Introduction
Hydatidiform mole is a gestational trophoblastic
disease, classified into partial mole (PM) and complete mole (CM),
which have the potential to develop into a persistent tumor
(1,2).
The incidence of hydatidiform mole varies worldwide, at 1/200
livebirths in Southeast Asia, 1/1,000 gestations in Europe and
1/2,000 gestations in the United States (3,4). It is
treated by surgical suction evacuation and serial serum human
chorionic gonadotropin (hCG) estimations. Spontaneous regression
following molar evacuation occurs in 80–90% cases, and such
patients do not require further treatment (5–7). In 10–20%
cases, development into invasive mole and choriocarcinoma occurs.
Choriocarcinoma is a malignant trophoblast-cell tumor,
characterized by infiltrative and destructive growth, and elevated
levels of β-human chorionic gonadotrophin (β-hCG) (2,5,6,8). However,
the molecular mechanism of its pathology remains unclear.
Previous findings have demonstrated that uterine
natural killer (uNK) cells serve a role in the control of
trophoblast invasion, placental development and remodeling of
spiral arteries (7,9,10). uNK
cells were originally identified by the presence of cytoplasmic
granules, thus, they were previously termed granular metrial gland
cells (9,11,12).
Although uNK cells are granular and express essential proteins
required for lysis, they exhibit significantly reduced cytotoxic
activity compared with peripheral blood NK cells (10,13,14).
Glycoconjugate contents in cytoplasmic granules of uNK cells have
been demonstrated to react with Shiff's reagent following periodate
treatment, therefore, Periodic acid-Schiff (PAS) staining was
established as a method of uNK-cell identification in the mouse and
human endometrium (11,12,15).
However, the more recently developed dolichos biflorus
agglutinin (DBA) lectin staining was demonstrated to be a more
sensitive method for the identification and localization of uNK
cells (16,17). DBA lectin can react with
glycoconjugates within the granules and at the surface of uNK
cells. Furthermore, the DBA lectin method has been used to classify
uNK cells into mature and immature uNK cell subtypes (18).
Prior investigators have demonstrated that uNK cells
are associated with gestational trophoblastic diseases (19–22).
However, it remains contradictory whether the number of uNK cells
is altered in gestational trophoblastic diseases (20–23). In
the present study, the distribution and number of uNK cells in
normal pregnancy (NP), hydatidiform molar and invasive molar
pregnancy were performed using DBA lectin binding activity.
Materials and methods
Tissue collection
Tissues and clinical information were obtained under
the approval of the Ethics Committee of Jiangxi Province People's
Hospital (Nanchang, China), and all participants provided informed
consent. Tissue samples of normal decidua and placenta of first
trimester normal pregnancies (n=15; average age of participant,
22.3 years, age range, 19–32 years) were collected from healthy
women undergoing suction termination of pregnancy (term, 6–11
weeks) between March 2012 and August 2012, for non-medical reasons.
Normal term placenta tissues were collected following uncomplicated
pregnancies and vaginal deliveries (n=15; average age of
participant, 25.7 years; age range, 20–32 years) between March 2012
and November 2012. Paraffin-embedded PM (n=22; average age of
participant, 26.1 years; age range, 20–33 years), complete
hydatidiform mole (n=20; average age of participant, 29.8 years,
age range, 22–33 years) and invasive mole (n=10; average age of
participant, 33.3 years, age range, 24–36 years) tissues, which had
been collected between January 2010 and December 2012, were
supplied by the Pathology Department of Jiangxi Province People's
Hospital. All the tissue sections were assessed by a gynecological
pathologist (Department of Pathology of Jiangxi Province People's
Hospital) to confirm diagnosis and ensure suitability for
experimental investigation. PM was diagnosed when there was
evidence of fetal development, including nucleated red blood cells,
or when hydropic change was observed in only a portion of chorionic
villi (1). CM was diagnosed by the
presence of hydatidiform mole without a fetus, characterized by the
transformation of chorionic villi into clusters of vesicles with
variable dimensions (1). Invasive
mole was diagnosed by the presence of hydatidiform mole, and,
microscopically, by the invasion of the myometrium, blood vessels
or extrauterine sites by hydropic villi (1). All the patients were physically examined
by ultrasound, and patients suffering from chronic disease, uterine
abnormalities, infection or genetic abnormalities were
excluded.
