17A | F:
CGATCCACCTCACCTTGGAA |
|
| R:
TAGTCCACGTTCCCATCAGC |
| GAPDH | F:
CAATGACCCCTTCATTGACC |
|
| R:
GACAAGCTTCCCGTTCTCAG |
Western blot analysis
HepG2 cells were treated with the 40 µmol/l ST2825
and 1 mg/l LPS for 24 h. The control cells were cultured with DMEM.
Next, cells were lysed in a radioimmunoprecipitation assay buffer
containing 1% NP-40, 150 mM NaCl, 50 mM Tris-HCl, 50 mM NaF and 1
mM Na3VO4. The protein concentration in the
cell lysate was determined using a BCA Protein Assay kit. A total
of 40 µg protein was then separated via 20% SDS-PAGE, followed by
transfer to a nitrocellulose membrane. The membranes were blocked
with 5% milk in Tris-buffered saline/Tween-20 (TBST) buffer at 4°C
overnight and probed with primary antibodies against TLR4, IL-23 or
IL-17A, and β-actin was used as the internal control. Membranes
were subsequently washed three times with TBST for 5 min each and
incubated with a horseradish peroxidase-conjugated secondary
antibody for 1 h at room temperature. Following washing five times
with 1X TBST for 5 min each time, the membranes were developed with
ECL detection reagents, and the signal intensity was quantified by
ImageJ (version 1.45s; National Institutes of Health, Bethesda, MD,
USA).
Statistical analysis
All statistical analyses were performed with SPSS
version 17.0 software (SPSS, Inc., Chicago, IL, USA). Quantitative
variables are presented as the mean ± standard deviation, and were
analyzed with the Student's t-test, one-way analysis of variance,
χ2 test or Pearson's correlation coefficient. P<0.05
was considered to indicate statistically significant
differences.
Results
TLR4 expression is upregulated in HCC
tissues
TLR4 is a receptor that serves an important role in
inflammation (28). The expression of
TLR4 has been identified in several types of cancer, including
liver, breast, colon, ovarian and prostate cancer, and melanoma,
and its expression contributes to carcinogenesis (6–10,13). In order to determine whether TLR4 is
involved in the development of HCC, the expression of TLR4 in HCC
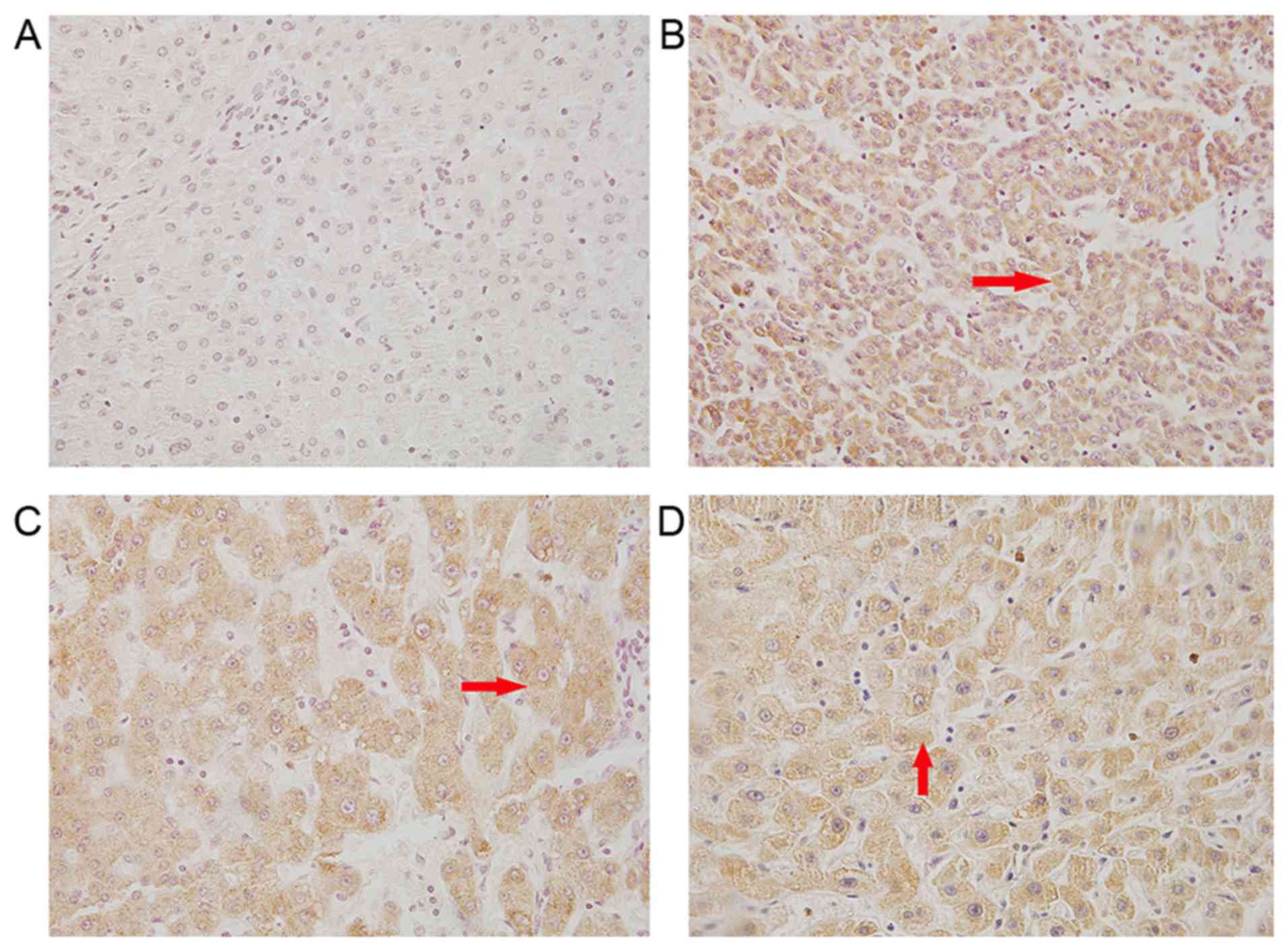
