Introduction
Ketogenic diets (KDs) are low carbohydrate, adequate
protein, high fat diets that mimic fasting and induce ketosis
(1). KDs are well-established,
effective and clinically approved dietary treatments for children
with drug-resistant epilepsy and are used therapeutically in other
neurological conditions such as Alzheimer's disease, traumatic
brain injury and stroke (2,3). Moreover, they have been shown to be
beneficial as adjuvant therapy in certain types of cancer,
including lung, stomach and ovarian cancer (4,5).
Although, preclinical evidence indicates that KDs are safe and
feasible for use in cancer treatment (6), the safety of KDs in combination with
chemotherapy is still unclear, so their potential use as adjuvant
cancer treatment remains controversial.
The liver is a major organ for the biotransformation
of xenobiotics and for the conversion of fatty acids into ketone
bodies, such as β-hydroxybutyrate, acetoacetate and acetone
(7). Therefore, hepatic metabolism
serves a major role in substrate availability and metabolic
alterations during fasting or with fasting mimicking diets such as
KDs, especially when given simultaneously with chemotherapy
(8–10). It was recently reported that a KD
enhances the anticancer effect of gemcitabine
(2′,2′-difluoro-2′-deoxycytidine), improves survival and mitigates
cachexia in
LSL-KrasLSL-G12D/+Trp53R172H/+Pdx-1-
Cre (KPC) tumor-bearing mice (11,12).
Although additional evidence on the efficacy of KDs in combination
with chemotherapy exists (4,13),
little is known regarding the safety of KDs with or without
concomitant treatment with gemcitabine, an antineoplastic drug
commonly used in pancreatic cancer treatment. While gemcitabine is
generally considered safe, it has been associated with elevations
in indicators of liver disease, such as serum aminotransferases
[aspartate aminotransferase (AST) and alanine aminotransferase
(ALT)] and with liver injury in patients with preexisting liver
disease (14). Furthermore, a small
number of case reports have indicated that this cytotoxic drug can
cause steatohepatitis and hepatotoxicity (15–17).
Given the potential of KDs as a therapeutic diet for
pancreatic cancer when combined with chemotherapy (12), comprehensive safety studies are
critical in order to advance this promising treatment strategy into
the clinic. Therefore, the present study performed a secondary
analysis using previously collected liver and serum samples from
our recent study (12), to evaluate
the liver safety profile of a KD in combination with gemcitabine in
KPC mice bearing pancreatic tumors.
Materials and methods
Animal studies
All animal experiments were performed according to
ethical guidelines and were approved by The Animal Care and Use
Committee of the University of California (approval no. 20555). KPC
mice, bred at The UC Davis Animal Facility in Meyer Hall, were
generated from three mouse parental strains
(LSL-KrasG12D/+,
LSL-Trp53R172H/+ and Pdx-1-Cre)
backcrossed on the C57BL/6 strain background, as previously
described (18). The genetic
background of each pup was confirmed by toe DNA extraction and PCR.
Briefly, genomic DNA was extracted using the Extract-N-Amp™ Tissue
PCR kit (cat. no. XNAT2R; MilliporeSigma) following the
manufacturer's instructions. PCR was performed for Kras, p53 and
Pdx-1-Cre using the following conditions: Initial denaturation at
95°C for 2 min, followed by 37 cycles at 95°C for 0.5 min, 58°C for
1 min and 68°C for 1 min. The oligonucleotide primer sequences used
were as follows: Kras 5′-CCTTTACAAGCGCACGCAGAG-3′ sense, and
5′-AGCTAGCCACCATGGCTTGAGTAAGTCTGCA-3′ anti-sense; Control p53
5′-CTTGGAGACATAGCCACACTG-3′, Mutated p53
5′-AGCTAGCCACCATGGCTTGAGTAAGTCTGCA-3′, WT p53;
5′-TTACACATCCAGCCTCTGTGG-3′, and Cre 5′-CTGGACTACATCTTGAGTTGC-3′
sense and 5′-GGTGTACGGTCAGTAAATTTG-3′ antisense. PCR products were
separated on a 2% agarose gel, stained with GelRed®
(cat. no. 41003; Biotium, Inc.) and visualized in a Chemidoc™
Imaging-System (Bio-Rad Laboratories, Inc.). After weaning and
confirmation of the genotype by PCR, KPC mice [3 months old;
weighing between 20 and 25 g (females), and 23 and 28 g (males)]
were individually housed in polycarbonate cages in a room with
controlled humidity (40–60%) and temperature (22–24°C), maintained
on a 12 h light-dark cycle and fed LabDiet 5001 chow diet
(LabDiet®) ad libitum until they were enrolled in
the studies.
Dietary and chemotherapy
interventions
The experimental design of the present secondary
analysis has been previously described (12). Briefly, following tumor size
determination using a high-resolution ultrasound imaging of the
pancreas using a Vevo 2100 System (Visual Sonics; FUJIFILM Wako
Pure Chemical Corporation), male and female KPC mice (4–5
mice/sex/group), with a confirmed tumor volume of ~250
mm3, were randomized to either a control group (CG; %
kcal: 20% fat, 65% carb, 15% protein + gemcitabine) or a ketogenic
group (KG; % kcal: 84% fat, 15% protein, 1% carb + gemcitabine).
Gemcitabine was administered to the CG and KG groups at 100 mg/kg
by intraperitoneal injection twice per week for 3.5 weeks (seven
total injections). Lard was the main fat source in the KD, whereas
in the control diet (CD) it was soybean oil (Table SI). The exact fatty acid
composition of the diets is presented in Table I. The mineral mix, TD94046 was used
for the CD and TD79055 was used for the KD due to the lower
carbohydrate content (both from Envigo Rms, Inc.; Bioanalytical
Systems, Inc.). For both diets, the CA40060 vitamin mix was used
(Inotiv; Table SI). Gemcitabine
(>99% 2′-deoxy-2′,2′-difluorocytidine; Gemzar; cat. no.
LY-188011; Thermo Fisher Scientific, Inc.) was administered at 100
mg/kg by i.p, injection 2×/week for 3.5 weeks (for a total of 7
injections). Throughout the study, KPC mice were weighed twice a
week and observed daily for signs of inactivity and presence of
abdominal ascites. Endpoint criteria included the development of
abdominal ascites, weight loss >20% of the initial weight,
extreme weakness and inactivity. After 2 months of dietary
treatment, mice were euthanized by carbon dioxide inhalation at a
displacement rate of 30% vol/min and the liver was dissected,
weighed and split into three parts that were either stored in
liquid nitrogen, RNAlater® or 10% buffered formalin.
 | Table I.Fatty acid composition in the CD and
the KD. |
Table I.
Fatty acid composition in the CD and
the KD.
| Fatty acids | CD, % of total
fatty acids | KD, % of total
fatty acids |
|---|
| SFA | 17.3 | 35.3 |
| cis-MUFA | 20.3 | 39.1 |
| n6-PUFA | 53.6 | 23.0 |
| n3-PUFA | 8.2 | 2.0 |
| n6/n3 ratio | 6.5 | 11.2 |
| Total PUFAs | 61.8 | 25.0 |
Metabolic measurements
Blood was collected via cardiac puncture after
euthanasia and serum was isolated following centrifugation at 3,000
× g for 10 min at room temperature. Total serum cholesterol (cat.
no. 03039773), triglycerides (cat. no. 20767107 322), ALT (cat. no.
20764957 322), AST (cat. no. 20764949 322), alkaline phosphatase
(ALP; cat. no. 03333752 190), total bilirubin (cat. no. 05795397
190), albumin (cat. no. 04469658 190), creatinine (cat. no.
03263991 190), total protein (cat. no. 03183734 190) and blood urea
nitrogen (cat. no. 04460715 190) were measured using COBAS INTEGRA
kits (Roche Diagnostics) according to the manufacturer's
instructions.
Histology
After necropsy, liver tissue was fixed in 10% (w/v)
buffered formalin overnight at 4°C. Tissues were processed,
embedded in paraffin and sectioned (5 µm) by routine methods.
Tissue sections were stained with hematoxylin and eosin or Masson's
trichrome (Chromaview; Thermo Fisher Scientific, Inc.) following
standard protocols (19,20), and analyzed, blind, by a
pathologist. Sections were examined using an Olympus BX46
microscope (Olympus Corporation), with ×20 and ×40 objective
lenses. The liver sections were scored for the presence of
macrovesicular and microvesicular steatosis and hepatocyte
hypertrophy according to the method reported by Liang et al
(21). Briefly, the severity of
macrovesicular steatosis and microvesicular steatosis was graded
based on the percentage of the total area affected, as follows: 0
(<5%), 1 (5–33%), 2 (34–66%) and 3 (>66%). Macrovesicular
steatosis was defined as the displacement of the nucleus to the
side by vacuoles and microvesicular steatosis was defined as the
absence of this displacement. Similarly, the level of
hepatocellular hypertrophy, defined as cellular enlargement
>1.5× the normal hepatocyte diameter, was scored, based on the
percentage of the total area affected, into the following
categories: 0 (<5%), 1 (5–33%), 2 (34–66%) and 3 (>66%). The
evaluation of hepatocellular hypertrophy was based on abnormal
enlargement of the liver cells, irrespective of rounding of the
hepatocytes and/or changes in cytoplasm or the number of vacuoles.