Mice
Adult Kunming white strain mice (n=6; 25–30 g; 9
weeks old; Obtained from Nanchang University, Jianxi, China) were
housed in a constant photoperiod (14/10 h light/dark cycle) and
relative humidity (55±10%) at room temperature with food and water
available ad libitum. All the animal procedures were
approved by the Institutional Animal Care and Use Committee of
Nanchang University (Nanchang, China). Adult female virgin mice
were mated with fertile males to induce pregnancy. The
identification of a vaginal plug was considered as day 1 of
pregnancy. On day 8 of pregnancy, mice were anesthetized with 45
mg/kg pentobarbital sodium prior to cervical dislocation and
uterine collection between 8:00 and 9:00 a.m. The uterine tissues
were frozen at −80°C for later use as positive controls of DBA
lectin staining (24).
Histopathological examination
Tissue samples were fixed in 4% neutral-buffered
formalin (4 h, room temperature) and embedded in paraffin. The
paraffin sections (5 µm) were mounted on glass slides coated with
poly-l-lysine, then deparaffinized in 100% xylene, rehydrated in
graded ethanol solutions (100, 95, 80 and 70%), and stained with
hematoxylin and eosin. The pathological diagnosis of partial,
complete molar and invasive molar pregnancies was based on the
aforementioned standard histopathological criteria (1), and performed by a gynecological
pathologist. The hydatidiform mole and invasive mole samples were
further stained with anti-human cytokeratin (dilution, 1:200;
sc-70906; Santa Cruz Biotechnology, Santa Cruz, USA) to identify
the presence and position of trophoblast cells overnight at 4°C.
The sections were incubated with a horseradish
peroxidase-conjugated secondary antibody (dilution, 1:200; ZB-2306)
for 50 min at 37°C followed by 0.05% fresh diaminobenzidine
solution for 4 min, together with counter-staining with 5% Harris'
hematoxylin solution for 5 sec at room temperature (all Zhongshan
Golden Bridge Biotechnology Co., Ltd., Beijing, China).
Brown-yellow staining in the cytoplasm was observed to indicate a
positive signal in a conventional light microscope (magnification,
×200; Olympus Corporation, Tokyo, Japan).
DBA lectin immunohistochemistry
DBA lectin staining was performed as previously
described (24). Endogenous
peroxidase activity was blocked by incubating the sections in 3%
H2O2 for 10 min at room temperature.
Non-specific antibody binding was blocked in 5% bovine serum
albumin (Zhongshan Golden Bridge Biotechnology Co., Ltd.) for 60
min at room temperature. The sections were then incubated in
biotinylated-DBA lectin (dilution, 1:1,500; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) overnight at 4°C. Following washing in
PBS, the sections were incubated with streptavidin-peroxidase for
another 60 min at 37°C. Positive signals were detected with 0.05%
3,3′-diaminobenzidine solution and counter-stained with 5% Harris'
hematoxylin solution (all Zhongshan Golden Bridge Biotechnology
Co., Ltd.) 2 min at room temperature. The addition of 0.1 M
N-acetyl-D-galactosamine (Sigma-Aldrich; Merck KGaA) to the DBA
lectin incubation provided a negative control.
Microscopic evaluation
Microscopic evaluation was performed blindly by two
independent pathologists using a light microscope. The number of
cells positive for DBA lectin staining was counted in 10
non-overlapping fields at magnification, ×400. The procedure was
repeated in 2 specimens from each subject. The mean number of cells
was calculated per patient, then per patient group.
Statistics analysis
The data are presented as the mean ± standard error
of the mean. The Mann-Whitney U test was used to determine
statistically significant differences. P<0.05 was considered to
indicate a statistically significant difference. All statistical
analyses were performed using SPSS 13.0 (SPSS, Inc., Chicago, IL,
USA).