tissues obtained from patients was assessed. Immunohistochemical
staining with TLR4 antibody revealed that TLR4 was expressed in the
cytoplasm and cell surface of cancer cells in the tissue sections,
as observed by yellow and brown staining (Fig. 1). The positive rate of TLR4 in HCC
tissues was 77.8% (35/45; Table II),
while this rate was only 20% in adjacent noncancerous tissues,
suggesting an upregulation of TLR4 expression in HCC.
| Table II.Statistical analysis of TLR4, IL-23
and IL-17A expression and clinicopathological factors in
hepatocellular carcinoma. |
Table II.
Statistical analysis of TLR4, IL-23
and IL-17A expression and clinicopathological factors in
hepatocellular carcinoma.
|
| TLR4 | IL-23 | IL-17A |
|---|
|
|
|
|
|
|---|
|
Characteristics | Patients | (+)a | χ2 | P-value | Patients | (+)a | χ2 | P-value | R (vs.TLR4) | Patients | (+)a | χ2 | P-value | R (vs.TLR4) |
|---|
| Cancer
patients | 45 | 35 (77.8%) | 8.916 | 0.003 | 45 | 32 (71.1%) | 14.347 | 0.0002 | 0.634 | 45 | 30 (66.7%) | 11.9 | 0.0005 | 0.583 |
| Control
tissues | 17 | 5 (29.4%) |
|
| 17 | 3 (17.6%) |
|
|
| 17 | 3 (17.6%) |
|
|
|
| Age, years |
|
≤50 | 20 | 15 | 0.265 | 0.607 | 19 | 12 | 1.013 | 0.314 |
| 17 | 10 | 0.756 | 0.384 |
|
|
>50 | 25 | 17 |
|
| 26 | 20 |
|
|
| 28 | 20 |
|
|
|
| Sex |
|
Male | 39 | 33 | 7.912 | 0.005 | 37 | 30 | 10.07 | 0.002 |
| 38 | 29 | 6.287 | 0.012 |
|
|
Female | 6 | 2 |
|
| 8 | 2 |
|
|
| 7 | 2 |
|
|
|
| Tumor size, cm |
| ≤5 | 27 | 23 | 2.143 | 0.143 | 25 | 18 | 0.022 | 0.883 |
| 25 | 18 | 0.72 | 0.396 |
|
|
>5 | 18 | 12 |
|
| 20 | 14 |
|
|
| 20 | 12 |
|
|
|
| Tumor
metastasis |
|
Positive | 20 | 15 | 5.513 | 0.019 | 30 | 25 | 6.545 | 0.01 |
| 34 | 25 | 5.01 | 0.025 |
|
|
Negative | 25 | 10 |
|
| 15 | 7 |
|
|
| 11 | 4 |
|
|
|
| TNM stage |
|
T1+T2 | 25 | 18 | 4.36 | 0.037 | 27 | 9 | 4.821 | 0.028 |
| 26 | 6 | 4.185 | 0.041 |
|
|
T3+T4 | 20 | 17 |
|
| 18 | 12 |
|
|
| 19 | 10 |
|
|
|
| Tumor
differentiation |
|
High | 25 | 18 | 1.086 | 0.297 | 27 | 21 | 6.682 | 0.409 |
| 26 | 20 | 2.915 | 0.088 |
|
|
Poor | 20 | 17 |
|
| 18 | 12 |
|
|
| 19 | 10 |