Inflammation was scored based on the number of inflammatory cell
clusters (consisting of ≥5 lymphocytes) averaged over five fields
at ×200 magnification, as follows: 0 (<0.5 foci), 1 (0.5-1.0
foci), 2 (1.0-2.0 foci) and 3 (>2.0 foci) (14).
Western blot analysis
Liver tissue homogenates were prepared using RIPA
buffer (Thermo Fisher Scientific, Inc.), as previously described
(22). The protein concentration
was determined using the Bradford method. Aliquots of total
homogenates containing 25–40 µg protein were separated by SDS
8–12.5% (w/v) polyacrylamide gel electrophoresis and electroblotted
onto nitrocellulose membranes. After blocking membranes in 5% (w/v)
nonfat milk for 1 h at room temperature, they were incubated
overnight at 4°C with phospho-extracellular signal-regulated
protein kinases 1 and 2 (Thr202/Tyr204; p-ERK1/2; cat. no. 4376;
RRID: AB_331772), extracellular signal-regulated protein kinases 1
and 2 (ERK1/2; cat. no. 9102; RRID: AB_330744), phospho-protein
kinase B (Ser473; p-Akt; cat. no. 4060; RRID: AB_2315049), protein
kinase B (AKT; cat. no. 9272; RRID: AB_329827), AMP-activated
protein kinase (AMPK; cat. no. 2795; RRID: AB_560856),
phospho-AMP-activated protein kinase (Thr172; p-AMPK; cat. no.
2535; RRID: AB_331250), p-IκBα (Ser32; cat. no. 2859; RRID:
AB_561111), IκBα (cat. no. 4814; RRID: AB_390781), phospo-p65
(Ser536; p-p65; cat. no. 3033; RRID: AB_331284), p65 (cat. no.
8242; RRID: AB_10859369), acetylated lysine (cat. no. 9441; RRID:
AB_331805), hexokinase 2 (HK2; cat. no. 2867; RRID: AB_2232946),
pyruvate dehydrogenase (PDH; cat. no. 3205; RRID: AB_2162926),
phosphofructokinase (PFK; cat. no. 13123; RRID: AB_2617178),
acetyl-CoA carboxylase (ACC; cat. no. 3676; RRID: AB_2219397) and
toll-like receptor 2 (TLR2; cat. no. 12276; RRID: AB_2797867)
primary antibodies from Cell Signaling Technology, Inc. In
addition, 3-hydroxymethyl-3-methylglutaryl-CoA synthase (HMGCS;
cat. no. sc-373681; RRID: AB_10947237), sterol regulatory element
binding protein 1 (SREBP1; cat. no. sc-13551; RRID: AB_628282),
peroxisome proliferator-activated receptor α (PPARα; cat. no.
sc-398394; RRID: AB_2885073), fatty acid synthase (FAS; cat. no.
sc-74540; RRID: AB_1121387), phospho-acetyl-CoA carboxylase (p-ACC;
cat. no. sc-271965; RRID: AB_10710517), fibronectin (cat. no.
sc-271098, RRID: AB_10608215), collagen type 1α1 chain (Col1A1;
cat. no. sc-59772; RRID: AB_1121787) and toll-like receptor 4
(TLR4; cat. no. sc-293072; RRID: AB_10611320) primary antibodies
were purchased from Santa Cruz Biotechnology, Inc. Finally,
3-hydroxymethyl-3-methylglutaryl-CoA lyase (HMGCL) (cat. no.
16898-1-AP; RRID: AB_2295304) primary antibodies were purchased
from Proteintech Group, Inc. and 4-hydroxynonenal (4-HNE; cat. no.
ab46545; RRID: AB_722490) primary antibodies were purchased from
Abcam. All antibodies were prepared using a 1:1,000 dilution. The
next day, membranes were washed three times with TBS-tween 0.05%
(v/v) and incubated with secondary antibodies [either HRP (cat. no.
7074) or biotinylated antibodies (cat. no. 14708) from Cell
Signaling Technology, Inc.; dilution, 1:2,500] for 1 h at room
temperature. Finally, following another set of washes, the
conjugates were incubated with the ProSignal® Pico ECL
reagent (cat. no. 20–300; Genesee Scientific Corporation),
visualized and quantified by chemiluminescence detection in a
Chemidoc™ Imaging-System (Bio-Rad Laboratories, Inc.). β-actin
(cat. no. A1978) from MilliporeSigma (Merck KGaA) or vinculin (cat.
no. 13901; RRID: AB_2728768) from Cell Signaling Technology, Inc.
were used as loading controls. The densitometric analysis was
performed using ImageJ version 2.3.0/1.53f (National Institutes of
Health).
Liver fatty acid analysis
The content of fatty acid in KPC livers of CG and
KG-treated mice was measured using gas chromatography (GC). Liver
samples were freeze-dried and direct-methylated with sodium
methoxide as previously described (23). Prior to the methylation step,
cis-10-17:1 methyl ester (Nu-Check Prep, Inc.) was added to the
samples as an internal standard. Fatty acid methyl esters (FAMEs)
were analyzed using a CP-Sil88 column (100 m; 25 µm ID; 0.2 µm film
thickness) in a TRACE 1310 gas chromatograph (Thermo Fisher
Scientific, Inc.), which was equipped with a flame-ionization
detector (GC-FID; Thermo Fisher Scientific, Inc.). Each sample was
analyzed twice using a 175°C plateau temperature program (23). FAMEs were quantified using
chromatographic peak area and internal standard-based calculations
(24).
Statistical analysis
Data are presented as the mean ± SEM;
(n=4-5/sex/group). Each experiment was conducted once. Statistical
analysis was performed by unpaired t-test or two-factor analysis of
variance followed by Tukey's test for multiple comparisons using
GraphPad Prism software (Dotmatics; version 9.2). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of a KD in combination with
gemcitabine on liver function tests in KPC mice
It has been recently reported that a KD plus
gemcitabine enhances overall survival and mitigates cachexia in KPC
mice compared with mice fed a CD (11,12).
Based on these promising results, the present secondary analysis
was performed to evaluate the impact of a KD in combination with
gemcitabine on the liver safety profile in KPC mice.
Initially, the safety profile of KG in female and
male KPC mice treated for 2 months was evaluated. CD + gemcitabine
was selected as the CG to specifically assess the effect of a KD.
No significant differences in body weight were observed between CG
and KG throughout the treatment (Fig.
1A and B). Moreover, no differences in liver weight were
observed between groups at the endpoint (Fig. 1C). No liver metastatic events were
observed in CG- or KG-treated KPC mice at euthanasia.
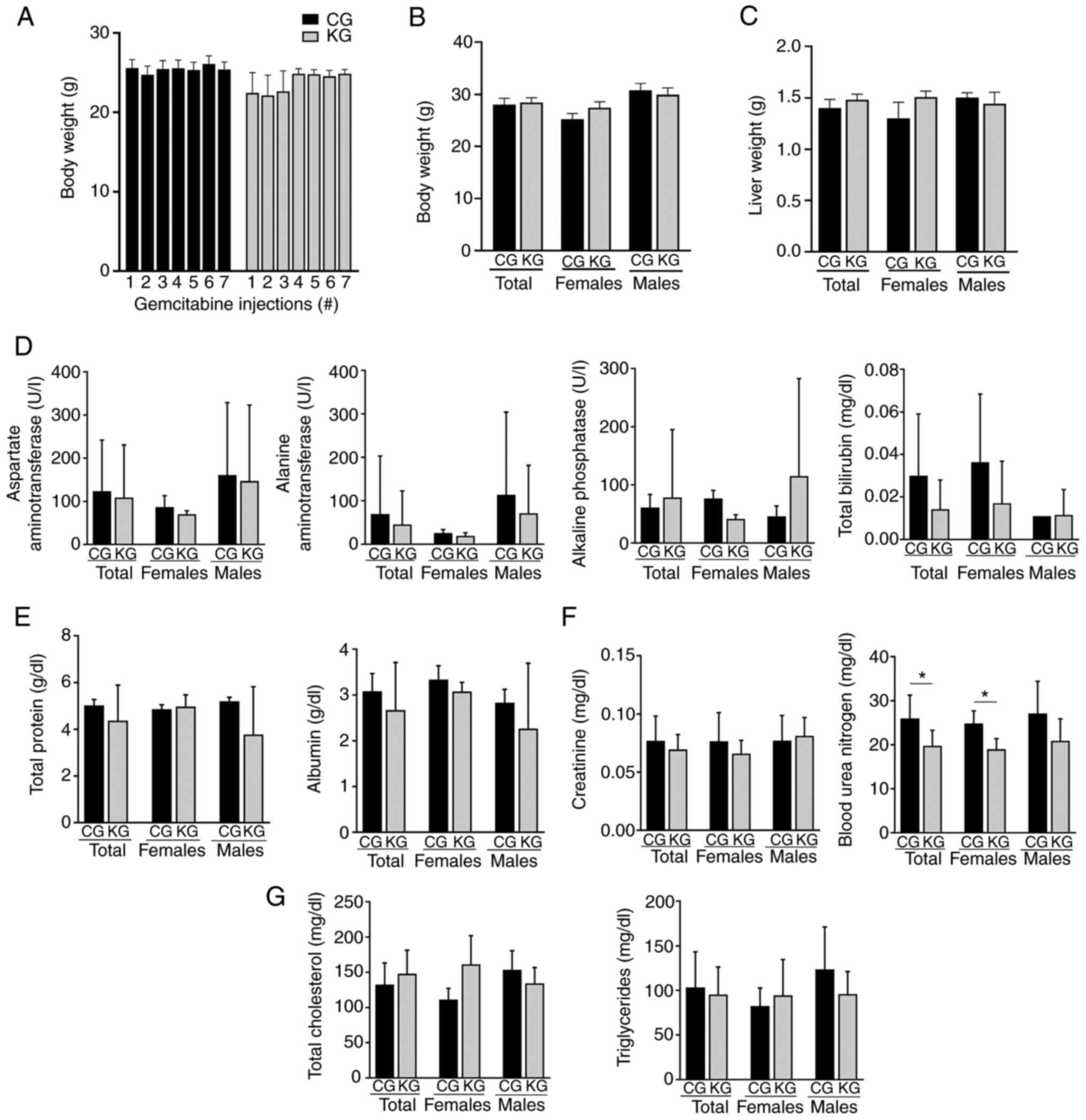 | Figure 1.Effect of a KD in combination with
gemcitabine on body weight, liver weight, liver enzymes, kidney
markers and serum lipids in KPC mice. (A) Body weight at each
gemcitabine injection, (B) final body weight and (C) liver weight
at euthanasia, (D) serum levels of liver enzymes aspartate
aminotransferase, alanine aminotransferase and alkaline phosphatase
and bilirubin levels. (E) Serum total protein and albumin levels.