Results
Distribution and number of uNK cells
in NP
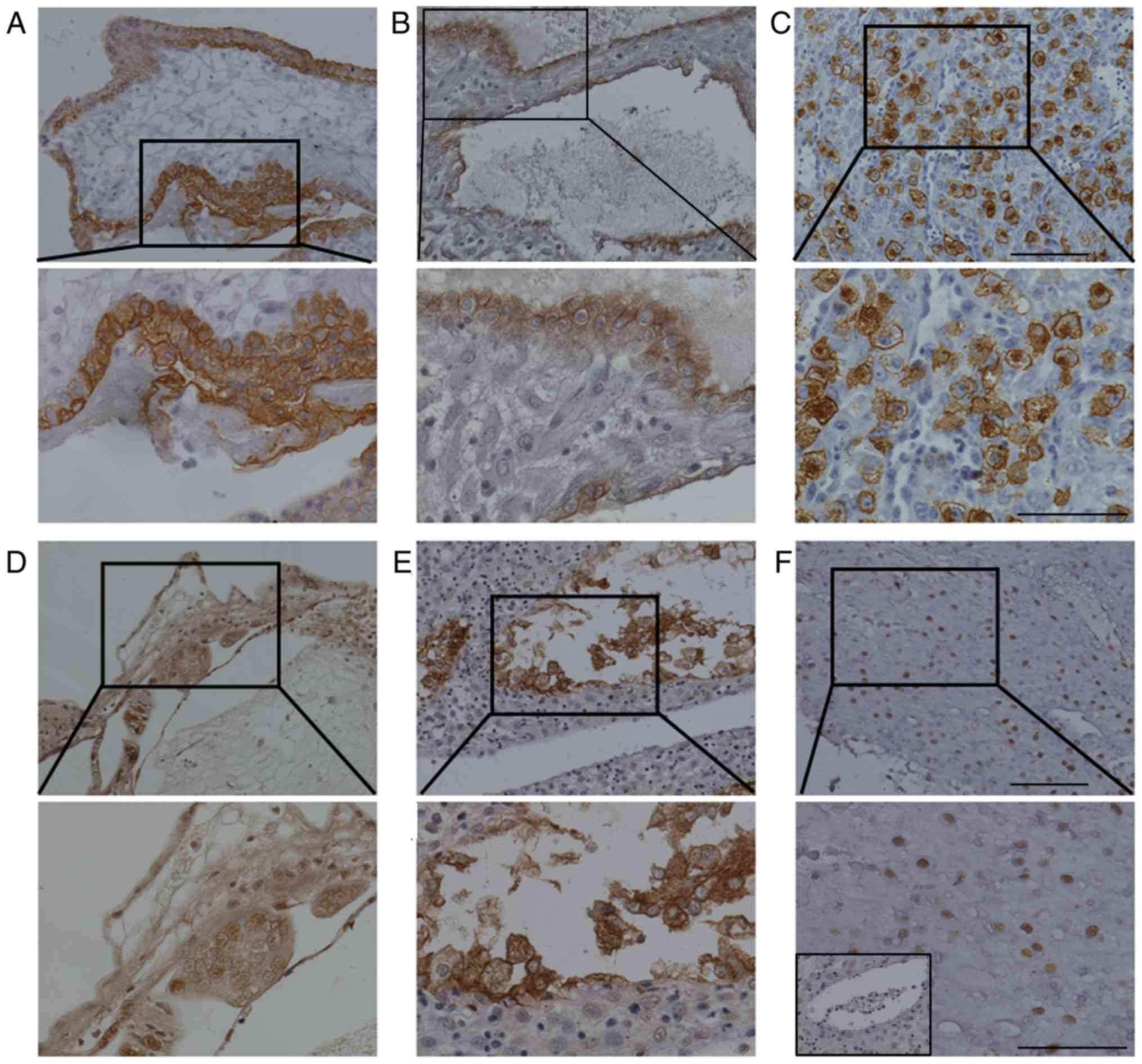
Firstly, the identity of trophoblastic cells in the
villous (Fig. 1A) and decidual tissue
(Fig. 1B) was confirmed by staining
for cytokeratin. Mouse uterine tissues from day 8 of NP were
stained with DBA lectin as a positive control (Fig. 1C). The results demonstrated that uNK
cells were located at the mesometrial pole of implantation sites.