|
|
|
TLR4 expression is correlated with HCC
clinicopathological characteristics
To further examine whether TLR4 expression in HCC
serves a role in the carcinogenesis, the correlation between TLR4
expression and the clinicopathological characteristics of HCC
patients was analyzed. As shown in Table
II, there was no significant correlation of TLR4 expression
with the age, tumor size or tumor differentiation of patients
(P>0.05). However, high expression of TLR4 was correlated with
sex and a lower TNM stage (P<0.05; Table II), suggesting that TLR4 expression
may contribute to the development of HCC.
TLR4 expression is correlated with the
expression levels of IL-17A and IL-23 in HCC
IL-17A and IL-23 are key mediators of inflammation
that contribute to carcinogenesis (29,30). In
order to establish whether TLR4 promotes carcinogenesis through
IL-17A and IL-23 regulation, the correlation between TLR4
expression and the levels of IL-17A and IL-23 was examined. The
results identified that the expression of TLR4 was associated with
the levels of IL-17A (R=0.583) and IL-23 (R=0.634) in HCC (Table II), indicating a possible regulation
effect between the expression of TLR4 and that of IL-17A and
IL-23.
Expression levels of IL-23 and IL-17A
are regulated by TLR4/MyD88 in HCC cells
TLR4 is a receptor expressed in all cell types in
the liver, including hepatocytes cells, and is able to recognize
LPS from the bacterial wall. Activation of TLR4 triggers an
inflammatory response through MyD88-mediated activation of
downstream signaling pathways (4,5). As TLR4
expression was correlated with IL-23 and IL-17A expression in HCC
tumors, as discussed earlier, the study further examined whether
activation of the TLR4/MyD88 pathway regulates the expression
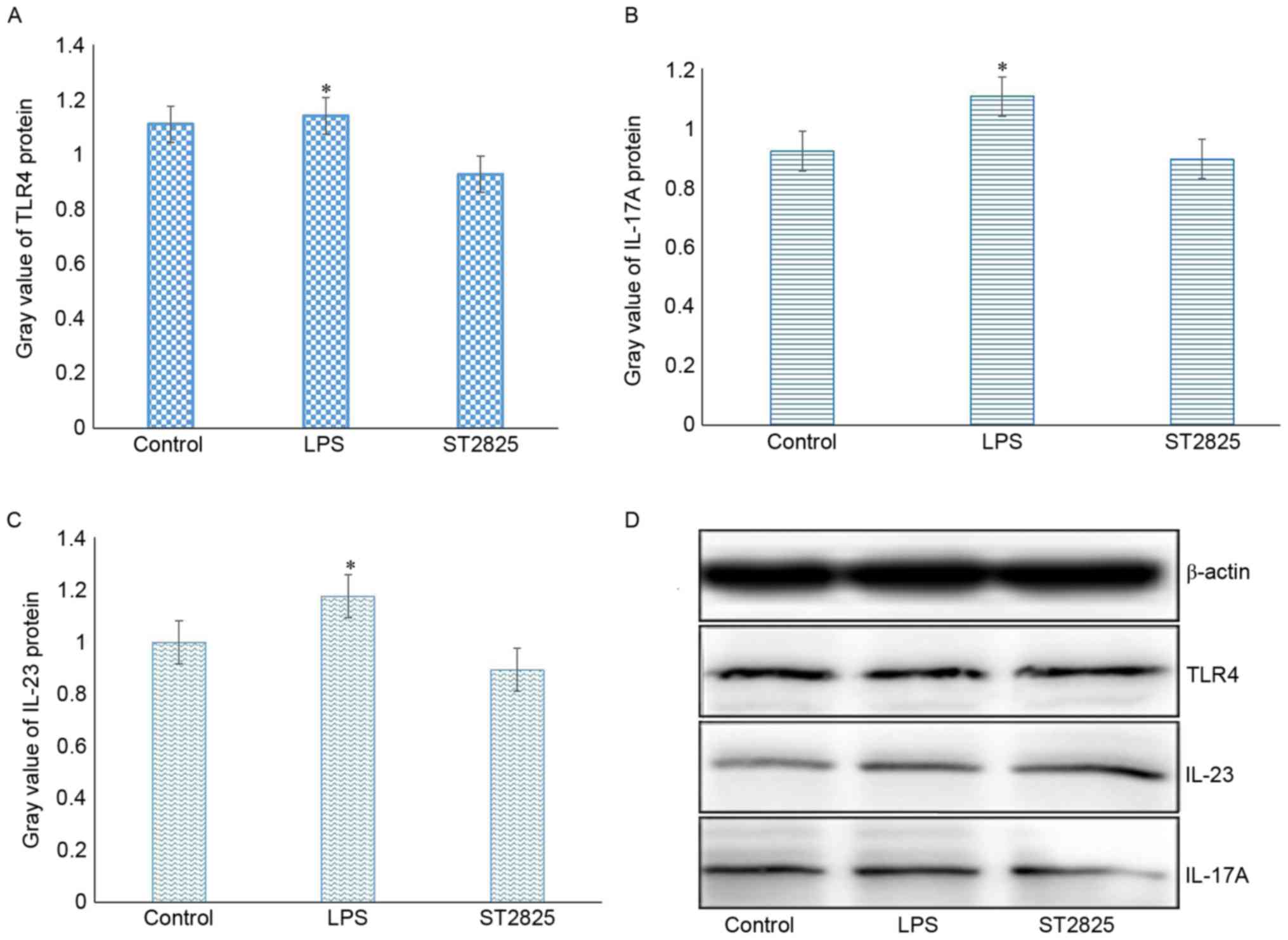
levels of IL-23 and IL-17A proteins. As determined by western blot
analysis, LPS induced the activation of TLR4, leading to enhanced
expression of IL-23 and IL-17A in HCC cells. The expression of
these interleukins was subsequently blocked by incubation with the
MyD88 inhibitor, ST2825, and showed a time-dependent effect