(F) Serum creatinine and blood urea nitrogen levels. (G) Serum
cholesterol and triglycerides levels. Values are presented as mean
± SEM, *P<0.05. n=4-5 animals/group/sex. KPC,
LSL-KrasLSL-G12D/+
Trp53R172H/+Pdx-1-Cre; CG, control plus
gemcitabine group; KG, ketogenic plus gemcitabine group. |
To determine whether KG affected normal liver
function, the levels of serum liver enzymes AST, ALT and ALP, as
well as bilirubin levels in female and male KPC mice at euthanasia
were determined (Fig. 1D). After 2
months of treatment, there was no significant difference in liver
enzyme and bilirubin levels between the CG- and KG-treated groups.
Of note, all the mice had values within normal ranges, except one
male mouse in the CG group and one male mouse in the KG group,
which showed liver enzyme levels higher than the physiological
range for C57BL/6 mice (physiological range, 46–221 U/l for AST,
22–133 U/l for ALT, 16–200 U/l for ALP and 0.4±0.5 mg/dl for
bilirubin) (25). Moreover, no
differences in serum total protein and albumin levels were observed
(Fig. 1E).
Kidney function indicators were also assessed, with
no significant differences in creatinine levels between the CG and
KG. In contrast, higher blood urea nitrogen levels were observed in
the serum of CG-treated KPC female mice, as well as when both sexes
were analyzed together, compared with the KG-treated group
(Fig. 1F). The serum lipid profile
was measured and no significant differences in total cholesterol or
triglycerides were observed between CG and KG groups (Fig. 1G).
Effect of a KD in combination with
gemcitabine on liver lipid accumulation
Whether a KD in combination with gemcitabine could
impact hepatic lipid accumulation/steatosis was subsequently
evaluated. For this purpose, histologic sections of liver from CG
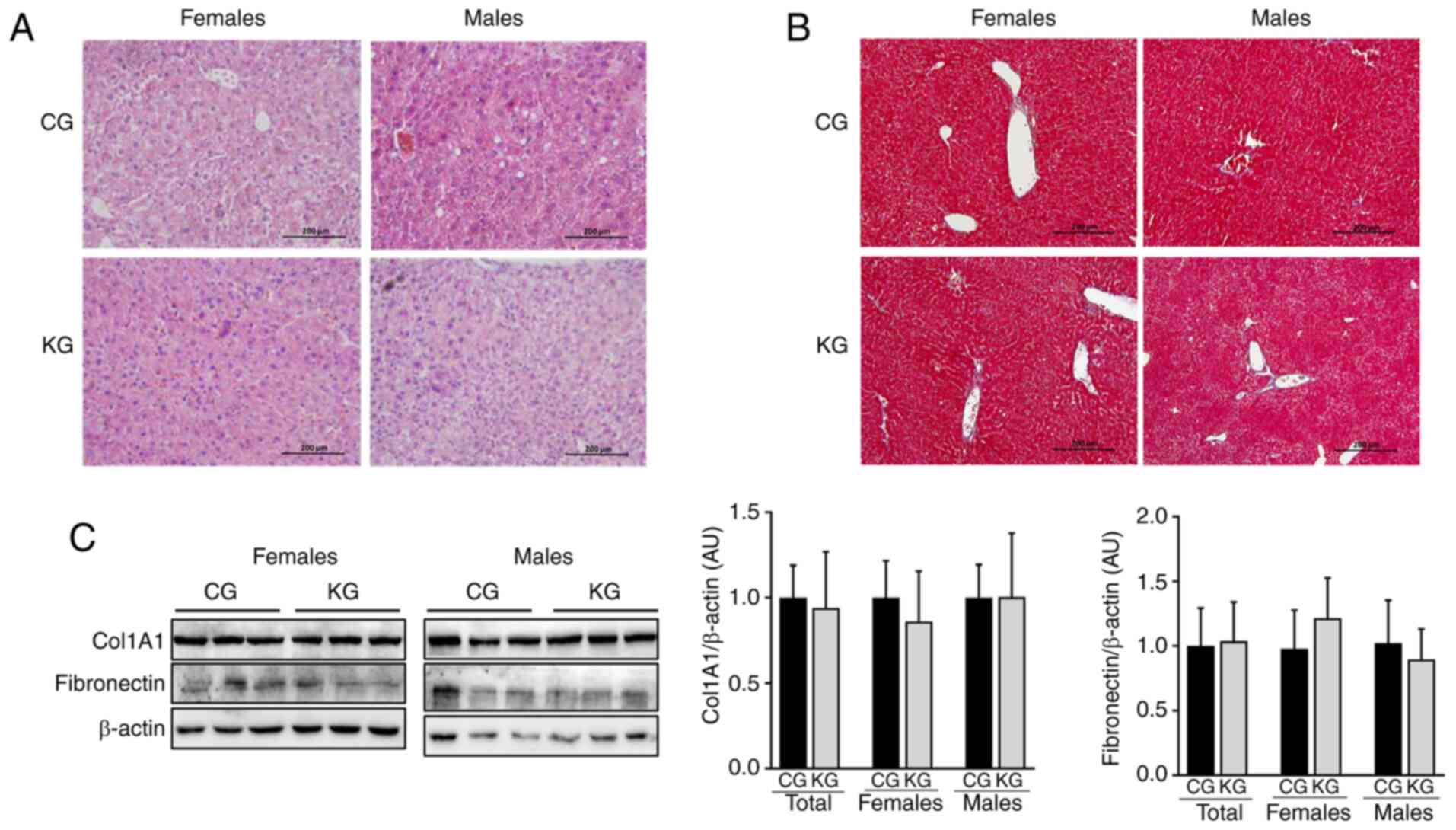
and KG-treated mice were evaluated (Fig. 2A and B) and the presence of
macrovesicular steatosis, microvesicular steatosis and hepatocyte
hypertrophy were assessed using a previously reported scoring
system (21). Overall, no notable
differences in macrovesicular steatosis, microvesicular steatosis,
hepatocyte hypertrophy or inflammation were observed between CG-
and KG-treated mice. Additionally, no histologic evidence of
fibrosis was observed in either group (Fig. 2B). This was confirmed by evaluating
the protein expression levels of fibronectin and Col1A1 using
immunoblotting. No significant differences in fibronectin and
Col1A1 levels were demonstrated in liver homogenates obtained from
CG- and KG-treated mice, together or when separated by sex
(Fig. 2C).
Effect of a KD in combination with
gemcitabine on liver fatty acid composition
Next, whether feeding a KD plus gemcitabine could
affect fatty acid profiles in the liver was evaluated. Compared
with CG-treated mice, KG significantly increased the proportion of
total saturated fatty acids (SFAs), mainly driven by the increased
levels of stearic acid (18:0) and margaric acid (17:0) and
significantly reduced the proportion of cis-monounsaturated fatty
acids (c-MUFAs), including reductions in palmitoleic acid
(cis9-16:1), oleic acid (cis9-18:1) and ascleptic acid
(cis11-18:1). In contrast, no significant differences were
demonstrated in the concentrations of n6-polyunsaturated fatty
acids (PUFAs), n3-PUFAs or total PUFAs in the liver tissue of the
KG mice compared with the CG mice (Fig.
3; Table SII).