The cells were large in size and exhibited heavy cytoplasmic
granules (Fig. 1C). In villi of early
NP, a number of small uNK cells were distributed in the villous
stroma (Fig. 1D). In the deciduas of
early NP, DBA lectin staining demonstrated that 2 forms of uNK
cells were located in different regions of the decidual tissues
(Fig. 1E). Mature uNK cells were
large in size and exhibited strong brown staining of the cellular
cytoplasm. The membranes were mainly distributed near endothelial
cells of decidual blood vessels (Fig.
1E). The smaller immature uNK cells exhibited weak brown
staining of the nucleus and were scattered among decidual cells
(Fig. 1F). The number of uNK cells in
the villi was significantly lower than that in the decidual samples
of early pregnancy (46.8±5.3 vs. 142.3±10.7; Table I). However, at full term NP, DBA
lectin staining was almost negligible in decidual and placental
chorionic villous tissue (Fig. 2;
Table I).
 | Table I.Number of uterine natural killer
cells in normal, hydatidiform molar and invasive molar
pregnancy. |
Table I.
Number of uterine natural killer
cells in normal, hydatidiform molar and invasive molar
pregnancy.
|
|
| Tissue
location |
|---|
|
|
|
|
|---|
| Pregnancy type | Number | Deciduas | Villi |
|---|
| Early normal | 15 | 142.3±10.7 |
46.8±5.3a |
| Full term
normal | 15 |
2.6±1.2a | ND |
| Partial
hydatidiform mole | 22 | 156.4±15.6 | ND |
| Complete
hydatidiform mole | 20 |
10.3±3.9a | ND |
| Invasive
hydatidiform mole | 10 |
4.1±2.2a | ND |
Distribution and number of uNK cells
in PM
H&E staining for deciduas (Fig. 3A) and villi (Fig. 3B) of PM was observed to identify the
mole histologically. The identity of trophoblastic cells in the
decidua (Fig. 3C) and villi (Fig. 3D) of PM was confirmed by staining for
cytokeratin. DBA lectin staining demonstrated that decidual uNK
cells were mainly located near blood vessel endothelial cells in PM
(Fig. 3E). No significant difference
was detected between the number of decidual uNK cells in PM and
early NP (156.4±15.6 vs. 142.3±10.7; P>0.05). In addition, DBA
lectin staining was not detected in the villi of PM (Fig. 3F).
Distribution and number of uNK cells
in CM
H&E staining for deciduas (Fig. 4A) and villi (Fig. 4B) of CM was observed to identify the
mole histologically. The identity of trophoblastic cells in the
decidua (Fig. 4C) and villi (Fig. 4D) of CM was also confirmed by staining
for cytokeratin. In CM, decidual uNK cells were also identified
near blood vessel endothelial cells (Fig.
4E). However, the number of decidual uNK cells was
significantly decreased in CM (10.3±3.9; P<0.01) compared with
early NP (142.3±10.7, Table I). In
addition, DBA lectin staining was also not detected in the villi of
CM (Fig. 4F).
Distribution and number of uNK cells
in IM
H&E staining for deciduas (Fig. 5A) and villi (Fig. 5B) of IM was observed to identify the
mole histologically. The identity of trophoblastic cells in the
decidua (Fig. 5C) and villi (Fig. 5D) of IM was also confirmed by staining
for cytokeratin. In IM, decidual uNK cells were also identified
near blood vessel endothelial cells (Fig.
5E). However, the number of decidual uNK cells was
significantly decreased in IM (4.1±2.2; P<0.01) compared with
early NP (142.3±10.7, Table I). No
significant difference was identified with regard to the number of
decidual uNK cells in CM and IM (10.3±3.9 vs. 4.1±2.2; P>0.05).