(Fig. 2). Analysis of IL-23 and
IL-17A levels using an ELISA assay further confirmed this result,
indicating that the expression levels of IL-23 and IL-17A were
regulated by the TLR4/MyD88 signaling pathway in HCC cells
(Fig. 2).
TLR4/MyD88 signaling pathway regulates
the transcription of IL-23 and IL-17A
Enhanced gene expression may be due to the
upregulation of gene transcription. Using RT-qPCR, it was observed
that LPS treatment of HepG2 cells resulted in enhanced
transcription of IL-23 and IL-17A, while this upregulation of gene
transcription was then blocked upon incubation with a MyD88
inhibitor (Fig. 3). These findings
indicated that activation of the TLR4/MyD88 signaling pathway
enhances the expression of IL-23 and IL-17A through the
upregulation of gene transcription.
Activation of TLR4/MyD88 enhances TLR4
expression
The present study further examined whether
activation of TLR4 regulates the expression of itself. RT-qPCR and
western blot analyses revealed that stimulation of HepG2 cells with
the TLR4-ligand LPS enhanced the expression levels of TLR4 mRNA and
protein, whereas application of the MyD88 inhibitor blocked the
upregulation of TLR4 expression induced by LPS. These results
indicate a positive feedback loop of TLR4 activation and its
expression (Fig. 4).
Expression levels of IL-23 and IL-17A
are correlated with the HCC clinicopathological
characteristics
Since the expression of TLR4 was correlated with the
clinical features of HCC, and activation of TLR4 was observed to
upregulate the expression levels of IL-23 and IL-17A, the current
study further investigated whether the levels of IL-23 and IL-17A
are also correlated with the clinical features of HCC. Analysis of
the protein expression levels of IL-23 and IL-17A in HCC tumor
tissues and adjacent normal tissues indicated that they were
associated with sex, lymph node metastasis and the TNM stages of
HCC (P<0.05). By contrast, no association was detected between
these protein levels and the patient age, tumor size and tumor
differentiation (P>0.05), as shown in Table II.