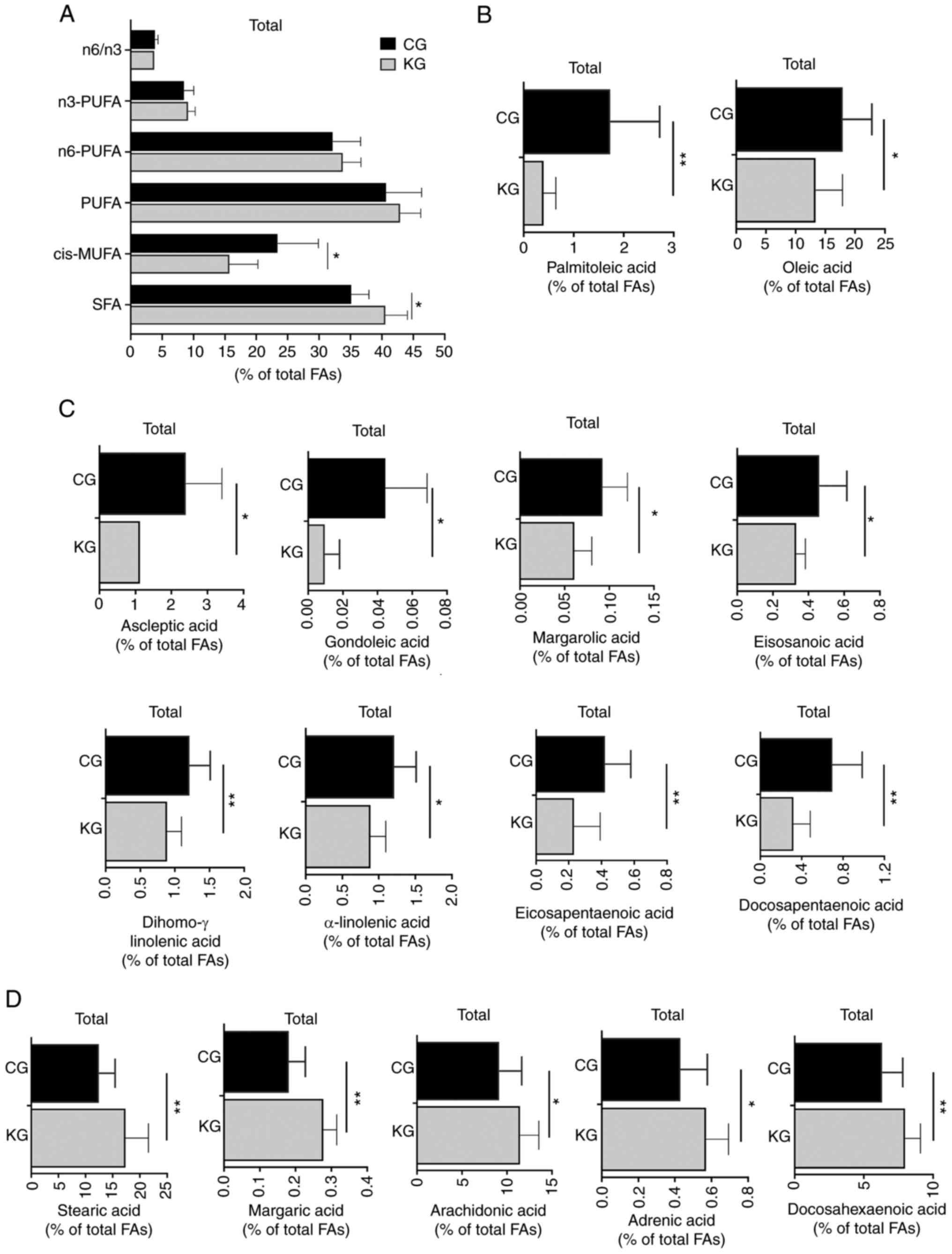 | Figure 3.Effect of a ketogenic diet in
combination with gemcitabine on liver FA composition. (A) SFAs,
MUFAs, n-6 and n-3 PUFAs, as well as the n-6/n-3 fatty acid ratio,
concentrations (% of total FAs) of select FAs, (B) palmitoleic acid
(cis9-16:1) and oleic acid (cis9-18:1), (C) ascleptic acid
(cis11-18:1), gondoleic acid (cis9-20:1), margarolic acid
(cis9-17:1), eicosanoic acid (cis11-20:1), dihomo-g-linolenic acid
(20:3, n-6), α-linolenic acid (18:3n-3), eicosapentaenoic acid
(20:5n-3) and docosapentaenoic acid (22:5n-3) and (D) stearic acid
(18:0), margaric acid (17:0), arachidonic acid (20:4n-6), adrenic
acid (22:4n-6) and docosahexaenoic acid (22:6n-3) in liver
homogenates isolated from CG- and KG-treated KPC female and male
mice following 2 months of treatment. Values are presented as mean
± SEM, *P<0.05 and **P<0.01 n=5 animals/group/sex. SFAs,
short chain fatty acids; FAs, fatty acids; MUFAs, monounsaturated
fatty acids; PUFAs, polyunsaturated fatty acids; CG, control plus
gemcitabine group; KG, ketogenic plus gemcitabine group; KPC,
LSL-KrasLSL-G12D/+Trp53R172H/+
Pdx-1-Cre. |
When analyzing individual fatty acids, it was
observed that dihomo-g-linolenic acid (20:3, n-6) from the n6
family, α-linolenic acid (18:3n-3), eicosapentaenoic acid (20:5n-3)
and docosapentaenoic acid (22:5n-3) from the n3 family were
significantly reduced in KG group in contrast to CG group. At the
same time, arachidonic acid (20:4n-6), adrenic acid (22:4n-6) from
the n6 family and docosahexaenoic acid (22:6n-3) from the n3 family
were significantly increased in KG-treated livers compared with the
CG group (Table SII).
Effect of a KD in combination with
gemcitabine on de novo lipogenesis regulatory proteins
Next, whether KG could influence the expression
levels of sterol regulatory element-binding proteins (SREBPs) were
evaluated, which together with FAS and ACC enzymes are central
regulators of de novo lipogenesis (26–28).
No significant differences were observed in either SREBP1 protein
expression levels or ACC activation between CG- and KG-treated
mice. However, KG treatment significantly reduced FAS protein
expression levels in the livers of male mice, compared with
CG-treated mice (Fig. 4). Moreover,
no significant differences were observed in the liver protein
expression levels of PPARα, a key nuclear receptor regulating de
novo lipogenesis (29), between
CG- and KG-treated mice, nor by sex (4–5 mice/group).
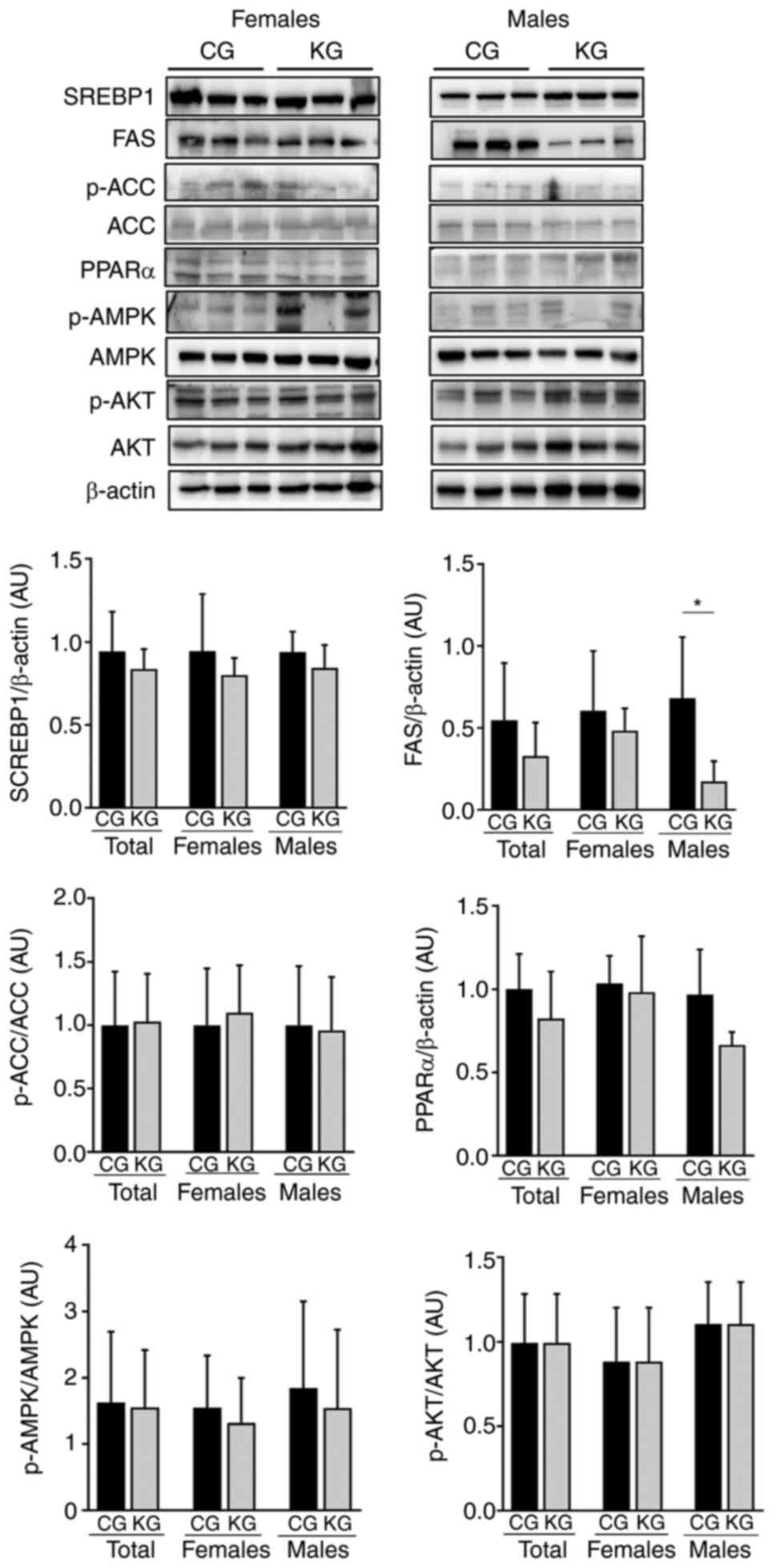 | Figure 4.Effect of a KD in combination with
gemcitabine on enzymes involved in de novo fatty acids synthesis.