In addition, DBA lectin staining was also not detected in the villi
of IM (Fig. 5F).
Discussion
The present study revealed that uNK cells were
mainly located in the uterine decidual tissue of early NP. With
progression of pregnancy, the number of uNK cells significantly
decreased. In the deciduas of hydatidiform mole (including PM, CM
and invasive mole), the DBA lectin staining demonstrated that uNK
cells were mainly distributed near blood vessel endothelial cells.
No significant difference was identified between the number of
decidual uNK cells in partial hydatidiform mole and early NP.
However, the number of decidual uNK cells was significantly
decreased in complete and invasive mole compared with early NP,
suggesting that CM has a high risk of developing malignant
trophoblast disease compared with PM. Bagshawe et al
(5) demonstrated that the risk of
developing a tumor was 15% in CM and 0.5% in PM.
A previous study demonstrated that the number of uNK
cells is dynamic within the menstrual cycle (25). uNK cells have been reported to be
absent in the proliferative endometrium, and the cell number has
been reported to increase in the mid- to late-luteal phase of the
menstrual cycle and in early pregnancy (9,25). The
present study demonstrated that uNK cells were highly abundant and
moderately scattered among uterine decidual cells in the first
trimester of pregnancy, which is in accordance with a previous
study (23). It was also identified
that a proportion of decidual uNK cells were distributed near blood
vessel endothelial cells in early pregnancy. It has been previously
suggested that blood vessel-adjacent uNK cells may be able to
secrete a series of cytokines, including interferon γ (IFN-γ),
angiopoietin (Ang)1, placental growth factor, transforming growth
factor-β1 (TGF-β1), and vascular endothelial growth factor (VEGF)C,
which are critical in angiogenesis and placental development
(26,27). Furthermore, it has been indicated that
mice with uNK-cell or IFN-γ signaling deficiencies (recombination
activating gene 2−/− or IFN-γ−/− mice,
respectively) exhibit decidual artery remodeling failure and have
implantation site abnormalities (28,29).
Contradictory data exist with respect to uNK cell
number in the third trimester of NP (9,20,23). A number of studies reported that uNK
cells expressing cluster of differentiation (CD)56+ (a
key marker for human uNK cells) were substantial in number in
decidual tissue in the third trimester of NP (20,23).
Williams et al (23)
demonstrated that substantial numbers of CD56+ uNK cells
remain in decidua at full term pregnancy. Other studies suggested
that uNK cells were virtually absent at full term NP
(20–22). In the present study, almost negligible
DBA lectin staining was evident in the decidual and villous tissue
at full term. These conflicting results may be due to variability
in samples and research techniques.
Cytogenetic studies demonstrated that the karyotype
of partial molar pregnancy is 69, XXY or 69, XXX, resulting from
the fertilization of an ovum by 2 sperm (1). Consequently, PM can be defined as a
partial allograft, as can NP. However, CM is defined as a complete
allograft seeing as all chromosomes are of paternal origin
(2,3).
An immunological response against foreign paternal antigens would
be predicted, therefore, researchers have explored immune cell
profiling in NP and molar pregnancy, including that of macrophages,
natural killer (NK) cells and T cells. Nagymanyoki et al
(22) revealed a significant increase
in cytotoxic T-cell and granzyme B (GrB)+ NK-cell
numbers at the implantation site of CM by immunostaining of CD8,
GrB and forkhead box P3. However, Knoeller et al (21) demonstrated that the number of CD8 T
cells, CD3 T cells and mast cells significantly increased, and
CD56+ NK cell number decreased, in choriocarcinoma and
hydatidiform mole compared with NP, using antibodies against CD8,
CD3, CD56, CD68 and mast cell tryptase. In the present study, it
was demonstrated that the number of decidual uNK cells was
significantly reduced in CM and invasive mole compared with early
NP. This suggests that uNK cells may serve a pivotal role in the
control of trophoblast invasion and proliferation. The likely
mechanisms of uNK cell regulation of trophoblast invasion are
cytokine and growth factor secretion, including IFN-γ, transforming
growth factor-β1, interleukin-1β, TNF-α, VEGFC, placental growth
factor, Ang-2, leukemia inhibitory factor and colony stimulating
factor-1 (30,31).