Discussion
Chronic inflammation is associated with the
initiation, development and progression of carcinomas, and may
contribute to cancer progression by activating angiogenesis and
oncogenic signaling pathways (31).
It is well known that HCC is a long-term consequence of chronic
inflammatory liver diseases (32). In
the present study, it was demonstrated that TLR4, IL-23 and IL-17A,
which are involved in inflammation, were highly expressed in
carcinoma tissues. In addition, the expression levels of TLR4,
IL-23 and IL-17A were associated with lymph node metastasis and the
TNM stages of HCC. It has previously been reported that the
expression of TLR4 and MyD88 may be associated with breast cancer
growth and distant metastases (6). In
another study, Grivennikov et al (33) observed that IL-23 signaling promoted
tumor growth and progression, as well as the development of a
tumoral IL-17 response. The current study finding further supported
the important role of the TLR4/MyD88 signaling pathway and
IL-23/IL-17A in cancer.
The TLR4/MyD88 pathway mediates the activation of
NF-κB and subsequent production of pro-inflammatory cytokines,
including IL-1β, IL-6 and TNF-α. These cytokines stimulate myeloid
dendritic cells to secret IL-23, which promotes Th17 cell
differentiation, proliferation and maintenance (34,35). It
has been reported that the IL-23/IL-17A axis promotes the formation
of lung metastases through tumor-endothelial transmigration
(36). In the present study, it was
observed that the expression of TLR4 was associated with the
expression levels of IL-17A (R=0.583) and IL-23 (R=0.634). To
determine the possible role of the TLR4/MyD88 pathway and the
IL-23/IL-17A axis in HCC, the study measured the TLR4, IL-23 and
IL-17A expression levels in HepG2 cells stimulated with LPS using
RT-qPCR and western blot analyses. The results revealed that LPS
promoted the mRNA transcription and protein expression levels of
TLR4, IL-23 and IL-17A in HepG2 cell lines, which were subsequently
inhibited by incubation with the MyD88 inhibitor ST2825. Teng et
al (37) previously reported that
IL-23 was induced in response to stimulation by multiple TLR
ligands, and this cytokine has been linked to autoimmune and
inflammatory diseases. IL-23 consists of p19 and p40 units, while
Brentano et al (38) further
observed that the induction of p19 was TLR-dependent. In the
present study, the data revealed that TLR4/MyD88 signaling
regulated the expression levels of IL-23 and IL-17A in
hepatocellular tumor cells.
In conclusion, the current study identified that the
TLR4/MyD88 signaling pathway may be involved in hepatocellular
carcinogenesis via upregulating the IL-23/IL-17A axis, and that it
may serve as an important therapeutic target of HCC.
Acknowledgements
The authors would like to thank all the members of
Key Laboratory of Pathogen Biology and Immunology (Ningxia Medical
University, Yinchuan, China) for their assistance. The present
study was supported by the Ningxia Natural Science Foundation
Program (grant no. NZ14139) and the West China First-Class
Discipline Construction Project in Basic Medicine funded by Ningxia
Medical University.
References
|
1
|
Eiró N, Altadill A, Juárez LM, Rodríguez
M, González LO, Atienza S, Bermúdez S, Fernandez-Garcia B,
Fresno-Forcelledo MF, Rodrigo L and Vizoso FJ: Toll-like receptors
3, 4 and 9 in hepatocellular carcinoma: Relationship with
clinicopathological characteristics and prognosis. Hepatol Res.
44:769–778. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shlomai A, de Jong YP and Rice CM: Virus
associated malignancies: The role of viral hepatitis in
hepatocellular carcinoma. Semin Cancer Biol. 26:78–88. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vescovo T, Refolo G, Vitagliano G, Fimia
GM and Piacentini M: Molecular mechanisms of hepatitis C
virus-induced hepatocellular carcinoma. Clin Microbiol Infect.