Immunoblots of SREBP-1, FAS, p-ACC/ACC, PPARα, pAMPK/AMPK and
pAKT/AKT from liver homogenates from CG- and KG-treated female and
male KPC mice following 2 months of treatment. Representative
images are shown. Each band represents an independent liver
homogenate sample obtained from either female or male KPC mice
treated with CD or KD. Bands were quantified and values normalized
to the non-phosphorylated protein (ACC, AKT and AMPK) or β-actin
levels (SREBP-1, FAS and PPARα). Results are expressed as a
proportion of the control, *P<0.05, values are presented as mean
± SEM with 4 animals/group/sex. CD, control diet; KD, ketogenic
diet; CG, control plus gemcitabine group; KG, ketogenic plus
gemcitabine group; KPC,
LSL-KrasLSL-G12D/+Trp53R172H/+Pdx-1-Cre;
SREBP-1, sterol regulatory element binding protein 1; FAS, fatty
acid synthase; ACC, acetyl-CoA carboxylase; ACC, acetyl-CoA
carboxylase; PPARα, peroxisome proliferator-activated receptor α;
AMPK, AMP-activated protein kinase; AMPK, AMP-activated protein
kinase; AKT, protein kinase B; AKT, protein kinase B; p,
phosphorylated. |
Finally, given that AMPK and AKT can stimulate de
novo lipid synthesis by activating SREBP (27,28),
whether KG could affect AMPK and AKT phosphorylation levels was
assessed. There were no significant differences in AMPK and AKT
phosphorylation levels demonstrated between CG- and KG-treated mice
(Fig. 4).
Effects of a KD in combination with
gemcitabine on ketogenesis and glucose metabolism
Given that the liver is a major organ involved in
the metabolism of ketone bodies and glucose (30), the regulation of enzymes involved in
ketogenesis (HMGCL and HMGCS) and glucose metabolism (HK2, PFK and
PDH) following KG treatment was investigated. No significant
differences in HMGCL, HMGCS, HK2, PFK and PDH hepatic protein
expression levels were demonstraed between CG- and KG-treated mice
together or separated by sex (Fig. 5A
and B).
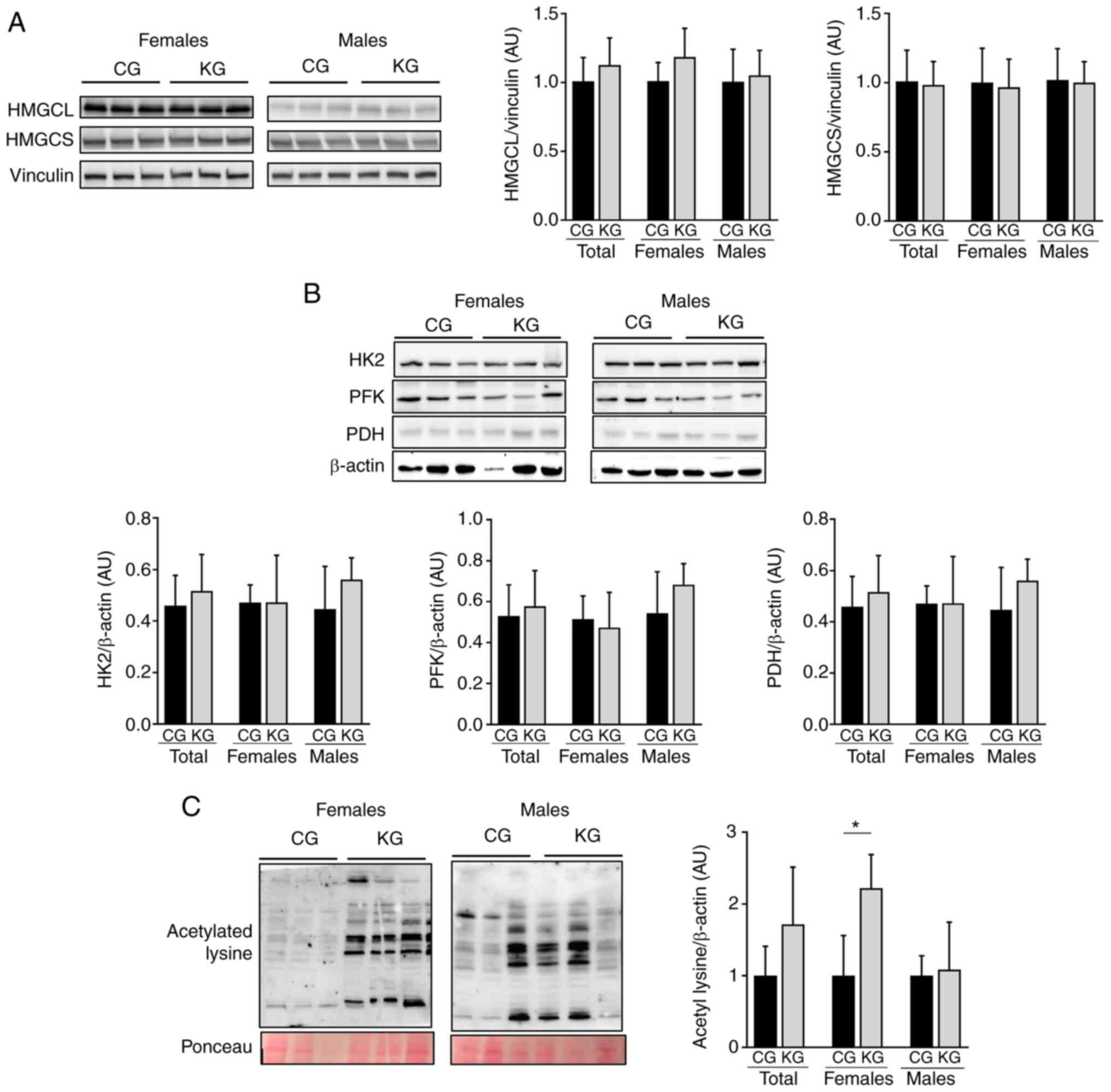 | Figure 5.Effect of a KD in combination with
gemcitabine on ketogenesis and glucose metabolism. Immunoblotting
of (A) HMGCL and HMGCS, (B) HK2, PFK and PDH and (C) acetylated
lysine from liver homogenates from CG- and KG-treated female and
male KPC mice following 2 months of treatment. The loading controls
were vinculin or β-actin. Representative images are shown. Each
band represents an independent liver homogenate sample obtained
from either female or male KPC mice treated with a CD or KD. Bands
were quantified and results are presented as a proportion of the
control; *P<0.05. Values are presented as mean ± SEM with 5
animals/group/sex. CD, control diet; KD, ketogenic diet; CG,
control plus gemcitabine group; KG, ketogenic plus gemcitabine
group; KPC,
LSL-KrasLSL-G12D/+Trp53R172H/+Pdx-1-Cre;
HMGCL, 3-hydroxymethyl-3-methylglutaryl-CoA lyase; HMGCS,
3-hydroxymethyl-3-methylglutaryl-CoA synthase, HK2, hexokinase 2;
PFK, phosphofructokinase; PDH, pyruvate dehydrogenase. |
Acetylation may serve a key role in the coordination
of different metabolic pathways in response to extracellular
conditions, including nutrient availability (31). Since the liver is highly exposed to
lysine acetylation (32), whether a
KD in combination with gemcitabine affected acetylation was
investigated. After 2 months of treatment, significantly increased
lysine acetylation levels in the liver of female, but not male,
KG-treated mice when compared with CG-treated mice were observed
(Fig. 5C).
Effect of a KD in combination with
gemcitabine on markers of inflammation and oxidative stress in the
liver of KPC mice
Activation of TLR signaling is key during liver
inflammation processes, with TLR4 and TLR2 serving crucial roles in
the progression of NASH (33).
Therefore, if a KD could impact the activation of TLR2 and TLR4 was
evaluated. Moreover, since TLR cascades can lead to the activation
of NF-κB and mitogen-activated protein kinases (MAPKs) pathways,
which serve central roles in inflammation, the phosphorylation
levels of IκBα, p65 and ERK1/2 were measured. After 2 months of
intervention, no significant differences in TLR4, TLR2 levels nor
in the downstream IκBα and ERK1/2 phosphorylation levels were
observed in the livers of KG- or CG-treated male and female mice
(Fig. 6A and B). A significant
increase in p65 phosphorylation was observed in all and male mice
in the KG when compared with the KG; however, no significant
difference was demonstrated between KG and CG within the female
group (Fig. 6B).