In summary, the present study demonstrates that the
number of decidual uNK cells was significantly reduced in full term
NP, CM and invasive mole compared with early NP. This suggests that
uNK cells may serve an important role in the regulation of
trophoblast invasion and proliferation. However, further research
is required to establish the role of uNK cells in the diagnosis and
treatment of gestational trophoblastic disease.
Acknowledgements
The authors would like to thank Dr Min Jiang at
Jiangxi Province People's Hospital for proofreading the
manuscript.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81671486), and the
555 project of Jiangxi Province Gan Po Excellence and Post-graduate
Innovation Project of Nanchang University (grant nos. cx2015176 and
cx2016355).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TZh, XX and HK conceived and designed the
experiment. TZh, XX, TZo and XY carried out all the experiments. YL
was responsible for collection of the samples. TZh and YL analyzed
the data. TZh, XX and HK drafted the paper. All authors revised the
paper, read and approved the final manuscript.
Ethics approval and consent to
participate
Patient tissues and clinical information were
obtained under the approval of Ethical Committee of Jiangxi
Province People's Hospital, and all participants provided informed
consent. All animal procedures were approved by the Institutional
Animal Care and Use Committee of Nanchang University.
Patient consent for publication
Written informed consent was obtained from each
patient for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cheung AN: Pathology of gestational
trophoblastic diseases. Best Pract Res Clin Obstet Gynaecol.
17:849–868. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Monchek R and Wiedaseck S: Gestational
trophoblastic disease: An overview. J Midwifery Womens Health.
57:255–259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Seckl MJ, Sebire NJ and Berkowitz RS:
Gestational trophoblastic disease. Lancet. 376:717–729. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tse KY, Chan KKL, Tam KF and Ngan HYS:
Gestational trophoblastic disease. Obstet Gynaecol Reprod Med.
19:89–97. 2009. View Article : Google Scholar
|
|
5
|
Bagshawe KD, Lawler SD, Paradinas FJ, Dent
J, Brown P and Boxer GM: Gestational trophoblastic tumours
following initial diagnosis of partial hydatidiform mole. Lancet.
335:1074–1076. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Berkowitz RS and Goldstein DP: Chorionic
tumors. N Engl J Med. 335:1740–1748. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sebire NJ, Fisher RA and Rees HC:
Histopathological diagnosis of partial and complete hydatidiform
mole in the first trimester of pregnancy. Pediatr Dev Pathol.
6:69–77. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turgut EN, Celik E, Celik S, Arikan DC,
Altuntas H, Leblebici C, Purisa S and Dansuk R: Could serum β-hCG
levels and gestational age be the indicative factors for the
prediction of the degree of trophoblastic invasion into tubal wall
in unruptured ampullary pregnancies? Arch Gynecol Obstet.
287:323–328. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bulmer JN and Lash GE: Human uterine
natural killer cells: A reappraisal. Mol Immunol. 42:511–521. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Santoni A, Carlino C and Gismondi A:
Uterine NK cell development, migration and function. Reprod Biomed
Online. 16:202–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stewart I and Peel S: Granulated metrial
gland cells at implantation sites of the pregnant mouse uterus.
Anat Embryol (Berl). 160:227–238. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bulmer D: Further studies on the
granulated metrial gland cells of the pregnant rat. J Anat.
103:479–489. 1968.PubMed/NCBI
|
|
13
|
Croy BA, He H, Esadeg S, Wei Q, McCartney
D, Zhang J, Borzychowski A, Ashkar AA, Black GP, Evans SS, et al:
Uterine natural killer cells: Insights into their cellular and
molecular biology from mouse modelling. Reproduction. 126:149–160.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van den Heuvel MJ, Xie X, Tayade C,
Peralta C, Fang Y, Leonard S, Paffaro VA Jr, Sheikhi AK, Murrant C
and Croy BA: A review of trafficking and activation of uterine
natural killer cells. Am J Reprod Immunol. 54:322–331. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hamperl H and Hellweg G: Granular
endometrial stroma cells. Obstet Gynecol. 11:379–387.