22:853–861. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jack CS, Arbour N, Manusow J, Montgrain V,
Blain M, McCrea E, Shapiro A and Antel JP: TLr signaling tailors
innate immune responses in human microglia and astrocytes. J
Immunol. 175:4320–4330. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zughaier SM, Zimmer SM, Datta A, Carlson
RW and Stephens DS: Differential induction of the toll-like
receptor 4-MyD88-dependent and independent signaling pathways by
endotoxins. Infect Immun. 73:2940–2950. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mehmeti M, Allaoui R, Bergenfelz C, Saal
LH, Ethier SP, Johansson ME, Jirström K and Leandersson K:
Expression of functional toll like receptor 4 in estrogen
receptor/progesterone receptor-negative breast cancer. Breast
Cancer Res. 17:1302015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takazawa Y, Kiniwa Y, Ogawa E, Uchiyama A,
Ashida A, Uhara H, Goto Y and Okuyama R: Toll-like receptor 4
signaling promotes the migration of human melanoma cells. Tohoku J
Exp Med. 234:57–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Semlali A, Parine Reddy N, Arafah M,
Mansour L, Azzi A, Al Shahrani O, Al Amri A, Shaik JP, Aljebreen
AM, Alharbi O, et al: Expression and polymorphism of toll-like
receptor 4 and effect on NF-κB mediated inflammation in colon
cancer patients. PLoS One. 11:e01463332016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo XZ, He QZ and Wang K: Expression of
Toll-like receptor 4 in ovarian serous adenocarcinoma and
correlation with clinical stage and pathological grade. Int J Clin
Exp Med. 8:14323–14327. 2015.PubMed/NCBI
|
|
10
|
Shui IM, Stark JR, Penney KL, Schumacher
FR, Epstein MM, Pitt MJ, Stampfer MJ, Tamimi RM, Lindstrom S, Sesso
HD, et al: Genetic variation in the toll-like receptor 4 and
prostate cancer incidence and mortality. Prostate. 72:209–216.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang HY, Zhang ZJ, Cao CB, Wang N, Liu
FF, Peng JQ, Ren XJ and Qian J: The TLR4/NF-κB signaling pathway
mediates the growth of colon cancer. Eur Rev Med Pharmacol Sci.
18:3834–3843. 2014.PubMed/NCBI
|
|
12
|
Chen X, Zhao F, Zhang H, Zhu Y, Wu K and
Tan G: Significance of TLR4-MyD88 expression in breast cancer. Int
J Clin Exp Pathol. 8:7034–7039. 2015.PubMed/NCBI
|
|
13
|
Wang L, Zhu R, Huang Z, Li H and Zhu H:
Lipopolysaccharide-induced toll-like receptor 4 signaling in cancer
cells promotes cell survival and proliferation in hepatocellular
carcinoma. Dig Dis Sci. 58:2223–2236. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu WT, Jing YY, Yu GF, Han ZP, Yu DD, Fan
QM, Ye F, Li R, Gao L, Zhao QD, et al: Toll like receptor 4
facilitates invasion and migration as a cancer stem cell marker in
hepatocellular carcinoma. Cancer Lett. 358:136–143. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Heinrich EL, Walser TC, Krysan K, Liclican
EL, Grant JL, Rodriguez NL and Dubinett SM: The inflammatory tumor
microenvironment, epithelial mesenchymal transition and lung
carcinogenesis. Cancer Microenviron. 5:5–18. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grivennikov SI: Inflammation and
colorectal cancer: Colitis-associated neoplasia. Semin
Immunopathol. 35:229–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kawanishi S, Ohnishi S, Ma N, Hiraku Y,
Oikawa S and Murata M: Nitrative and oxidative DNA damage in
infection-related carcinogenesis in relation to cancer stem cells.
Genes Environ. 38:262017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rasic I, Radovic S and Aksamija G:
Relationship between chronic inflammation and the stage and
histopathological size of colorectal carcinoma. Med Arch.
70:104–107. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Langrish CL, Chen Y, Blumenschein WM,
Mattson J, Basham B, Sedgwick JD, McClanahan T, Kastelein RA and
Cua DJ: IL-23 drives a pathogenic T cell population that induces
autoimmune inflammation. J Exp Med. 201:233–240. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kolls JK and Lindén A: Interleukin-17
family members and inflammation. Immunity. 21:467–476. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Toussirot É: The IL23/Th17 pathway as a
therapeutic target in chronic inflammatory diseases. Inflamm
Allergy Drug Targets. 11:159–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kortylewski M, Xin H, Kujawski M, Lee H,
Liu Y, Harris T, Drake C, Pardoll D and Yu H: Regulation of the
IL-23 and IL-12 balance by Stat3 signaling in the tumor
microenvironment. Cancer Cell. 15:114–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu D, Wu P, Huang Q, Liu Y, Ye J and Huang
J: Interleukin-17: A promoter in colorectal cancer progression.