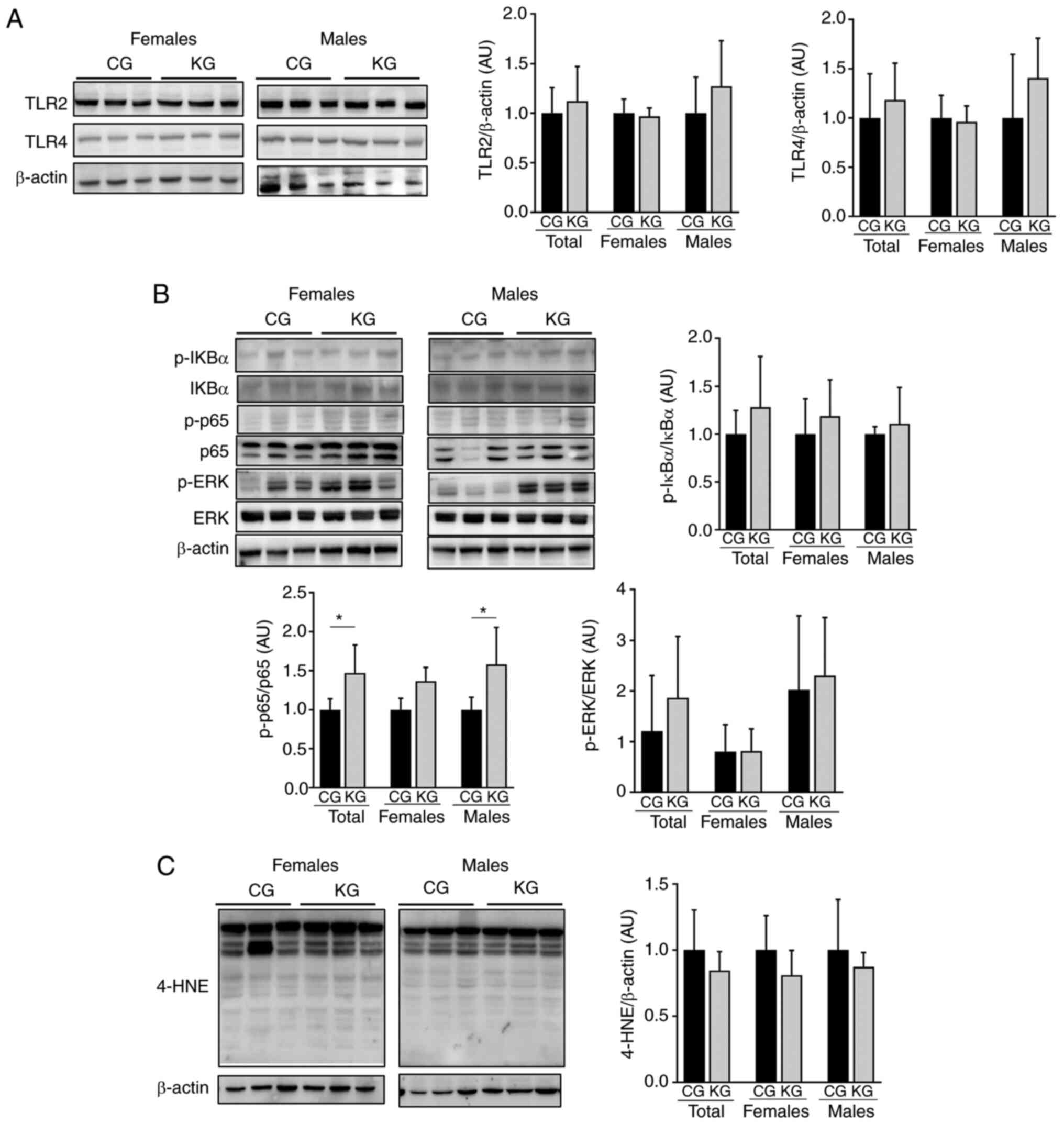 | Figure 6.Effect of a KD in combination with
gemcitabine on markers of inflammation and oxidative stress.
Immunoblotting of (A) TLR2 and TLR4, (B) p-IκBα, IκBα, p-p65, p65,
p-ERK and ERK and (C) 4-HNE from liver homogenates isolated from
CG- and KG-treated female and male KPC mice following 2 months of
treatment. The loading control used was β-actin. Representative
images are shown. Each band represents an independent liver
homogenate sample obtained from either female or male KPC mice
treated with a CD or KD. Bands were quantified and values
normalized to (A) the non-phosphorylated protein and (B and C)
β-actin levels. Results are expressed as a proportion of the
control. *P<0.05. Values are presented as mean ± SEM. n=4
animals/group/sex. CD, control diet; KD, ketogenic diet; CG,
control plus gemcitabine group; KG, ketogenic plus gemcitabine
group; KPC, LSL-KrasLSL-G12D/+
Trp53R172H/+Pdx-1-Cre; TLR, toll-like
receptor; ERK, extracellular signal-regulated protein kinase; ERK,
extracellular signal-regulated protein kinase; 4-HNE,
4-hydroxynonenal; p, phosphorylated. |
Given that oxidative stress serves a major role in
the development of liver injury (34), next 4-HNE levels, a main lipid
peroxidation product that displays increased levels with oxidative
stress (35), were measured. No
significant differences in 4-HNE levels were observed between KG-
and CG for all mice or those separated by sex (Fig. 6C).
Discussion
The characterization of the safety profile of KDs
concomitant with chemotherapeutic treatments for pancreatic cancer
is critical in order to develop clinical recommendations. Data on
the overall safety and feasibility of KDs indicate that this diet
can be tolerated by patients with cancer (36). Nevertheless, the safety of KDs when
administered with standard cancer treatment modalities in patients
with pancreatic cancer is still unknown. Gemcitabine is a main
chemotherapeutic agent used for the treatment of pancreatic cancer
(37). Although it is considered
generally safe, it can cause adverse hepatic events such as
elevations in serum ALT, ALP and bilirubin (38,39). A
case report previously reported that gemcitabine monotherapy caused
hepatic failure in a patient with advanced pancreatic cancer after
pancreaticoduodenectomy (40).
Several human studies have reported the benefits of
using a KD in pancreatic cancer treatment. For instance, certain
randomized controlled trials have reported that a KD may improve
the quality of life and overall survival of patients (41,42).
Additionally, it has been suggested that a KD might help to reduce
tumor growth and slow disease progression in certain cases
(43–45). It was recently reported that when
combined with the chemotherapeutic agent gemcitabine, a KD
increases overall survival in the autochthonous-clinically relevant
KPC mouse model, without any signs of intolerance to the fat
content of the diet (12).
Therefore, the liver safety profile of a KD in combination with
gemcitabine in KPC tumor-bearing mice was evaluated. To the best of
our knowledge, this is the first study to evaluate the liver safety
of a KD in combination with gemcitabine.
In the present study, multiple parameters of liver
toxicity were assessed in pancreatic tumor-bearing mice treated
with a KD plus gemcitabine. Overall, there were no changes in liver
enzymes nor in markers of kidney function with the KG regimen.
Consistent with the findings of the present study, a previous study
reported normal liver and kidney function tests were observed in
rats after being fed a KD for 60 days (46). A recent study reported that liver
function markers differ among groups fed lard, soybean oil or a
blend of both, with the levels of AST and ALT being lower in
subjects that consumed the blend oil for 12 weeks compared with
those in subjects in the pure soybean oil or lard groups (47). The KD composition used in the
present study was prepared primarily with lard (84%) and combined
with soybean oil, while there was no lard in the CD, which should
be taken into consideration for further research.
The effect of KDs on serum lipids has been
inconsistent among previous studies. In a study where healthy male
mice were given a KD for 22 weeks, cholesterol and triglycerides
levels were increased and signs of hepatic steatosis were observed
(48). On the other hand, a KD led
to improvements in total cholesterol and triglycerides in women who
were overweight (49). In the
present study, a KD plus gemcitabine had no effect on serum
cholesterol nor on triglyceride levels.
Long term high-fat diets can induce liver steatosis,
which is a precursor of non-alcoholic fatty liver disease (NAFLD)
(50). Given the high fat content
of a KD, this could be a potential concern. Nevertheless, some
controversy exists as to whether a KD predisposes NAFDL
development. In patients with NAFLD, a KD given for 6 days improved
metabolic abnormalities (51).
Furthermore, a KD prevented the development of steatosis in obese
mice (52). In the present study, a
KD did not alter hepatic lipid accumulation, nor did it affect the
protein or enzyme levels related to de novo lipogenesis,
ketogenesis or glucose metabolism. Moreover, a KD plus gemcitabine
treatment failed to induce macrovesicular or microvesicular
steatosis, hepatocyte hypertrophy or fibrosis. Inflammatory and
oxidative stress markers, two known factors that can contribute to
the pathogenesis of NAFLD (53,54),
were not affected either.
Hepatic fatty acid composition may affect steatosis
development (55,56). In particular, low hepatic n6- and
n3-PUFAs could contribute to steatosis and steatohepatitis
(57). In the present study, it was
observed that a KD plus gemcitabine did not affect hepatic levels
of n6-PUFA, n3-PUFA nor total PUFAs (n6 + n3) compared with CG
mice. On the other hand, significant differences in the composition
of numerous intrahepatic fatty acids following KG-treatment, with
lower MUFA levels, but higher SFAs were observed. Additional
studies are warranted to elucidate the potential impact of each
individual fatty acid affected by KG and their potential role in
steatosis risk or prevention.
Lysine acetylation/deacetylation mediated by histone
acetylases and histone deacetylases (HDACs) is a regulator of
signaling pathways involved in cancer progression (58). Moreover, it serves a role in
metabolic processes involved in liver disease, with certain HDAC
inhibitors considered candidates for hepatocellular carcinoma
treatment (59). β-hydroxybutyrate
is a known HDAC inhibitor (60). In
aging mice, both survival and total acetyl-lysine levels have been
reported to be significantly increased in the liver of KD-fed males
compared with controls (61).