1958.PubMed/NCBI
|
|
16
|
Stewart IJ and Webster AJ: Lectin
histochemical studies of mouse granulated metrial gland cells.
Histochem J. 29:885–892. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stewart J, Bebington CR and Mukhtar DD:
Lectin binding characteristics of mouse placental cells. J Anat.
196:371–378. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Paffaro VA Jr, Bizinotto MC, Joazeiro PP
and Yamada AT: Subset classification of mouse uterine natural
killer cells by DBA lectin reactivity. Placenta. 24:479–488. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
El Costa H, Tabiasco J, Berrebi A, Parant
O, Aguerre-Girr M, Piccinni MP and Le Bouteiller P: Effector
functions of human decidual NK cells in healthy early pregnancy are
dependent on the specific engagement of natural cytotoxicity
receptors. J Reprod Immunol. 82:142–147. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Haller H, Radillo O, Rukavina D, Tedesco
F, Candussi G, Petrović O and Randić L: An immunohistochemical
study of leucocytes in human endometrium, first and third trimester
basal decidua. J Reprod Immunol. 23:41–49. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Knoeller S, Lim E, Aleta L, Hertwig K,
Dudenhausen JW and Arck PC: Distribution of immunocompetent cells
in decidua of controlled and uncontrolled
(choriocarcinoma/hydatidiform mole) trophoblast invasion. Am J
Reprod Immunol. 50:41–47. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagymanyoki Z, Callahan MJ, Parast MM,
Fulop V, Mok SC and Berkowitz RS: Immune cell profiling in normal
pregnancy, partial and complete molar pregnancy. Gynecol Oncol.
107:292–297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Williams PJ, Searle RF, Robson SC, Innes
BA and Bulmer JN: Decidual leucocyte populations in early to late
gestation normal human pregnancy. J Reprod Immunol. 82:24–31. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuang H, Peng H, Xu H, Zhang B, Peng J and
Tan Y: Hormonal regulation of uterine natural killer cells in mouse
preimplantation uterus. J Mol Histol. 41:1–7. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Igarashi T, Konno R, Okamoto S, Moriya T,
Satoh S and Yajima A: Involvement of granule-mediated apoptosis in
the cyclic changes of the normal human endometrium. Tohoku J Exp
Med. 193:13–25. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lash GE, Schiessl B, Kirkley M, Innes BA,
Cooper A, Searle RF, Robson SC and Bulmer JN: Expression of
angiogenic growth factors by uterine natural killer cells during
early pregnancy. J Leukoc Biol. 80:572–580. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lobo SC, Huang ST, Germeyer A, Dosiou C,
Vo KC, Tulac S, Nayak NR and Giudice LC: The immune environment in
human endometrium during the window of implantation. Am J Reprod
Immunol. 52:244–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ashkar AA, Di Santo JP and Croy BA:
Interferon gamma contributes to initiation of uterine vascular
modification, decidual integrity, and uterine natural killer cell
maturation during normal murine pregnancy. J Exp Med. 192:259–270.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Acar N, Ustunel I and Demir R: Uterine
natural killer (uNK) cells and their missions during pregnancy: A
review. Acta Histochemica. 113:82–91. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li XF, Charnock-Jones DS, Zhang E, Hiby S,
Malik S, Day K, Licence D, Bowen JM, Gardner L, King A, et al:
Angiogenic growth factor messenger ribonucleic acids in uterine
natural killer cells. J Clin Endocrinol Metab. 86:1823–1834. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jokhi PP, King A and Loke YW: Cytokine
production and cytokine receptor expression by cells of the human
first trimester placental-uterine interface. Cytokine. 9:126–137.
1997. View Article : Google Scholar : PubMed/NCBI
|