Clin Dev Immunol. 2013:4363072013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qin S: Primary Liver Cancer Diagnosis and
Treatment Expert Panel of the Chinese Ministry of Health:
Guidelines on the diagnosis and treatment of primary liver cancer
(2011 edition). Chin Clin Oncol. 1:2304–3865. 2012.
|
|
25
|
Chen ZH, Hong YF, Lin J, Li X, Wu DH, Wen
JY, Chen J, Ruan DY, Lin Q, Dong M, et al: Validation and ranking
of seven staging systems of hepatocellular carcinoma. Oncol Lett.
14:705–714. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song Y, Zhou M, Cao Y, Qi J, Geng J and
Liu X: Expression of GLP-1 receptor and CD26 in human thyroid
C-cells: The association of thyroid C-cell tumorigenesis with
incretin-based medicine. Oncol Lett. 13:2684–2690. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ruifeng G, Yunhe F, Zhengkai W, Ershun Z,
Yimeng L, Minjun Y, Xiaojing S, Zhengtao Y and Naisheng Z:
Chlorogenic acid attenuates lipopolysaccharide-induced mice
mastitis by suppressing TLR4-mediated NF-κB signaling pathway. Eur
J Pharmacol. 729:54–58. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Langowski JL, Zhang X, Wu L, Mattson JD,
Chen T, Smith K, Basham B, McClanahan T, Kastelein RA and Oft M:
IL-23 promotes tumour incidence and growth. Nature. 442:461–465.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou Y, Wu PW, Yuan XW, Li J and Shi XL:
Interleukin-17A inhibits cell autophagy under starvation and
promotes cell migration via TAB2/TAB3-p38 mitogen-activated protein
kinase pathways in hepatocellular carcinoma. Eur Rev Med Pharmacol
Sci. 20:250–263. 2016.PubMed/NCBI
|
|
31
|
Sakurai T, Kashida H, Watanabe T, Hagiwara
S, Mizushima T, Iijima H, Nishida N, Higashitsuji H, Fujita J and
Kudo M: Stress response protein cirp links inflammation and
tumorigenesis in colitis-associated cancer. Cancer Res.
74:6119–6128. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang H, Liu J, Hu X, Liu S and He B:
Prognostic and therapeutic values of tumor necrosis factor-alpha in
hepatocellular carcinoma. Med Sci Monit. 22:3694–3704. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grivennikov SI, Wang K, Mucida D, Stewart
CA, Schnabl B, Jauch D, Taniguchi K, Yu GY, Osterreicher CH, Hung
KE, et al: Adenoma-linked barrier defects and microbial products
drive IL-23/IL-17-mediated tumour growth. Nature. 491:254–258.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Blauvelt A, Lebwohl MG and Bissonnette R:
IL-23/IL-17A dysfunction phenotypes inform possible clinical
effects from anti-IL-17A therapies. J Invest Dermatol.
135:1946–1953. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
González-Reyes S, Marín L, González L,
González LO, del Casar JM, Lamelas ML, González-Quintana JM and
Vizoso FJ: Study of TLR3, TLR4 and TLR9 in breast carcinomas and
their association with metastasis. BMC Cancer. 10:6652010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kulig P, Burkhard S, Mikita-Geoffroy J,
Croxford AL, Hövelmeyer N, Gyülvészi G, Gorzelanny C, Waisman A,
Borsig L and Becher B: IL17A-mediated endothelial breach promotes
metastasis formation. Cancer Immunol Res. 4:26–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Teng MW, Bowman EP, McElwee JJ, Smyth MJ,
Casanova JL, Cooper AM and Cua DJ: IL-12 and IL-23 cytokines: From
discovery to targeted therapies for immune-mediated inflammatory
diseases. Nat Med. 21:719–729. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brentano F, Ospelt C, Stanczyk J, Gay RE,
Gay S and Kyburz D: Abundant expression of the interleukin (IL)23
subunit p19, but low levels of bioactive IL23 in the rheumatoid
synovium: Differential expression and Toll-like receptor-(TLR)
dependent regulation of the IL23 subunits, p19 and p40, in
rheumatoid arthritis. Ann Rheum Dis. 68:143–150. 2009. View Article : Google Scholar : PubMed/NCBI
|