Hutfles et al (62), also
reported an increase in cytosolic and mitochondrial acetylation in
the livers of KD-fed male C57Bl/6J mice in contrast to those fed
standard chow diet. To the best of our knowledge, the present study
is the first to evaluate lysine acetylation in the livers of KD
fed, gemcitabine treated, pancreatic tumor bearing, male and female
mice. It was observed that a KD combined with gemcitabine
significantly increased liver lysine acetylation in females, but
not males, when compared with CG-treated mice. Interestingly,
previously published data elucidated sex-dependent effects of KG
treatment in the skeletal muscle of KPC animals (11), so the results of the present study
highlight the importance of researching the multi-organ effects of
treatments in both males and females. Unfortunately, as the
research design of the present study did not include KPC mice fed
with a KD alone, the exploration of the exact contribution of the
KD to this effect was impeded. In addition, another limitation is
that the analysis was performed using previously collected samples
and the number of samples/group/sex used were not powered for this
particular study. Being underpowered potentially explains why
additional differences between the CGs and KGs were not
demonstrated. Nevertheless, further studies are warranted to
explore the specific role of KDs, without the confounding effect of
gemcitabine, in the hepatic modulation of protein acetylation and
the potential impact of biological sex suggested by these data.
In summary, the findings of the present study
indicate that a KD in combination with gemcitabine appears to be
safe in male and female mice bearing pancreatic tumors. However, it
is important to stress that the liver safety profile of a specific
KD with a specific dose of gemcitabine over two months was
evaluated. Differences in diet composition (macronutrient
distribution and/or amount and type of fats), length of the
treatment and the type of adjuvant drug administered could alter
the liver safety profile when incorporating a KD to the treatment
strategy and require further, future study. Moreover, the present
study evaluated the safety of a KD in combination with gemcitabine.
Additional studies are warranted to evaluate the safety of this
dietary regimen in combination with other chemotherapeutics used
clinically (such as, nab-paclitaxel, 5-FU, irinotecan and
oxaliplatin). Another limitation of the present study is the lack
of evaluation of the liver safety of a KD alone, without the
confounding effect of the chemotherapeutic agent, that would allow
for the direct hepatic safety effect of this dietary intervention
to be evaluated in mice bearing pancreatic tumors. Thus, future
studies should focus on identifying a standardized treatment
protocol that includes the composition, length and regimen for a KD
(63), which would assist in
translating this promising dietary treatment strategy safely into
the clinic.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was funded by The University of California, Davis;
The UCD Comprehensive Cancer center (ELEMENTS initiative); The
University of California Cancer Research Coordinating Committee
(award no. C23CR5560) and NIFA-USDA (grant no. CA-D-NTR-2397-H).
This research was also supported by the Biorepository Shared
Resources, funded by the UC Davis Comprehensive Cancer Center
Support Grant awarded by the National Cancer Institute (grant no.
P30CA093373).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NEC and CRL performed conceptualization,
experiments, visualization, investigation, writing of the original
draft, reviewing and editing. PV analyzed the fatty acids and
related data, and reviewed and edited the manuscript. KM performed
histological analysis, reviewed and edited the manuscript. GGM
performed conceptualization, investigation, writing, reviewing and
editing of manuscript, and acquired funding. NEC and CRL confirm
the authenticity of all the raw data. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
All animal experiments were performed according to
ethical guidelines and were approved by the University of
California, Davis Animal Care and Use Committee (Davis, USA;
approval no. 20555).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhu H, Bi D, Zhang Y, Kong C, Du J, Wu X,
Wei Q and Qin H: Ketogenic diet for human diseases: The underlying
mechanisms and potential for clinical implementations. Signal
Transduct Target Ther. 7:112022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tong X, Deng Y, Liu L, Tang X, Yu T, Gan
J, Cai Q, Luo R and Xiao N: Clinical implementation of ketogenic
diet in children with drug-resistant epilepsy: Advantages,
disadvantages, and difficulties. Seizure. 99:75–81. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thomas JG and Veznedaroglu E: Ketogenic
diet for malignant gliomas: A review. Curr Nutr Rep. 9:258–263.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cortez NE and Mackenzie GG: Ketogenic
diets in pancreatic cancer and associated cachexia: Cellular
mechanisms and clinical perspectives. Nutrients. 13:32022021.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang WH, Wang WQ, Han X, Gao HL, Li TJ,
Xu SS, Li S, Xu HX, Li H, Ye LY, et al: Advances on diagnostic
biomarkers of pancreatic ductal adenocarcinoma: A systems biology
perspective. Comput Struct Biotechnol J. 18:3606–3614. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li J, Zhang H and Dai Z: Cancer treatment
with the ketogenic diet: A systematic review and meta-analysis of
animal studies. Front Nutr. 8:5944082021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kennedy AR, Pissios P, Otu H, Roberson R,
Xue B, Asakura K, Furukawa N, Marino FE, Liu FF, Kahn BB, et al: A
high-fat, ketogenic diet induces a unique metabolic state in mice.
Am J Physiol Endocrinol Metab. 292:E1724–E1739. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bertholdt L, Gudiksen A, Jessen H and
Pilegaard H: Impact of skeletal muscle IL-6 on regulation of liver
and adipose tissue metabolism during fasting. Pflugers Arch.
470:1597–1613. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cunha GM, Guzman G, Correa De Mello LL,
Trein B, Spina L, Bussade I, Marques Prata J, Sajoux I and
Countinho W: Efficacy of a 2-month very low-calorie ketogenic diet
(VLCKD) compared to a standard low-calorie diet in reducing
visceral and liver fat accumulation in patients with obesity. Front
Endocrinol (Lausanne). 11:6072020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Watanabe M, Tozzi R, Risi R, Tuccinardi D,
Mariani S, Basciani S, Spera G, Lubrano C and Gnessi L: Beneficial
effects of the ketogenic diet on nonalcoholic fatty liver disease:
A comprehensive review of the literature. Obes Rev. 21:e130242020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cortez NE, Pathak S, Rodriguez Lanzi C,
Hong BV, Crone R, Sule R, Wang F, Chen S, Gomes AV, Baar K and
Mackenzie GG: A ketogenic diet in combination with gemcitabine
mitigates pancreatic cancer-associated cachexia in male and female
KPC mice. Int J Mol Sci. 24:107532023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cortez NE, Rodriguez Lanzi C, Hong BV, Xu
J, Wang F, Chen S, Ramsey JJ, Pontifex MG, Müller M, Vauzour D, et
al: A ketogenic diet in combination with gemcitabine increases
survival in pancreatic cancer KPC mice. Cancer Res Commun.
2:951–965. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Plotti F, Terranova C, Luvero D, Bartolone
M, Messina G, Feole L, Cianci S, Scaletta G, Marchetti C, Di Donato
V, et al: Diet and chemotherapy: The effects of fasting and
ketogenic diet on cancer treatment. Chemotherapy. 65:77–84. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hailan WAQ, Abou-Tarboush FM, Al-Anazi KM,
Ahmad A, Qasem A and Farah MA: Gemcitabine induced cytotoxicity,
DNA damage and hepatic injury in laboratory mice. Drug Chem
Toxicol. 43:158–164. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stellman A, Loke MM and Mann S: Acute
liver failure secondary to gemcitabine. BMJ Case Rep.
2010:bcr12200813712010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Coeman DC, Verbeken EK, Nackaerts KL,
Demedts MG and Vansteenkiste JF: A fatal case of cholestatic liver
failure probably related to gemcitabine. Ann Oncol. 11:15032000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dobbie M, Hofer S, Oberholzer M and
Herrmann R: Veno-occlusive disease of the liver induced by
gemcitabine. Ann Oncol. 9:6811998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hingorani SR, Wang L, Multani AS, Combs C,
Deramaudt TB, Hruban RH, Rustgi AK, Chang S and Tuveson DA:
Trp53R172H and KrasG12D cooperate to promote chromosomal
instability and widely metastatic pancreatic ductal adenocarcinoma
in mice. Cancer Cell. 7:469–483. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cardiff RD, Miller CH and Munn RJ: Manual
hematoxylin and eosin staining of mouse tissue sections. Cold
Spring Harb Protoc. 2014:655–658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Van De Vlekkert D, Machado E and d'Azzo A:
Analysis of generalized fibrosis in mouse tissue sections with
masson's trichrome staining. Bio Protoc. 10:e36292020.PubMed/NCBI
|
|
21
|
Liang W, Menke AL, Driessen A, Koek GH,
Lindeman JH, Stoop R, Havekes LM, Kleemann R and van den Hoek AM:
Establishment of a general NAFLD scoring system for rodent models
and comparison to human liver pathology. PLoS One. 9:e1159222014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rodriguez Lanzi C, Perdicaro DJ,
Antoniolli A, Fontana AR, Miatello RM, Bottini R and Vazquez Prieto
MA: Grape pomace and grape pomace extract improve insulin signaling
in high-fat-fructose fed rat-induced metabolic syndrome. Food
Funct. 7:1544–1553. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dugan MER, Kramer JKG, Robertson WM,
Meadus WJ, Aldai N and Rolland DC: Comparing subcutaneous adipose
tissue in beef and muskox with emphasis on trans 18:1 and
conjugated linoleic acids. Lipids. 42:509–518. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vahmani P, Rolland DC, McAllister TA,
Block HC, Proctor SD, Guan LL, Prieto N, López-Campos Ó, Aalhus JL
and Dugan MER: Effects of feeding steers extruded flaxseed on its
own before hay or mixed with hay on animal performance, carcass
quality, and meat and hamburger fatty acid composition. Meat Sci.
131:9–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Loeb WF and Quimby FW: The clinical
chemistry of laboratory animals. Taylor and Francis; Philadelphia:
1999
|
|
26
|
Gosmain Y, Dif N, Berbe V, Loizon E,
Rieusset J, Vidal H and Lefai E: Regulation of SREBP-1 expression
and transcriptional action on HKII and FAS genes during fasting and
refeeding in rat tissues. J Lipid Res. 46:697–705. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo S: Insulin signaling, resistance, and
the metabolic syndrome: Insights from mouse models into disease
mechanisms. J Endocrinol. 220:T1–T23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Y, Xu S, Mihaylova MM, Zheng B, Hou X,
Jiang B, Park O, Luo Z, Lefai E, Shyy JY, et al: AMPK
phosphorylates and inhibits SREBP activity to attenuate hepatic
steatosis and atherosclerosis in diet-induced insulin-resistant
mice. Cell Metab. 13:376–388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Softic S, Cohen DE and Kahn CR: Role of
dietary fructose and hepatic de novo lipogenesis in fatty liver
disease. Dig Dis Sci. 61:1282–1293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rui L: Energy metabolism in the liver.
Compr Physiol. 4:177–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao S, Xu W, Jiang W, Yu W, Lin Y, Zhang
T, Yao J, Zhou L, Zeng Y, Li H, et al: Regulation of cellular
metabolism by protein lysine acetylation. Science. 327:1000–1004.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Wang T, Xia J, Yao W and Huang F:
Enzymatic and nonenzymatic protein acetylations control glycolysis
process in liver diseases. FASEB J. 33:11640–11654. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Roh YS and Seki E: Toll-like receptors in
alcoholic liver disease, non-alcoholic steatohepatitis and
carcinogenesis. J Gastroenterol Hepatol. 28 (Suppl 1):S38–S42.
2013. View Article : Google Scholar
|
|
34
|
Cichoż-Lach H and Michalak A: Oxidative
stress as a crucial factor in liver diseases. World J
Gastroenterol. 20:8082–8091. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Castro JP, Jung T, Grune T and Siems W:
4-Hydroxynonenal (HNE) modified proteins in metabolic diseases.
Free Radic Biol Med. 111:309–315. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tan-Shalaby J: Ketogenic diets and cancer:
Emerging evidence. Fed Pract. 34 (Suppl 1):37S–42S. 2017.PubMed/NCBI
|
|
37
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
So E, Crees ZD, Crites D and Wang-Gillam
A: Digital ischemia and necrosis: A rarely described complication
of gemcitabine in pancreatic adenocarcinoma. J Pancreat Cancer.
3:49–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
National Institute of Diabetes and
Digestive Kidney Diseases: Gemcitabine LiverTox: Clinical and
research information on drug-induced liver injury. National
Institute of Diabetes and Digestive and Kidney Diseases; Bethesda,
MD: 2012
|
|
40
|
Okada T, Egawa S, Motoi F, Yamamoto K,
Ottomo S, Sakata N, Rikiyama T, Katayose Y and Unno M: Severe
cholestatic liver failure associated with gemcitabine adjuvant
monotherapy for pancreatic cancer. Clin J Gastroenterol. 4:391–395.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Klement RJ, Weigel MM and Sweeney RA: A
ketogenic diet consumed during radiotherapy improves several
aspects of quality of life and metabolic health in women with
breast cancer. Clin Nutr. 40:4267–4274. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Klement RJ, Champ CE, Kämmerer U,
Koebrunner PS, Krage K, Schäfer G, Weigel M and Sweeney RA: Impact
of a ketogenic diet intervention during radiotherapy on body
composition: III-final results of the KETOCOMP study for breast
cancer patients. Breast Cancer Res. 22:942020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Schmidt M, Pfetzer N, Schwab M, Strauss I
and Kämmerer U: Effects of a ketogenic diet on the quality of life
in 16 patients with advanced cancer: A pilot trial. Nutr Metab
(Lond). 8:542011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang L, TeSlaa T, Ng S, Nofal M, Wang L,
Lan T, Zeng X, Cowan A, McBride M, Lu W, et al: Ketogenic diet and
chemotherapy combine to disrupt pancreatic cancer metabolism and
growth. Med. 3:119–136. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zahra A, Fath MA, Opat E, Mapuskar KA,
Bhatia SK, Ma DC, Rodman SN III, Snyders TP, Chenard CA,
Eichenberger-Gilmore JM, et al: Consuming a ketogenic diet while
receiving radiation and chemotherapy for locally advanced lung
cancer and pancreatic cancer: The university of iowa experience of
two phase 1 clinical trials. Radiat Res. 187:743–754. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Arsyad A, Idris I, Rasyid AA, Usman RA,
Faradillah KR, Latif WOU, Lubis ZI, Aminuddin A, Yustisia I and
Djabir YY: Long-term ketogenic diet induces metabolic acidosis,
anemia, and oxidative stress in healthy wistar rats. J Nutr Metab.
2020:36420352020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu Z, Yuan J, Wen P, Guo X, Li K, Wang Y,
Liu R, Guo Y and Li D: Effect of lard or plus soybean oil on
markers of liver function in healthy subjects: A randomized
controlled-feeding trial. Foods. 12:18942023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ellenbroek JH, van Dijck L, Töns HA,
Rabelink TJ, Carlotti F, Ballieux BE and de Koning EJ: Long-term
ketogenic diet causes glucose intolerance and reduced β- and α-cell
mass but no weight loss in mice. Am J Physiol Endocrinol Metab.
306:E552–E558. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tragni E, Vigna L, Ruscica M, Macchi C,
Casula M, Santelia A, Catapano AL and Magni P: Reduction of
cardio-metabolic risk and body weight through a multiphasic
very-low calorie ketogenic diet program in women with
overweight/obesity: A study in a real-world setting. Nutrients.
13:18042021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ben-Yakov G, Alao H, Haydek JP, Fryzek N,
Cho MH, Hemmati M, Samala V, Shovlin M, Dunleavy K, Wilson W, et
al: Development of hepatic steatosis after chemotherapy for
non-hodgkin lymphoma. Hepatol Commun. 3:220–226. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Luukkonen PK, Dufour S, Lyu K, Zhang XM,
Hakkarainen A, Lehtimäki TE, Cline GW, Petersen KF, Shulman GI and
Yki-Järvinen H: Effect of a ketogenic diet on hepatic steatosis and
hepatic mitochondrial metabolism in nonalcoholic fatty liver
disease. Proc Natl Acad Sci USA. 117:7347–7354. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Okuda T and Morita N: A very low
carbohydrate ketogenic diet prevents the progression of hepatic
steatosis caused by hyperglycemia in a juvenile obese mouse model.
Nutr Diabetes. 2:e502012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Masarone M, Rosato V, Dallio M, Gravina
AG, Aglitti A, Loguercio C, Federico A and Persico M: Role of
oxidative stress in pathophysiology of nonalcoholic fatty liver
disease. Oxid Med Cell Longev. 2018:95476132018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gao B and Tsukamoto H: Inflammation in
alcoholic and nonalcoholic fatty liver disease: Friend or foe?
Gastroenterology. 150:1704–1709. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Berná G and Romero-Gomez M: The role of
nutrition in non-alcoholic fatty liver disease: Pathophysiology and
management. Liver Int. 40 (Suppl 1):S102–S108. 2020. View Article : Google Scholar
|
|
56
|
Musso G, Cassader M, Paschetta E and
Gambino R: Bioactive lipid species and metabolic pathways in
progression and resolution of nonalcoholic steatohepatitis.
Gastroenterology. 155:282–302.e8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Allard JP, Aghdassi E, Mohammed S, Raman
M, Avand G, Arendt BM, Jalali P, Kandasamy T, Prayitno N, Sherman
M, et al: Nutritional assessment and hepatic fatty acid composition
in non-alcoholic fatty liver disease (NAFLD): A cross-sectional
study. J Hepatol. 48:300–307. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ding P, Ma Z, Liu D, Pan M, Li H, Feng Y,
Zhang Y, Shao C, Jiang M, Lu D, et al: Lysine
acetylation/deacetylation modification of immune-related molecules
in cancer immunotherapy. Front Immunol. 13:8659752022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhao Q, Zhang Z, Li J, Xu F, Zhang B, Liu
M, Liu Y, Chen H, Yang J and Zhang J: Lysine acetylome study of
human hepatocellular carcinoma tissues for biomarkers and
therapeutic targets discovery. Front Genet. 11:5726632020.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Newman JC and Verdin E: Ketone bodies as
signaling metabolites. Trends Endocrinol Metab. 25:42–52. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Roberts MN, Wallace MA, Tomilov AA, Zhou
Z, Marcotte GR, Tran D, Perez G, Gutierrez-Casado E, Koike S,
Knotts TA, et al: A ketogenic diet extends longevity and healthspan
in adult mice. Cell Metab. 26:539–546.e5. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hutfles LJ, Wilkins HM, Koppel SJ,
Weidling IW, Selfridge JE, Tan E, Thyfault JP, Slawson C, Fenton
AW, Zhu H and Swerdlow RH: A bioenergetics systems evaluation of
ketogenic diet liver effects. Appl Physiol Nutr Metab. 42:955–962.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Allen BG, Bhatia SK, Anderson CM,
Eichenberger-Gilmore JM, Sibenaller ZA, Mapuskar KA, Schoenfeld JD,
Buatti JM, Spitz DR and Fath MA: Ketogenic diets as an adjuvant
cancer therapy: History and potential mechanism. Redox Biol.
2:963–970. 2014. View Article : Google Scholar : PubMed/NCBI
|